
Abstract
For diseases of the brain, the pig (Sus scrofa) is increasingly being used as a model organism that shares many anatomical and biological similarities with humans. We report that pig induced pluripotent stem cells (iPSC) can recapitulate events in early mammalian neural development. Pig iPSC line (POU5F1high/SSEA4low) had a higher potential to form neural rosettes (NR) containing neuroepithelial cells than either POU5F1low/SSEA4low or POU5F1low/SSEA4high lines. Thus, POU5F1 and SSEA4 pluripotency marker profiles in starting porcine iPSC populations can predict their propensity to form more robust NR populations in culture. The NR were isolated and expanded in vitro, retaining their NR morphology and neuroepithelial molecular properties. These cells expressed anterior central nervous system fate markers OTX2 and GBX2 through at least seven passages, and responded to retinoic acid, promoting a more posterior fate (HOXB4+, OTX2−, and GBX2−). These findings offer insight into pig iPSC development, which parallels the human iPSC in both anterior and posterior neural cell fates. These in vitro similarities in early neural differentiation processes support the use of pig iPSC and differentiated neural cells as a cell therapy in allogeneic porcine neural injury and degeneration models, providing relevant translational data for eventual human neural cell therapies.
Introduction
L
To increase the probability of successful human iPSC-based neural cell trials, closely matched animal iPSC-derived neural cells and subtypes should be tested in species that are more representative of the human central nervous system (CNS) in size and other physiological characteristics [10].
The pig is an increasingly relevant model organism for biomedical research with the pig CNS demonstrating significant similarities to humans relative to the rodent model, which suggests pig study results may translate to human patients more readily [15].
In contrast to the rodent, the pig and human brain are gyrencephalic, possessing significantly higher levels of white matter, and they are larger in size, all important factors when considering the development of a cell therapy [16 –18]. For example, brain size is likely to affect the number of cells to be transplanted, the number and location of injection sites, supporting components such as extracellular matrices, and the ability of cells to be vascularized. In addition, recent genomic and proteomic data characterizing the pig at the molecular level will be indispensable in establishing the pig as an appropriate biomedical model [19,20]. Pigs are available for biomedical research through many of the same commercial vendors that supply rodents to the research community and can be obtained as miniature swine that are more comparable to humans [20].
The generation of pig iPSC opens up the possibility of evaluating allograft and autologous iPSC generated neural cell therapies in pig neurodegeneration and injury models [21 –23]. However, the level of translational relevance to humans will be determined in part after studies determine whether pig iPSC are capable of undergoing spatial, temporal, cellular, and molecular neural differentiation events that are similar to human iPSC neural differentiation [24].
Human pluripotent stem cell (PSC) neural developmental paths can be recapitulated in vitro. Initially, these cells form neural rosette (NR) structures, the in vitro equivalent to the neuroepithelium of the developing neural tube. The NR consists of a radial arrangement of neuroepithelial cells with the capacity to differentiate into neurons and glia [5,25,26]. Human NR structures have a broad differentiation potential propitious for development of the central and peripheral nervous systems, allowing regional specification and expansion in vitro without losing rosette properties [27].
Although NR structures have been observed during the transition of pig ESC through neural differentiation [16,28] rosette morphology was only maintained briefly, and their capacity to recapitulate neural differentiation, specifically neural regionalization, was not examined. We previously reported that pig iPSC can differentiate into all three neural lineages but the presence or absence of NR and neuroepithelial cells was not investigated [24]. Because the NR is a distinctive cellular aggregate structure during the initial neuralization of human PSC [25], it is critical to demonstrate that pig iPSC not only form NR but also differentiate through a process similar to human NR-derived cells, supporting the use of pig iPSC as a translational model for human regenerative therapies.
Several groups have generated pig iPSC that have been characterized using pluripotent markers such as stage-specific embryonic antigen 4 (SSEA4) and POU domain class 5, transcription factor (POU5F1) [29 –31],without examining the relative expression of both pluripotency markers and whether this influenced subsequent in vitro differentiation. POU5F1 is the main regulatory gene in governing self-renewal, it also plays a role in determining lineage commitment in the differentiation of rodent and human PSC, while SSEA4 is associated with pluripotency in human but not murine PSC [32,33]. SSEA4 has been used to characterize pig iPSC lines, however, expression varied among studies, accentuating the need to utilize multiple pluripotent marker expression profiles to fully characterize the pluripotency of pig iPSC lines.
The use of multiple marker and differential expression profiles may be correlative with any given pig iPSC lines in vitro lineage differentiation profiles. Human PSC have demonstrated distinct lineage differentiation efficiencies, and one proposal for overcoming this limitation was developed, a lineage scorecard analysis that permits the evaluation of multiple markers to rapidly assess the differentiation propensities of multiple starting populations to predict the quality and utility of any pluripotent cell line [34].
In this study, we determined whether expression of pluripotent markers (POU5F1 and SSEA4) in morphologically uniform pig iPSC lines was correlative with neural development and from this expression profile determined line propensity to form NR structures. Moreover, we assessed the spatial, temporal, molecular, and cellular differentiation patterns of pig iPSC-derived NR development to ascertain parallels to human PSC NR differentiation. Ultimately, we tested the ability of pig iPSC-derived NR to be isolated, expanded in vitro while maintaining a homogenous NR phenotype, and their capability to differentiate into all three neural lineages.
Materials and Methods
Cell culture
Pig iPSC lines were generated as previously described [23,24]. Preliminarily, we generated three putative pig iPSC lines. All lines were <20 passages from the derivation and were continuously maintained on Matrigel (diluted 1:100 in Dulbecco's modified Eagle's medium/F12; BD Biosciences)-coated dishes in mTeSR1 (Stemcell Technologies) medium. Cells were passaged using 1 mg/mL dispase (Stemcell Technologies) every 3–4 days upon reaching 80% confluency.
Immunocytochemistry
Pig iPSC and NR were plated onto glass four-well chamber slides and fixed with 4% paraformaldehyde (PFA; Electron Microscopy Sciences) for 15 min. For intracellular staining, cells were permeabilized with 0.1% Triton X-100 and 1% Polyvinylpyrrolidone (PVP; Sigma-Aldrich) in blocking solution containing 4% normal donkey serum (Jackson Immuno Research). For extracellular staining, cells were blocked in phosphate-buffered saline (PBS) containing 4% normal donkey serum. Primary antibodies were diluted in blocking solution and incubated with the cells at 4°C overnight. Primary antibodies used were POU5F1 (Santa Cruz; 1:500), SSEA4 (Developmental Studies Hybridoma Bank; 1:200), tight junction protein ZO-1 (ZO-1, Mid; Invitrogen, 1:50), PAX6 (Millipore; 1:1000), SOX1 (R&D Systems; 1:25), NESTIN (Millipore, 1:1000), beta III-TUBULIN (βIII-TUB; Neuromics, 1:200), microtubule-associated protein 2 (MAP2, Invitrogen; 1:200), glial fibrillary acidic protein (GFAP, Chemicon; 1:200), and oligodendrocyte marker 4 (O4, Chemicon; 1:100). Primary antibodies were detected using a fluorescently conjugated secondary antibody, Alexa Fluor (Invitrogen; 1:1000) and incubated for 1 h at room temperature before washing and subsequent microscopic inspection. Cells plated on slides were imaged on IX81 microscope with Disc-Spinning Unit (Olympus, Inc.) using Slide Book Software (Intelligent Imaging Innovations). Three dimensional structures were imaged using Z-stack capabilities of the Slide Book Software. Protein expression was quantified using Image-pro plus 5.1 software. Quantitative evaluation was performed by counting immune-labeled cells from three independent experiments.
Neural induction
POU5F1low/SSEA4low, POU5F1high/SSEA4low, and POU5F1low/SSEA4high pig iPSC lines were disassociated using dispase and plated on Matrigel in mTeSR1 medium at a density of 12,500 cells/cm2 and cultured at 37°C in 5% CO2. After 24 h, pig iPSC were then placed in a chemically defined neural induction (NI) medium: DMEM/F12, N2 (Gibco), 2 mM
Differentiation of pig iPSC-derived NR
To evaluate the differentiation potential of pig iPSC-derived NR into neurons and glial cells, NR at passage 3 were manually disassociated and seeded at a ratio of 1:3 on Matrigel-coated dishes and at a ratio of 1:10 on Matrigel-coated four well Permanox chamber slides (Lab-Tek). For neuronal differentiation, pig iPSC-derived NR were maintained for 20 days in medium containing basal neural medium AB2 (ArunA), 1X ANS (ArunA), 2 mM
RNA isolation and polymerase chain reaction
Total RNA was extracted using the RNeasy mini kits (Qiagen) according to the manufacturer's protocol. The RNA quantity and quality was measured on a NanoDrop 8000 (Thermo Scientific). Five hundred nanograms of total RNA was used for reverse transcription with the iScript cDNA synthesis kit (BioRad) following the manufacturer's protocol. For reverse transcription polymerase chain reaction (RT-PCR) analysis, PCR amplification was performed using GoTaq Green master mix (Promega), primers were designed using Primer-BLAST software [35] or based on literature (Supplementary Table S1; Supplementary Data are available online at
Statistical analysis
Immunocytochemistry data were analyzed using one-way ANOVA (SAS ® 9.3) followed by Tukey's LSD post hoc test to determine significant differences between groups. Statistically significant differences are defined at the 95% confidence index (P<0.01).
Results
POU5F1high/SSEA4low pig iPSC demonstrate a higher rate of differentiation toward NR
Initially, we assessed the pluripotent characteristics of three porcine iPSC lines. Pig iPSC exhibited typical characteristics associated with human ESC and iPSC including polygonal shape, a high nuclear-to-cytoplasmic ratio, and a single nucleus with multiple prominent nucleoli (Fig. 1A–D). Pig iPSC line L2 exhibited high expression of POU5F1 with 94.10% positive cells versus 64.67% and 63.00% of cells in lines L1 and L3 respectively (P<0.01; Fig. 1F–H, M). Expression of SSEA4 was higher (P<0.01) in line L3 with 82.60% of cells being positive relative to lines L2 and L1, which were 32.33% and 29.20% respectively (Fig. 1J–L, N). All pig iPSC were negative for NR markers PAX6 and SOX1 before neural induction (data not shown). Based on these results, we will subsequently identify lines L1, L2, and L3 as line POU5F1low/SSEA4low, line POU5F1high/SSEA4low, and line POU5F1low/SSEA4high respectively.
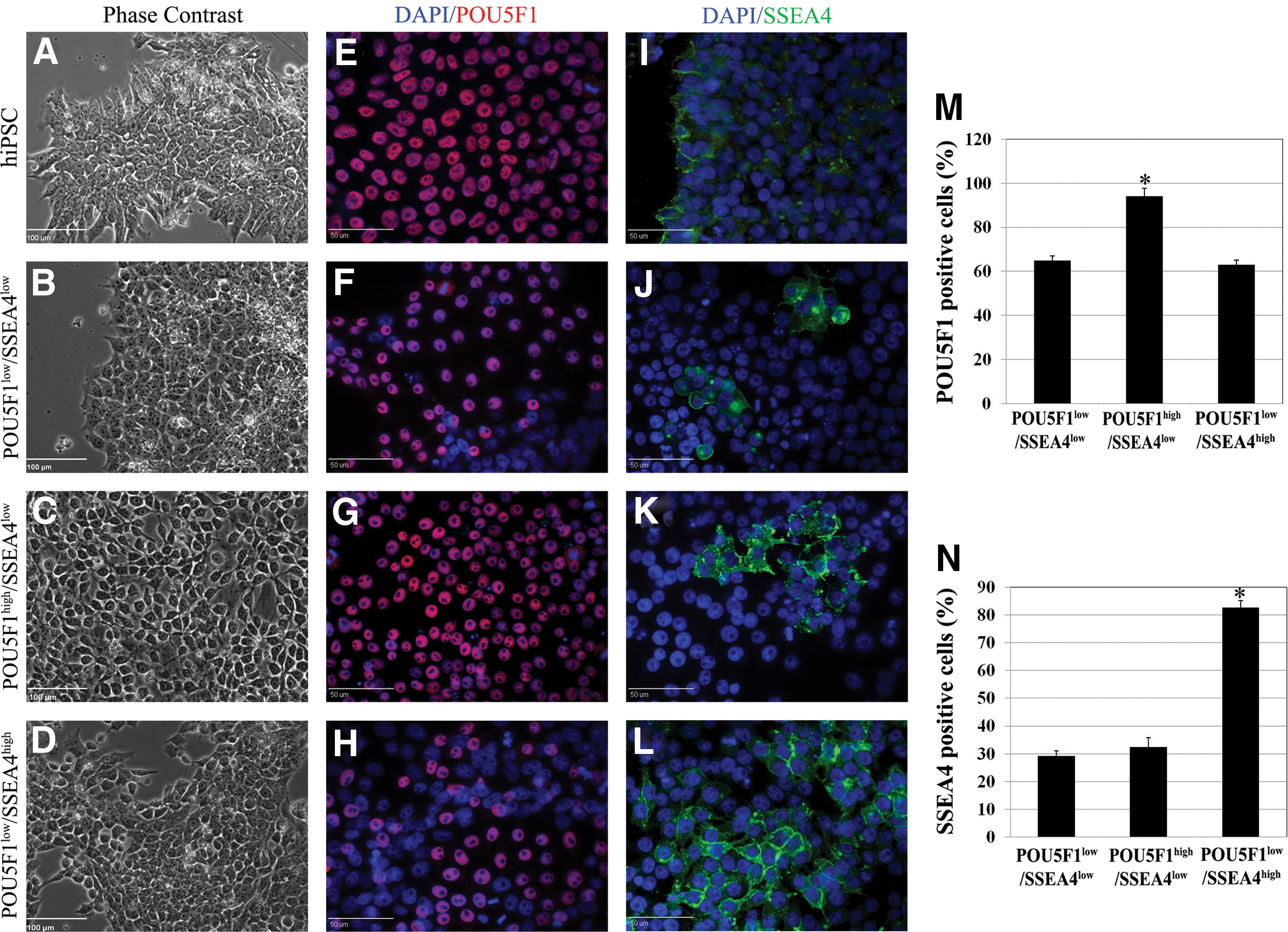
Pig induced pluripotent stem cell (iPSC) lines POU5F1low/SSEA4low, POU5F1high/SSEA4,low and POU5F1low/SSEA4high differ in POU5F1 and SSE4A protein expression. Morphology and pluripotency were evaluated among all three pig iPSC lines and compared to human iPSC (hiPSC)-positive controls before initiating the neural induction (NI) process.
Although all lines phenotypically resembled pluripotent cells superficially, they differed in POU5F1 and SSEA4 expression suggesting potential differences in plasticity and neural differentiation. To test whether pig iPSC populations with different POU5F1 and SSEA4 expression profiles demonstrated differing early neural induction potential as evidenced by formation of NR, we compared all three pig iPSC lines after transitioning into NI medium. After approximately 5 days of neural induction, the three pig iPSC lines began to form colonies but only colonies from POU5F1high/SSEA4low pig iPSC line were surrounded by fibroblast-like cells (Supplementary Fig. S1B).
After 9 days of induction, cells in the colonies elongated and organized into columnar epithelial cells (Fig. 2A and Supplementary Fig. S1E), followed by further organization of the epithelial cells into NR after an additional 4–5 days in culture (Fig. 2B, C and Supplementary Fig. S1H). Pig iPSC line POU5F1high/SSEA4low produced more colonies at day 9 and 13 (4.8X) than the other lines (Fig. 2D). By day 13, 63.5% of POU5F1high/SSEA4low line-derived colonies contained rosette structures (Fig. 2E). Pig iPSC line POU5F1low/SSEA4low failed to form NR (Supplementary Fig. S1G) while POU5F1low/SSEA4high generated limited NR formation in one instance, but did not produce the replicable, robust colony formation characteristic of POU5F1high/SSEA4low cells (Supplementary Fig. S1I).
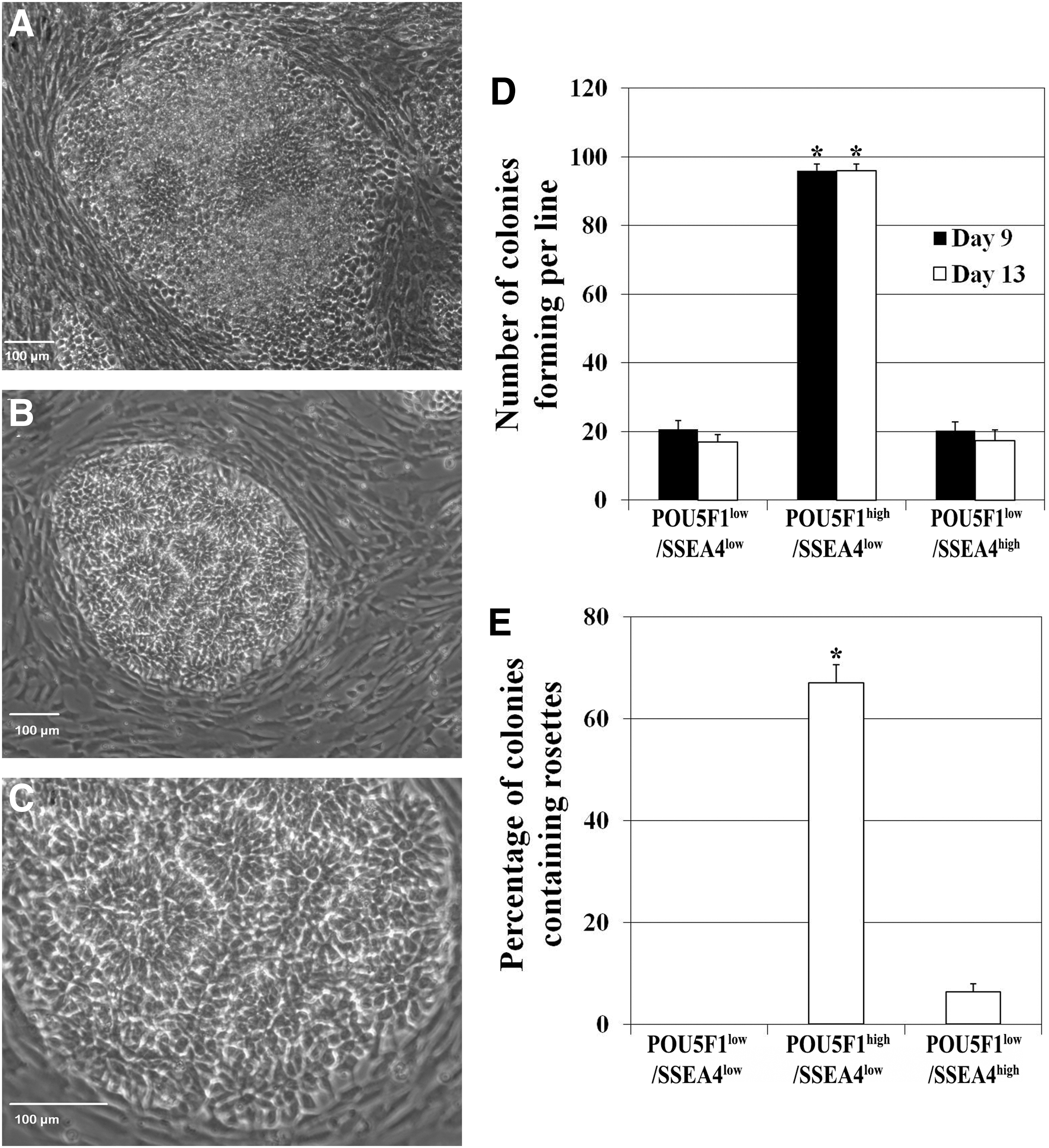
Pig iPSC line POU5F1high/SSEA4low is more suitable for neural rosette differentiation.
Two developmental stages have been identified during neuroepithelial differentiation of human PSC, the early primitive NR, that is characterized by columnar epithelial cells positive for PAX6 and negative for SOX1, and the later definitive neuroepithelial NR, that is characterized by columnar cells forming neural tube-like rosette expressing PAX6 and SOX1 [5,36,37]. To test whether our cultures transition through these two early stages during early neural induction, protein expression was evaluated by immunocytochemistry.
Immunostaining data indicated that before neural induction, POU5F1 (Fig. 3A)-positive pig iPSC were PAX6 and SOX1 negative (Fig. 3D) and ZO-1 (Fig. 3G), a tight junction marker, was evenly expressed on the surface of pig iPSC indicative of an undifferentiated state. However, 9 days postneural induction, immunocytochemistry indicated that the columnar epithelial cells in NR uniformly expressed PAX6 but not SOX1 and that ZO-1 marker started to localize at the apical border of NR (Fig. 3E, H). At day 13, NR cells were positive for both PAX6 and SOX1, and ZO-1 was localized to the lumen of NR (Fig. 3F, I). Moreover, our results showed a pronounced decrease of POU5F1 within the NR structures after 13 days in neural induction (Fig. 3A–C). These results demonstrate that pig iPSC transition through two neural developmental stages similar to human PSC.
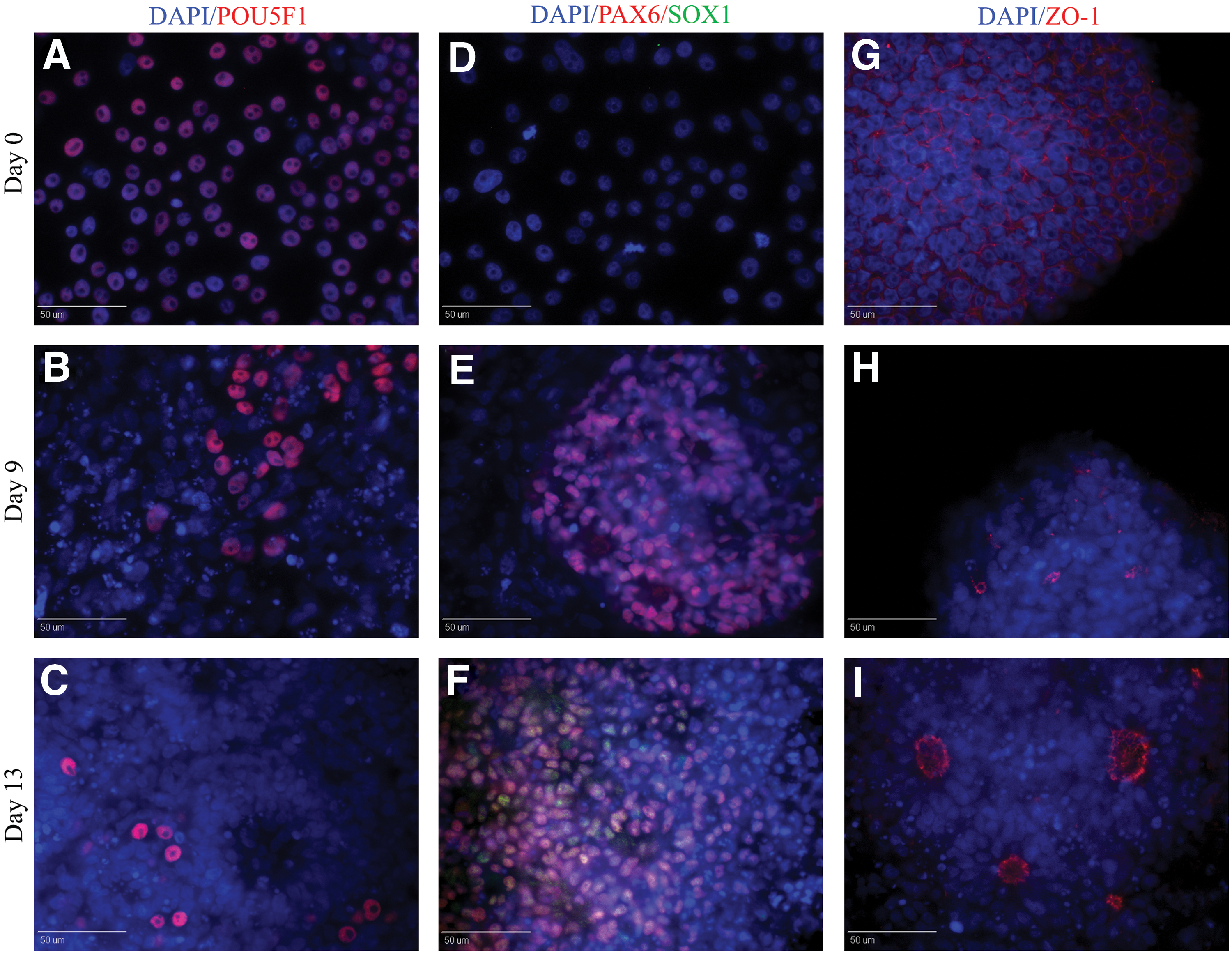
Pig iPSC-derived NR cells follow the same temporal course of neural differentiation as human pluripotent stem cells. At day zero, pig iPSC were positive for POU5F1
Pig iPSC-derived NR cells maintain NR phenotype after multiple passages
We next investigated whether pig iPSC-derived NR share features of developing human neuroepithelium, including polar organization and expression of NR-associated transcripts and proteins, and if these characteristics were conserved through multiple passages and after cryopreservation [27]. The cultures exhibited pronounced continuous self-organization into NR structures at early passages (Fig. 4A), but were less prevalent by passage 7 (Fig. 4E). However, at passage 3 and 7 the NR cells expressed the NR transcription factors PAX6 and SOX1 (Fig. 4B–F) and were positive for NESTIN (Fig. 4D, H). In addition, the ZO-1 marker was detected at the luminal surface of the rosettes in both passages, reflecting polarized organization (Fig. 4C–G).
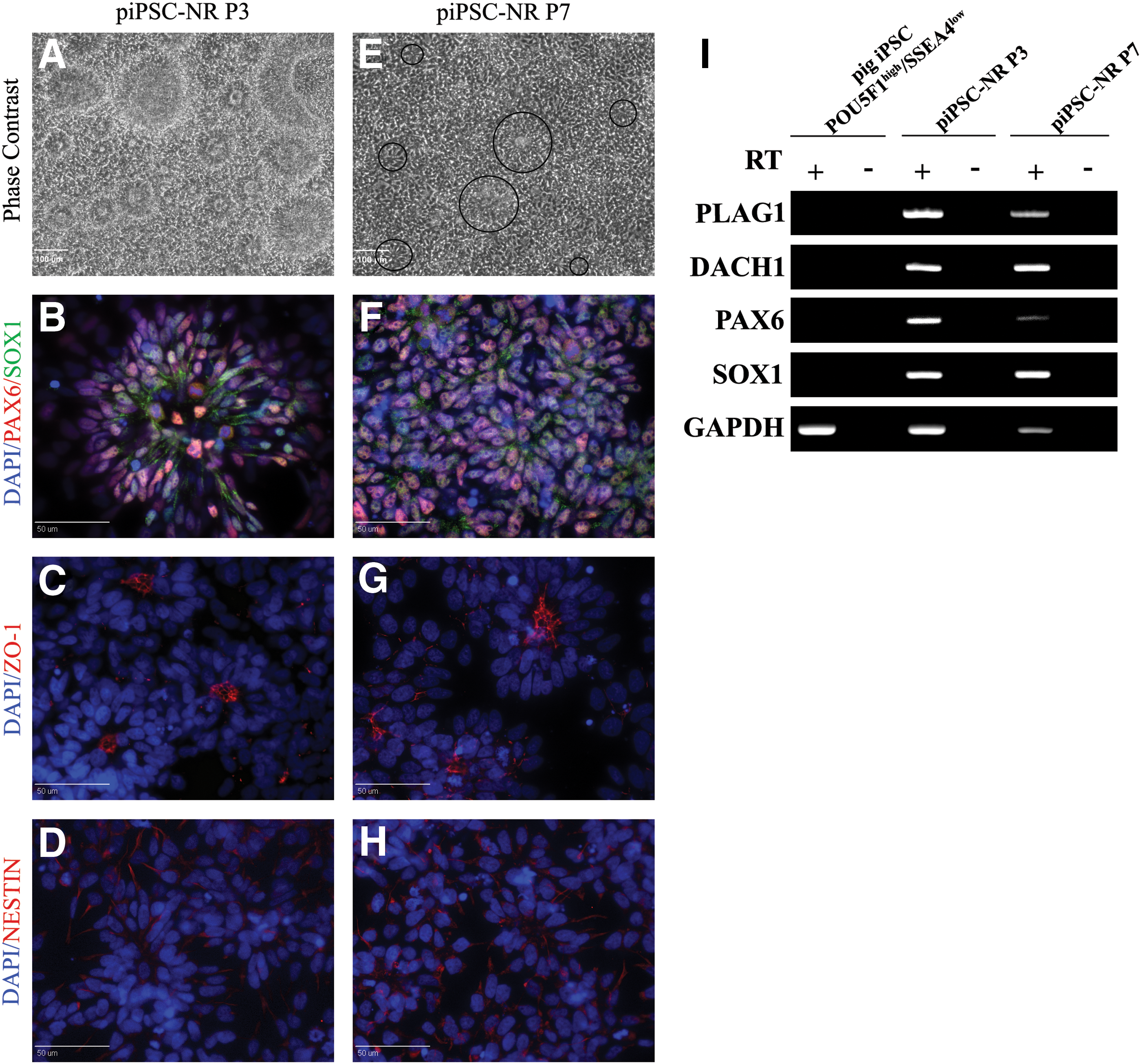
Pig iPSC-derived NR maintain rosette properties over multiple passages.
Moreover, pig iPSC-derived NR could be cryopreserved and thawed without alteration in polarity (ZO-1) or NESTIN, PAX6, and SOX1 all of which are expressed in human NSC and NR cell [36,38] (Supplementary Fig. S2A–D). PLAG1 and DACH1 genes, mouse and human NR-specific markers, were expressed at equivalent levels in pig iPSC-derived NR passage 3 and 7, yet these were negative in the POU5F1high/SSEA4low pig iPSC and no reverse transcriptase (RT) negative controls. In addition, transcript expression for PAX6 and SOX1 in pig iPSC-derived NR cells concurred with the immunocytochemical data (Fig. 4I). These results demonstrated that pig iPSC-derived NR could be maintained for multiple passages without loss of NR phenotype.
Enriched pig iPSC-derived NR cells can be efficiently patterned to posterior fate
Pig iPSC-derived NR cell cultures differentiated in neural induction medium initially expressed anterior transcription factors OTX2 and GBX2, but not posterior-associated HOXB4 and HOXB6 genes (Fig. 5A). Dorsal marker PAX7 was detected at passage 7 but not at early passage; and ventral marker NKX6.1 was not detected (Fig. 5A). We next determined whether pig iPSC-derived NR passage 3 and 7 remain receptive to instructive regionalization cues and could be induced to form other regional phenotypes, such as the posterior hindbrain or spinal cord fate. To analyze the potential induction of more posterior cells fates, we exposed the cells to 1 μM retinoic acid for 6 days. This study resulted in an induction of posterior HOX genes, HOXB4 and HOXB6 (Fig. 5B). These data show that the restricted anterior regionalization of pig iPSC-derived NR can be effectively patterned to caudal progenitors using a defined morphogen.
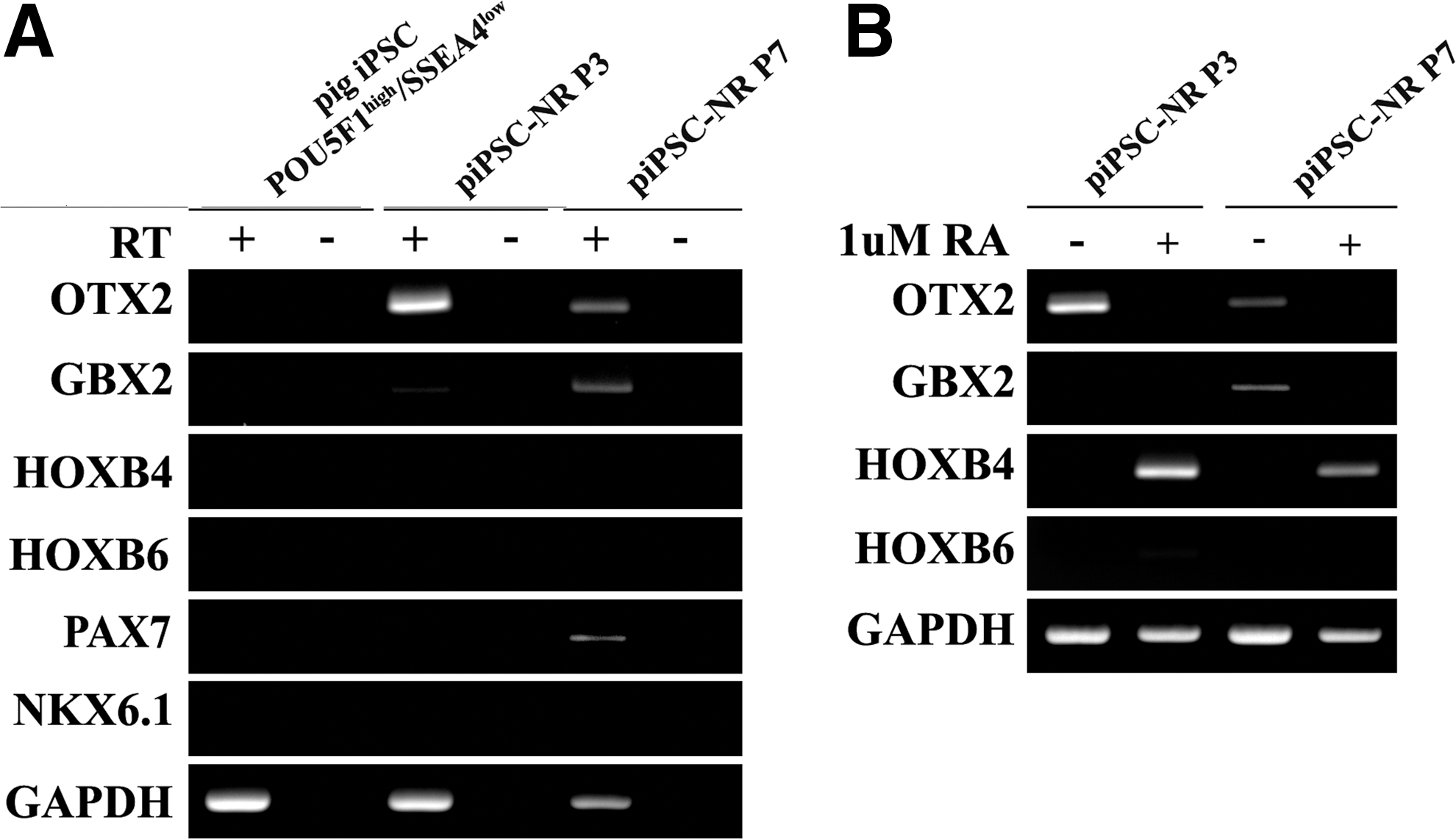
Pig iPSC-derived NR cells adopt polarized neuroepithelial structures of anterior central nervous system (CNS) and remain responsive to instructive regionalization cues
Pig iPSC-derived NR cells differentiated into neurons and glial cells
To determine pig iPSC-derived NR neural differentiation potential, FGF2 was removed from the medium of passage 3 and 7 pig iPSC-derived NR cultures using the same method we previously used with pig iPSC [24]. As expected, pig iPSC-derived NR populations were negative for the pluripotent marker POU5F1 from day 0 to day 20 (Fig. 6A–C), and highly positive for the neural marker βIII-TUB (Fig. 6D–F). At 10 day post-FGF2 removal, MAP2-positive cells were detected (Fig. 6E) and visibly increased through day 20 (Fig. 6F). A subset of pig iPSC-derived NR differentiated into GFAP-positive astrocytes (Fig. 6I); however, O4-positive oligodendrocytes were not detected.
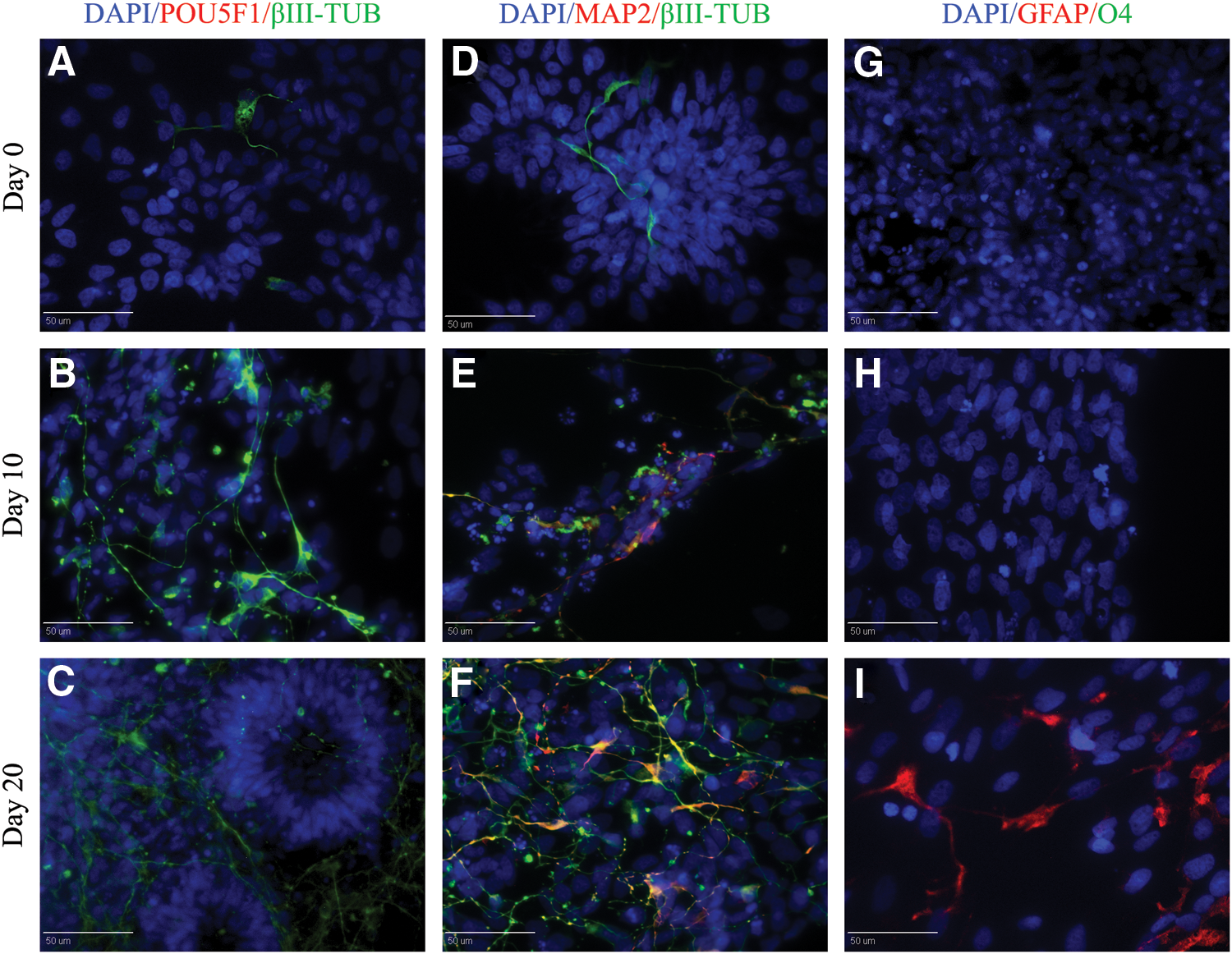
Differentiation of pig iPSC-derived NR. Upon growth factor withdrawal pig iPSC-derived NR predominantly differentiated into βIII-TUB and MAP2-positive cells.
Discussion
Porcine iPSC cell in vitro characterization has widely varied among labs [23,24,29 –31]; therefore, we set out to determine whether basic initial pluripotency marker profiles could provide a means of screening cell lines for early neural differentiation potential. Indeed, the POU5F1high/SSEA4low pig iPSC were more suitable for NR formation than POU5F1low/SSEA4low or POU5F1low/SSEA4high lines. We also found that our culture system allows direct inspection, analysis, and control of the process of neural specification of pig iPSC without the confounding of coculture [28,39] or uncharacterized media constituents [24,28] that hinder analysis of developmental mechanisms or results in components that are not compatible with clinical applications [40].
We have demonstrated for the first time the generation of NR from pig iPSC, producing cells that can be isolated, regionally specified, and expanded in vitro without losing rosette-forming properties. This finding not only supports similarities in neural development between human and pig cells, but also opens the possibility of generating diverse region-specific lineages of cells from the pig nervous system. These results strengthen the potential for the use of the pig animal model in allograft and/or autologous cell transplantation studies for neurological disorders.
Differentiation potential of mouse and human PSCs into a desired cell lineage varies across cell lines [41,42]. Moreover, a lineage score card has been described as an algorithm test that predicts the differentiation potential of human PSCs [34]. The transcription factor POU5F1 plays an important role in the maintenance of the PSC, but it can also regulate early lineage differentiation [43]. The pig iPSC line with POU5F1high expression led to a robust contribution to early neuroectoderm. This result was consistent with a recent report where efficient mouse PSC in vitro and in vivo differentiation into neural lineages was observed in constitutive POU5F1 expressing lines [44]. Several studies have reported the involvement of POU5F1 in ESC differentiation in vitro [45,46].
Our results are consistent with a model in which the starting porcine iPSC population should have POU5F1high expression before the initial stages of neural differentiation [47]. Here, pig iPSC lines with POU5F1high/SSEA4low expression were more suitable for NR formation. SSEA4 is an embryonic ganglioside antigen with globe-series carbohydrate chains as main structure [48], which is expressed in the inner cell mass (ICM) in preimplantation human embryos [49]. Although SSEA4 is routinely used as surface marker for human ESC [49,50] and iPSC [51,52], the functional significance of this antigen is unclear since SSEA4 downregulation appears to be dispensable for the pluripotent stage [53].
In pig, the SSEA4 expression varies among iPSC lines and studies. While this antigen has not been reported during pig embryo development [54], SSEA4 expression has been detected inconsistently and at various levels in pig ESC [55,56] and iPSC [23,24,29,31]. We found that SSEA4 expression was not required for NR formation and perhaps low expression in iPSC may lead to more directed NR formation. Taken together, we believe that high levels of POU5F1 expression is critical when choosing lines specifically for neural differentiation, while SSEA4low could indicate that its function is more important for cellular differentiation, in agreement with the reports for human PSCs [53,57].
During human ESC differentiation, POU5F1 is downregulated before PAX6, an early neural transcription factor, becomes highly expressed [43,58]. Our study has demonstrated that during neural differentiation of pig iPSC, POU5F1 is progressively downregulated while neural marker expression is gradually upregulated. (Fig. 3). Moreover, NR derived from line POU5F1high/SSEA4low followed the same temporal protein expression progression as observed in human PSCs. Our study revealed the uniform expression of PAX6 before SOX1, similar to human neuroepithelial cells [5,36,37,58,59]. This differs from the patterned expression of PAX6 after SOX1 expression in other animals, including mouse [60,61]. The pig iPSC-derived NR displayed the same polarity as human NR [27,59] and in the in vivo neural tube [62].
Pig iPSC-derived NR displayed characteristic and distinctive luminal distribution of the ZO-1 tight junction protein in the center of NR, revealing that pig iPSC-derived NR were able to acquire apicobasal polarity. These findings suggest an analogy between human and pig PSC in response to neural differentiation conditions, further corroborated by a comparison of gene and proteomic expression profile in pig and human ESC [19,20]. Given these recent findings the use of pig iPSC differentiation has developmental parallels with human PSCs, and it represents an additional in vitro model for investigating the molecular mechanisms of neural induction in mammals.
Previous studies have shown that continued culture of NR cells in FGF/EGF resulted in spontaneous differentiation and loss of rosette morphology [27]. However, we observed that passage 3 and 7 pig iPSC-derived NR retained their characteristic morphological and immunocytochemical properties in the presence of FGF2. These cells did not express POU5F1 and continued expressing early neural markers such as NESTIN, PAX6, and SOX1 and maintained expression of ZO-1. Moreover, the presence of PLAG1 and DACH1 genes, which have been identified as mouse-associated NR-specific markers [62] and human NR cells with the highest levels of plasticity [27]; and both were strongly expressed in pig NR out to passage 7. These findings suggest that neural development is a conserved program among mice, human, and pig PSCs and that pig NR can be maintained in extended culture without loss of NR phenotype.
During neural differentiation of human PSC, it was demonstrated that NR adopt anterior CNS fate indicated by the expression of the anterior markers BF1 and OTX2 and absence of posterior homeobox protein HOXB4. In addition, pig iPSC-derived NR lacked the expression of markers that define specific dorsal-ventral domains during neural tube formation [5,27]. The anterior marker expression and absence of defined dorsal-ventral polarity of NR suggest that NR may resemble the neuroepithelium of neural plate stage neural development before dorsal-ventral specification [58,63].
Our data also suggest that pig iPSC-derived NR respond to standard in vitro conditions utilized to induce anterior CNS fate, exhibiting a regional transcription profile characteristic of anterior CNS cells, which could be respecified toward caudal fate upon retinoic acid treatment. However, GBX2 (anterior hindbrain marker) and the dorsal marker PAX7 were only detected at low levels in passage 7 cells suggesting a posteriorization and dorsalization of pig iPSC-derived NR at late passages. These findings could be due to FGF2 used in cell culture. This growth factor, used to stimulate proliferation of NSC culture, induces posterior cell fates during nervous system development [64,65], altering regulation of normal dorsal ventral fate [66]. Therefore, longer exposure to FGF2 could promote posteriorization and a dorsal pattern in late passage of NR.
Although NR structures have been reported during pig ESC differentiation, these structures were only transient and lost after three passages [16,28]. Therefore, the downregulation of pluripotent markers and full patterning and wide plasticity of NR toward CNS lineages were not previously reported in the pig. Our findings suggest that, under standard conditions pig iPSC-derived NR progress toward an anterior patterning state, retain rosette properties and can be modulated toward more posterior fate using defined morphogens similar to human NR [36,67].
During embryonic development, the formation of major neuronal cell types precedes formation of glial cells, including astrocytes and oligodendrocytes [68]. In our study, the same neurogenesis to gliogenesis sequence was observed (Fig. 6). After withdrawal of FGF2, we noted that NR predominantly differentiated into neurons, found mostly in the periphery of NR. This finding suggests that cells undergoing differentiation migrate away from the expanding NR, similar to newborn neurons migrating away from the germinal layers (ventricular zone and subventricular zone) during human neurogenesis [37].
By the end of the differentiation process, mature neurons co-expressed βIII-TUB and MAP2, and GFAP-positive astrocytes were observed, which confirmed the temporal sequence of neuronal and glial differentiation previously reported in vivo in pig and human [68,69]. However, O4-positive oligodendrocytes were not detected within 20 days of culture. Previous reports indicated that the oligodendrocyte lineage is infrequently observed or not reported during ESC neural differentiation [25,70,71]. This suggests that oligodendrocyte derivation potentially requires extended cultures for differentiation or specific growth factors have to be added to the culture medium to develop oligodendrocyte precursor cells (O-2A) [72]. Nevertheless, in our previous study [24], O4+ oligodendrocytes were directly differentiated from pig iPSC using similar culture conditions as the current study but did not transition through obvious NR stages.
This information corroborates the observation in vivo during early neural development, when early neuroepithelial cells are responsible for initiating neurogenesis in the developing CNS before they give rise to glial cells [73]. Pig iPSC-derived NR have the capacity to reconstitute the normal timing program of the developing CNS observed in mammals [68,69], suggesting that both neurogenesis and gliogenesis are conserved across species. Taken together, these studies indicate that there are many parallels in the development of neural cell types in pig and human iPSC. This supports other recent findings where human iPSC differentiation protocols have been used to differentiate pig iPSC into neurons, astrocytes, and oligodendrocytes—and more recently into even more specialized cell types, including photoreceptors [74] and motor neurons [75].
Conclusion
In this study, we demonstrated expression of POU5F1 at an iPSC level and its interaction with differential expression of SSEA4 was a key determinant to initiate development of early neural events in pig iPSC. The POU5F1high/SSEA4low pig iPSC demonstrated the ability to undergo robust and developmentally ordered neural differentiation into NR. Furthermore, the pig iPSC-derived NR shared many characteristics with human NR, such as expression of specific NR markers, anterior CNS phenotype, respecification to more posterior fate by retinoic acid, and the ability to differentiate into both neurons and glia cells. Taken together, these morphological and functional features indicate that rosettes derived from pig iPSC are indeed a close in vitro representation of early neurulation events in human cells and support the use of pig iPSC-derived NR and NSC derived from them are a reliable large animal model for the development of neural iPSC therapies and an attractive tool for translational research in regenerative medicine.
Footnotes
Acknowledgment
This work was supported by the Georgia Research Alliance endowment.
Author Disclosure Statement
The authors declare no conflicts of interest.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.