
Abstract
The 3′ untranslated region (UTR)-associated RNAs (uaRNAs) have important roles in various biological processes, especially in development. However, since they overlap with protein-coding mRNAs, uaRNAs are difficult to study by RNA interference techniques. We recently identified a chemical molecule, 3-benzyl-5-((2-nitrophenoxy) methyl)-dihydrofuran-2(3H)-one (3BDO), that could efficiently induce human embryonic stem cells (hESCs) differentiation, and meanwhile selectively and efficiently downregulate the uaRNA FLJ11812. By acting as a competing endogenous RNA, downregulated FLJ11812 by 3BDO further increased miR-4459 level in hESCs. miR-4459 could decrease the expression of its targets, CDC20B and ATG13, and thus altered stemness via cell cycle and autophagy. Our results revealed that FLJ11812 played a key role in maintenance of stemness of hESCs for the first time. The findings provide new clues and a powerful tool for investigating the action mechanism of FLJ11812 in early development.
Introduction
I
Many uaRNAs have been identified, but the cellular functions of most remain to be determined [1]. Due to the unique position of uaRNAs in the transcript—overlapping protein-coding mRNAs—designing specific siRNA sequences for their knockdown or knockout is difficult. In some cases, only the candidates that could be knocked down by using RNA interference (RNAi) were selected for further study [5]. Therefore, the functions of some important RNAs are not known.
To address this question, we identified a small molecule, 3-benzyl-5-((2-nitrophenoxy) methyl)-dihydrofuran-2(3H)-one (3BDO), that could selectively and efficiently downregulate the uaRNA FLJ11812, derived from the 3′UTR of transforming growth factor, beta 2 (TGFB2) [6]. Homo sapiens cDNA FLJ11812 has been completely sequenced and characterized and was found expressed in whole embryos, mainly in the head [7]. In our previous work, we found that 3BDO activated a serine/threonine kinase:mechanistic target of rapamycin (MTOR) by targeting FKBP1A and further phosphosphorylatd TIA1, which was responsible for the processing of FLJ11812 by interacting with TGFB2. TIA1 phosphorylation reduced this interaction, and then selectively and efficiently downregulated FLJ11812 in human umbilical vein endothelial cells (HUVECs). Furthermore, FLJ11812 acted as a competing endogenous RNA (ceRNA) that regulated miR-4459 and the expression of its target gene-ATG13 thus regulated autophagy level in HUVECs [6].
In previous reports, a ceRNA, linc-RoR, was found to regulate self-renewal and differentiation of human embryonic stem cells (hESCs) [8]. ESCs, derived from the inner cell mass of blastocysts, are established models for investigating embryonic development and cell differentiation, especially in humans [9,10]. The uaRNAs can be independently expressed as developmentally regulated non-coding RNAs (ncRNAs) and may participate in regulating differentiation and developmental processes [1]. However, the functions and molecular mechanisms of FLJ11812 during embryonic development and maintenance of hESCs stemness have not been elucidated. In this study, we used hESCs as models to study the role of FLJ11812 in maintaining the stemness and differentiation of hESCs by using 3BDO to selectively inhibit FLJ11812.
Materials and Methods
Cell culture
hESC (H1 cell line) was obtained from Prof. Jiaxi Zhou (Chinese Academy of Medical Sciences and Peking Union Medical College, Tianjin, China) and maintained as routine under a feeder-independent culture system as described [11]. Experiments involved H1 cells between passages 40 and 60. HEK293 cells were grown in Dulbecco's modified Eagle's medium (Gibco) with 10% fetal bovine serum (Gibco), penicillin (50 U/mL), and streptomycin (50 μg/mL) (Gibco; 10378-016).
Fluorescent in situ hybridization
FLJ11812 expression in hESCs was detected by use of a FISH kit (Roche Applied Science). Briefly, hESCs treated with or without 3BDO were fixed in 4% paraformaldehyde, then prehybridized with hybridization solution, and incubated with a digoxigenin-labeled FLJ11812 probe. Cell nuclei were stained with 4′,6-diamidino-2-phenylindole (DAPI) for 5 min at room temperature. Fluorescence images were obtained by use of an LSM 700 Confocal Laser Scanning Microscope (Carl Zeiss).
Quantitative real-time polymerase chain reaction
Total RNA was extracted from ESCs by the Trizol reagent method (Invitrogen) and underwent quantitative reverse transcription PCR (RT-PCR) (Roche; Light Cycler 2.0 system) with the primer pair sequences for genes (Supplementary Table S1; Supplementary Data are available online at
Western blot analysis
Treated hESCs were lysed in protein lysis buffer (Beyotime; P0013), and protein content was determined by use of the BCA Protein Assay Kit (Beyotime; P0011). Proteins were separated by 15% or 12% SDS-PAGE and transferred to PVDF membrane (Millipore; IPVH00010), which was incubated with primary antibodies (1:1,000) for octamer-binding transcription factor 4 (Oct4; Abcam, ab18976), Nanog (Abcam; ab80892), and sex determining region Y (SRY)-box 2 (Sox2; Abcam, ab97959); ATG13 (Sigma; SAB4200100); CDC20B (Santa Cruz Biotechnology; sc-137387), and ACTB (Santa Cruz Biotechnology; sc-47778), or LC3B (Cell Signaling Technology; 2775) at 4°C overnight and detected with corresponding horseradish peroxidase (HRP)-conjugated secondary antibody (1:10,000) at room temperature for 1 h. The membrane was incubated with Immobilon Western Chemiluminescent HRP Substrate (Millipore; WBKLS0500) for 5 min at room temperature and exposed to X-ray film (Kodak). The relative protein content was analyzed by use of ImageJ software and normalized to loading controls.
Cell cycle analysis
hESCs were treated with or without 3BDO for 72 h, then trypsinized by adding 0.05% Trypsin/EDTA solution at 37°C for 10 min. Cells were fixed overnight with ice-cold 70% ethanol and stained with Guava Cell Cycle Reagent (Millipore; 4500-0220) at room temperature for 30 min in the dark, then analyzed by use of the Guava EasyCyte Mini flow cytometry system (Millipore). A minimum of 5,000 events was counted.
Immunofluorescence assay
Treated hESCs were fixed with 4% paraformaldehyde for 15 min, blocked with 5% normal goat serum for 20 min at room temperature, and incubated with primary antibodies (1:100) for Oct4 (Abcam; ab18976), Nanog (Abcam; ab80892), or Sox2 (Abcam; ab97959); or LC3B (Cell Signaling Technology; 2775) at 4°C overnight, then corresponding Alexa 488-labeled secondary antibody (1:200) at 37°C for 1 h. Negative control (NC) cells were incubated with normal IgG. Cells were examined under a Leica TCS SP2 confocal laser scanning microscope (Leica).
Quantitative real-time PCR for miRNA
Total RNA was isolated by the Trizol reagent method (Invitrogen). RNA content was determined by use of the NanoDrop (Thermo). miR-4459 was converted into cDNA with use of the TaqMan MicroRNA Reverse Transcription Kit (Applied Biosystems; PN4366596) and a specific stem-loop primer, 5′-GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACTCCACCT-3′. The expression of mature miR-4459 in hESCs was quantified by use of the KAPA SYBR FAST qPCR Kit (Kapa Biosystems; KK4601). Quantitative real-time PCR involved the LightCycler 480 Real-Time PCR System (Roche). Small RNA U6 was used as an internal control for small RNAs. Primer sequences were for miR-4459, forward, 5′-ATTATCCAGGAGGCGGAGG-3′, and reverse, 5′-TCGTATCCAGTGCAGGGTC-3′; and U6, forward, 5′-CTCGCTTCGGCAGCACATATACT-3′, and reverse, 5′-ACGCTTCACGAATTTGCGTGTC-3′. miR-4459 expression was calculated relative to U6 expression, by three independent experiments.
RNA mimics transfection
HEK293 cells or hESCs were grown on six-well plates. The mimics of miR-4459 was designed to mimic endogenous mature miR-4459. RNA mimics for miR-4459 and NC were designed and purchased from Invitrogen and transfected into HEK293 or hESCs with use of Lipofectamine RNAiMAX Transfection Reagent (Invitrogen; 13778150) for 48 h. The final concentrations of the mimics and NC were 10 or 50 nM.
Luciferase reporter transfection and dual luciferase activity assay
HEK293 cells were seeded into 96-well plates at 6,000 cells per well for incubation overnight, then cotransfected with plasmids of dual-luciferase (containing firefly and Renilla luciferase) reporters for CDC20B 3′UTR (Luc-CDC20B 3′UTR) and miR-4459 mimics or NC (final concentration, 10 or 50 nM) with Lipofectamine 2000 (Invitrogen; 11668-019) for 24 h. Dual luciferase activity was measured by use of the Dual-Glo Luciferase Assay System (Promega; E2920). After reagent was added, firefly luciferase or Renilla luciferase activity was measured by use of a VICTOR X2 Multilabel Plate Reader (PerkinElmer). Firefly luciferase activity was normalized to that of Renilla.
Statistical analysis
The number of LC3B puncta was acquired by using ImageJ 1.47 (US National Institutes of Health). Data are presented as mean±SEM, and analysis involved use of SPSS 20.0.0 (SPSS, Inc.). Images were processed by use of Adobe Photoshop CS5 (Adobe). P<0.05 was considered statistically significant.
Results
3BDO could efficiently induce hESCs differentiation into three layers
The chemical small molecule, 3BDO, that inhibited autophagy in HUVECs and neuronal cells [12,13], but its function in hESCs was unknown. After 3BDO treatment, we observed the morphological changes of hESCs, whose loss of ESC morphologic features presented a differentiated state in maintenance medium for hESCs (Fig. 1A). We wondered whether 3BDO influence the maintenance of hESCs stemness. The mRNA levels of core transcript factors Oct4, Sox2, and Nanog were significantly decreased in hESCs with 3BDO treatment (Fig. 1B). Their protein levels were decreased with 30 or 60 μM 3BDO for 72 h (Figs. 1C and 2). Furthermore, we analyzed the cell cycle of hESCs with or without 3BDO treatment, and found that 3BDO-treated hESCs lost the ESC-specific cell cycle (short G1 phase) (Fig. 3A), which is crucial for maintenance of ESCs stemness [14]. Therefore, 3BDO could efficiently induce the differentiation of hESCs.
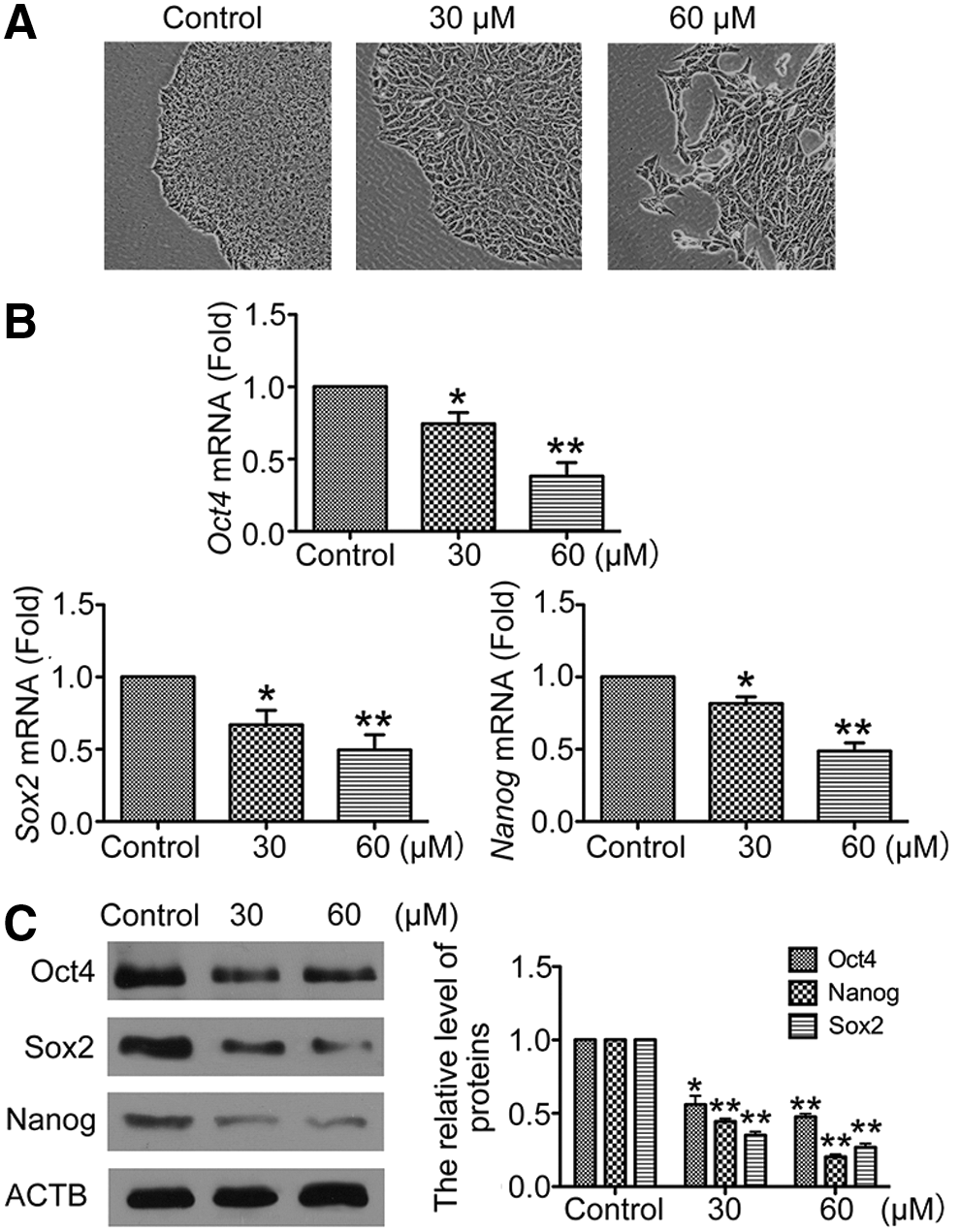
3-Benzyl-5-((2-nitrophenoxy) methyl)-dihydrofuran-2(3H)-one (3BDO) induced human embryonic stem cells (hESCs) differentiation.

3BDO downregulated core transcript factors Oct4, Sox2, and Nanog in hESCs. Immunofluorescent assay of Oct4, Nanog, or Sox2 protein level in hESCs with or without 3BDO (30 or 60 μM) for 72 h (scale bar=40 μm).

hESCs lost the ESC-specific cell cycle and differentiated into three layers after 3BDO treatment.
To understand which germ layer differentiation was induced by 3BDO in hESCs, we examined the expression of markers for three germ layers [Nestin, Zic1 and Sox1 for ectoderm; Nkx2.5, Brachyury (T gene) and Flk1 for mesoderm; Gata6, Gata2, Foxa2, and Sox17 for endoderm; and SSEA4 for self-renewal] in hESCs cultured with or without 3BDO. The mRNA levels of markers for the three germ layers were upregulated with 3BDO treatment (Fig. 3B). Collectively, our data showed that 3BDO could rapidly lead hESCs to lose the stemness and the ESC-specific cell cycle, and drive it to differentiate into three layers.
FLJ11812 RNA level was selectively decreased with 3BDO treatment in hESCs
In our previous work, we found that 3BDO selectively downregulated FLJ11812 in HUVECs [6]. To understand how 3BDO promote hESCs differentiation, we detected the RNA level of FLJ11812 in hESCs with or without 3BDO treatment. The result showed that the RNA level of FLJ11812 was significantly decreased by 3BDO at 30 or 60 μM for 72 h, but mRNA level of TGFB2 was not affected (Fig. 4A). On fluorescent in situ hybridization, 3BDO at 30 or 60 μM for 72 h significantly inhibited the expression of FLJ11812 (Fig. 4B). Therefore, 3BDO could specifically decrease FLJ11812 RNA level in hESCs.
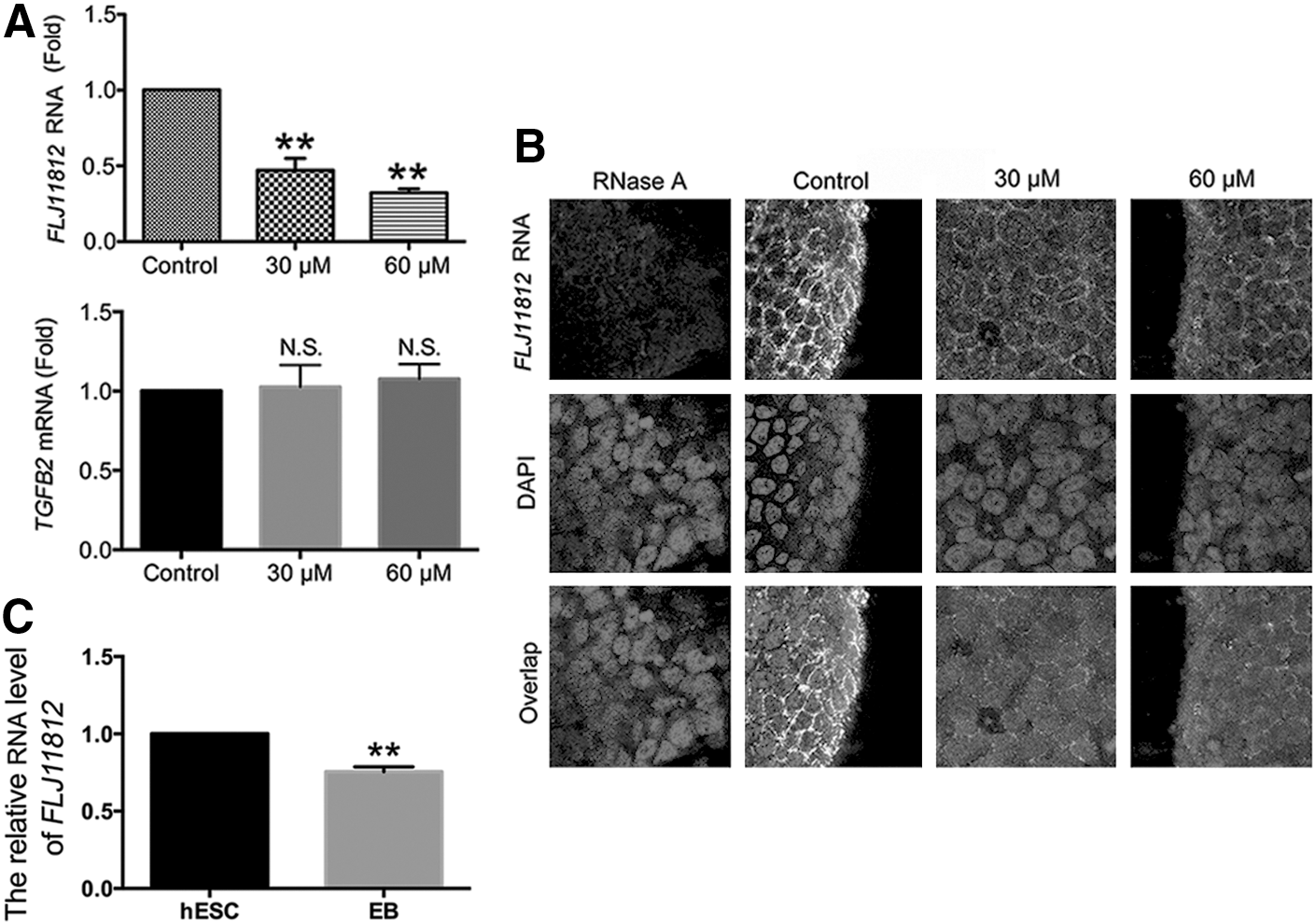
3BDO treatment selectively decreased FLJ11812 RNA level in hESCs.
To confirm the role of FLJ11812 in maintaining stemness of hESCs, we examined the changes in FLJ11812 RNA level during hESCs differentiation. FLJ11812 RNA level was reduced in embryoid bodies (EBs) from hESCs as compared with undifferentiated hESCs (Fig. 4C). The reduction of FLJ11812 RNA level in EBs indicated that FLJ11812 expression was positively related with the pluripotent state. Thus, according to these results, the 3BDO-induced differentiation of hESCs might be due to the selectively decreased FLJ11812 RNA level.
FLJ11812 functioned as an endogenous sponge of miR-4459 in hESCs, which directly bind with the 3′UTR of CDC20B
FLJ11812 acts as an endogenous “sponge” of miR-4459 and downregulates miR-4459 expression levels in HUVECs [6], so we evaluated the level of miR-4459 in hESCs after 3BDO inhibition of FLJ11812 RNA level. The mRNA level of miR-4459 was increased with 3BDO treatment (Fig. 5A). Therefore, FLJ11812 also functioned as an endogenous sponge of miR-4459 in hESCs.

miR-4459 was upregulated after 3BDO treatment and directly bound with the 3′ untranslated regions (UTR) of CDC20B.
We had identified ATG13 as a target of miR-4459 in our previous work [6]. An miRNA may have many different mRNA targets [15]. To find out other target genes of miR-4459, we analyzed the 3′UTR of each predicted target mRNA sequence except ATG13 by using RNAhybrid software, and found that the 3′UTR of CDC20B showed a highly conserved miR-4459-predicted binding sequence (Fig. 5B). The approximate 200 bp fragment of 3′UTR, including the best predicted binding sites of miR-4459 from CDC20B, was cloned into the pGL3 vector. Luciferase assay showed that miR-4459 mimics greatly reduced the luciferase activities of the reporter vectors containing the 3′UTR of CDC20B (Fig. 5C). Together, these results indicated that FLJ11812 was able to downregulate miR-4459 expression in hESCs; in addition, CDC20B was another downstream target of miR-4459.
miR-4459 regulated by FLJII812 participates in the maintenance of hESCs stemness through decreasing the protein level of CDC20B and ATG13
To investigate the regulatory role of FLJ11812 in the expression of CDC20B and ATG13, targets of miR-4459, after downregulation of FLJ11812 by 3BDO in hESCs, we measured the protein levels of CDC20B and ATG13 with or without 3BDO treatment. The result showed that the protein levels of CDC20B and ATG13 were decreased in hESCs treated with 3BDO (Fig. 6A). Due to the distinctiveness of hESCs, we further overexpressed FLJ11812 in HEK293T with or without 3BDO treatment and founded that FLJ11812 could eliminate the effect of 3BDO treatment (Supplementary Fig. S1). These data suggested that FLJ11812 could regulate the expression of CDC20B and ATG13 by binding with miR-4459, which targets their 3′UTR.
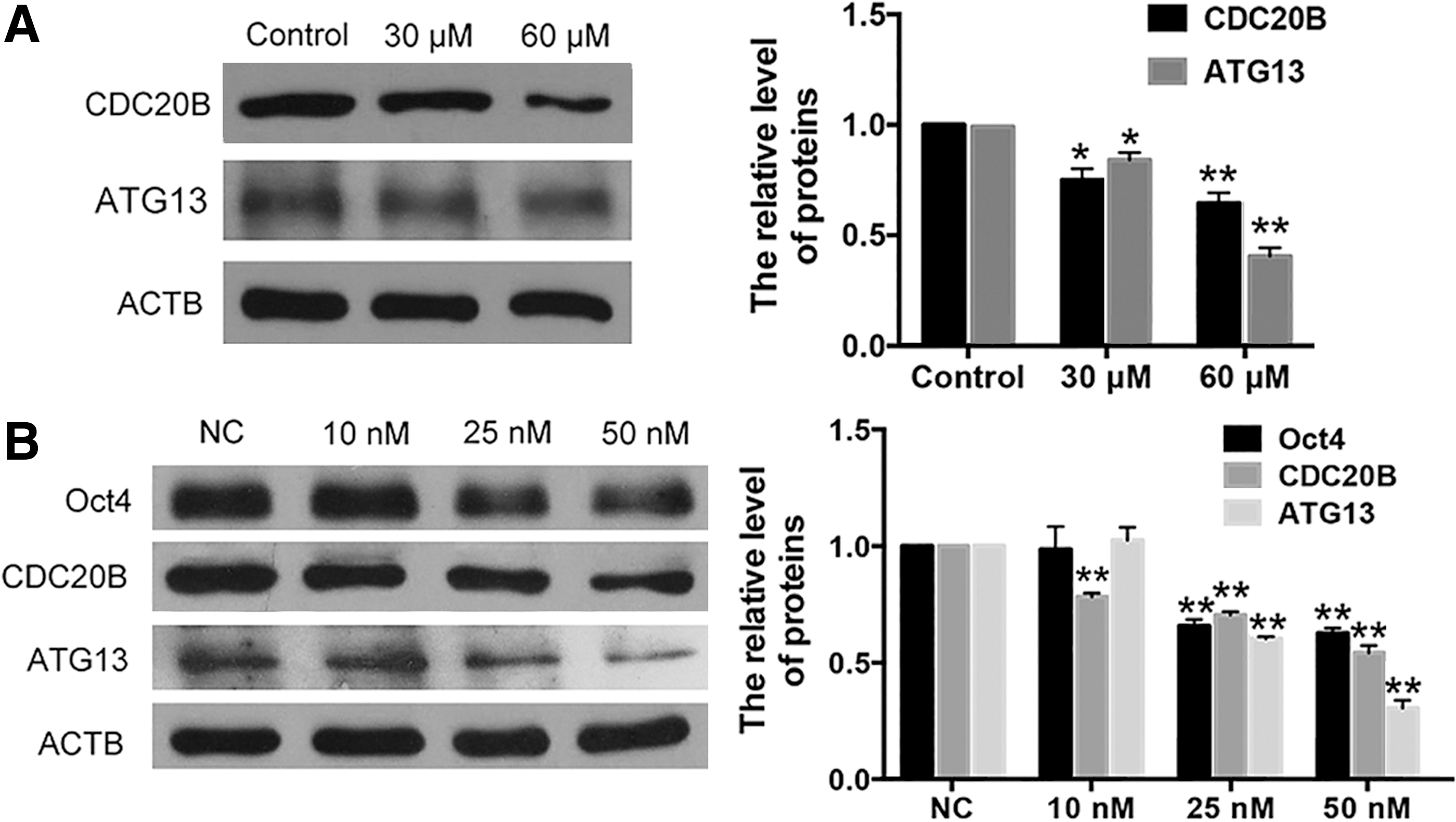
miR-4459 regulated the maintenance of hESCs stemness via its targets—CDC20B and ATG13.
To verify whether miR-4459 participates in the maintenance of stemness of hESCs, we detected the protein levels of its targets and Oct4 after transduction of miR-4459 mimics. Western blot showed that miR-4459 mimics significantly inhibited protein levels of CDC20B and ATG13 as well as Oct4 (Fig. 6B). Meanwhile, we predicted the potential binding site of miR-4459 in the whole sequence of Oct4 and did not find any binding site. Oct4 is critically involved in the self-renewal of undifferentiated ESCs and frequently used as a marker for undifferentiated cells [16]. Therefore, these results indicated that miR-4459 indeed directly regulate the expression of CDC20B and ATG13, and indirectly regulate pluripotency in hESCs. Taken together, miR-4459, which is downregulated by FLJ11812, plays an important role in the maintenance of hESCs stemness through decreasing the protein level of CDC20B and ATG13.
Meanwhile, we previously found that 3BDO could inhibit autophagy via a FLJ11812–miR-4459–ATG13 signal pathway in HUVECs. To validate this regulatory mechanism in hESCs, we detected autophagy by western blot and immunofluorescence in hESCs after treatment with 3BDO for 72 h. The protein level of LC3B-II and number of LC3B puncta were decreased with 3BDO treatment (Supplementary Fig. S2). Furthermore, we also found that 3BDO-treated hESCs lost the ESC-specific cell cycle. Therefore, 3BDO could inhibit cell cycle and autophagy in hESCs (Fig. 7).
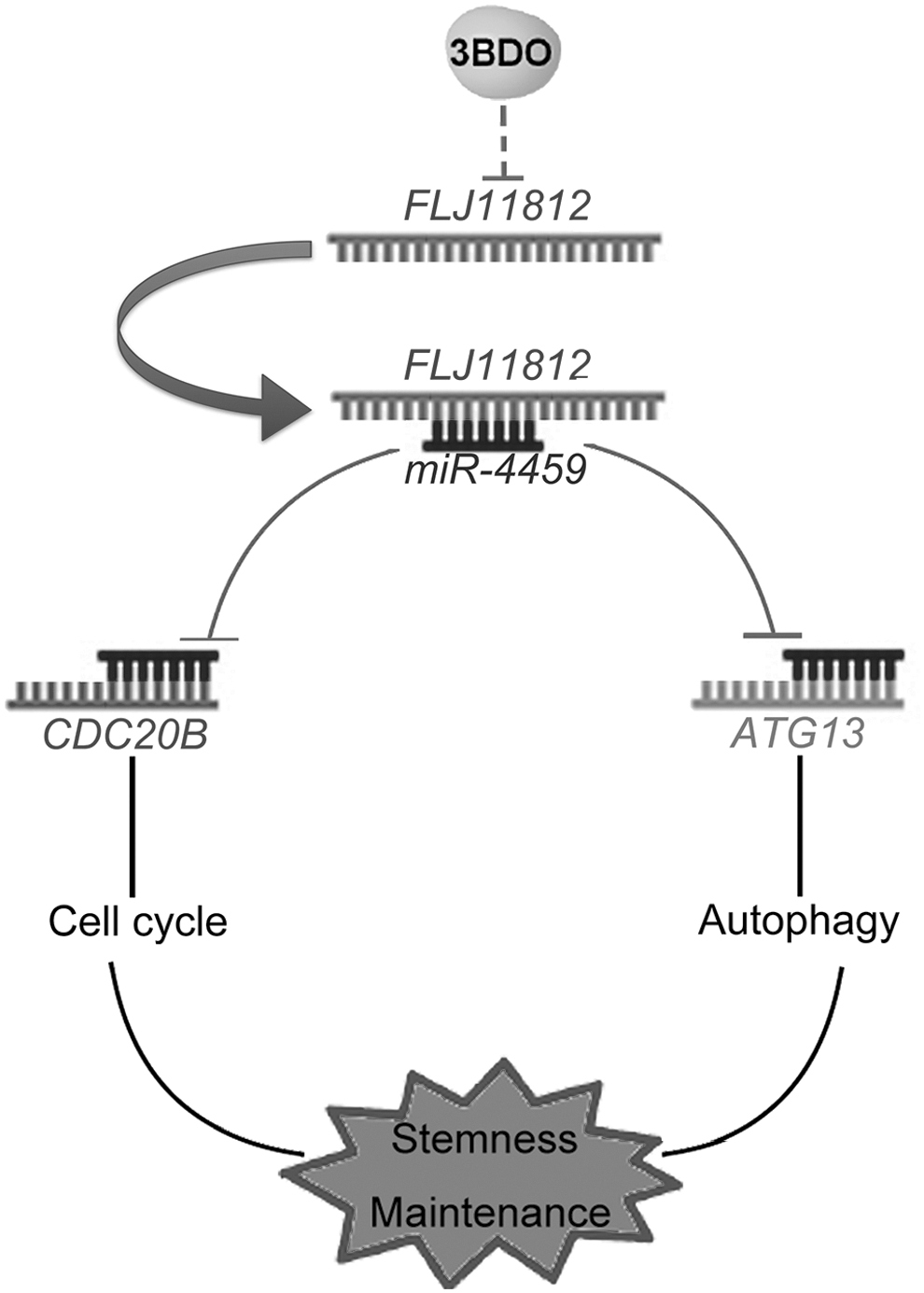
A conceptual schematic of FLJ11812-related regulatory mechanism in hESCs stemness. FLJ11812 functions as a competing endogenous RNA (ceRNA) to bind with miR-4459, which regulates the expression of its targets (CDC20B and ATG13), thus controlling hESCs stemness by regulating the cell cycle and autophagy.
Discussion
The uaRNAs have important roles in various biological processes, especially development. We recently identified a chemical molecule, 3BDO, that could efficiently induce hESCs differentiation into three layers, and selectively and significantly downregulate a uaRNA, FLJ11812. By acting as a ceRNA, FLJ11812 had positive roles in the pluripotent state and maintenance of hESCs stemness. FLJ11812 could bind with miR-4459 to regulate the expression of its targets, CDC20B and ATG13, and thus control stemness via different biological effects.
In our previous study, 3BDO was identified as a new activator of MTOR in HUVECs [6]. MTOR also played an intricate role in the maintenance of hESCs stemness. Inhibition of MTOR impairs pluripotency by rapamycin treatment, prevents cell proliferation, and enhances mesoderm and endoderm activities in hESCs [17]. However, MTOR-mediated activation of p70 S6K induces differentiation of hESCs [18]. Therefore, our findings provided new clues and a powerful tool for investigating the dual role of MTOR in the maintenance of hESCs stemness.
ncRNAs could function as ceRNAs in regulating the concentration and biological functions of miRNAs in stem cells [8,19]. In previous research, linc-MD1 was found to be a ceRNA that controls muscle differentiation by binding miR-133 and miR-135 and regulates mRNA targets of miRNAs, including MAML1 and MEF2C [19]. A large intergenic ncRNA, lincRNA-RoR, was identified as a ceRNA that regulates the expression of core transcript factors (Oct4, Sox2, and Nanog) and differentiation-related miRNAs, including miR-145-5p, miR-181a-5p, and miR-99b-3p, in hESCs [8]. linc-RoR shared these miRNAs with the core transcript factors and “directly” regulated self-renewal and pluripotency of hESCs through miRNAs targeting core transcript factors. This work showed that indirect regulation of a ceRNA, FLJ11812, regulated miR-4459 and its targets in the stemness of hESCs.
Here, we identified that the uaRNA FLJ11812 has ceRNA activity for miR-4459 and regulated its targets (CDC20B and ATG13), thereby affecting the stemness of hESCs. We speculated that the two targets of miR-4459 coregulated the hESCs stemness and differentiation: CDC20B and ATG13. CDC20B protein belongs to the WD repeat CDC20/Fizzy family and may participate in cell cycle progression. ATG13 is required for phagosome formation and autophagy induction [20]. Some reports have shown that autophagy plays a crucial role in maintaining self-renewal and pluripotency capacity and differentiation of ESCs [21,22]. In addition to CDC20B and ATG13, there may be some other targets of miR-4459 that are detectable through RNAhybrid software analysis, such as TPPP and LARP1, that was found to be required for spermatogenesis and embryogenesis [23,24].
Taken together, FLJ11812 regulated the stemness of hESCs through two different biological processes—cell cycle and autophagy—by controlling miR-4459 (Fig. 7). Our findings illuminate the different mechanisms of ceRNA in stemness regulation of hESCs.
A previous report demonstrated that FLJ11812 was expressed in the whole embryo, mainly in the head [7], and may be important for embryogenesis and neurogenesis. However, because of overlapping TGFB2 mRNA, FLJ11812 is difficult to study by common RNAi. In this work, by using a chemical small molecule, 3BDO, that could selectively downregulate FLJ11812, we showed that FLJ11812 played a key role in maintenance of stemness of hESCs for the first time. Therefore, our findings provide new clues and a powerful tool for investigating the action mechanism of FLJ11812 in early development.
Conclusion
We identified 3BDO as a new tool for hESCs differentiation via selectively downregulating a uaRNA level—FLJ11812. FLJ11812 functions as an endogenous sponge of miR-4459 in hESCs, further upregulated the protein level of its targets—CDC20B and ATG13, so as to maintain stemness of hESCs through two different biological processes—cell cycle and autophagy.
Footnotes
Acknowledgments
This work was financially supported by the National Natural Science Foundation of China (nos. 31270877, 81321061, 91313303, 20972088, and 31070735), the National 973 Research Project (no. 2011CB503906), and Specialized Research Fund for the Doctoral Program of Higher Education (no. 20120131130010).
Author Disclosure Statement
The authors indicate no potential conflicts of interest.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.