
Abstract
When mesenchymal stem cells (MSCs) are used for therapy of immunological pathologies, they get into an inflammatory environment, altering the effectiveness of the treatment. To establish the impact of environmental inflammatory factors on MSCs' immunofunction in the mirror of intrinsic heterogeneity of mouse MSC population, individual MSC clones were generated and characterized. Adipogenic but not osteogenic differentiation and pro-angiogenic activity of five independent MSC cell lines were similar. Regarding osteogenic differentiation, clones MSC3 and MSC6 exhibited poorer capacity than MSC2, MSC4, and MSC5. To study the immunosuppressive heterogeneity, in vitro and in vivo experiments have been carried out using T-cell proliferation assay and delayed-type hypersensitivity (DTH) response, respectively. A remarkable difference was found between the clones in their ability to inhibit T-cell proliferation in the following order: MSC2≥MSC5>MSC4>MSC3 >> MSC6. Nevertheless, the differences between the immunosuppressive activities of the individual clones disappeared on pretreatment of the cells with pro-inflammatory cytokines, a procedure called licensing. Stimulation of all clones with IFN-γ and TNF-α resulted in elevation of their inhibitory capability to a similar level. Nitric oxide (NO) and prostaglandin E2 (PGE2) were identified as major mediators of immunofunction of the MSC clones. The earlier findings were also supported by in vivo results. Without licensing, MSC2 inhibited DTH response, while MSC6 did not affect DTH response. In contrast, prestimulation of MSC6 with inflammatory cytokines resulted in strong suppression by this clone as well. Here, we have showed that MSC population is functionally heterogeneous in terms of immunosuppressive function; however, this variability is largely reduced under pro-inflammatory conditions.
Introduction
M
Therapy of severe refractory acute graft versus host disease (GVHD) [6] has already received conditional approval in several countries. Moreover, treatment of myocardial infarction is one of the most successful clinical improvements being at phase II clinical trial [7,8]. Benefits of MSC therapy originate from the fact that the MSCs promote tissue regeneration via differentiation and engraftment into the injured recipient tissue and/or production of a wide panel of bioactive molecules supporting angiogenesis, stimulating endogenous stem cell recruitment and differentiation, and modulating immune response by diminishing inflammation [9]. Although a growing body of data has been accumulated, suggesting clonal heterogeneity of MSCs, it is still not well established how the heterogeneity of MSC population affects the therapeutical application.
The heterogeneity of MSCs has been demonstrated regarding morphology, cell surface marker expression, differentiation potential, and transcriptome [10 –12]. It was first proposed by Friedenstein and his colleagues, who described variable osteogenic potential of bone marrow-derived fibroblastoid colonies in vivo [13]. This study was followed by more than a dozen studies showing that MSC clones displayed various capacities of multilineage differentiation both in vitro and in vivo (reviewed in Pevsner-Fischer et al. [10]). On the other hand, immunosuppressive functions of single cell-derived MSC populations have been improperly compared. The few studies available resulted in controversial data showing either similar or different immunomodulatory capacities of single cell-derived MSC lines [14 –16]. Recent findings have provided evidence that the immunosuppressive function of these cells is modulated by an inflammatory microenvironment [15,17]. Whether licensing by pro-inflammatory cytokines affects the immunosuppressive function at the clonal level has not yet been revealed.
This work aims at exploring the heterogeneity of mouse bone marrow-derived MSC population with a special focus on the consequence of the heterogeneity to the immune function. Differentiation properties and pro-angiogenic activities are characterized at the clonal level. Our results show that the analyzed five MSC clones are similar in immunophenotype, adipogenic differentiation, and pro-angiogenic activity. Two clones, MSC3 and MSC6 show poorer capacity to differentiate toward osteogenic direction. Most importantly, remarkable differences are observed in immunosuppressive capabilities between the individual clones both in vitro and in vivo. However, the variability in the immune function disappears when MSCs are licensed by inflammatory cytokines, indicating that the functions of individual cells are largely dependent on their surrounding environment.
Materials and Methods
Cell culture
MSCs were isolated from bone marrow of one C57BL/6 mouse and characterized as previously described [18,19]. The cells were cultured in DMEM (Gibco®) supplemented with 10% fetal bovine serum (FBS) (Gibco), penicillin-streptomycin (Sigma-Aldrich), and 2 mM
MSC culture at passage 10 was cloned with limiting dilution in DMEM/F-12 (Gibco) supplemented with 10% FBS and 5% horse serum (Gibco) by plating 1 cell/well in a 96-well culture plate (Orange Scientific). Five monoclonal MSC cultures labeled as MSC2-6 were randomly selected under a CKX41 inverted light microscope (Olympus Holding Europa GmbH) and further cultured till 5–10 passages.
Flow cytometry
Cells (5×105 cells/sample) were labeled for 30 min at 4°C with various antibodies: anti-mouse Sca-1 labeled with phycoerythrin (PE), CD44-PE, biotin-conjugated CD119 (BD Pharmingen) followed by streptavidin-PE (Sigma-Aldrich). Mouse Mesenchymal Stem Cell Marker Antibody Panel (BD Pharmingen) was used to analyze CD11b, CD45, CD29, and CD106 according to the manufacturer's instructions. The proper isotype-matched immunoglobulins were used as controls. Cell analysis was performed by a FACSCalibur flow cytometer (Becton Dickinson). Evaluation of the data was carried out with CellQuest™ software (Becton Dickinson).
Detection of adipogenic and osteogenic differentiation
Adipogenic medium consisted of DMEM/F-12 supplemented with 10% FBS, penicillin-streptomycin, 0.5 mM 3-isobutyl-1-methylxanthine (IBMX; Sigma-Aldrich), 10 μM dexamethasone (Sigma-Aldrich), 66 μM indomethacin (Sigma-Aldrich), and 2 μL/mL insulin (insulin lispro injection, 100 U/mL, Humalog®) [20]. MSCs were maintained in adipogenic medium for 3 days, detached with 0.05% trypsin-EDTA, and fixed with 4% paraformaldehyde (Sigma-Aldrich) and lipid droplets were stained with fluorescent AdipoRed™ according to the manufacturer's instructions (Lonza). Fluorescent lipid droplets were detected with the Axioskop 2 Mot fluorescence microscope (Carl Zeiss) using EC Plan-Neofluar 20×/0.5 objective (Carl Zeiss). For quantitation of lipid accumulation in adipocytes, the cells (5×105 cells/sample) were labeled with AdipoRed as described earlier and quantified by the FACSCalibur flow cytometer using CellQuest software.
Osteogenic medium was composed of CM and 10 mM β-glycerophosphate (Sigma-Aldrich), 50 μg/mL L-ascorbic acid 2-phosphate (Sigma-Aldrich), and 0.1 μM hydrocortisone (Sigma-Aldrich). MSCs were cultured in osteogenic medium for 14 days, fixed with 8% formaldehyde, and stained with Alizarin Red S (Sigma-Aldrich) solution (pH 4.1) to detect calcium deposition. Photomicrographs were taken with an Olympus CKX41 inverted light microscope and an Olympus Camedia C-5060 camera (Olympus Holding Europa GmbH).
In vitro prevascular structure assay
Co-culture of MSCs and mouse heart endothelioma cells, H5V (kindly provided by Vizler C of HAS, BRC, Hungary) [21] was initiated at a ratio of 1: 1 in a 24-well plate at a cell density of 2×104 cells/well. Prevascular structure formation was evaluated as follows: Five randomly selected nonoverlapping areas of co-plated cells were examined by an Olympus Cell-R fluorescence microscope (Olympus Holding Europa GmbH) using UPlanSApo 4×/0.16 objective. The lengths of prevascular structures were quantified by measuring with CellR Imaging Software (Olympus Holding Europa GmbH) after 3 days of cell culture.
T-cell proliferation assay
First, MSCs were seeded on a 96-well plate at a cell density of 5×103 cells/100 μL CM in a well. Mesenteric, inguinal, and brachial lymph nodes from Balb/C mice were dissected. Lymph nodes were disrupted mechanically to release most of the leukocytes. Leukocytes were harvested by centrifugation and resuspended in phosphate-buffered saline (PBS) containing 5% FBS at a final concentration of 2×106 cells/mL. Carboxyfluorescein diacetate succinimidyl ester (CFSE, CellTrace™ CFSE Cell Proliferation Kit; Life Technologies) was added to leukocytes at a final concentration of 2.5 μM, and cells were incubated in the dark for 5 min at room temperature. Leukocytes were washed twice with fresh RPMI culture medium (Gibco); then, 2×105 cells were added to the preseeded MSCs in 100 μL of RPMI containing 10% FBS, 50 μM β-mercaptoethanol (Sigma-Aldrich), and 5 μg/mL Concanavalin A (Con A; Sigma-Aldrich). In blocking experiments, nitric oxide synthase (NOS2), prostaglandin-endoperoxide synthase 2 (cyclooxygenase 2) (PTGS2) or indolamine-2,3-dioxygenase (IDO) inhibitors, 1 mM NG-methyl-L-arginine acetate salt, L-NMMA (Sigma-Aldrich), 10 μM indomethacin (Sigma-Aldrich), or 1 mM 1-methyl-L-tryptophan, 1-MT (Sigma-Aldrich), respectively, were added to MSC/leukocyte co-cultures. After 72 h of incubation, nonadherent leukocytes were collected, washed with PBS, settled by centrifugation, and resuspended in PBS supplemented with 1% FBS, 0.1% sodium-azide (FACS-buffer), and 10 μg/mL propidium-iodide. Cell proliferation was measured by flow cytometry. Evaluation of the data was carried out using CellQuest software (Becton Dickinson) and ModFit LT™ Version 2.0. software (Verity Software House, Inc.). T-cell proliferation was evaluated as proliferating fraction (PF) calculated as follows: PF%=number of proliferating T-cells/total number of T-cells ×100.
In some experiments, MSCs were pretreated with a mixture of 100 ng/mL mouse recombinant IFN-γ (R&D Systems) and 50 ng/mL TNF-α (kindly provided by Duda E of HAS, BRC Hungary) [22] for 24 h, then washed twice with CM prior T-cells were added, and finally T-cell proliferation was tested as described earlier.
Western blot analysis
Mesenchymal stromal cells were lysed (107 MSCs/mL) in RIPA buffer containing 10 mM Tris•HCl and 150 mM NaCl (Molar Chemicals), 1% Triton X-100 (Sigma-Aldrich), 0.5% sodium deoxycholate (Reanal), 0.1% SDS (Sigma-Aldrich), 2 mM EDTA (Reanal), and 1 mM PMSF (Sigma-Aldrich). The lysates were analyzed by western blotting after running the samples on a 10% SDS-polyacrylamide gel and then electroblotted onto a nitrocellulose membrane (Whatman® Protran®). After blocking with 3% cold fish gelatin (Sigma-Aldrich) in Tris-buffered saline (TBS) and 0.05% Tween® 20 (Sigma-Aldrich), the membranes were incubated with rabbit anti-Gal-1 (produced in our laboratory, [19]) and then with HRP-conjugated anti-rabbit immunoglobulin (Dako). Rabbit anti-β-actin antibody (Abcam) was used as a loading control. Immunoreactive proteins were visualized with Amersham™ ECL Prime Western Blotting Detection Reagent (GE Healthcare Life Sciences) and analyzed using the EC3 Imaging System furnished with a CCD Chemi 410 Camera (Ultra-Violet Products). Page Ruler™ Prestained Protein Ladder (Fermentas) was used as a molecular mass standard. Density of the bands was measured by VisionWorks®LS Image Acquisition and Analysis Software (Ultra-Violet Products).
Quantitative real-time PCR
MSCs (105 cells/well) were plated in 35 mm-diameter culture dishes in CM. After 24 h, fresh CM was added with or without 100 ng/mL IFN-γ and 50 ng/mL TNF-α and then further incubated for 24 h. Finally, total RNA was extracted using Nucleospin RNA II isolation kit (MACHEREY-NAGEL GmbH) according to the manufacturer's instructions. RevertAid H Minus First-Strand cDNA Synthesis Kit (Thermo Fisher Scientific, Inc.) was used to obtain cDNA from 2 μg of total RNA/reaction. Quantitative real-time PCR (qPCR) was performed using TaqMan® Gene Expression Master Mix (Life Technologies) for Ptgs2, Nos2, and Ido1 and AccuPower® 2X Greenstar qPCR Master Mix (Bioneer) for angiopoietin-1 (Angpt1), vascular endothelial growth factor A (Vegfa), transforming growth factor beta 1
Induction of delayed-type hypersensitivity response
Ovalbumin (1 mg/mL in PBS; Sigma-Aldrich) was emulsified at a 1: 1 ratio in complete Freund's adjuvant (Sigma-Aldrich). Male C57BL/6 mice (6–8 weeks old) were immunized by a subcutan injection of 100 μg ovalbumin/animal. A subcutaneous injection of PBS with complete Freund's adjuvant served as a control. Cells (106 MSCs/animal) or PBS were injected intraperitoneally in 200 μL volume. After 4 days, delayed-type hypersensitivity (DTH) was initiated by challenging with 250 μg heat-aggregated ovalbumin in 12.5 μL PBS or PBS alone injected into the right or left hind footpad, respectively. After 48 h, ovalbumin-induced footpad thickness was measured using a caliper and calculated as follows: (T48h−U48h)−(T0h−U0h), where T and U are thickness of ovalbumin-treated and ovalbumin-untreated footpads, respectively. All animal studies were approved by the Animal Care and Use Committee of the National Medical Center (Budapest, Hungary) in accordance with the national and international law and regulations of animal experiments.
Statistical analysis
Statistical analysis was performed using one-way analysis of variance test (ANOVA) in GraphPad Prism Version 6.03 (GraphPad Software). When significant (P<0.05) by ANOVA, pairwise comparisons of experimental groups were carried out using Fisher's least-significant difference (LSD) method as a post hoc test. For statistical analysis of qPCR data, the nonparametric Kruskal–Wallis test followed by Mann–Whitney test was used. Differences were considered statistically significant with a P-value less than 0.05 (set at *P<0.05, **P<0.01, ***P<0.001).
Results
Characterization of monoclonal MSC cell lines
Each clone uniformly expressed Sca-1, CD29, CD44, CD106, and CD119 (Fig. 1) at a similar degree and none of the clones expressed lineage-specific markers CD11b and CD45 (Fig. 1).
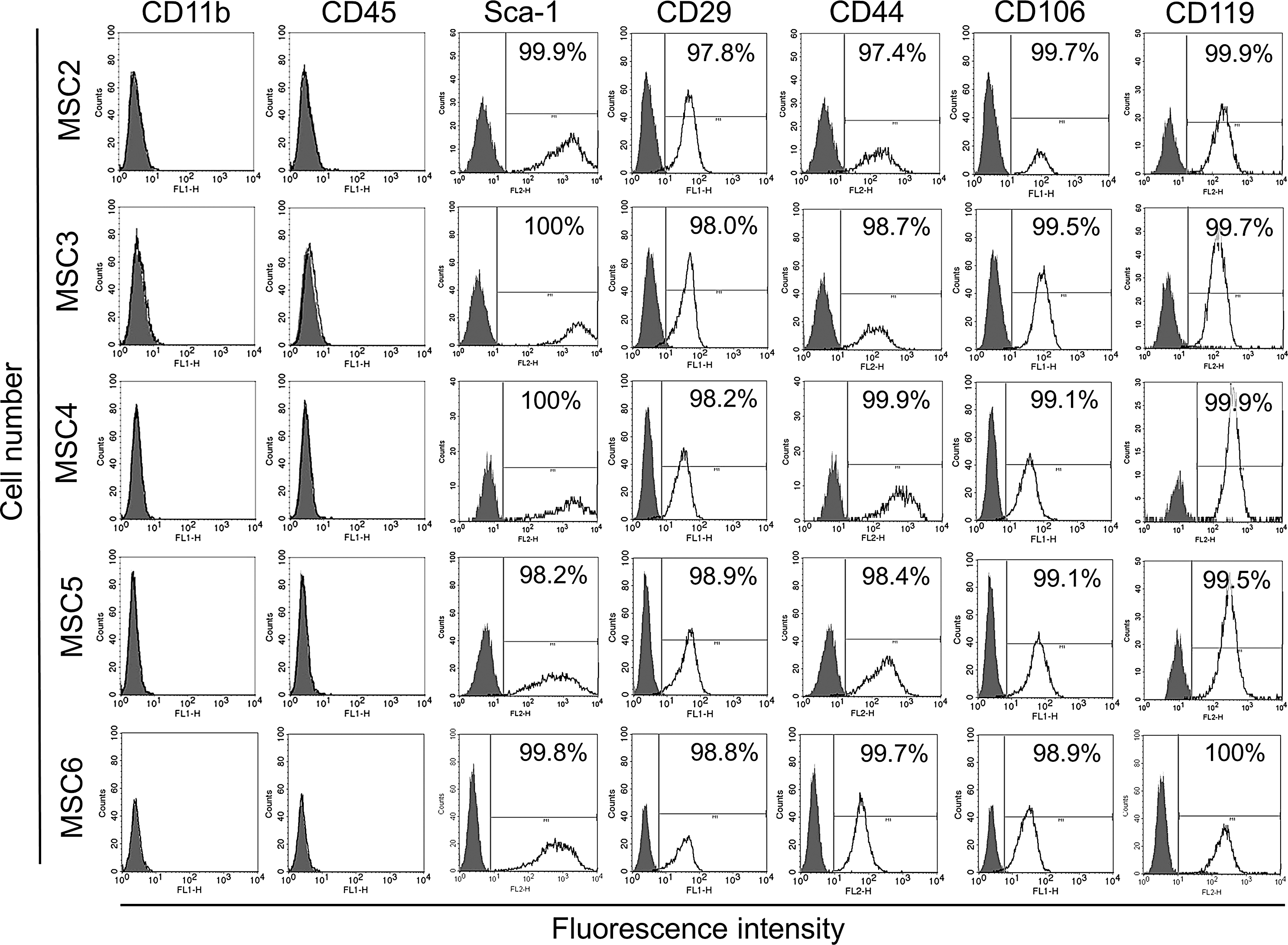
Expression of cell surface markers of monoclonal MSC lines. MSC2-6 were labeled with antibodies to CD11b, CD45, Sca-1, CD29, CD44, CD106, or CD119 (black lines). The negative controls were isotype matching immunoglobulins (gray shadowed area). Samples were then analyzed with flow cytometry.
In vitro osteoblast differentiation was assayed with Alisarin Red S staining showing that MSC2, MSC4, and MSC5 exhibited strong, while MSC3 and MSC6 underwent weak in vitro osteoblast differentiation (Fig. 2A). During adipocyte differentiation, all MSC clones accumulated a comparable amount of neutral lipids into intracellular lipid droplets (Fig. 2B, C), indicating that these cells had similar adipogenic differentiation potential.
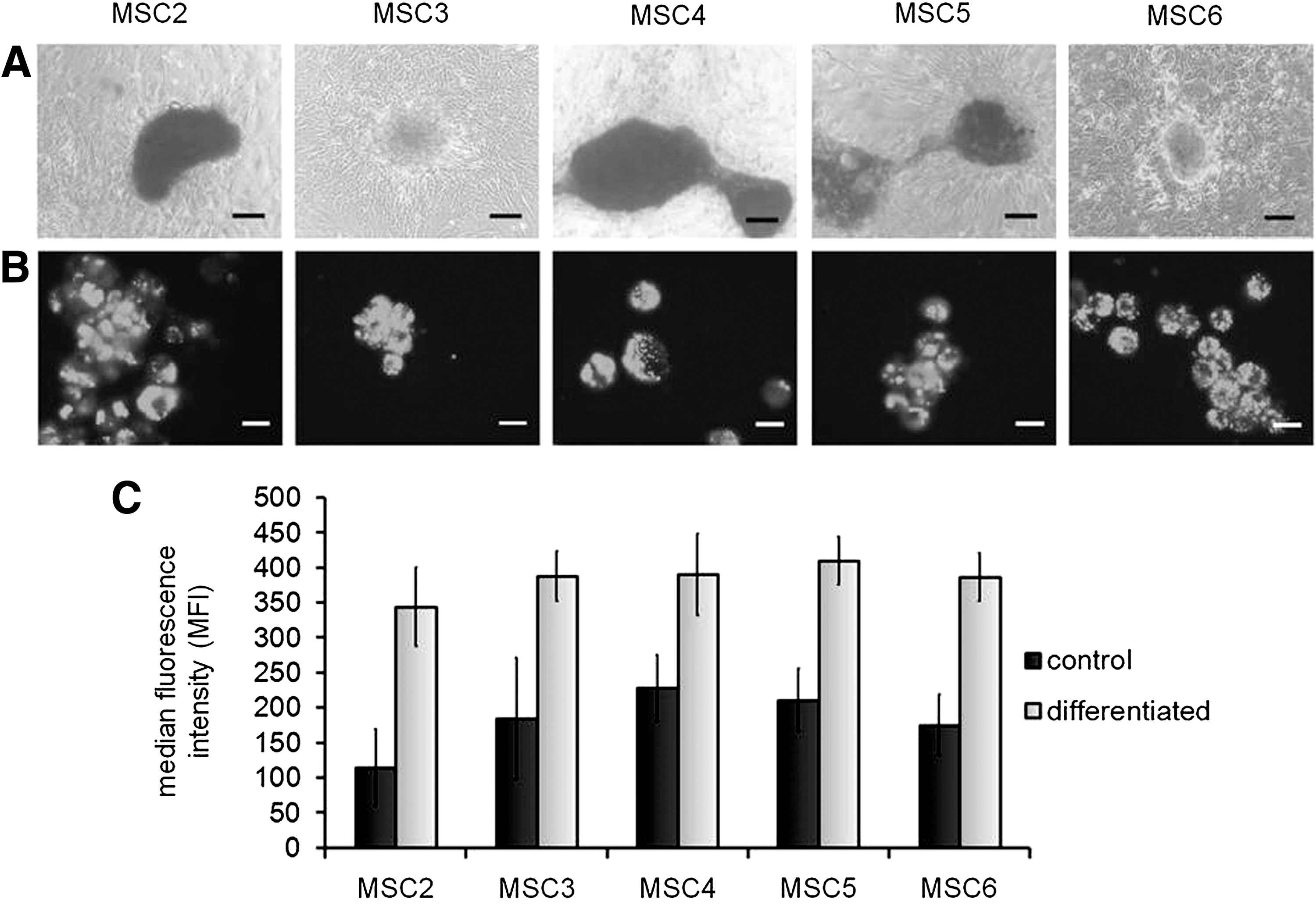
In vitro adipogenic and osteogenic differentiation of MSC clones. Cells were induced to differentiate toward osteoblastic
In vitro pro-angiogenic activity of MSC clones
We have previously reported that MSCs and H5V endothelial cells spontaneously arranged into vessel-like structures within 3 days of co-culture [19]. Moreover, deficiency in galectin-1, a pro-angiogenic lectin in MSCs, resulted in reduced formation of prevascular structures [19]. To determine whether individual MSC clones supported similarly or differently the shaping of these structures, in vitro pro-angiogenic assays were performed with the monoclonal MSC cultures. While neither MSC clones nor H5V endothelial cells formed prevascular structures alone, co-culturing any of the clones with H5V resulted in arrangement into prevascular structures (Fig. 3A) to a similar extent (Fig. 3B).
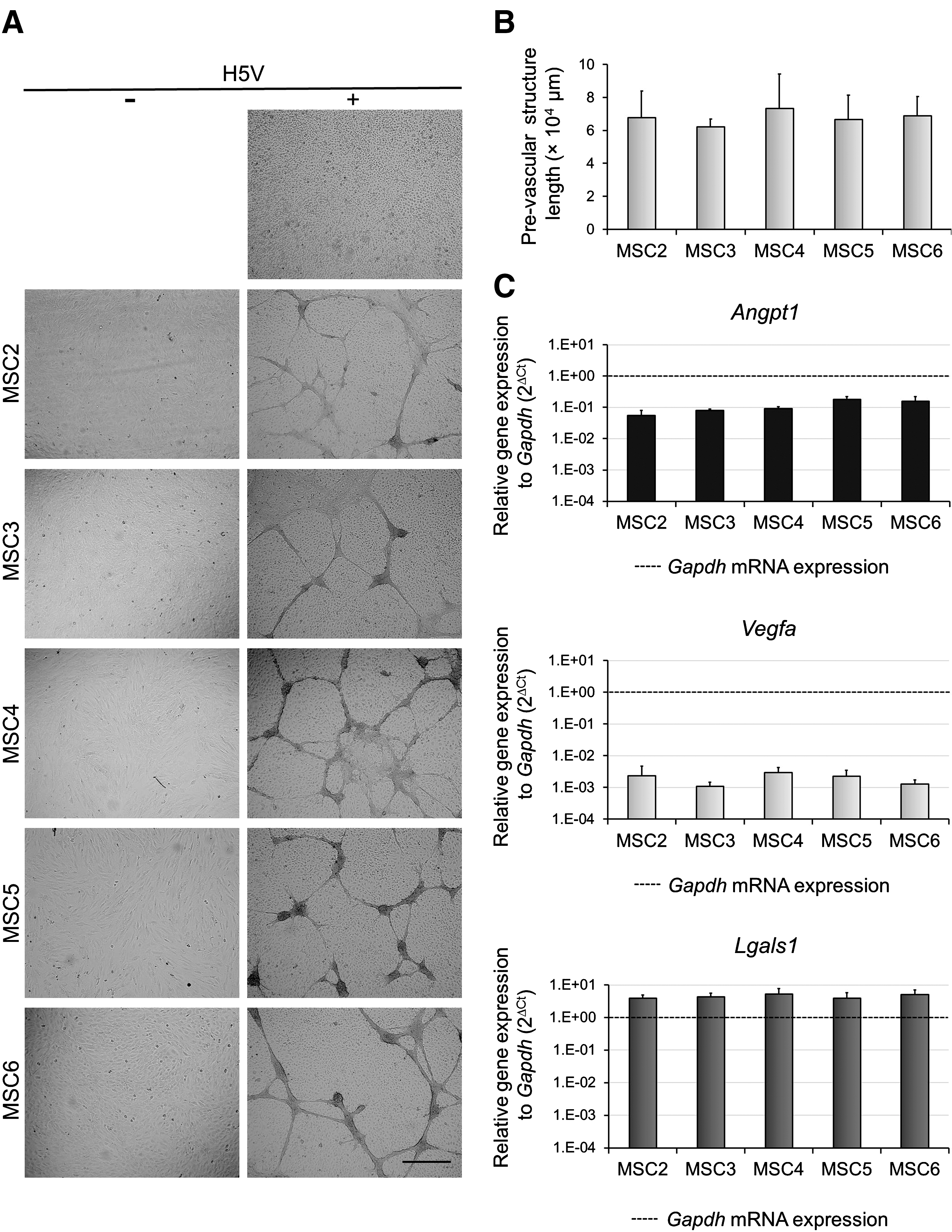
Monoclonal MSC lines exhibit similar pro-angiogenic activity. MSCs were co-cultured with or without H5V at a 1: 1 ratio for 3 days. Prevascular structures were visualized with an Olympus Cell-R fluorescence microscope using UPlanSApo 4×/0.16 objective (representative images selected from three independent experiments)
Analysis of pro-angiogenic factor expression by qPCR showed that all clones expressed a similar level of Angpt1, Vegfa, and Lgals1 mRNAs (Fig. 3C). Results of qPCR experiments (Fig. 3C) were validated by western blot analysis of galectin-1 protein (Supplementary Fig. S1; Supplementary Data are available online at
MSC clones vary in in vitro immunosuppressive activity
Immunomodulatory properties of MSC clones were assessed in vitro by analyzing their effect on Con A-activated T-cell proliferation. Proliferation of T-cells was robustfully diminished in the presence of MSC2, MSC4, and MSC5 while it was poorly inhibited by MSC3 and MSC6 (Fig. 4A, B). Differences were significant, and the order of their inhibitory capacity was as follows: MSC2≥MSC5>MSC4>MSC3 >> MSC6 (Fig. 4B).
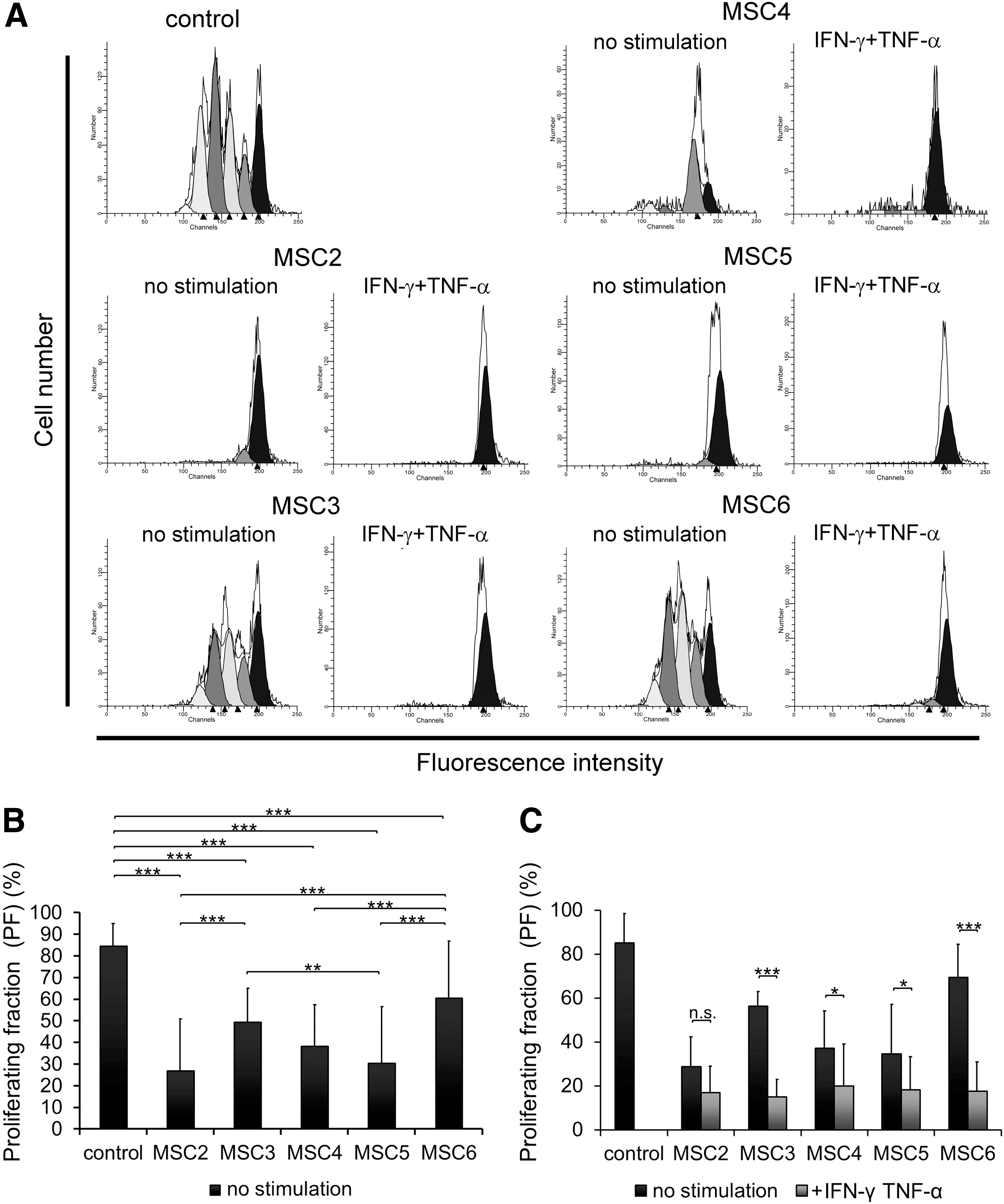
MSC cell lines vary in immunosuppressive activity. Cytofluorimetric profiles of CFSE-labeled T-cells under different conditions are presented (representative experiment)
Our aim was to determine whether variability between the clones regarding their inhibitory activity was reflected in the expression levels of immunomodulatory factors. Therefore, the expression levels of mRNAs encoding NOS2, PTGS2 responsible for the production of anti-inflammatory mediators, NO and PGE2, respectively, and IDO1 creating a tryptophan-deprived environment were analyzed. Relative Nos2 mRNA level well coincided with anti-proliferative function of the clones, as it was expressed at higher levels in MSC2, MSC5 (significant), and MSC4 (not significant) than in MSC3 and was undetectable in MSC6 (Fig. 5A). Expression of Ptgs2 did not show any remarkable difference between the clones (Fig. 5B). The two enzymes, NOS2 and PTGS2 seemed to be important in the inhibitory function of the most immunosuppressive clone MSC2. The inhibition of T-cell proliferation by MSC2 was blocked entirely with L-NMMA and partially with indomethacin, inhibitors of NOS2 and PTGS2, respectively (Fig. 6, Unstimulated MSC). In contrast, the presence of these inhibitors did not affect the activity of the least immunosuppressive clone MSC6 (Fig. 6, Unstimulated MSC). Expression of other immunomodulatory factors such as Tgfb1 (Supplementary Fig. S2) and Lgals1 (Fig. 3C) was similar, and Ido1 mRNA expression could not be detected in any of the clones (Supplementary Fig. S3A).
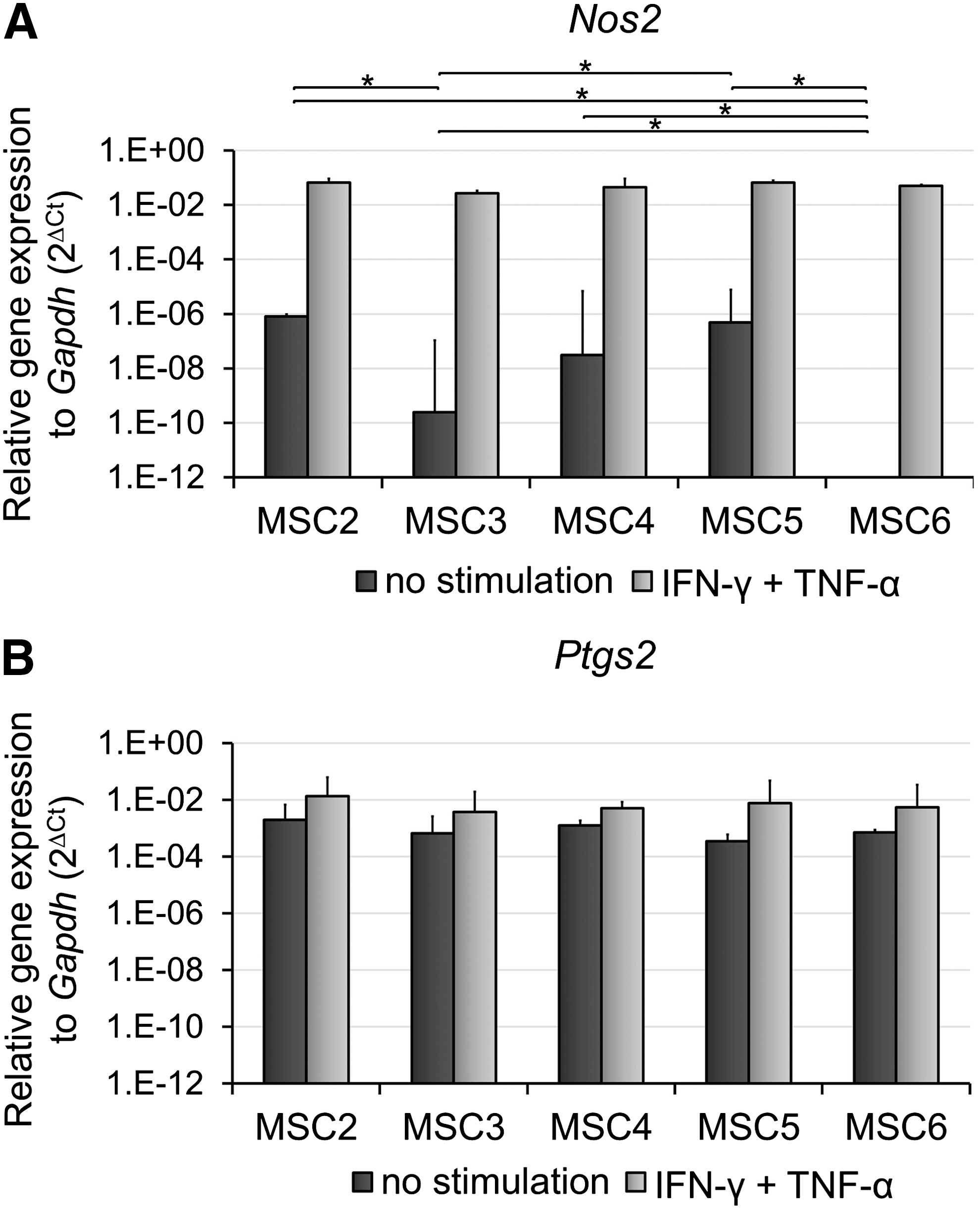
Expression levels of genes affecting immunosuppressive function of MSCs. Total RNA was extracted from cells cultured for 24 h in the absence or presence of 100 ng/mL IFN-γ and 50 ng/mL TNF-α and expression levels of Nos2
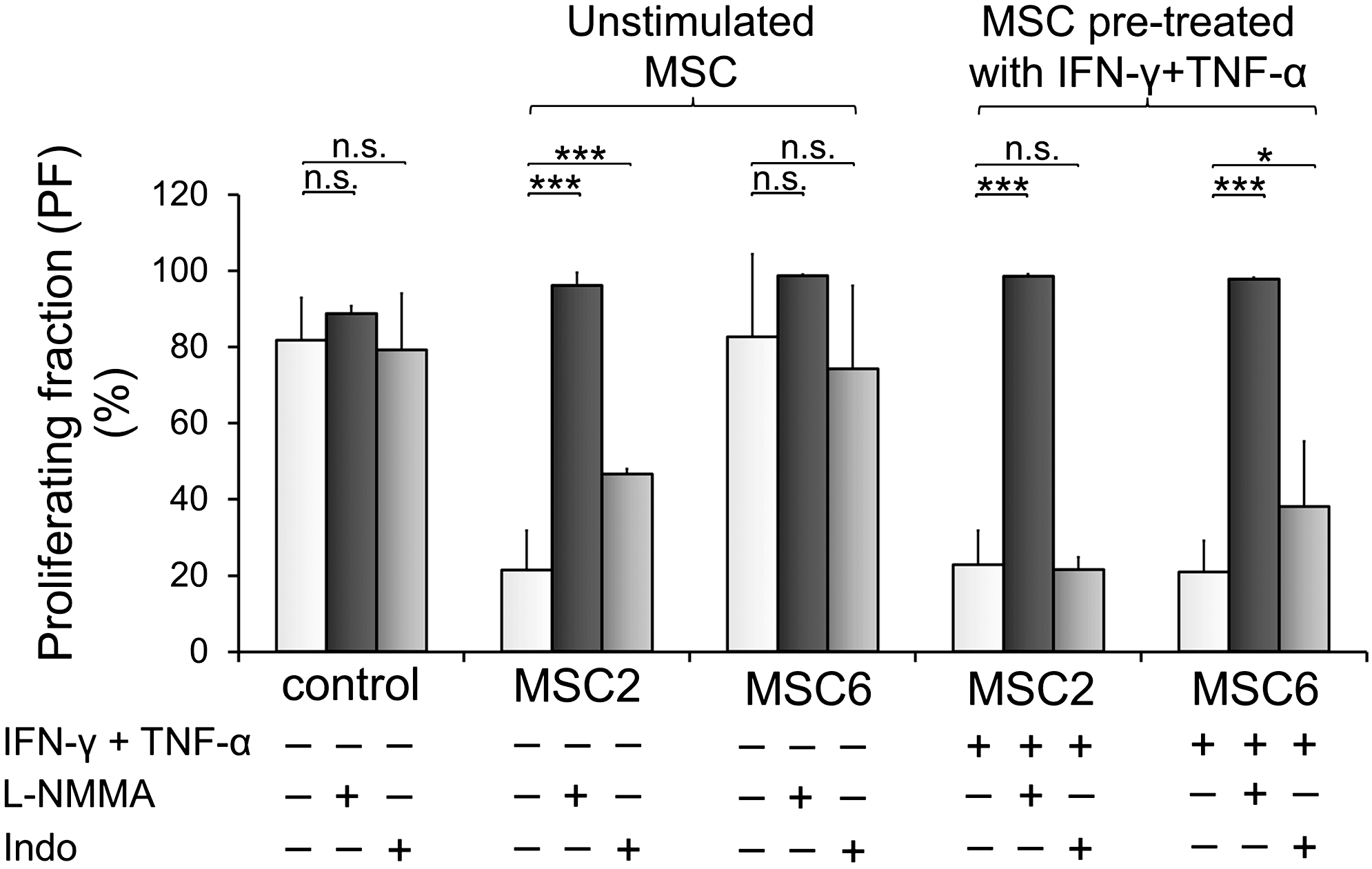
The role of NO and PGE2 in the inhibition of T-cell proliferation by MSC2 and MSC6. MSCs were plated on 96-well plates, left unstimulated, or preactivated with 100 ng/mL IFN-γ and 50 ng/mL TNF-α for 24 h. After 24 h, MSCs were washed with DMEM, co-cultured with freshly isolated, CFSE-labeled lymph node cells at a 1 to 40 ratio with or without 1 mM L-NMMA or 10 μM indomethacin (Indo). Proliferation of T-cells was stimulated by Con A. After 72 h of co-culture of MSCs and T-cells, proliferation of T-cells was assessed by cytofluorometry and data were analyzed by ModFit LT software. The figure shows the percentage of proliferating T-cells. Con A-activated T-cells cultured with or without L-NMMA or indomethacin in the absence of MSCs served as positive controls. The data are the representative of three independent experiments+SD. The experiments were performed in triplicate. n.s.=not significant P>0.05; *P<0.05; ***P<0.001.
To model the in vivo inflammatory environment in which MSCs act as anti-inflammatory cells, the monoclonal cell lines were pretreated with Th1-type cytokines, IFN-γ, and TNF-α and the effect of the licensed MSC on T-cell proliferation was analyzed. Licensing by pro-inflammatory cytokines, MSC3 and MSC6 clones, which exerted poor inhibitory activity, blocked the T-cell proliferation at a similar extent to the clones with strong inhibition. In contrast, licensing was either not or barely able to further enhance immunosuppressive activity of MSC2, MSC4, and MSC5 (Fig. 4C). Expression of Nos2 gene largely increased in all clones to an equal level on IFN-γ and TNF-α treatment. Nevertheless, the degree of elevation was the most remarkable in clones with poor inhibitory activity (Fig. 5A). On the other hand, Ptgs2 expression was also upregulated in all clones, although much less than that of Nos2 gene (Fig. 5B). Furthermore, unstimulated MSC clones did not express Ido1, unless they were preactivated with IFN-γ and TNF-α. Pro-inflammatory cytokine treatment triggered Ido1 mRNA expression as well (Supplementary Fig. S3A).
To determine the potential role of anti-inflammatory mediators in the inhibition of T-cell proliferation under inflammatory conditions, anti-proliferative activities of the clones were blocked with inhibitors. Immunosuppressive activity of licensed MSC2 and MSC6 were assayed in the presence of L-NMMA and confirmed the crucial role of NO. On the other hand, the presence of indomethacin slightly affected MSC6 but not MSC2 activity (Fig. 6, MSC pretreated with IFN-γ + TNF-α), suggesting a less pronounced function of PGE2. In addition, neither MSC2 nor MSC6 activities were influenced by IDO1 inhibitor 1-MT, independently from IFN-γ and TNF-α-activation (Supplementary Fig. S3B), indicating that IDO1 did not participate in the immunosuppression by these clones.
Ovalbumin-induced DTH model supports in vitro findings
The effect of the most and least immunosuppressive MSC2 and MSC6 clones, respectively, was tested in ovalbumin-induced DTH response in mice. MSC2 significantly prevented the thickening of footpad, while MSC6 injection resulted in no change compared with the control (treatment without MSCs) (Fig. 7). Nevertheless, when pretreated with IFN-γ and TNF-α before injection, MSC6 diminished DTH reaction similarly to nontreated MSC2 clone while immunosuppressive activity of MSC2 was not affected by licensing (data not shown).

Effect of an intraperitoneal injection of MSC2 and MSC6 in the ovalbumin-induced delayed-type hypersensitivity (DTH) model. Hypersensitivity was triggered in male C57BL/6 mice with subcutaneous administration of ovalbumin. Simultaneously, mice were injected intraperitoneally with 106 MSC2 or 106 MSC6 untreated or pretreated with IFN-γ and TNF-α in PBS or with PBS alone. After 4 days, heat-aggregated ovalbumin was injected into the right hind footpad and PBS was injected into the left hind footpad. After 48 h, ovalbumin-induced footpad thickness increment was measured. RFI, relative footpad increment, was calculated as follows: RFI=(treated48h−untreated48h)−(treated0h−untreated0h). The data are the means of two independent experiments. One-way ANOVA, Fisher's LSD multiple-comparison test was used for statistical analysis, n.s. P>0.05, *P<0.05; **P<0.01. Black lines mark the average footpad thickness increment in each experimental group.
Discussion
It has been proposed that the heterogeneity of MSCs may be partially responsible for the divergent outcomes of MSC-based clinical trials [11]. Several studies reported that various MSC clones differed in differentiation potential into mesenchymal lineages [10]. Other functional properties, such as pro-angiogenic effect, has not yet been extensively investigated at the clonal level. In addition, the data regarding heterogeneity in immunosuppressive properties of MSC population are insufficient and meet discrepancy. While Xu et al. argued that single cell-derived MSC populations possessed uniform immunomodulating capacities [14], others demonstrated large differences in immunosuppression at the clonal level [15,16].
In this study, clonal diversity of murine bone marrow MSCs was examined with a special focus on both the in vitro and in vivo immunosuppression. Moreover, in vitro osteoblast and adipocyte differentiation and pro-angiogenic effect of the MSC clones were also compared as a part of the characterization of the clones.
In according to previous results regarding variable osteogenic differentiation of single cell-derived human or murine MSCs [15,23 –25], we found that the clones exhibited different osteoblast potential in vitro. On the other hand, the analyzed clones similarly differentiated toward adipocytes. It has recently been described that MSCs produce a wide array of factors influencing endothelial cell growth and angiogenesis [26], thereby inducing capillary-like structure formation in vitro [27,28]. We previously demonstrated that MSCs spontaneously formed interconnected prevascular structures in a galectin-1 dependent fashion when co-cultured with H5V endothelial cells [19]. However, only one paper analyzed angiogenic activity of MSCs at the clonal level [29]. Burns et al. reported that two telomerase-immortalized human MSC (hMSC-TERT20) clones distinctly formed cord-like structures due to their different extracellular matrix protein production and the key role of galectin-1 had been proved in both in vitro and in vivo angiogenesis in serum-deprived condition in spite of equivalent expression of two major pro-angiogenic factors (VEGFA, ANGPT1) [29]. We analyzed the in vitro pro-angiogenic activity of MSC2-6 clones in a prevascular structure assay in which MSCs were co-cultured with H5V cells. The clones expressed a comparable level of pro-angiogenic factors, including Vegfa, Angpt1, and Lgals1 and accordingly, formed similar prevascular structures. The controversy between our and Burns' results may derive from the different experimental conditions, mainly from the fact that human MSCs were genetically modified and cultured under serum-depriving condition.
The major aspect of this work has been the comparison of the immunomodulatory activity of MSCs at the clonal level. The isolated MSC clones differently influenced the proliferation of Con A-activated T-cells ranging from 30% to 70% of inhibition. In this term, MSC2, MSC4, and MSC5 strongly inhibited T-cell proliferation while inhibitory activities of MSC3 and MSC6 were significantly weaker. Our results were in accordance with those published by others [15,16], reporting a difference between the immunosuppressive activities of single cell-derived MSC clones. The diversity in the inhibition of T-cell proliferation could be explained by the different expression of immunomodulatory gene Nos2, responsible for the generation of a crucial mediator, NO. An elevated level of Nos2 expression was found in naïve MSC2, MSC4, and MSC5, the clones representing high inhibitory activity. However, it either was expressed at a lower level or was undetectable in MSC3 and MSC6, respectively, the clones slightly inhibiting T-cell proliferation. The central role of NO in inhibition of T-cell proliferation was also confirmed using NOS2 inhibitor, L-NMMA, which blocked the inhibitory effect of the most immunosuppressive clone, MSC2. These results were in good agreement with previous experimental data [15,16,30]. Expression of Ptgs2 playing a role in PGE2 synthesis showed no significant difference among the clones. Studies examining the function of PGE2 in immunosuppression by murine MSCs produced controversial data, namely PGE2 had important [31], minor [30], or no role [14 –16]. However, the importance of PGE2 was unambiguously shown here by using an inhibitor of PTGS2, indomethacin. Interestingly, blocking NOS2 resulted in full inhibition of T-cell anti-proliferative effect of MSC2, the most immunosuppressive clone while indomethacin only incompletely reversed T-cell growth. Our recent result [32] might serve as an explanation for this phenomenon; PGE2 secretion by MSC seems to be promoted by an NO-dependent signaling pathway, since L-NMMA not only inhibits NO production but also partially diminishes PGE2 secretion, resulting in complete recovery of T-cell proliferation.
Recently, a growing body of data supports the idea that the biological functions of MSCs are strongly influenced by the surrounding microenvironment, especially the inflamed milieu. Activation, termed as licensing [33], is triggered by pro-inflammatory cytokines released from immune cells at the site of inflammation [15,34], although the polarization toward anti-inflammatory phenotype may depend on the concentration and duration of the stimulating agent or the type of the factor itself [33]. The site of inflammation abounds in pro-inflammatory mediators, including IFN-γ and TNF-α [35]. For priming of MSCs' immunosuppressive function, the combination of IFN-γ and TNF-α or IL-1α/β was recommended [15]. The production of numerous cytokines and chemokines involved in MSCs' T-cell inhibitory machinery in mouse or in human (IL-6, IL-8, IL-10, HGF, PD-L1, PGE2, NO, and IDO1) is induced by priming MSCs with IFN-γ and TNF-α [15,31,36,37]. In this study, the combination of IFN-γ and TNF-α was applied for the priming of MSC inhibitory activity to model an inflammatory milieu. The pretreatment of MSC clones with IFN-γ and TNF-α equated the inhibition of T-cell proliferation; hence, clones, MSC3 and MSC6, poorly inhibiting the T-cell proliferation, acquired similarly strong inhibitory activities to those of MSC2, MSC4, and MSC5. This result indicated that stimulation of MSC clones with IFN-γ and TNF-α enhanced the immunosuppressive effect of otherwise nonfunctional single cell-derived MSC cultures. Indeed, licensing MSC clones resulted in a remarkable upregulation of Nos2 and Ido1 and various moderate increases of Ptgs2 gene expression in every MSC clone. On examining the role of inflammatory cytokines, we found that the NOS2 expression was essential since its inhibitor L-NMMA reversed T-cell proliferation in licensed MSC2 and MSC6 clones. Inhibition of PGE2 synthase only partially reversed the suppression by MSC6 but did not affect that by MSC2. These results might allow the conclusion that the MSC population has an intrinsic heterogeneity regarding the mediators of the immunosuppressive function, such as PTGS2.
Induction of Ido1 mRNA expression under inflammatory conditions was the most striking, since naïve clones did not express the gene at all whereas remarkable increase occurred on IFN-γ and TNF-α treatment. In spite of the high gene expression of Ido1, IDO1 inhibitor, 1-MT did not affect the activities of licensed MSC clones. Although IDO1 is an important factor in human MSC [38], its function in mouse MSCs is inconsistent. Even though Ido1 mRNA expression is inducible with pro-inflammatory cytokines, the IDO1 protein expression is undetectable in mouse MSC [39]. One possible explanation was served by Hucke et al., who showed that an accelerated proteosomal degradation of IDO protein occurred on NO production [40]. Accordingly, chemical inhibition of IDO1 in mouse MSCs did not affect MSC function in T-cell proliferation [15,30,38].
Experimental animal models clearly revealed that MSCs could be effective cell-based therapies in immunological pathologies [17,18,41 –43]. Whether the in vivo immunoregulatory property of MSCs was reflected at the clonal level, the effects of the most and least immunosuppressive clones, MSC2 and MSC6, respectively, were examined in the ovalbumin-induced delayed-type hypersensitivity (DTH) model. The in vitro results were validated in vivo, since MSC2 diminished whereas MSC6 did not affect DTH response when naïve MSC clones were applied. In contrast, pretreatment of MSC6 with licensing cytokines IFN-γ and TNF-α before an intraperitoneal injection resulted in a similar blocking effect of MSC6 to that of MSC2, indicating the increasing immunosuppressive function of an otherwise poorly acting clone. Our results were supported by the data obtained in the GVHD model, since preactivation of MSCs resulted in more effective prevention of the disease [17].
Taken together, our experiments clearly show that murine MSC population possesses intrinsic heterogeneity regarding the immunosuppressive function of single cell-derived MSC clones. Variations between the clones can be, at least partly, explained by the different expression of immunoregulatory factors. In this sense, NO is the key mediator while PGE2 is involved and IDO1 has no role in the suppressive activity of MSC clones. An important novel message of this work is that variability in the immune function of the different MSC clones can be abolished by licensing the cells by inflammatory cytokines before application for immunological assays or therapy of immune disorders.
Footnotes
Acknowledgments
The authors would like to thank Mrs. Andrea Gercsó for her excellent technical assistance, Edit Kotogány and Nóra Fehér for flow cytometric analysis. Csaba Vizler, Ernő Duda, and János Haskó are acknowledged for their generous gifts of the H5V cell line and recombinant TNF-α and for helping the authors with the animals, respectively. This work was supported by TÁMOP-4.2.2.A-11/1/KONV-2012-0035. Roberta Fajka-Boja has been supported by János Bolyai Research Fellowship of the Hungarian Academy of Sciences.
Author Disclosure Statement
The authors declare no competing financial interests.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.