
Abstract
During human embryonic stem cell (ESC) hematopoietic differentiation, the description of the initial steps of lymphopoiesis remains elusive. Using a two-step culture procedure, we identified two original populations of ESC-derived hematopoietic progenitor cells (HPCs) with CD34+CD45RA+CD7− and CD34+CD45RA+CD7+ phenotypes. Bulk cultures and limiting dilution assays, culture with MS5 cells in the presence of Notch ligand Delta-like-1 (DL-1), and ex vivo colonization tests using fetal thymic organ cultures showed that although CD34+CD45RA+CD7− HPCs could generate cells of the three lymphoid lineages, their potential was skewed toward the B cell lineages. In contrast, CD34+CD45RA+CD7+ HPCs predominantly exhibited a T/natural killer (NK) cell differentiation potential. Furthermore these cells could differentiate equivalently into cells of the granulo-macrophagic lineage and dendritic cells and lacked erythroid potential. Expression profiling of 18 markers by quantitative reverse transcription-polymerase chain reaction (qRT-PCR) revealed that CD34+CD45RA+CD7− and CD34+CD45RA+CD7
Introduction
T
Human embryonic stem cells (ESCs) are pluripotent stem cells that are capable of indefinite self-renewal in vitro and differentiation into all cell types of the body, making them uniquely valuable for the development of novel research tools and, potentially, cell-based therapies [13,14]. Hematopoietic differentiation of ESCs reproduces many aspects of embryonic hematopoiesis and provides a powerful in vitro model to elucidate the mechanisms of early hematopoietic commitment [15 –23] that is otherwise practically inaccessible in the human embryo [24]. In recent years, a number of protocols have been successfully developed to promote ESC differentiation into cells of the lymphoid system [25 –31]. However, while these studies provided important guidelines, enabling final production of T, B, or NK cells starting from ESCs, the initial steps of lymphopoiesis from ESCs remain an elusive issue, and the description of multipotent lymphoid progenitors emerging from ESCs has not been addressed so far.
Regarding human fetal and adult early lymphopoiesis, we previously characterized umbilical cord blood (UCB)- and bone marrow (BM)-derived CD34+CD45RAhiCD7+ and CD34+CD45RAhiCD10+Lineage (Lin)− hematopoietic progenitor cells (HPCs) as being multipotent early lymphoid progenitors (MELPs) polarized toward T/NK and B cell lineages, respectively [7]. We provided evidence for their in vitro independent emergence from CD34+CD45RAloCD7− lympho-myeloid progenitors and shed light on the physiological mechanisms by which these cells contribute to early lymphopoiesis during human hematopoietic ontogeny [7,8]. Later, Doulatov et al. provided a comprehensive analysis of the human hematopoietic hierarchy and identified from UCB and BM multi-lymphoid progenitors, as a distinct population of CD45RA+ cells within the CD34+CD38− adult stem cell compartment. This population was shown to give rise to all lymphoid cell types, as well as to monocytes, macrophages, and dendritic cells (DCs) [10]. These results indicated that the lymphoid lineage arises concomitantly with the myeloid lineage within early lymphoid progenitors specified by the cell surface acquisition of CD45RA. Furthermore, Kohn et al. recently identified the earliest stage of lymphoid priming in human BM based especially on the cell surface expression of CD45RA [12]. Interestingly, we have recently shown that embryoid body (EB) cells co-cultured with MS5 stromal cells could generate hematopoietic cells, comprising CD34+CD45RA+ HPCs, which were endowed with a strong NK cell differentiation potential [32].
Building on these findings, we identified CD34+CD45RA+CD7− and CD34+CD45RA+CD7+ cells as two original populations of ESC-derived HPCs. Here, ESC-derived CD34+CD45RA+CD7− and CD34+CD45RA+CD7+ HPCs were compared for their lymphoid potentials and gene expression profiles. We show that, although both populations retain some degree of multipotency, the CD34+CD45RA+CD7+ HPCs are skewed toward the T- and NK-cell lineages, whereas the CD34+CD45RA+CD7− HPCs are enriched in pro-B cells. Our study provides evidence with regard to the development of early multi-lymphoid progenitors from ESCs, thereby establishing valuable tools to explore early ontogenetic events that direct the initiation of lympho-hematopoietic cell fate and enabling efforts to generate therapeutically useful cell types.
Materials and Methods
Cell lines
ESCs from H1 cell line (WiCell Research Institute,
Human UCB cells
Normal UCB samples were collected with the informed consent of the mother, according to approved institutional guidelines. After Pancoll (Dutscher) centrifugation, CD34+ cells were purified by immunomagnetic selection using a CD34 MicroBeads kit (Myltenyi Biotec).
Mice
Experiments of thymus colonization were carried out with NOD.Cg-PrkdcscidIl2rgtm1Wjl /SzJ (NOG) mice embryos (Charles River Laboratories) in accordance with European Directives (86/609/CEE), and with French regulations for animal experimentations following Inserm guidelines. NOG mice were maintained in Animal Core Facility in micro-isolator cages.
Formation of EBs
As described [32], H1 colonies were harvested and cultured in ultra-low attachment six-well plates (Costar) in Iscove's modified Dulbecco's medium (IMDM) (Invitrogen) supplemented with 15% FCS (Biowest), 1% penicillin/streptomycin, 1 mM L-glutamine, 1% nonessential amino-acids, 100 μM 2-ME, and 10 ng/mL basic fibroblast growth factor (bFGF; Miltenyi Biotec). After 24 h, mesoderm specification and generation of HPCs was achieved by replacing the bFGF by the following cytokine cocktails [17,32]: stem cell factor (SCF, 100 ng/mL), fms-like tyrosine kinase 3 ligand (Flt3-L, 10 ng/mL), IL-3 and IL-6 (10 ng/mL), granulocyte colony-stimulating factor (G-CSF, 50 ng/mL), vascular endothelial growth factor (10 ng/mL), and bone morphogenetic protein-4 (BMP4, 10 ng/mL; all from Peprotech). The culture was maintained during 19 days at 37°C in a humidified atmosphere, 5% CO2 and the medium was changed every 4–5 days.
Generation of ESC-derived lymphoid progenitor cells
EBs were dissociated at culture day 19 by collagenase IV (Invitrogen) treatment. EB-derived cells were co-cultured during 2 weeks at 37°C in a humidified atmosphere, 5% CO2 with the mouse stromal cell line MS5 (15×103–50×103 cells/cm2). The cells were cultured in EB medium supplemented with 50 ng/mL SCF, 50 ng/mL Flt3-L, and 50 ng/mL recombinant human thymic stromal lymphopoietin (TSLP; Peprotech) to generate lymphoid progenitor cells as described [7].
Flow cytometry analysis and cell sorting
Immunolabeling of cells was performed by incubating the cells for 30 min at 4°C in PBS containing 2% FCS with the following mAbs (1:50 final): CD34-allophycocyanin (APC, clone 581), CD7-FITC (clone 8H8.1), CD7-PE (clone 8H8.1), CD10- phycoerythrocyanin 5 (PECy5, clone ALB1), CD127-PE (clone R34.34,) CD4-PECy5 (clone 13B8.2), CD8-APC (clone B9.11), TCRαβ-PE (clone IP26A), CD56-PE/CD3-FITC (clones NKH-1/UCHT1), IgM-PE (clone UHB), HLA-DR-APC (clone Immu-357), CD62L-FITC (clone DREG56), CD207-PE (clone DCGM4; all from Beckman Coulter), CD14-FITC (clone M5E2), CD15-FITC (clone HI98), CD45RA-PE (clone HI100), CD45RA-PECy5 (clone HI100), CD19-APC (clone HIB19), CD64-FITC (clone 10.1), CD1a-APC (clone HI149; all from BD Biosciences), and CD1a-PE (clone WM-35; ImmunoTools). Cells were then washed and analyzed with an FACScalibur flow cytometer (Becton Dickinson) using CellQuest software. After 14 days of primary co-culture (Fig. 1A, step II), HPC subsets were sorted based on CD34, CD45RA, CD7 expression (purity>93%), using the FACSDiva instrument (Becton Dickinson). Dead cells were excluded using 7AAD (BD Biosciences) staining. Isotype-matched FITC-, PE-, PECy5-, and APC-conjugated irrelevant mAbs were from BD Biosciences and Beckman Coulter.

Schematic representation of the protocol for lympho-hematopoietic differentiation of ESC-derived HPCs. EB cells derived from H1 ESC line (Step I) were co-cultured with MS5 cells during 2 weeks in the presence of SCF/Flt3-L/TSLP (Step II). Cell-sorted HPC subsets derived from the second step of co-culture were then submitted to specific lymphoid or myeloid culture conditions (Step III). BMP4, bone morphogenetic protein-4; DC, dendritic cell; EB, embryoid body; ESC, embryonic stem cell; Flt3-L, fms-like tyrosine kinase 3 ligand; G-CSF, granulocyte colony stimulating factor; HPC, hematopoietic progenitor cell; NK, natural killer; SCF, stem cell factor; TSLP, thymic stromal lymphopoietin; VEGF, vascular endothelial growth factor.
Gene expression analysis by real-time quantitative reverse transcription-polymerase chain reaction
RNA samples were treated with RNAse-free DNAse (Qiagen) and reverse transcribed into cDNA with oligo-dT primer using Superscript III reverse transcriptase (Invitrogen). One part of the quantitative reverse transcription-polymerase chain reaction (qRT-PCR) was performed using SYBR-Green I Master Mix (Roche diagnostics) on an Mx3005P instrument (Stratagene). The other part of qRT-PCR was performed using TaqMan reagents (LifeTechnolgies) according to the recommendation of the provider. Reverse and forward primer sequences are shown in Supplementary Table S1 (Supplementary Data are available online at
Assessment of NK, B, and DC differentiation potentials in bulk cultures
For NK cell culture conditions, HPCs (1,000 cells/cm2) were co-cultured for 3 weeks as described [7,8], in 48-well plates (Dutscher) with MS5 cells, in RPMI 1640 (Invitrogen), 1% penicillin/streptomycin, 1% L-glutamine, 5% FCS (StemCell Technologies), 10% human AB serum (Jack Boy), 0.1 mM 2-ME with human recombinant cytokines: 50 ng/mL SCF, 50 ng/mL Flt3-L, 5 ng/mL IL-2, 20 ng/mL IL-7, and 20 ng/mL IL-15 (Peprotech). B-cell differentiation of HPCs was assessed, as described [7,8], by growing cells (104 cells/cm2) for 2 weeks in MS5 cell-coated plates in IMDM complemented with 3% FCS, 1% penicillin/streptomycin, 1% L-glutamine, 0.1 mM 2-ME, plus 50 ng/mL SCF, 100 ng/mL human thrombopoietin, and 20 ng/mL IL-7. Half of the complete medium was replaced weekly. Finally, to achieve DC differentiation of the two HPC populations, cells were cultured for 10 days as described [37], in RPMI medium supplemented with 10% FCS, 1% penicillin/streptomycin, 1% L-glutamine, and the following cytokines: 50 ng/mL SCF, 50 ng/mL Flt3-L, 50 ng/mL TNF-α, and 20 ng/mL granulocyte-macrophage colony-stimulating factor (GM-CSF, Peprotech). Half of the culture medium was changed every 3 days. Cultures in NK, B, and DC conditions were conducted at 37°C in a humidified atmosphere, 5% CO2.
Clonogenic assays
Erythroid and granulo-macrophagic potential of each HPC subset was assessed by seeding 1,000 cells in 35-mm duplicate dishes (Corning), in 1 mL complete Methocult GF+H4435 medium (StemCell Technologies), supplemented with IL-3, IL-6, G-CSF, SCF, GM-CSF, and erythropoietin (EPO). Dishes were incubated at 37°C in a humidified atmosphere, 5% CO2. The erythroid burst and colony-forming units (BFU/CFU-E), granulocyte, macrophage, and granulo-macrophagic (CFU-G/M/GM) colony-forming units were counted under an inverted microscope on culture day 14.
Limiting-dilution assays
To assess NK- and B-cell progenitor frequencies, cells were seeded at 300, 100, 30, 10, 3, and 1 cells/well into MS5 cell-coated 96-well plates (ATGC) and cultured as earlier. Cultures lasted for 2 to 3 weeks with half medium changes and fresh cytokines added every 6 to 7 days. Plates were examined weekly, and cell-containing wells were scored under the microscope as previously described [7]. At the end of the cultures, B and NK cells were identified by a fluorescence activated cell sorting analysis of CD19 and CD56 expression, respectively. The maximum likelihood estimate of NK-cell or B-cell precursors was calculated according to the single-hit Poisson model.
Assessment of T-cell differentiation potential
ESC-derived CD45RA+CD7− and CD45RA+CD7+ HPCs were co-cultured with MS5 cells for 4 weeks in αMEM supplemented with 10% human AB serum, 5% FCS (StemCell Technologies), 1% penicillin/streptomycin, 1% L-glutamine, 0,1 mM 2-ME, 250 ng/mL Delta-like-1 (DL-1; R&D Systems Europe), 50 ng/mL SCF, and 50 ng/mL IL-7. Half of the culture medium was changed weekly. Cultures were conducted at 37°C in a humidified atmosphere, 5% CO2. The MS5 stromal cells that express ectopic DL-1 (MS5/DL-1) were also used in some experiments as described [38 –40].
Thymus colonization assays
ESC-derived CD45RA+CD7− and CD45RA+CD7+ sorted populations were independently labeled in PBS with CFSE according to the manufacturer's instructions (Molecular Probes®; Invitrogen). CFSE-labeled cell populations were then resuspended in IMDM, 10% human AB serum. Hanging drops were prepared in Terasaki plates by adding 25 μL of cell suspension to individual thymic lobe freshly isolated from NOG mice embryos (5×102 cells/lobe). Inverted plates were subsequently incubated for 48 h at 37°C, 5% CO2. Lobes were washed by gentle dipping in PBS, transferred onto floating Nucleopore filters (Millipore SA), and immediately processed for microscopy. Specimens were gently layered on glass coverslips that were subsequently inverted onto single concave microscope slides containing 50 μL RPMI 1640 without phenol red and maintained at 37°C throughout the experiment as previously described [8]. Two-photon excitation confocal analysis of samples was carried out with a Leica TCS SP2 AOBS multiphoton CLSM (Leica) as described [8].
Statistical analysis
Data are expressed as mean±SEM. Statistical significance was assessed using the unpaired Student's t-test and Mann–Whitney Rank Sum test. Differences with P<0.05 were considered statistically significant.
Results
In vitro generation of ESC-derived CD34+ HPC sub-populations by a two-step culture procedure
To assess whether ESCs give rise to HPCs endowed with lymphoid potential, we designed a multi-step culture procedure that first comprised the generation of EBs from ESCs (Fig. 1, step I) followed by a co-culture of EB-derived cells with MS5 cells, classically used to study human hemato-lymphopoiesis [41 –43], in the presence of SCF, Flt3-L, and TSLP (Fig. 1, step II). Since the cell surface expression of CD45RA, CD7, CD127 (IL-7Rα), CD10, and CD62L could be considered as a relevant marker of early lymphoid specification of human CD34+ HPCs [3,5,7,8,10,12,32], cells were harvested at the end of the 2-week co-culture period, and analyzed for the expression of these markers. As shown in Fig. 2, co-culture of EB-derived cells with MS5 cells in the presence of cytokines promoted the development, among the CD34+ cells, of CD45RA+ HPCs, and particularly that of CD45RA+CD7− and CD45RA+CD7+ HPCs (Fig. 2A). As already shown [32], this step of co-culture of EB-derived cells with MS5 cells is indispensable to enhance the generation of CD34+CD45RA+ HPCs, as CD45RA+CD7− HPCs derived from day-0 fresh EBs (that is w/o any co-culture) or from co-cultures represent 0.8%±0.6% and 26%±11% of CD34+ cells, respectively (mean±SEM, P<0.001). Similarly, CD45RA+CD7+ HPCs derived from fresh EBs or from co-cultures represented 1.5%±1.6% and 29%±7.8%, respectively (P<0.001) (Fig. 2B). This resulted in an increase of the absolute numbers of CD45RA+CD7− and CD45RA+CD7+ HPCs after co-culture with MS5 cells (Fig. 2B). We next analyzed the expression of the CD10 and CD62L markers within CD34+CD45RA+CD7− and CD34+CD45RA+CD7+ HPCs and found that CD10 was expressed in 12.4%±2.3% of CD34+CD45RA+CD7− HPCs and in 11.3%±3.6% of CD34+CD45RA+CD7+ HPCs (Fig. 2C). We also found that CD62L was expressed in 59.7%±0.5% of CD34+CD45RA+CD7− HPCs and in 42%±13.5% of CD34+CD45RA+CD7+ HPCs (Fig. 2C). Moreover, it is noteworthy that CD45RA− population cells which represented 97.7%±2% of fresh EB-derived CD34+ HPCs represented only 48%±14.7% after co-culture on MS5 cells (P<0.001), which led to a decrease in the absolute number of CD45RA− HPCs after co-culture (Fig. 2B). These CD34+CD45RA− HPCs comprised CD10+ and CD62L+ cells that represented, respectively, 26.4%±6.5% and 15.5%±0.5% of total CD34+CD45RA− HPCs. Finally, HPCs derived from EB cell co-cultures with MS5 displayed a very weak cell surface expression of CD127 and never expressed CD19 (not shown). Taken together, the results show that in our conditions, co-culture of EB-derived cells with MS5 cells, in the presence of cytokines, fosters the emergence of CD34+CD45RA+CD7− and CD34+CD45RA+CD7+ HPCs which express early lymphoid cell surface markers.
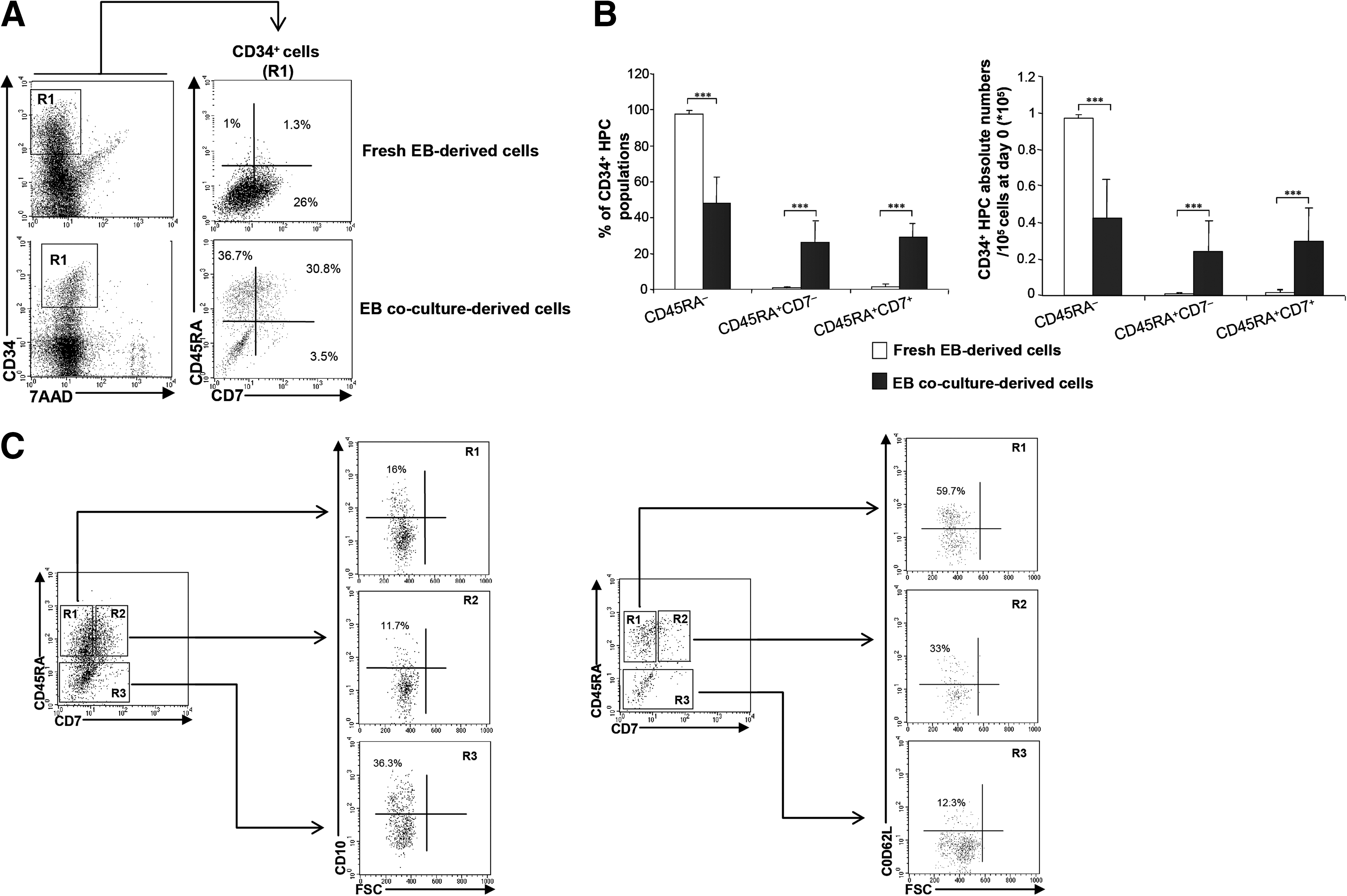
Characterization of CD34+ HPC populations according to CD45RA, CD7, CD10, and CD62L expression.
Molecular characterization of ESC-derived CD34+ HPCs according to CD45RA and CD7 expression
We studied genes associated to early lymphoid lineage, such as IKAROS, RAG1, RAG2, CD127, and DNTT. We also studied B- and T-cell lineage-related genes such as CD79a PAX5, Pre-Tα, TCRγ, TCRβ, and RORC. Finally, we analyzed the expression of PU.1 involved in early lympho-myeloid specification and that of the myeloid gene MPO (Fig. 3A). We compared gene expression levels in CD45RA−, CD45RA+CD7−, and CD45RA+CD7+ HPCs. The expression levels of IKAROS, RAG2, DNTT, and RORC, and in CD45RA+CD7− and CD45RA+CD7+ cell populations were significantly increased in comparison to their expression levels in CD45RA− HPCs but no difference in gene expression level was observed when comparing CD45RA+CD7− and CD45RA+CD7+ cell populations (Fig. 3B). We found no significant difference in the expression of RAG1 when comparing CD45RA+CD7− and CD45RA+CD7+ HPCs (Fig. 3A). The expression level of CD127 in CD45RA+CD7− cell populations was 2.5±0.3 times more elevated than in CD45RA+CD7+ HPCs (P<0.01); inversely, the expression levels of Pre-Tα, TCRβ, and TCRγ in CD45RA+CD7+ HPCs were, respectively, 3.3±0.5, 1.7±0.8, and 1.8±0.9 times more elevated than in CD45RA+CD7− HPCs (P<0.01) (Fig. 3B). CD79a and PAX5 transcripts were detected in CD45RA+CD7− HPCs (Fig. 3A). Furthermore, the relative expression level of PU.1 in CD45RA+CD7+ HPCs was half as much as in CD45RA− HPCs (0.35±0.28, P<0.05), while no significant difference was observed between CD45RA+CD7− HPCs and CD45RA− HPCs (Fig. 3B). Regarding the expression level of MPO, no significant difference was observed between the three subpopulations. Finally, CD45RA−, CD45RA+CD7−, and CD45RA+CD7+ HPCs did not express genes related to the erythro-megakaryocytic lineage such as EPOR, KLF1, GATA1, RUNX1/AML1, and MPL/TPOR (Fig. 3A) [44]. Absolute values of gene expressions are shown in Supplementary Fig. S1. Taken together, these data show that the ESC-derived CD34+ HPCs expressing CD45RA carry a substantial lymphoid molecular profile. Our results show that CD45RA+CD7+ HPCs preferentially exhibit a T-cell molecular profile and that CD45RA+CD7− HPCs are more related to the B-cell lineage. These findings led us to focus more specifically on the identity of CD45RA+CD7− and CD45RA+CD7+ HPCs.
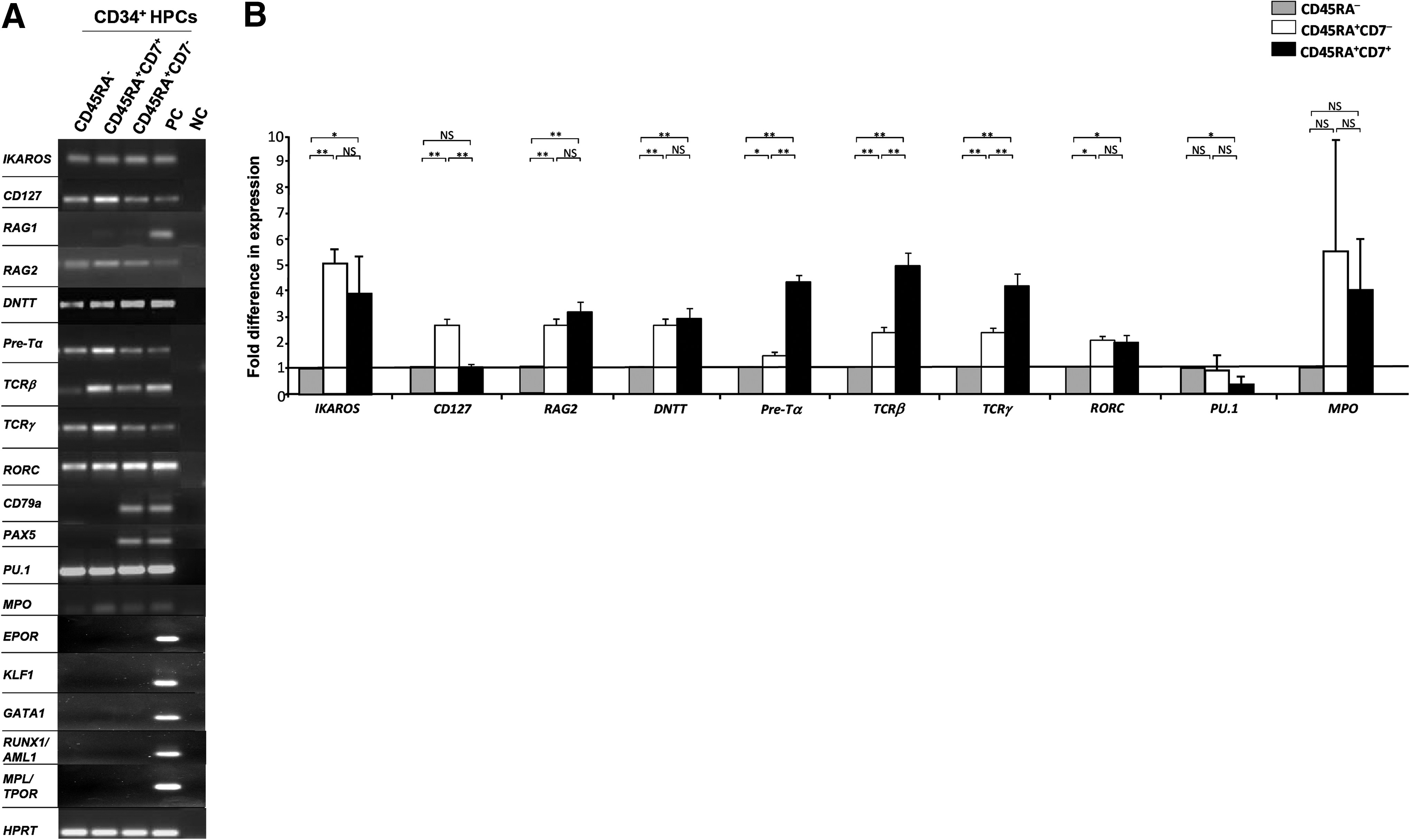
Gene expression analysis of ESC-derived HPCs.
CD34+CD45RA+CD7− and CD34+CD45RA+CD7+ ESC-derived HPCs differ in their capacity to generate NK cells
We first analyzed NK-cell differentiation potential of CD45RA+CD7− and CD45RA+CD7+ HPCs (Fig. 1, step III and Fig. 4A, B). In bulk culture, CD45RA+CD7+ HPCs' capacity at generating CD56+ NK cells was superior to that of CD45RA+CD7− HPCs with 81.2%±17.5% versus 51.1%±11.9% (P=0.05), with 0.6- to 11.6 and 1.4- to 7.5-fold expansion for CD45RA+CD7− and CD45RA+CD7+ HPCs, respectively (Fig. 4A). In limiting-dilution analysis, NK cell precursor frequency varied between 1:89 and 1:75 among CD45RA+CD7− HPCs relative to 1:338 and 1:264 for CD45RA+CD7+ HPCs (Fig. 4B). This difference was due to the overall cloning efficiency under NK condition that varied between 1:98 and 1:40 among CD45RA+CD7− HPCs relative to 1:271 and 1:208. These results underline the fact that CD45RA+CD7− HPCs are more enriched in clonogenic precursors when compared with CD45RA+CD7+, which means that CD45RA+CD7− HPCs are more proliferative than CD45RA+CD7+ HPCs, leading to a higher NK cell precursors' frequency among CD45RA+CD7− than among CD45RA+CD7+ HPCs. However, we found that almost all the clonogenic precursors in NK condition corresponded to NK precursors starting either from CD45RA+CD7− or from CD45RA+CD7+ HPCs. However, the two populations markedly differed regarding their intrinsic capacity to generate NK cells. The analysis of the percentages of NK cells derived from CD45RA+CD7− and CD45RA+CD7+ HPCs in limiting-dilution analysis enables the appreciation of the intrinsic NK-cell differentiation potential of these cells. As shown in Fig. 4B, NK-cell percentages were equal to 76%±13% in wells seeded with 300 CD45RA+CD7+ HPCs relative to 55.7%±19.9% in CD45RA+CD7− HPC cultures (P=0.05), which is in accordance with data obtained from bulk culture under NK condition (Fig. 4A). This analysis of the percentage of NK cells derived from CD45RA+CD7− or CD45RA+CD7+ HPCs gives a qualitative approach of the NK-cell differentiation that enables a better appreciation of the NK polarization of CD45RA+CD7+ HPCs in comparison to CD45RA+CD7− HPCs. Altogether, these data indicate that CD45RA+CD7+ HPCs display a better NK-cell differentiation potential than CD45RA+CD7− HPCs. Interestingly, 100% of wells seeded with 300 CD45RA+CD7− HPCs contained mixed cells (NK and non-NK cells), whereas 60% of cells seeded with 300 CD45RA+CD7+ HPCs contained NK cells only (Fig. 4C), which could suggest that CD45RA+CD7− HPCs are enriched in more immature and proliferative precursors than CD45RA+CD7+.

NK-cell differentiation potential of CD34+CD45RA+CD7− and CD34+CD45RA+CD7+ HPCs.
CD34+CD45RA+CD7− and CD34+CD45RA+CD7+ ESC-derived HPCs differ in their capacity to generate B cells
In bulk culture under B lymphoid conditions (Fig. 1, step III and Fig. 5A, B), the CD45RA+CD7− and CD45RA+CD7+ HPCs displayed a similar capacity to generate CD19+ B cells, representing 25.2%±11.7% and 16%±8.4% of cells, respectively (P=0.1), with 0.8- to 3.5- and 1.9- to 11.4-fold expansion, respectively (Fig. 5A). In limiting-dilution analysis, B-cell precursor frequency varied between 1:163 and 1:57 among CD45RA+CD7− HPCs relative to 1:5177 and 1:302 for CD45RA+CD7+ HPCs (Fig. 5B). This difference is also due to the overall cloning efficiency under B conditions that varied between 1:116 and 1:19 among CD45RA+CD7− HPCs relative to 1:5177 and 1:19 for CD45RA+CD7+ HPCs, showing that under B condition, CD45RA+CD7− HPCs could generate more clonogenic precursors that correspond to B precursors than CD45RA+CD7+ HPCs did. Furthermore, the two populations markedly differed regarding their intrinsic capacity to generate B cells: CD19+ B cells represented 63%±23.2% and 65.4%±20.8% of the cells in wells seeded with 300 and 100 CD45RA+CD7− HPCs as compared with 25.3%±18% and 31.6%±22.2% in CD45RA+CD7+ HPC cultures (P<0.01and P<0.05) (Fig. 5B). Furthermore, 83% of wells seeded with 300 CD45RA+CD7− HPCs contained CD19+ B cells, whose proportion varied between 50% and 96%; whereas only 33% of wells seeded with 300 CD45RA+CD7+ HPCs contained B cells, whose proportion varied between 21% and 60% (Fig. 5C). In the same way, 83% of wells seeded with 100 CD45RA+CD7− HPCs contained CD19+ B cells, whose proportion varied between 30% and 98%; whereas only 33% of wells seeded with 100 CD45RA+CD7+ HPCs contained B cells, whose proportion varied between 8% and 67% (not shown). These results show that CD45RA+CD7− HPCs exhibit a major B-cell differentiation potential. However, the B cells generated from CD45RA+CD7− HPCs were not mature, as they did not express IgM (Supplementary Fig. S2).
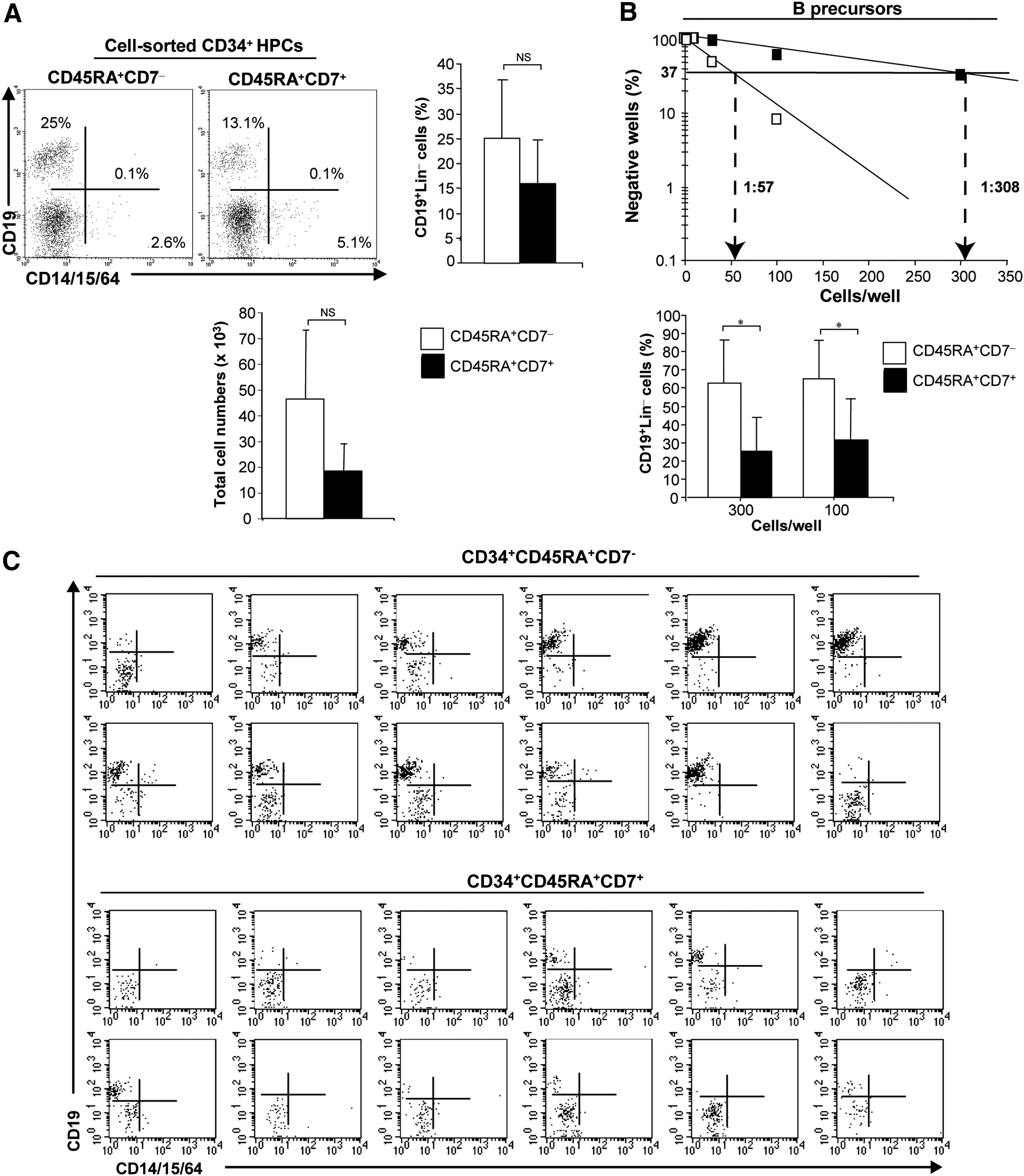
B-cell differentiation potential of CD34+CD45RA+CD7− and CD34+CD45RA+CD7+ HPCs.
Finally, whether CD45RA+CD7− cells represented precursors of CD45RA+CD7+ HPCs was examined. CD45RA+CD7− HPCs were sorted and cultured onto MS-5 cells in the presence of Flt3-L, SCF, and TSLP [7]. After 1 week of co-culture, we could identify the presence of CD45RA+CD7+ HPCs, which indicates that CD7+ HPCs could derive from CD7− cells (Supplementary Fig. S3). This result suggests that CD45RA+CD7− cells are more multipotent than CD7+ HPCs, which is in accordance with the results presented in Fig. 4C.
Taken together, these data indicate that CD45RA+CD7− and CD45RA+CD7+ HPCs are lymphoid progenitors which display mixed NK- and B-cell differentiation potential and that CD45RA+CD7+ HPCs are effectively endowed with an important NK-cell differentiation potential, whereas CD45RA+CD7− HPCs exhibit a high B-cell differentiation potential.
CD34+CD45RA+CD7− and CD34+CD45RA+CD7+ ESC-derived HPCs do not differ in their capacity to generate myeloid cells and DCs
In clonogenic assays, CD45RA+CD7− and CD45RA+CD7+ HPCs gave rise to an equivalent number of colony-forming cells (23±6.7 and 21.8±11.9 respectively, P=0.46) (Fig. 1, step III and Fig. 6A) that corresponded to granulo-macrophagic or macrophagic colonies. In these conditions, no erythroid colonies could be detected, which is in accordance with the lack of erythroid gene expressions. We next examined their capacity to differentiate into classical DC and Langherans cell (LC)-type DCs (Fig. 1, step III and Fig. 6B, C) by monitoring cell surface expression of CD1a, HLA-DR, and CD207. Growth of CD45RA+CD7− and CD45RA+CD7+ HPCs in DC culture condition was equivalent and limited (1.1- to 2.5- and 1.7- to 2-fold expansion, respectively). Furthermore, CD45RA+CD7− and CD45RA+CD7+ HPCs could generate CD1a+HLA-DR+ DCs in the same proportions that is, 63.3%±24.5% and 79%±12.5%, respectively (P=0.2). The proportion of CD1a+CD207+ LCs that developed from CD45RA+CD7− and CD45RA+CD7+ HPCs was also similar and represented 55.9%±7.2% and 40.3%±19.8%, respectively (P=0.2) (Fig. 6B). Finally, expansion of CD45RA+CD7− and CD45RA+CD7+ HPCs in DC culture condition was equivalent (1.1- to 2.5- and 1.7- to 2-fold expansion, respectively) (Fig. 6C).

Myeloid and dendritic cell differentiation potential of CD34+CD45RA+CD7− and CD34+CD45RA+CD7+ HPCs.
Taken together, these results show that CD45RA+CD7− and CD45RA+CD7+ HPCs exhibit an equivalent differentiation potential toward myeloid and DC lineages.
CD34+CD45RA+CD7− and CD34+CD45RA+CD7+ HPCs differ in their capacity to colonize embryonic thymus and to generate T cells
We used CFSE fluorochrome tracking to compare the capacity of CD45RA+CD7− and CD45RA+CD7+ HPCs to enter alymphoid thymus lobes from NOG mice embryos. Two-photon confocal microscopy and reconstructions of serial optical sections showed a massive surface invasion of lobes seeded with CD45RA+CD7+ HPCs, which did not enter the thymus parenchyma; whereas only a few CFSE+ cells were detected at the surface of CD45RA+CD7− HPC-seeded lobes (Fig. 7A). T-cell generation assays were used to assess whether the differences noted between CD45RA+CD7− and CD45RA+CD7+ HPCs extended to the capacity to generate thymocytes (Fig. 1, step III and Fig. 7B). Using MS5 co-culture in the presence of exogenously added DL-1, CD45RA+CD7+ HPCs displayed substantial T-cell potential, with CD4+CD8+TCRαβ−, CD4+CD8+TCRαβ+ and CD4+CD8− TCRαβ+ thymocytes representing on average of about 76.4%±11.5% of hematopoietic cells recovered from co-culture as compared with 41.2%±24.7% from cultures initiated with CD45RA+CD7− HPCs (see representative results in Fig. 7B). However, cell expansion in cultures seeded with CD45RA+CD7− and CD45RA+CD7+ HPCs was similar (1.7 vs. 0.9). Using MS5/DL-1 stromal cells, we confirmed that CD45RA+CD7+ HPCs disclosed a better capacity to generate CD4+CD8+ thymocytes, which were TCRαβ− in comparison to CD45RA+CD7− HPCs that did not generate CD4+CD8+ thymocytes (Supplementary Fig. S4), UCB-derived CD34+ HPCs were used as a positive control of T-cell differentiation (Supplementary Fig. S5). Finally, no TCRγδ+ thymocytes could be detected in our experimental procedure (not shown). Altogether, these results demonstrate that CD45RA+CD7+ HPCs are preferentially endowed with a T-cell differentiation potential.

T-cell differentiation potential of CD34+CD45RA+CD7− and CD34+CD45RA+CD7+ HPCs.
Discussion
This work gives new insights into the early steps of lymphopoiesis from human ESCs. Using the EB culture system followed by co-culture on MS5 stromal cells and based on the expression of CD34, CD45RA, and CD7 markers, we demonstrated here the development of two populations of ESC-derived CD34+ early multi-lymphoid progenitors: the CD45RA+CD7− and CD45RA+CD7+ cells, both of which expressed CD10 and CD62L cell surface markers and typical early lymphoid lineage genes such as IKAROS, CD127, RAG1, RAG2, and DNTT. Interestingly, CD45RA+CD7− HPCs expressed B-cell-lineage associated genes such as PAX5, CD79a. Thus we cannot exclude that among the CD45RA+CD7− HPCs, more committed HPCs such as CD10+(CD19−) (Fig. 2C), which correspond to the phenotypic homologues of UCB and BM-derived CD45RA+CD10+Lin− multipotent early pro-B cells [5,7,8], might probably contribute to the substantial B-cell differentiation potential supported by the ESC-derived CD45RA+CD7− HPCs. On the other hand, CD45RA+CD7+ HPCs express T-cell-lineage associated genes such as Pre-Tα, TRGC1, and TRBC2. It seems that the rearrangement machinery is operational in CD45RA+CD7− and CD45RA+CD7+ HPCs, as we detected an increased expression of RAG1 and RAG2 genes (Fig. 3).
However, the expression of lymphoid-associated genes in CD45RA− cells (Fig. 3), even at a lower level than in CD45RA+ cells, underlines the fact that ESC-derived CD45RA− cells correspond to heterogeneous cell populations, an observation in accordance with previous results on their mixed potential obtained from works on adult hematopoiesis [7,10]. Cell populations are never truly homogeneous; individual cells exist in biochemical states that define functional differences between them and qRT-PCR measure gene expression from pools of cells masking heterogeneities within cell populations, which may be important for understanding underlying biological processes. In contrast, single-cell qRT-PCR measurements would be preferable for the population analysis, because the control population being referenced is impure. New technology based on microfluidic arrays combined with multiplexed quantitative polymerase chain reactions now enables high-throughput single-cell gene expression measurement, enabling assessment of cellular heterogeneity [45,46].
Cell cultures under NK, B, and T conditions showed that although CD45RA+CD7− and CD45RA+CD7+ HPCs could generate cells of the three lymphoid lineages, they were skewed toward B and T/NK cell lineages, respectively (Figs. 3 –5 and 7).
Particularly, the analysis of the percentage of NK cells derived from CD45RA+CD7− and CD45RA+CD7+ HPCs in limiting dilution analysis enabled the appreciation of the intrinsic NK-cell differentiation potential of these cells and showed that CD45RA+CD7+ cells exhibited a more important NK-cell differentiation potential than CD45RA+CD7− cells. This analysis of the percentage of NK cells obtained from CD45RA+CD7− and CD45RA+CD7+ HPCs gives a qualitative approach of the NK-cell differentiation that enables a better appreciation of the NK polarization of CD45RA+CD7+ HPCs in comparison to CD45RA+CD7− cells [7]. Furthermore, the analysis of the clonogenic potential enables a quantitative approach of the NK-cell differentiation, which does not necessary lead to an appreciation of the intrinsic lymphoid potential but gives an idea of the proliferative potential of the cells and to what extent one population could be more immature than another (Fig. 4C). The absence of TCRγδ+ in our T-cell culture (not shown) could be compatible with an instructive role for the TCR in αβ versus γδ lineage commitment [47].
Likewise, we showed that CD45RA+CD7+ HPCs were preferentially recruited into fetal thymic lobes (Fig. 6A). In our hands, ESC-derived MELPs could not generate mature IgM+ B cells (Supplementary Fig. S2), which is in agreement with the work that Carpenter et al. [48] conducted with induced pluripotent stem cells. Moreover, CD45RA+CD7− and CD45RA+CD7+ MELPs were devoid of erythro-megakaryocytic potential but could differentiate along the myeloid pathway, giving rise to granulo-macrophagic cells and DCs (Figs. 3 and 5) and suggesting, in the model of human ESCs, the close association of myeloid and lymphoid potentials in human MELPs as already described for adult multi-lymphoid progenitors [7,8,10 –12]. Interestingly, these two subsets of HPCs could generate LCs without exogenous addition of TGF-β (Fig. 5), strengthening the lymphoid “nature” of CD45RA+CD7− and CD45RA+CD7+ MELPs derived from ESCs [37,49]. Finally, limiting dilution analysis suggested that CD45RA+CD7− HPCs are enriched in more immature and proliferative precursors than CD45RA+CD7+ (Fig. 4). This is also attested by PU.1 expression (Fig. 3B) and by the fact that CD45RA+CD7+ HPCs could be derived from CD45RA+CD7− cells (Supplementary Fig. S3). Furthermore, it is now clear that human multi-lymphoid progenitors are not lymphoid restricted, and possess myeloid, but not erythro-megakaryocytic potential [7,10,12]. Thus, since no erythro-megakaryocytic gene expressions is detectable in the CD45RA− cell populations, and because these cells express lower levels of lymphoid-associated genes in comparison to CD45RA+ cell populations (Fig. 3B), we cannot exclude the fact that CD45RA− cells generated ex vivo from EB cells after a co-culture with MS5 cells could correspond to multipotent progenitors which are specified toward lympho-myeloid lineage and that could be located upstream of CD45RA+CD7− and CD45RA+CD7+ cells.
The potential of human ESCs to differentiate into hematopoietic lineage has been classically demonstrated during EB differentiation or on co-culture with OP9 or S17 BM stromal cell lines. In these conditions, rare data are available regarding the early stages of hemato-lymphopoiesis. Vodyanik et al. performed a detailed analysis of precursors generated from ESCs in OP9 co-culture and described a CD34+CD43+ population committed toward the lympho-myeloid lineage [21,25]. Although T precursor activity was not addressed, these data suggested that the CD34+CD43+ population could contain multipotent lympho-hematopoietic progenitors. However, a report by Martin et al. indicated that such HPCs from ESCs did not derive T cells when co-cultured with OP9 cells expressing DL-1 or in fetal thymus organ culture [50]. Later, Timmermans et al. showed that CD34hiCD43lo cells, present in so-called “hematopoietic zones” generated from ESCs when co-cultured on OP9 stromal cells, could differentiate into T-lineage cells when transferred on OP9/DL-1 monolayers [28]. In the EB system, the work by Galic et al. demonstrated that CD34+CD45+ hematopoietic precursors could efficiently generate thymocytes in the SCID-hu mice model [31]. Furthermore, Kennedy et al. recently identified CD34+CD43lo/− progenitor populations from EB cells, with characteristics of hemogenic endothelium that along with myeloid and erythroid cells, display the capacity to generate T cells [23]. Nevertheless, the multi-lymphoid potential analysis of these HPCs described by Timmermans et al., Galic et al., and Kennedy et al. along with the characterization of human ESC-CD34+CD45RA+ HPCs have not been performed [23,28,31]. With regard to B lymphopoiesis, a study by Dravid et al. reported that B-lymphoid potential is defective in CD34+ cells derived directly from day 8 EB cells [51], which underlines that the culture conditions (EB vs. co-culture) and the time of analysis are major criteria to be taken into account to enable ESCs to disclose their lympho-hematopoietic differentiation potential.
Footnotes
Acknowledgments
The authors would like to thank X. Baudin and A. Jobart-Malfait (technological platform ImagoSeine of Institute Jacques Monod, University Paris Diderot, CNRS) for their expert assistance in photonic imaging, F. Pflumio (Inserm U967, Fontenay aux Roses) for the kind gift of MS5/DL-1 stromal cells, M. Point (SFR Andre Lwoff, Villejuif) for technical assistance, E. Oberlin (Inserm U935, Villejuif) for providing RNA from fetal thymocytes, Ph. Duquesnoy (Inserm U933, Hospital Armand Trousseau, Paris) for the primers design and for providing RNA from BM cells, and L. Renou (Inserm U967, Fontenay aux Roses) for informatic assistance. This work was supported by Inserm, Université Paris Sud, Medicen-Paris Region Ile de France (IngeCELL), PRES UniverSud, and the Association NRB (nouvelle recherche biomédicale) Vaincre le cancer.
Author Disclosure Statement
The authors have no conflicting financial interests.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.