
Abstract
A low-efficiency yield hinders the use of stem cells as a source of endothelial cells (ECs) for therapeutic vascularization, and the diversity of the transforming growth factor-β (TGF-β) superfamily has undermined understanding the effects of its potent vascularization-inducing. Herein, we studied the role of the TGF-β superfamily in EC differentiation of rat bone marrow mesenchymal stem cells (MSCs) induced by Smad2/3 and Smad1/5/8 signaling. MSCs that had been sorted by flow cytometry as CD31-negative were cultured for 14 days in medium supplemented with vascular endothelial growth factor (VEGF) and basic fibroblast growth factor (bFGF) as the control. The Smad2/3 pathway was activated by TGF-β1 and Smad1/5/8 by bone morphogenetic proteins (BMPs). In the early phase in the Smad2/3-activated group, there were 10% CD31-positive cells, which was significantly higher than in the control group. A low Smad1/5/8 phosphorylation level after BMP4 activation doubled the number of CD31-positive cells, while a higher phosphorylation level after BMP9 activation showed no effect. A Smad2/3 inhibitor initially blocked differentiation but later promoted it, while a Smad1/5/8 inhibitor reversed the induction observed with BMPs. Moreover, the positive effects of R-Smad on differentiation were weakened by the VEGF neutralizing antibody, and a Smad3 inhibitor decreased VEGF expression and blocked differentiation in both the early and late phases. In conclusion, differentiation of ECs from MSCs via Smad2/3 signaling is stage dependent. Activation, particularly by Smad3, significantly promotes differentiation at an early phase but later is suppressive. A low Smad1/5/8 phosphorylation level has a positive effect, and R-Smad effects are partly mediated by VEGF.
Introduction
A
Most transforming growth factor-β (TGF-β) superfamily ligands bind to heteromeric complexes of types I and II serine/threonine kinase receptors. Upon ligand binding, the type II receptor phosphorylates and activates the type I receptor activin receptor-like kinase (ALK). The latter, in turn, phosphorylates two classes of receptor Smad proteins (R-Smad), Smad2/3, and Smad1/5/8, and the phosphorylated R-Smads dimerize with Smad4; the complex translocates to the nucleus where it regulates target gene transcription [11,12]. Based on the R-Smads involved, the TGF-β superfamily of ligands is classified into two groups: one including TGF-β, which is involved in activin/nodal signaling and activates Smad2/3 via ALK4, ALK5, and ALK7; and a second one including bone morphogenetic proteins (BMPs), which activate Smad1/5/8 signaling via ALK1, ALK2, ALK3 (BMPR1A), and ALK6 (BMPR1B) [13,14]. Notably, although TGF-β/ALK1/Smad1/5/8 signaling also has been observed in ECs, chondrocytes, and neurons, ALK5 is the predominant type I receptor that mediates responses to TGF-β in most cell types [15 –17]. Although in vitro and in vivo studies offer strong evidence that TGF-β and BMP signaling pathways play important roles in EC differentiation, vasculogenesis, and angiogenesis, both positive and negative effects of TGF-β, BMP9, BMP4, and their ALK5, ALK1, ALK3, and ALK6 receptors have been reported [7,18 –21]. Beyond the remarkable diversity of the TGF-β superfamily of ligands and the multistep, complex, and regulated nature of the vascularization process much of the confusion likely arises from downstream effector R-Smad proteins and origin cells.
Therefore, the aim of this study was to investigate the direct role of Smad2/3 and Smad1/5/8 signaling in endothelial differentiation of MSCs via induction by TGF-β1, BMP4, and BMP9 or by blockage of induction by specific inhibitors.
Materials and Methods
Antibodies and reagents
The following antibodies and reagents were used: low-glucose Dulbecco's modified Eagle's medium (LG-DMEM), fetal bovine serum (FBS), bovine serum albumin (BSA), and trypsin/EDTA (Gibco-BRL); adipogenic and osteogenic induction media (Cambrex); insulin-transferrin-selenium-sodium pyruvate (ITS; Invitrogen); the EGM-2 Bullet Kit (mixture of FGF2, VEGF, heparin, IGF-I, EGF, hydrocortisone, and ascorbic acid; Lonza); rat recombinant VEGF/FGF2 and human recombinant TGF-β1 (99% identity with rat; PeproTech); mouse recombinant BMP4 (98% amino acid identity with rat) and mouse recombinant BMP9 (96% amino acid identity with rat) (R&D Systems); SB431542; dorsomorphin (EMD Biosciences); SIS3 (EMD Millipore); PE-CY7 CD29 (eBioscience); Alex Fluor647 CD90 (AbDSerotec); PE CD34 (Santa Cruz Biotechnology); PE-CY™5 CD45 and PE CD31 (BD Pharmingen); mouse anti-rat KDR and rabbit anti-rat Von Willebrand factor (vWF; Abcam); goat anti-mouse IgG-Alexa488 and goat anti-rabbit IgG-Alexa Fluor 594 (Invitrogen); 4,6-diamidino-2-phenylindole (DAPI; Molecular Probes); reverse transcription–polymerase chain reaction (RT-PCR) kit, Q-PCR kit, and TRIzol (Takara); protein lysis buffer kit (KeyGen Biotech); anti-p-Smad1/5/8, anti-p-Smad3, anti-Smad3, anti-p-Smad2, anti-Smad2, and control IgG (Cell Signaling Technology); anti-GAPDH and horseradish peroxidase (HRP)-conjugated goat anti-rabbit IgG (CW Biotech); Nitric Oxide (NO) Test Kit (Nanjing Jiancheng Bioengineering Institute); Alexa Fluor 594-acetyl-low-density lipoprotein (LDL) (Invitrogen); anti-VEGF neutralizing antibody AF564; and quantitative sandwich enzyme-linked immunosorbent assay (ELISA) kits (R&D Systems).
Bone marrow MSC isolation and expansion
Young male Sprague–Dawley rats were used for bone marrow MSC isolation. After administration of sodium pentobarbital anesthesia (50 mg/kg, given intraperitoneally), both femurs were excised and an 18-gauge needle attached to a 1.0-mL syringe was inserted at the epiphysis. The bone marrow was aspirated and resuspended in LG-DMEM containing 15% FBS at a cell concentration of 5×107 cells/mL. The cells were then plated in 25-cm2 flasks (Corning, Inc.) and incubated with 5% CO2 at 37°C. After incubation for 48 h, the nonadherent hematopoietic cells were removed by a wash with medium. During the expansion phase, the medium was renewed every 3 days. When the rBMSCs became confluent, they were detached with 0.25% trypsin/0.02% EDTA and replated at a dilution of 1:3. After three passages, a homogeneous population of rBMSCs was generated.
MSC adipogenic and osteogenic differentiation potential
To confirm the multipotency of the cultured MSCs, we tested whether cells in passage 3 could differentiate into adipocytes and osteoblasts, as reported previously [2].
Flow cytometric analysis and sorting
The immunophenotypes of MSCs were identified using an antibody panel in combination with flow cytometry. Antibodies against CD29 and CD90 specific for MSCs, with the exclusion of CD34 and CD45 specific for hematopoietic cells, were used. MSC-derived ECs were identified using a CD31 antibody, which was also used for cells sorting by flow cytometry. Isotype-identical antibodies served as controls. Analyses and sorting were performed using a BD FACS Calibur flow cytometer and CellQuest software (Becton Dickinson). Histograms of cell numbers versus fluorescence intensity were recorded for further analysis.
Differentiation of MSCs into the endothelial lineage
MSCs (CD31 negative) were plated at a cell density of 2×104 cells/cm2 and cultured in the presence of medium containing EGM-2, 2% ITS, and EGM-2 Bullet Kit reagents with the VEGF and bFGF concentrations adjusted to 10 and 2 ng/mL, respectively, at 37°C for 14 days. Human TGF-β1 (10 ng/mL), mouse BMP4 (20 ng/mL), and mouse BMP9 (10 ng/mL) were added to the cells at the time of induction, and the cells were cultured for 4, 7, 10, and 14 days, respectively. Next, to block Smad2/3, Smad1/5/8, and Smad3 phosphorylation, 10 μM SB431542, 5 μM dorsomorphin, and 3 μM SIS3 were added to the corresponding differentiation medium. The medium were changed every 3 days. The gene and cell surface markers for EC expression were assessed.
Immunofluorescence study
Differentiated endothelial-like cells from rat bone marrow MSCs were fixed with 4% paraformaldehyde for 15 min, permeabilized with 0.1% Triton X-100 for 20 min, and immersed in a solution of 5% BSA for 40 min at room temperature to block nonspecific binding. Subsequently, the cells were probed with the primary antibodies mouse anti-rat KDR and rabbit anti-rat vWF at 4°C overnight. After washing, the cells were incubated with the secondary antibody goat anti-mouse IgG-Alexa488 or goat anti-rabbit IgG-Alexa594 for 1 h, and then DAPI (0.1 mg/mL) was used for cell nuclear staining for 3–5 min at room temperature. Photographs were taken with a DP70 digital camera (Olympus) and analyzed by Image-Pro Plus software (Media Cybernetics).
RT-PCR and real-time PCR analyses
Total RNAs from undifferentiated/differentiated MSCs at different time points were extracted using the TRIzol kit and subjected to reverse transcription into cDNA. PCR amplifications of specific DNA fragments were performed in triplicate with TaqDNA polymerase and specific primers. All RNA samples were adjusted to yield equal amplification of β-actin as an internal standard. For real-time PCR, synthesized cDNA was amplified by a standard PCR protocol with SYBR Green Super Mix (Bio-Rad) and rat-specific primers. Parallel amplifications with primers for β-actin were performed. The cycling conditions were as follows: preincubation at 95°C for 3 min, followed by 40 cycles of denaturation at 95°C for 30 s, annealing at 55°C for 30 s, and extension at 72°C for 30 s with an iCycler instrument (Bio-Rad). The oligonucleotide primers and PCR conditions used are listed in Table 1.
TGF-βR2, transforming growth factor-β type II receptor; BMPR1A, bone morphogenetic protein type I receptor A; BMPR2, bone morphogenetic protein type II receptor; ALK1, activin receptor-like kinase-1; ALK5, activin receptor-like kinase-5; KDR, kinase insert domain receptor; PECAM-1, platelet/endothelial cell adhesion molecule-1.
Vascular Endothelial Growth Factor Enzyme-Linked Immunosorbent Assay
MSCs were plated at a density of 20,000 cells/cm2 in 12-well plates and cultured with EGM-2 Bullet Kit reagents, including 2% ITS and cytokines or inhibitors, according to the group. The medium was replaced every other day. The VEGF levels were assessed in the medium using a quantitative sandwich ELISA kit, according to the manufacturer's instructions. The standard induction medium acted as a negative control.
Western blotting
Different groups of induced MSCs were lysed in lysis buffer, and the protein concentrations were determined using a Bradford protein assay (Bio-Rad), separated by 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (60 μg/lane) and transferred to polyvinylidene difluoride membranes (Millipore). After blocking with 5% BSA in Tris-buffered saline containing 0.05% Tween-20 for 1 h, the membranes were incubated overnight at 4°C with anti-p-Smad1/5/8 (1:500), anti-p-Smad3 (1:500), anti-Smad3 (1:500), anti-p-Smad2 (1:500), anti-Smad2 (1:500), anti-GAPDH (1:1000), or 5 mg/mL control IgG (1:500), respectively. The bound antibodies were detected by HRP-conjugated IgG (1:2500) and visualized by enhanced chemiluminescence (Kodak). The levels of each protein relative to the control GAPDH were determined by densitometric analysis.
Characterization of cell activity in vitro and in vivo
Tubular formation assay
A total of 200 μL of Matrigel (BD Biosciences) was added to each well of a 24-well plate on ice and allowed to gel in a thin layer for 1 h at 37°C. Cells (25,000 per well) were seeded on the gel surface and photographed after 6, 24, and 48 h, respectively.
NO secretion
NO levels were indirectly assessed by a Nitric Oxide Test kit (Nanjing Jiancheng Bioengineering Institute, Jiangsu, China), according to the manufacturer's instructions.
Incorporation of Dil-acetyl-LDL
Induced cells were grouped into CD31-negative and CD31-positive cells by flow cytometric sorting. Then, the cells were incubated at 37°C with 2.0 μg/mL Alexa Fluor 594-conjugated acetyl-LDL for 4 h, followed by fluorescence microscopy analysis.
Subcutaneous cultures of tissue-engineered blood vessels
Acellular matrix vascular grafts were rinsed in EGM-2 overnight before cell seeding. MSCs obtained by high-density centrifugation were seeded into acellular matrix vascular grafts and cultured in vitro according to assignment to negative control, standard control, or VFT induction groups for 2 weeks. Sprague–Dawley rats (250 g) were anesthetized with sodium pentobarbital (50 mg/kg, given intraperitoneally). Then, two small dorsal subcutaneous pockets were created near each hind limb. One graft was placed into each pocket, and each group contained five rats. The incision was closed with a 4-0 nonabsorbable suture. A week after implantation, the rats were euthanized by an overdose of pentobarbital. Cell grafts were removed from the subcutaneous space with some surrounding tissue, fixed in 4% formaldehyde, and processed for immunochemical analysis.
Animal research ethics
The experimental protocols were approved by the Sun Yat-sen University Animal Care and Use Committee (approval ID: SCXK Guangdong 2009-0011). This study was approved by the Research Ethics Committee of the First Affiliated Hospital, Sun Yat-sen University.
Statistical analysis
Data are expressed as mean±standard error of the mean, statistical analysis was performed by SPSS 13.0 (SPSS, Inc.) and GraphPad Prism 5 software (GraphPad). The differences among groups were assessed by analysis of variance. When the f-statistic was significant, the least significant difference procedure was used to compare differences between two groups. The level of significance was set at a P-value less than 0.05.
Results
Isolation and identification of MSCs
MSCs were isolated using the plastic adherence method described by Friedenstein et al. [22]. Cells in passage 3 (P3) showed a fibroblast-like, spindle-shaped morphology (Supplementary Fig. S1A; Supplementary Data are available online at
Differentiation of MSCs into endothelial-like cells
First, CD31 (also known as platelet/EC adhesion molecule-1; PECAM-1)-negative MSCs were selected by flow cytometric sorting. Next, EGM-2 Bullet Kit medium was added (2% ITS replaced FBS) with the VEGF and bFGF concentrations adjusted to 10 and 2 ng/mL, respectively, as standard induction conditions. In the latter conditions, expression of the early EC marker VEGF receptor 2 (KDR or Flk-1) [23,24] became apparent after 1 day of induction and reached a peak at day 7 (95%±4%), retaining the latter level. The expression of vWF, a marker of mature ECs [25], started at day 7, continued to increase to approximately 72%±3% until day 10 (Fig. 1A), and began to decline at day 14 (data not shown). The MSCs exhibited no detectable morphological change until 2 weeks after induction (Fig. 1B). Furthermore, the MSCs showed tube-forming potential in Matrigel at 14 days after induction (Fig. 1C). Finally, we examined the functionality of the induced cells by measuring the LDL uptake. The differentiated CD31-positive cells showed the ability to uptake Alexa594-conjugated acetyl-LDL (Fig. 1D).
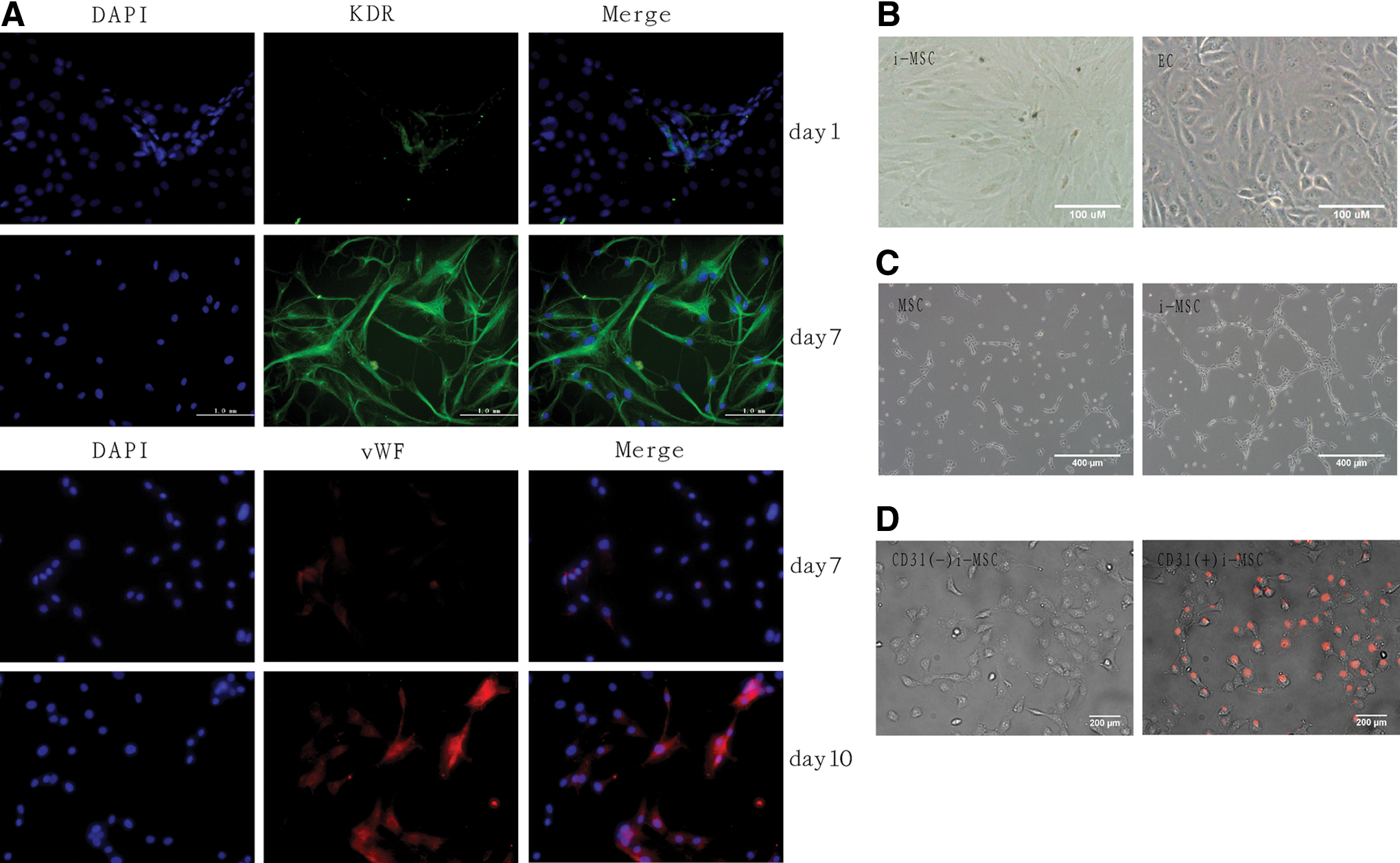
Dynamic changes in marker expression, morphology, and function of MSCs as they differentiate into endothelial-like cells after standard induction.
Kinetics of mRNA expression of TGF-β superfamily receptors and EC markers during differentiation
To study TGF-β receptor expression, RT-PCR and real-time PCR analyses were performed on RNA extracted from CD31-negative MSCs during days 0 to 14 of differentiation; the type II TGF-β1 receptor and type II BMP receptor were present with no difference in undifferentiated MSCs throughout the differentiation process (Fig. 2A). In contrast, the other TGF-β type I receptor, ALK1, showed very weak expression in undifferentiated MSCs, increasing up to day 5, and decreasing on day 14 (Fig. 2B). In terms of the two endothelial markers KDR and PECAM-1, KDR gene expression was found in CD31-negative MSCs, reaching a plateau on day 2; whereas PECAM-1 expression was apparent on day 2, peaked on day 5, and then declined notably on day 7 (Fig. 2B).

Addition of TGF-β1 or BMP4 at an earlier phase and SB431542 at a later phase promotes EC differentiation
Based on standard induction conditions, Smad2/3 was activated by 10 ng/mL TGF-β1, and Smad1/5/8 was activated by 20 ng/mL BMP4 or 10 ng/mL BMP9 for 4, 7, 10, and 14 days. MSCs cultured in standard medium (VF group) served as the control. The percentage of CD31-positive (PECAM-1-positive) cells on day 14 was analyzed. We observed that cells cultured in the presence of TGF-β1 during the first 7 days (VFT group) showed the highest percentage of CD31-positive staining (9.65%±2.75%), namely 7.7-fold higher (P<0.01) than the control (1.25%±0.12%) (Fig. 3A, B). The percentage of CD31-positive cells was 2.56%±0.35% when the cells were cultured with BMP4 for 14 days (VFB4 group), twice that of the control group (P<0.05). The percentage of CD31-positive cells decreased slightly (but not significantly) when BMP9 was added to the induction medium (Fig. 3A, B). Combining the data of TGF-β superfamily receptor expression and endothelial markers during differentiation, we divided the induction process into two distinct phases. The early phase spans the first 7 days of vascular progenitor cell differentiation from MSCs. The second or later phase, taking place during the next 7 days, corresponds to endothelial-like cell differentiation from vascular progenitor cells. Because the positive effect of TGF-β1 occurs only at an early phase, we added the Smad2/3 inhibitor SB431542 at different time points. CD31-positive cells were undetectable when SB431542 was added at day 0 (data not shown). This finding suggests that the early phase of endothelial differentiation from MSCs depends on Smad2/3 signaling, which is in accordance with the literature [26]. When SB431542 was added at the later phase (VFS group), it enhanced the percentage of CD31-positive cells to 2.21%±0.05%, that is, 1.8-fold higher (P<0.01) than in the VF group (Fig. 3A, B). Conversely, the addition of TGF-β1 slightly reduced the number of CD31-positive cells, with no significant difference versus the VF group at the later phase (data not shown). To demonstrate the function of the cultured endothelial-like cells, we compared NO production in cell culture supernatant fluids obtained 14 days after induction. The VFT and BMP4 groups had significantly higher NO production than the control group. In addition, the VFS group had a higher NO production than the control group, but the difference did not achieve statistical significance (Fig. 3C).

Regulation R-smad induces EC differentiation.
VEGF expression by MSC-derived endothelial-like cells
VEGF is a direct TGF-β target gene that has been previously shown to be endogenously expressed by lung fibroblasts and by proximal tubular epithelial cells [27,28]; furthermore, it is involved in BMP-mediated induction of osteoblast angiogenesis [29]. Thus, we tested the effects of TGF-β1, BMP4, BMP9, and SB431542 on VEGF expression in MSC endothelial differentiation. VEGF quantification in culture supernatant fluids of each group was performed by ELISA at days 0, 1, 2, 3, 4, 6, 8, 10, and 12. Compared with the VF group, the early-phase culture of MSCs with TGF-β1 had significantly enhanced VEGF secretion at all time points. Interestingly, the addition of SB431542 at the later phase enhanced VEGF secretion at days 8, 10, and 12 (Fig. 4A). BMP9 stimulation significantly enhanced VEGF expression during the first 2 days, while it notably decreased VEGF expression later. BMP4 significantly stimulated VEGF secretion during the first 4 days and maintained levels higher than those in the VF group over time (Fig. 4B).

VEGF production during differentiation. VEGF levels were measured in supernatant fluids by ELISA at the time points indicated, VEGF production was in accordance to impact on differentiation in each group;
Smad2/3 and Smad1/5/8 regulate EC differentiation of MSCs partly via VEGF production
To examine the effects of R-Smad expression, endogenous VEGF production, and EC differentiation, MSCs were cultured with standard induction medium for 3 days and were further treated with TGF-β1, BMP4, or BMP9 for 1–6 h. TGF-β1 induced Smad2/3 phosphorylation particularly at 1 h after induction (Fig. 5A). The addition of BMP4 or BMP9 led to an increased Smad1/5/8 phosphorylation level, with the change being particularly notable at 6 and 1 h, respectively. However, the effect of BMP9 was more remarkable (Fig. 5A, B). Moreover, the phosphorylation levels of Smad1/5/8, Smad2, or Smad3 did not show any changes after induction by VEGF and bFGF (Fig. 5B). TGF-β1 did not significantly affect Smad1/5/8 phosphorylation, which was also the case for BMP4, BMP9, and Smad2/3 phosphorylation (Fig. 5C). As expected, when 10 μM SB431542 was incubated in the standard induction medium for 1 h, the levels of Smad2 and Smad3 phosphorylation were decreased and the phosphorylation of Smad1/5/8 was not affected (Fig. 5C). Next, we added an anti-VEGF neutralizing antibody (AF564: 1.0 μg/mL) to the VFT and VFB4 groups, respectively. As shown in Fig. 5D and E, anti-VEGF significantly reduced the VEGF concentration in both the VFT and VFB4 groups (407.3±21.1 vs. 379.1±16.7 pg/mL, respectively); a reduction in the percentage of PECAM-1-positive cells (2.73% vs. 1.15%, respectively) was also observed, and the percentage in the VFT group after treatment was higher than that in the VF group. These data demonstrate that the effect on differentiation for each group is partly mediated by VEGF. Finally, to further define the roles of Smad2/3 and Smad1/5/8 in VEGF production and EC differentiation, we added the Smad2/3 inhibitor SB431542 to the VF group or the Smad1/5/8 inhibitor dorsomorphin to the VFB4 and VFB9 groups. SB431542 reduced VEGF secretion (day 1) compared to the VF group (457.4±16.1 vs. 755.1±35.3 pg/mL), and CD31-positive cells were hardly detected (data not shown), consistent with previous reports, indicating that SB431542 can block the onset of EC differentiation [26]. In contrast, dorsomorpin blocked the inhibitory effect of BMP9 and the promoting effect of BMP4 on VEGF production and EC differentiation (Fig. 5F, G).

R-Smad phosphorylation effect on the regulation of endothelial cell differentiation partly via VEGF production.
Smad3 phosphorylation plays a positive role in endothelial differentiation of MSCs by promoting VEGF production
To further demonstrate the effect of Smad2/3 signaling on endothelial differentiation of MSCs and to identify the contributions of Smad3 and Smad2, we investigated the impact of Smad3 on VEGF protein production and endothelial differentiation. First, 3 μM SIS3, an inhibitor of Smad3 phosphorylation, was added to both the VF and VFT groups at the early phase and to the VF group at the later phase, respectively, and at 1 h before cytokine addition. As expected, SIS3 only decreased Smad3 phosphorylation (Fig. 6A). The VEGF concentration was significantly reduced in the VFT group when SIS3 was added at day 0 and incubated with the cells for 24 h (Fig. 6B). Similar findings were observed in both the VFS and VF groups when they were treated with SIS3 at day 8 for 24 h (Fig. 6C). Consistent with the impact on VEGF production, the percentage of PECAM-1-positive cells after SIS3 addition decreased notably in the VFT group during the early phase (from 9.6% to 1.1%; P<0.01, Fig. 6D). When SIS3 was added at a later phase, the percentage was further significantly reduced to 0.8%, compared to the VF and VFS groups (Fig. 6E). Taken together, these data show that Smad3 plays a key role in the promotion of Smad2/3 signaling at the early phase and that the positive effect of SB431542 at the later phase was not secondary to Smad3 inhibition.

Smad3 phosphorylation showed positive effect on the regulation of endothelial cell differentiation and VEGF production.
Application to tissue-engineered blood vessels with optimized endothelial induction
Tissue-engineered vessels with optimized induction (VFT group) had the highest proportion of ECs, with (+++) vWF expression, which was higher than the standard control group (++) and the MSC group (+). Moreover, the intimal thicknesses of undifferentiated MSCs, induced by VEGF, bFGF, and VFT, were 606.0±21.76, 358.0±26.45, and 10.2±2.36 μm, respectively, indicating that the optimized induction grafts had significantly reduced intimal hyperplasia (Fig. 7).

Tissue-engineered blood vessels with optimized endothelial induction improved endothelialization subcutaneously in vivo. MSC versus VF: P<0.002, VF versus VFT: P<0.0005. MSC: without induction group; VF: standard control group (VEGF+bFGF); VFT: experimental group (VEGF+bFGF+TGF-β1 [days 0–7]). Color images available online at
Discussion
This study was designed to evaluate the efficiency of endothelial differentiation of MSCs via Smad2/3 and Smad1/5/8 signaling and to define their roles. To the best of our knowledge, this is the first study to show the possibility of inducing MSC differentiation into the endothelial phenotype, generating a higher yield by targeting R-Smad. Our findings indicate that the effect of Smad2/3 signaling in the differentiation of MSCs into ECs is stage dependent, with activation especially by Smad3 being required and positive at the early phase. Inhibition at the late phase of induction can also promote differentiation. The effect of Smad1/5/8 is related to the phosphorylation level, with a low level considered as positive and a high level as negative. Furthermore, all effects are mediated by the regulation of endogenous VEGF expression by the cells. The current study pointed out an effective and viable means to generate therapeutically relevant numbers of ECs, offering a valid method to fulfill the need for therapeutic vascularization and tissue-engineered grafts.
We evaluated the effects of TGF-β1, BMP4, and BMP9 on MSC endothelial differentiation by quantifying the PECAM-1 (CD31)-positive cell percentage, excluding any spontaneous differentiation into PECAM-1-positive cells. For this very reason, the differentiation factors were included in the induction medium for 4, 7, 10, and 14 days from the initiation of induction. Our findings showed that the presence of TGF-β1 at 0–7 days increased the PECAM-1-positive cell population and contributed to Smad2/3 phosphorylation activation. Furthermore, we demonstrated that blocking Smad2/3 phosphorylation with SB431542 at the very start of the cell culture led to almost total blockade of the differentiation process. These data demonstrate that Smad2/3 phosphorylation induces early MSC endothelial differentiation. Previous reports have shown that the ALK5 receptor, one of the main activators of Smad2/3 phosphorylation, is provasculogenic at early developmental stages in an embryonic stem cell differentiation model [19]. Moreover, we demonstrated that Smad2/3 signaling pathways cause an opposite effect during the later differentiation stage, similar to what is seen with ECs in blood-vessel formation [26,30]. In contrast to Smad2/3, the effect of Smad1/5/8 during EC differentiation is dependent on the phosphorylation level. At a lower level of phosphorylation, BMP4-activated Smad1/5/8 promoted EC differentiation, while a higher level of phosphorylation activated by BMP9 had an opposite effect. The addition of the Smad1/5/8 phosphorylation inhibitor dorsomorphin throughout the culture period reversed both effects significantly. Furthermore, the induced endothelial-like cells acquired tubule-forming function in vitro.
To obtain further insight into the mechanisms determining EC differentiation from MSCs, we distinguished among the differentiation stages by investigating the expression of relevant EC markers. The receptor 2 of VEGF Flk1 is expressed in early mesodermal cells before any morphological evidence of EC differentiation, and it is relatively stable; therefore, it is defined as the earliest EC differentiation marker [23,24]. Conversely, vWF and PECAM-1 are relatively mature EC markers [25]. MSC immunofluorescence staining during the course of differentiation showed that KDR expression appeared on day 1 and rose steadily for 6 days, consistent with a previous report [25]. vWF expression was seen on day 7, then rose rapidly, remained elevated through day 10, and then declined at day 14. Therefore, we divided EC differentiation into two phases: an early phase, spanning the first 7 days, when differentiation of vascular progenitor cells from MSCs occurs; and a later phase, spanning the following 7 days, as differentiation into ECs progresses from vascular progenitor cells.
Previous reports have suggested that VEGF could be a target gene for TGF-β1 [19,31]. Deckers MM et al. [29] found that BMP2/4/6 stimulates angiogenesis through VEGF in the preosteoblast-like cell line KS483, while TGF-β inhibits angiogenesis while stimulating VEGF production. In contrast, BMP9/10 inhibits angiogenesis by inhibiting VEGF and bFGF [32]. To investigate whether TGF-β1, BMP4, or BMP9 and their effects were mediated by R-Smads via VEGF, we evaluated VEGF production during differentiation. We found that VEGF production was in accordance with the impact on differentiation in each group. In contrast to the findings by Mallet et al. [19], we demonstrated that the anti-rat VEGF antibody inhibited the promotion effects in both the VFT and VFB4 groups. The contradictory results may be due to the specificity and efficiency of the VEGF neutralizing antibody [33]. AF-293-NA had been chosen by Mallet et al. [19] as a target for human but not mouse target cells (there was approximately 10% cross-reactivity with recombinant mouse VEGF165). Our results suggest that the effects of TGF-β, BMPs, Smad2/3, and Smad1/5/8 on endothelial differentiation are partly mediated by VEGF expression.
Previous studies have shown that TGF-β/Smad2 induces the production of antiangiogenic factors, while the TGF-β/Smad3 pathway induces VEGF expression in proximal tubular epithelial cells and breast cancer cells [28,34]. Therefore, we hypothesized that Smad2/3 may exert a biphasic role in EC differentiation; one can separate the roles of Smad2 and Smad3 in EC differentiation and focus on the role of Smad3. The addition of a Smad3 phosphorylation inhibitor appeared to block EC differentiation in both culture phases by sharply reducing VEGF production. Smad2/3, mainly mediated by Smad3 phosphorylation, has a positive effect in the early phase; however, Smad2 has a negative role that dominates at the later phase. The positive effect of SB431542 might be secondary to blockage of Smad2 phosphorylation; however, the mechanism requires further investigation. In the future, it also might be interesting to study whether Smad1/5/8 have distinct effects. Moreover, if Smad inhibitors (Smad6/7) or Smads with DNA-specific transcription factors, transcription coactivators, or corepressors have any specific effects over EC differentiation of MSCs should be analyzed. It is also relevant to explore whether dominant crosstalk exists between R-Smads and other signaling pathways such as MAPK/ERK, which has a VEGF-inducing effect [35], in addition to TAK1, Jnk, and PI3K signaling pathways activated or inhibited by TGF-β and BMPs [30,36,37].
In conclusion, our data strongly suggest that in the differentiation of ECs from MSCs, the role of Smad2/3 is stage dependent. The activation of this pathway, especially by Smad3 phosphorylation, is required and positive at the early phase; while its inhibition can increase the proportion of ECs at the later phase. The effect of Smad1/5/8 is related to the phosphorylation level; a low level promotes EC differentiation, while a high one may be inhibitory. All positive effects are, at least partially, mediated by VEGF production (Supplementary Fig. S2). Furthermore, tissue-engineered blood vessels with an optimized endothelial induction program show better endothelialization both in vitro and in subcutaneous cultures. Our approach for EC monitoring and differentiation may help to identify not-yet recognized EC differentiation modulators for preclinical studies aimed at facilitating vascularization by tissue-engineered grafts.
Footnotes
Acknowledgments
This study was supported by grants from the Yat-sen Innovative Talents Cultivation Program for Excellent Tutors (80000-3126202) and the Guangdong Provincial Science and Technology Fund of China (2012B031800087).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.