
Abstract
Podocyte depletion is a key event in the progression of end-stage kidney disease (ESKD) resulting in nephrotic proteinuria and renal failure, but the treatment options are limited to dialysis and renal transplantation. So there is an urgent need for renal regenerative therapies. Generation of podocytes from human stem cells is regarded as a promising therapeutic strategy to repair or regenerate the damaged kidneys; however, the reliable induction system remains a challenge. In this study, we established a two-stage induction protocol for podocyte generation from human adipose-derived mesenchymal stem cells (hAD-MSCs). We initially established a condition that induces hAD-MSCs toward intermediate mesoderm cells with activin A and high concentration of retinoic acid (RA). Subsequently, by using the combination of activin A and low concentration of RA and BMP7, we generated podocyte-like cells expressing multiple podocyte-specific markers and able to integrate into a developing nephron of embryonic kidney explant culture and ameliorate proteinuria and kidney injure in adriamycin-treated mice. Furthermore, we identified that miRNA-498 inhibitor has potential to improve the differentiation of hAD-MSCs into podocyte-like cells and established a robust induction protocol. Thereby, our study advocated an efficient method for the induction of kidney podocyte-like (iPod) cells from hAD-MSCs and provided an ideal candidate for regenerative therapies of the kidney.
Introduction
P
Mesenchymal stem cells (MSCs) were known as the most attractive adult stem cells for regenerative medicine in recent years, which are multipotent low-immunogenic cells with easy accessibility from various tissues [9]. Also, numerous researches, including clinical studies, have confirmed the promise of MSC-based therapy for a broad spectrum of human diseases [10 –12]. As we know, kidney is a mesodermal organ that differentiates from the intermediate mesoderm (IM), and podocytes arise from metanephric mesenchymal cells by a mesenchymal to epithelial transformation (MET) process [13,14]. Thus, MSCs, which are able to differentiate into a variety of mesodermal lineage cells [15], might be an ideal promising candidate for kidney podocyte generation in regenerative medicine.
Biological events, such as development, are regulated and controlled by an extremely complicated network. More and more evidences have shown that cell fate does not only rely on the regulation of proteins, for example cytokines, but also some other regulatory molecules, such as microRNAs (miRNAs). MicroRNAs are endogenous noncoding RNAs that negatively regulate the gene expression at the posttranscriptional level, predominantly by binding to the 3′-untranslated region (UTR) of target mRNAs through sequence pairing [16 –18], and play an important role in various biological processes, including organ development and cellular differentiation [19 –21]. We considered that it might be an ideal strategy to increase the efficiency of a specific cell generation by adding miRNA mimics or inhibitors, which could faithfully mimic the physiological processes of development and indeed be a safe method for regenerative medicine.
In this study, we successfully induced kidney podocyte-like (iPod) cells from human adipose-derived mesenchymal stem cells (hAD-MSCs) with kidney development-related growth factors, and by adding miR-498 inhibitor to this system, we have established a robust induction protocol. This finding provided an ideal strategy to generate podocytes for the kidney regenerative medicine and proposed a novel continuing optimization solution.
Materials and Methods
Isolation and culture of MSCs from adipose tissue
Human adipose tissue was obtained from liposuction aspirates with informed consent of the donors and was performed according to procedures provided by the Ethics Committee at the Chinese Academy of Medical Sciences and Peking Union Medical College. The isolation and culture procedures were described in our previous report [22]. hAD-MSCs were resuspended in a 12 mL culture medium and seeded at a density of 3 × 106 cells in a 75-cm2 culture flask. Cultures were maintained at 37°C in a humidified incubator with 5% CO2 and passaged with trypsin/EDTA when cells were confluent.
Differentiation of hAD-MSCs into podocyte-like cells
The stage 1 basic medium (BM1) was the DMEM/F12 (Gibco) supplemented with 2% fetal bovine serum (FBS; Gibco) and 10 ng/mL activin A (Peproptech). hAD-MSCs of third passage were seeded at a cell density of 5 × 105 cells/well in six-well plates. After adherence, the culture medium was switched to the BM1 with different concentrations of retinoic acid (RA) (0, 0.1, 1, 10 μM) for 3 days. The effect of RA concentration on the generation of kidney podocytes from hAD-MSCs was evaluated by measuring the expression of IM genes. Based on the preliminary results, 10-μM RA was used in the stage 1 differentiation. After 3 days of culture, cells were divided into three groups. In the first group, cells were cultured in the stage 2 basic medium (BM2) (DMEM/F12 + 2% FBS + 10 ng/mL activin A + 0.1 μM RA) with 20 ng/mL BMP7 (Peprotech). In the second group, cells were cultured in the BM2 with 20 ng/mL GDNF (R&D). In the third group, cells were cultured in the BM2 with 50 ng/mL Wnt4 (R&D). All of these groups were cultured for an additional 6 days after stage 1. The effects of the growth factors on podocyte generation were evaluated by measuring the expression of podocyte-specific genes. Based on the preliminary results, 20 ng/mL BMP7 was used in the following experiments.
Transfection of miRNAs
The synthetic miR-498 mimics, miR-498 inhibitor, negative control of mimics (NC), and the inhibitor control (NCI) were purchased from GenePharma. The synthetics were transfected into cells with lipofectamine 2000 (Invitrogen) according to the manufacturer's instruction. Transfection efficiency was assessed by the percentage of fluorescent-positive cells 24 h after transfection. After transfection, the medium was changed into an induction medium for differentiation.
RNA isolation and quantitative real-time polymerase chain reaction
Total RNA was extracted with TRIzol reagent (Invitrogen), according to the manufacturer's instructions. Two micrograms of total RNA from each sample was reverse transcribed using M-MLV (Takara) in a final volume of 30 μL. Quantitative real-time polymerase chain reaction (qRT-PCR) analyses were performed in triplicate using SYBR Green master mixture (Takara) and gene expression data were detected using ABI StepOnePlus System (Life Technologies). The gene expression level was normalized to β-actin. The expression of mature miRNAs was determined using miRNA-specific qRT-PCR. The expression level was normalized to U6. Relative expression of mRNA or miRNA was evaluated by the 2−ΔΔCt method. The primer sequences used in our study are listed in Supplementary Table S1 (Supplementary Data are available online at
Immunofluorescence staining
The cultured cells were fixed at 4°C in ice-cold methanol for 10 min, washed thrice in phosphate-buffered saline (PBS), and then permeabilized in 0.1% Triton X-100/PBS for 10 min at room temperature. Nonspecific binding was blocked with 0.5% Tween-20/PBS containing 1% bovine serum albumin for 30 min. The primary antibodies were incubated at 4°C overnight. The secondary antibodies were incubated for 1 h at room temperature. The incubated cells were washed in PBS, and Hoechst 33342 (Sigma-Aldrich) was used to visualize nuclei. Antibodies used were as follows: Osr1 (Rabbit IgG; Abcam), WT1 (Goat IgG; R&D), Pax2 (Rabbit IgG; Abcam), Podocin (Rabbit IgG; Abcam), Nephrin (Sheep IgG; R&D), Synaptopodin (Rabbit IgG; Abcam), Laminin (Rabbit IgG; Abcam), human nuclear antibody (HNA, Mouse IgG; Merck), Chicken anti-rabbit IgG fluorescein isothiocyanate (FITC; Santa Cruz), Alexa Fluor 488 Chicken anti-mouse IgG (Molecular Probe), Alexa Fluor 555 Goat anti-rabbit IgG (Molecular Probe), Alexa Fluor 555 Donkey anti-goat IgG (Molecular Probe), Alexa Fluor 555 Donkey anti-sheep IgG (Molecular Probe). Conditionally, immortalized mouse podocytes were purchased from the Cell Resource Center at the Chinese Academy of Medical Sciences (CAMS, Beijing, China) and used as the positive control.
Flow cytometry assays
The cultured cells were analyzed for cell antigen expression by fluorescence-activated cell sorting (FACS). To detect intracellular Ags, the Cytofix/Cytoperm Fixation/Permeabilization kit (BD Biosciences) was used according to the manufacturer's instructions. Briefly, resuspend the cells and add 100 μL Fixation/Permeabilization solution for 20 min at 4°C. Wash cells twice in the 1× BD Perm/Wash™ buffer. Cells were incubated at 4°C for 1 h with the following primary antibodies or isotype antibodies, which served as negative controls to set the quadrants in each graph. Then, cells were incubated for 30 min in secondary antibodies conjugated with FITC at 4°C. After washing, cells were fixed and the fluorescence intensity was detected using BD Accuri C6 flow cytometer.
Western blot analysis
Cells were harvested in the RIPA lysis buffer (Beyotime) with 1 mM phenylmethanesulfonyl fluoride, quantified by the BCA Protein Assay Kit (Beyotime), separated by 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis, and transferred to polyvinylidene fluoride membranes (Millipore). The membranes were blocked in 5% nonfat milk in TBST for 1 h, and incubated in primary antibodies overnight at 4°C, followed by horseradish peroxidase (HRP)-conjugated secondary antibodies for 1 h at room temperature. Signals were visualized with an Immobilon Western Chemiluminescent HRP Substrate (Millipore) and detected by ImageQuant LAS 4000 mini imaging system (GE Healthcare). β-actin was used as an internal control. In some instances, the membranes were stripped and incubated with different antibodies. Primary antibodies used were as follows: WT1 (Goat IgG; R&D), Podocin (Rabbit IgG; Abcam), Nephrin (Sheep IgG; R&D), Synaptopodin (Rabbit IgG; Abcam), and β-actin (mouse IgG; Santa Cruz); and HRP-conjugated anti-rabbit (NeoBioscience), HRP-conjugated anti-goat (NeoBioscience), HRP-conjugated anti-mouse (NeoBioscience), and HRP-conjugated anti-sheep (R&D) secondary antibodies were applied.
Reaggregation assay
All animal experiments were approved by the Animal Care and Use Committee of the Chinese Academy of Medical Sciences and Peking Union Medical College. The reaggregation assay was performed as previously described [23]. Kidney rudiments were dissected from E12.5 embryo of pregnant mice and digested with trypsin/EDTA solution for 4 min at 37°C, 5% CO2. The enzyme was then quenched by adding kidney culture media (DMEM+10% FBS) and incubating at 37°C for 10 min for recovery. Digested kidney rudiments were then transferred to a microcentrifuge tube with an additional kidney culture medium and dissociated by repeated trituration. 8 × 105 embryonic kidney cells were recombined with 10% hAD-MSC-derived cells and then centrifuged at 800 g for 2 min to form a pellet and placed on a 5-μm polycarbonate filter supported by a metal grid in the kidney culture medium [DMEM+10% FBS+1.25 μM Glycyl-H-1152 dihydrochloride (H-1152) (Tocris)] for 24 h. After the initial incubation, the media were replaced without H-1152 for an additional 3 days of culture. The pellets were fixed with 4% formaldehyde embedded in paraffin and sectioned with 5-μm thickness. Sections were blocked with horse serum for 1 h at room temperature, and then antigen retrieval was performed. Primary antibodies were incubated overnight at 4°C, and secondary antibodies were incubated for 1 h at room temperature and counterstained with Hoechst 33342.
Animal model
All animal experiments were approved by the Animal Care and Use Committee of the Chinese Academy of Medical Sciences and Peking Union Medical College. Male BALB/c mice aged 8 weeks were divided into control, adriamycin model (ADR), and iPod-treated ADR model groups. Renal damage was induced through a single tail-vein injection of 10 mg/kg adriamycin. In the iPod-treated group, 2 × 105 iPod cells were injected intravenously 3 days after the adriamycin injection. On day 17, urine was collected for 24 h using a metabolic cage (n = 6 per group). Proteinuria was measured using the Bradford method according to a previously described protocol [24]. On the following day, mice were killed under chloral hydrate anesthesia and the kidneys were removed and fixed in 4% paraformaldehyde and then embedded in paraffin. Histological sections from the mice kidneys were stained with periodic acid-Schiff stain (PAS).
Statistical analysis
Each experiment was performed at least thrice. Data are presented as mean ± SD. Statistical analysis was tested by two-tailed Student's t-test using SPSS statistics 17.0. The value of P < 0.05 was considered statistically significant.
Results
Activin A and high RA efficiently induced hAD-MSCs to differentiate into IM
To establish a platform for the differentiation of IM from hAD-MSCs, we examined the effects of the addition of 10 ng/mL activin A with different concentrations of RA (0, 0.1, 1, 10 μM), as the factors that have previously been shown to be critical for the early stages of kidney development [25 –28]. Adding 10 μM RA in combination with activin A dramatically increased the expression of IM genes (Osr1, Wt1, Pax2, Six2) [29] without significant upregulation of endoderm-related genes (Foxa2 and Sox17) [30] and ectoderm-related genes (Pax6 and Sox1) [31] on day 3 (Fig. 1A). Furthermore, RA treatment resulted in the transcriptional activation of Osr1 gene in a dose-dependent manner (Fig. 1A), and Osr1 has been proved to be a key factor for renal lineage differentiation of pluripotent cells, as well as a specific markers of IM [28]. This finding suggested that 10 μM RA might be an optimal concentration during this induction process. To further confirm this hypothesis, immunofluoresence and flow cytometry analysis were performed. Results showed that the induced cells expressed Osr1 with 50%–60% efficiency on day 3 by 10 ng/mL activin A and 10 μM RA treatment (Fig. 1B, C). Taken together, our observations indicated that the combination of 10 ng/mL activin A and 10 μM RA was optimal for induction of IM from hAD-MSCs.
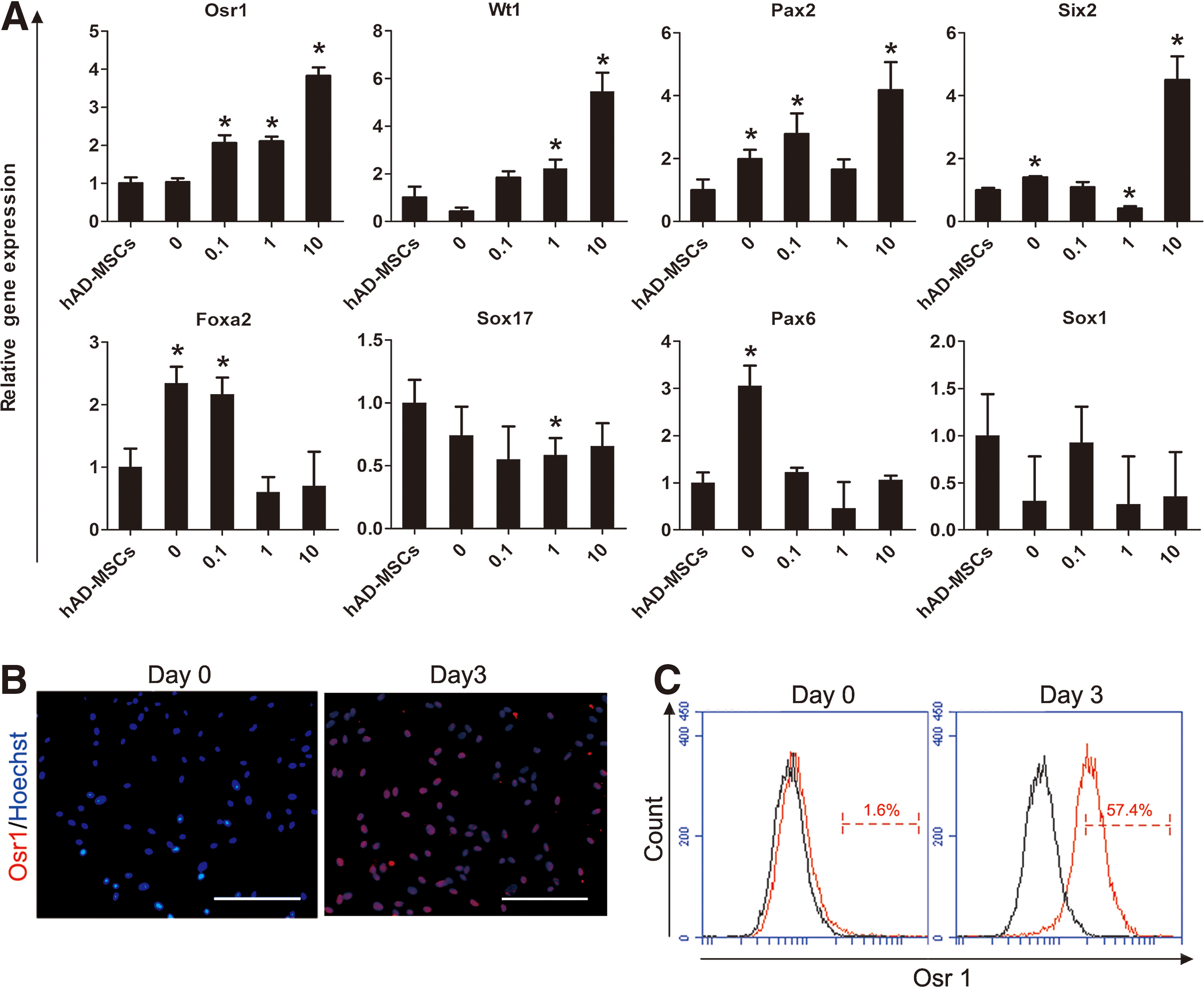
High concentration of RA efficiently induced hAD-MSCs to differentiate into intermediate mesoderm-like cells together with activin A.
Establishment of induction method of kidney podocyte-like cell generation from hAD-MSCs
Although we did not see that low-dose RA in combination with activin A has a significant increase in the Osr1 expression, we observed that the addition of 0.1 μM RA to 10 ng/mL activin A resulted in a marked increase in the expression of some other renal lineage genes (data not shown). To further establish a protocol for the differentiation of podocyte-like (iPod) cells from hAD-MSC-derived IM, we tested the effects of low concentration RA and activin A, together with three different growth factors: BMP7, GDNF and Wnt4, which were evaluated with the addition of the nephrogenic factors previously reported to enhance the expression of renal markers [7,8] (Fig. 2A). We examined the expressions of podocyte-specific maker genes (Nephrin, Podocin, Podocalyxin, Synaptopodin) [32] and transcription factors (Wt1 and Pax2), which have been identified in developing and mature podocytes of each culture group using qRT-PCR analysis. By 9 days of culture, podocyte-specific genes Nephrin and Podocin showed strongly upregulated by BMP7 treatment, much higher than the addition of either GDNF or Wnt4 (Fig. 2B). As expected, the BMP7 could also enhance the expression of Podocalyxin, Synaptopodin, Wt1, and Pax2 (Fig. 2B). These data suggested that the subsequent addition of BMP7 to 10 ng/mL activin A and 0.1 μM RA treatment in Stage 2 was the most potent inducer of Nephrin and Podocin expression. Thus, we chose BMP7 at stage 2 for podocyte generation from hAD-MSCs.
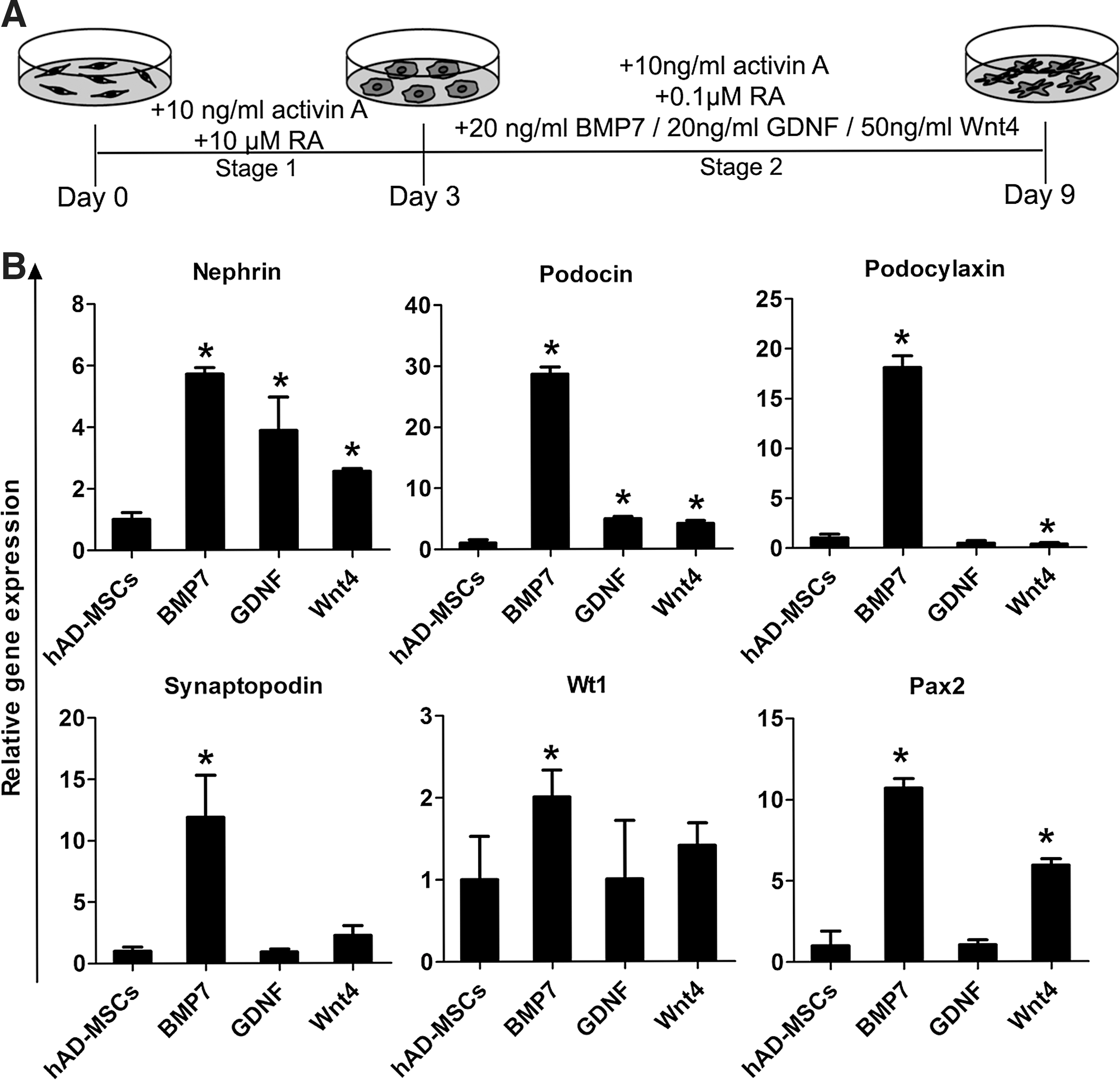
Establishing a method for differentiating hAD-MSCs into kidney podocyte-like cells.
Characterization of iPod cells
Cells differentiated from hAD-MSCs showed a significant morphological change from fibroblast-like cells to flat and round morphology after 3 days of culture (Fig. 3A). By day 9, the cells differentiated into large and arborized cells with processes (Fig. 3A) similar to those previously described for cultured glomerular podocytes [33]. To further characterize iPod cell generation from hAD-MSCs, podocyte-specific makers were examined by western blot and immunofluorescence. We observed that the protein expression of nephrogenic transcription factor WT1 was upregulated on day 3 and persisted to day 9, while podocyte-specific protein (Nephrin, Podocin, Synaptopodin) gradually increased as time extended and achieved a peak on day 9 (Fig. 3B). Immunofluorescence of iPod cells showed nuclear coexpression of WT1 and Pax2 (Fig. 3C), furthermore, the podocyte-specific protein, Nephrin and Podocin, distributed in cytoplasmic and cell surface (Fig. 3D), as would be expected; Synaptopodin was strongly expressed particularly in the cytoplasm and extending into cell processes by 9 days of differentiation (Fig. 3E). These findings confirmed that our differentiation protocol could successfully generate kidney podocyte-like cells from hAD-MSCs and indicated the nephrogenic potential of hAD-MSCs.
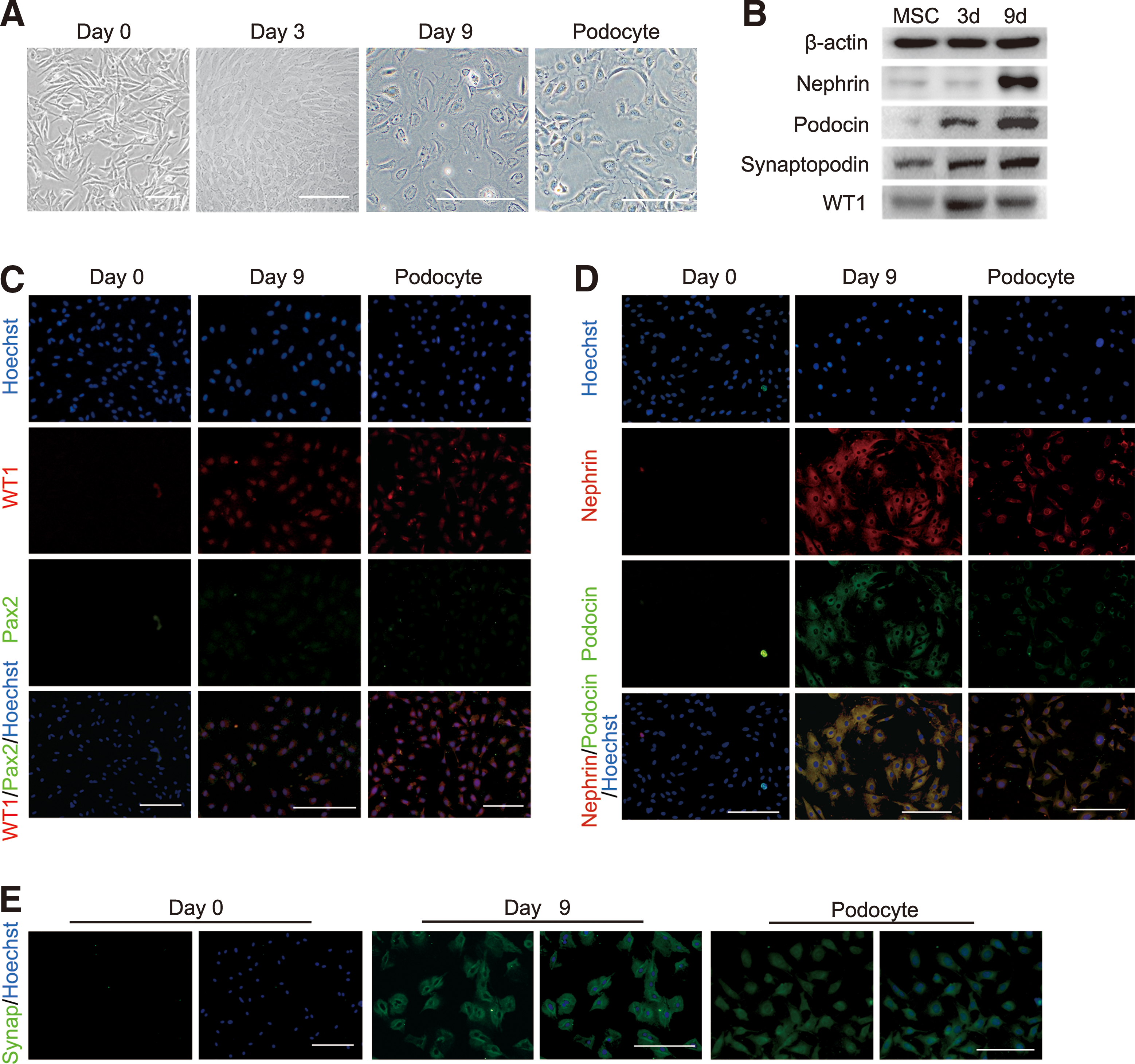
Characterization of iPod cells.
iPod cells have a potential to integrate into developing chimeric kidney cultures and ameliorate proteinuria in adriamycin-treated mice
To investigate the integration capacity of iPod cells, we subjected them to an in vitro reaggregation assay, previously described by Jamie A. Davies et al. [23]. Adapting this technique, E12.5 mouse embryonic kidneys were dissociated to single cells and then reaggregated with iPod cells or undifferentiated hAD-MSCs and grown as organoids at an air–medium interface (Fig. 4A). Numerous glomerulus-like structures were observed in reaggregations, including iPod cells, by histological analysis (Fig. 4B). Immunofluorescence staining showed that these structures contain clusters of cells that expressed WT1, and some laminin-bounded structures were also formed in the reaggregations (Fig. 4C). iPod cells derived from hAD-MSCs were identified using an antibody against HNA. And HNA-positive cells were viable and located in the region of glomerulus-like structures where they were shown to express podocyte protein, Nephrin (Fig. 4C). Furthermore, these structures contained clusters of cells that coexpressed typical podocyte markers (Nephrin and Podocin) (Fig. 4C). Such an integration did not occur in undifferentiated hAD-MSC reaggregations (Fig. 4B).
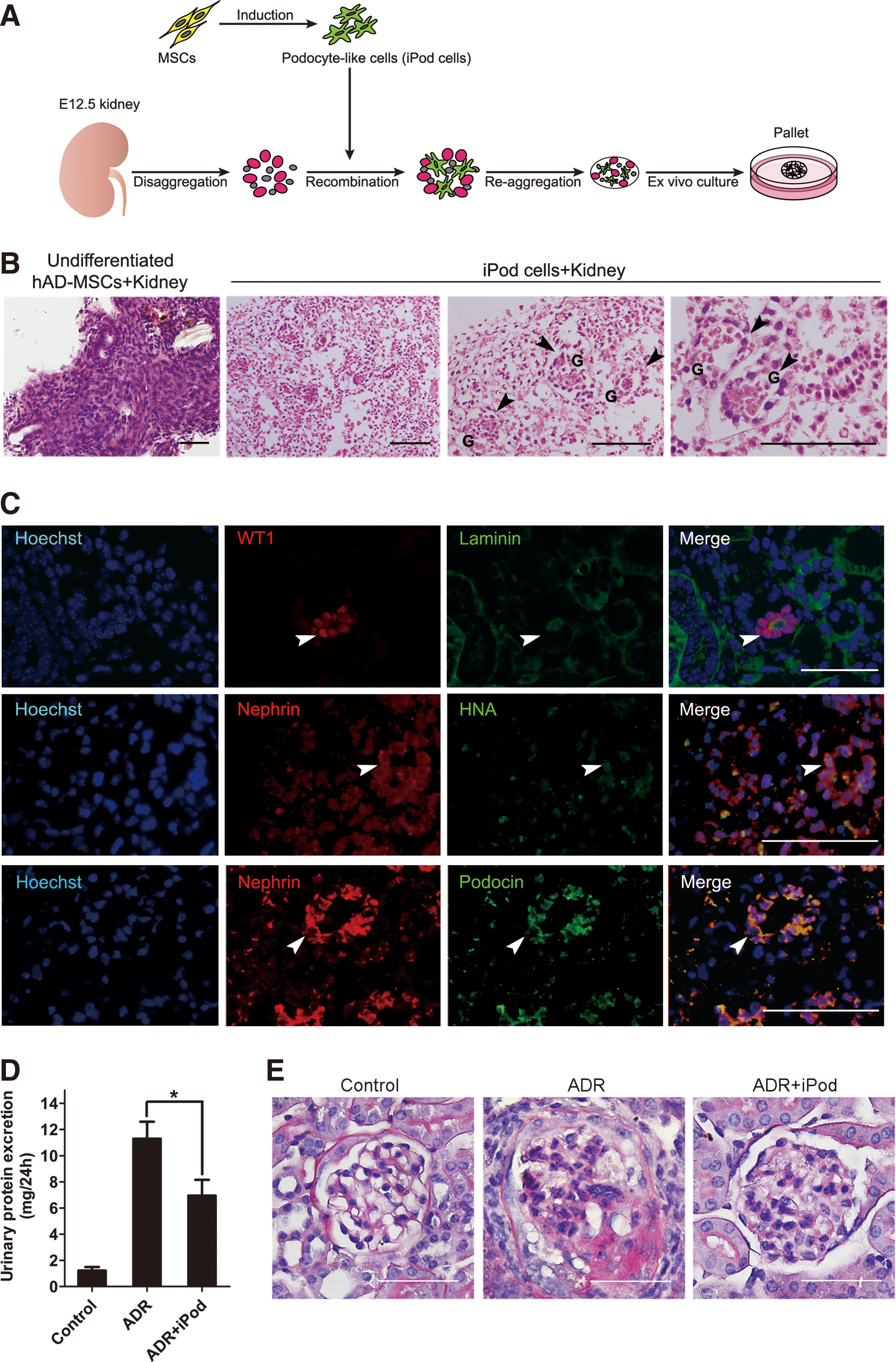
The integration of iPod cells into reaggregates of mouse embryonic kidney cells.
Next, to evaluate the effects of iPod cells in vivo, iPod cells were injected to ADR mice 3 days after drug injection. As shown in Figure 4D and E, adriamycin injection caused a significant proteinuria and histological signs of glomerular injure, and even glomerulosclerosis was observed. Also, treatment of iPod cells significantly reduced urinary protein excretion and attenuated glomerular lesions in ADR mice. Therefore, we concluded that iPod cells, differentiated from hAD-MSCs, were able to integrate into developing kidney tissues and ameliorate proteinuria and kidney injure in ADR mice.
Inhibition of endogenous miR-498 improves the differentiation of hAD-MSCs into podocyte-like cells
MicroRNAs play an important role in a wide range of biological processes and also have been demonstrated to be critical for the development of podocytes [34 –37]. Since Wt1 is an essential kidney development gene, it plays a crucial role in nephron development and podocyte differentiation [13]. To identify miRNAs that specifically regulated the podocyte differentiation, we predicted miRNAs that target Wt1 by silico analyses using TargetScan and PicTar algorithms and selected miRNA-498 as a candidate. To determine the role of miRNA-498 during podocyte differentiation of hAD-MSCs, RNA samples were harvested on days 0, 3, 6, and 9. miRNA-specific qRT-PCR analysis showed that the expression levels of miR-498 were significantly downregulated during this process (Fig. 5A). According to these results, we then speculated that miR-498 might be a negative regulator of podocyte generation.
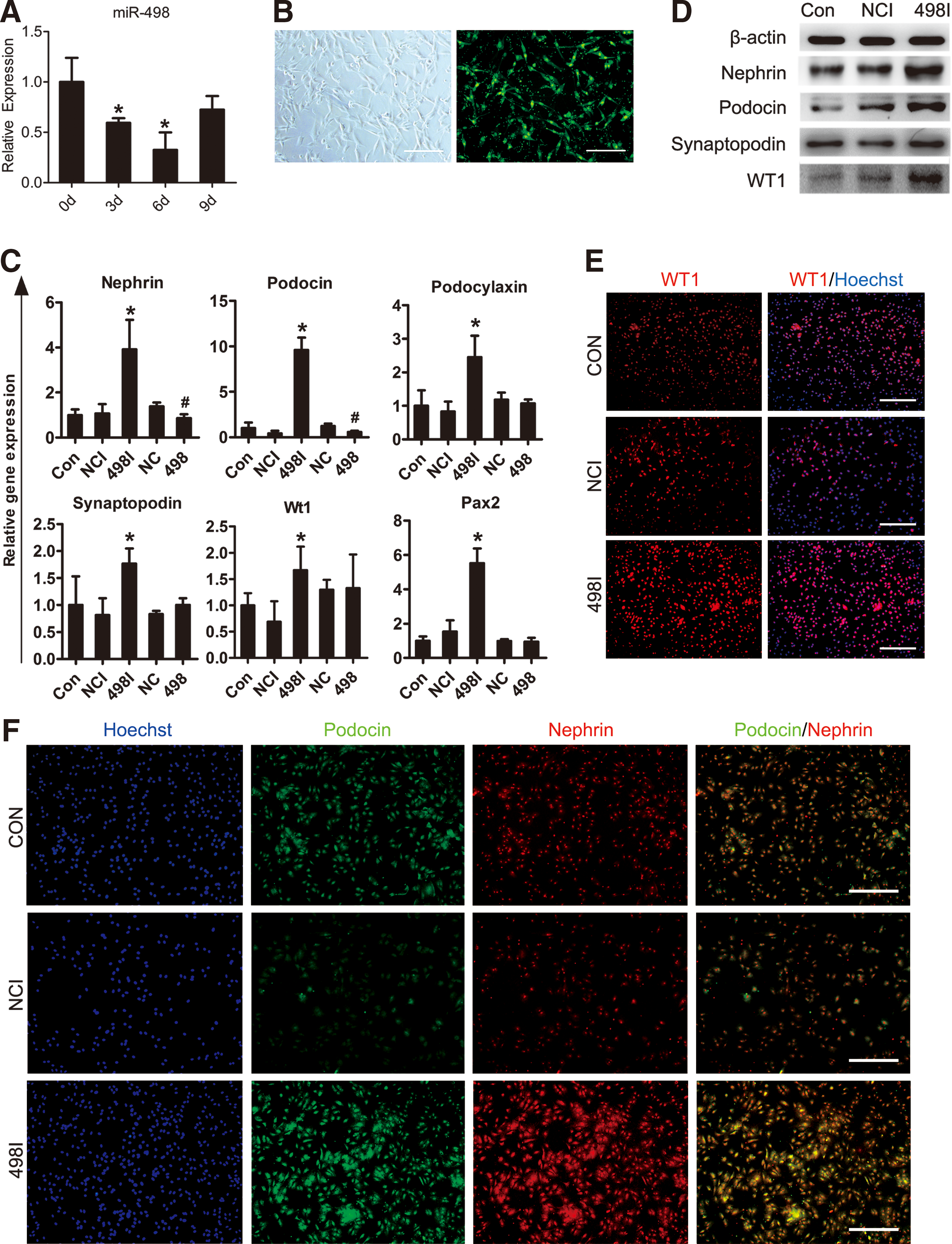
Inhibition of endogenous miR-498 in hAD-MSCs promote differentiation of hAD-MSCs into podocyte-like cells.
To further examine the effect of miR-498, miR-498 mimics (miR-498) or specific inhibitor (miR-498I) and the respective negative control (miR-NC or miR-NCI) were transfected into hAD-MSCs, and hAD-MSCs were then to differentiate toward kidney podocyte-like cells by our two-stage protocol. A high transfection efficiency (80%–90%) was confirmed by a fluorescent microscope (Fig. 5B). Treatment of hAD-MSCs by miR-498 inhibitor significantly increased the expression of marker genes of podocytes (Nephrin, Podocin, Podocalyxin, and Synaptopodin) and transcription factors (Wt1 and Pax2) compared with the control group (iPod cells without transfection) and the miR-NCI-treated group (Fig. 5C), whereas, overexpression of miR-498 significantly downregulated the expression of Nephrin and Podocin, but had no effect on Wt1 at the mRNA level (Fig. 5C).
We also observed that the transfection of miR-498 inhibtor strongly upregulated the protein expression of WT1 and podocyte-specific markers than the negative control after induction (Fig. 5D). Immunostaining also showed a higher expression for WT1 in the nuclei and coexpression of typical podocyte markers Nephrin and Podocin in miR-498I treated group (Fig. 5E, F). Furthermore, the mean number of iPod cells obtained from miR-498-inhibited (3.81 ±0.18 × 105, n = 6) hAD-MSCs per well (six-well plates) on day 9, were much higher than the control group (2.15 ± 0.30 × 105, n = 6) and the miR-NCI-treated group (1.66 ± 0.15 × 105, n = 6) (Fig. 6A).
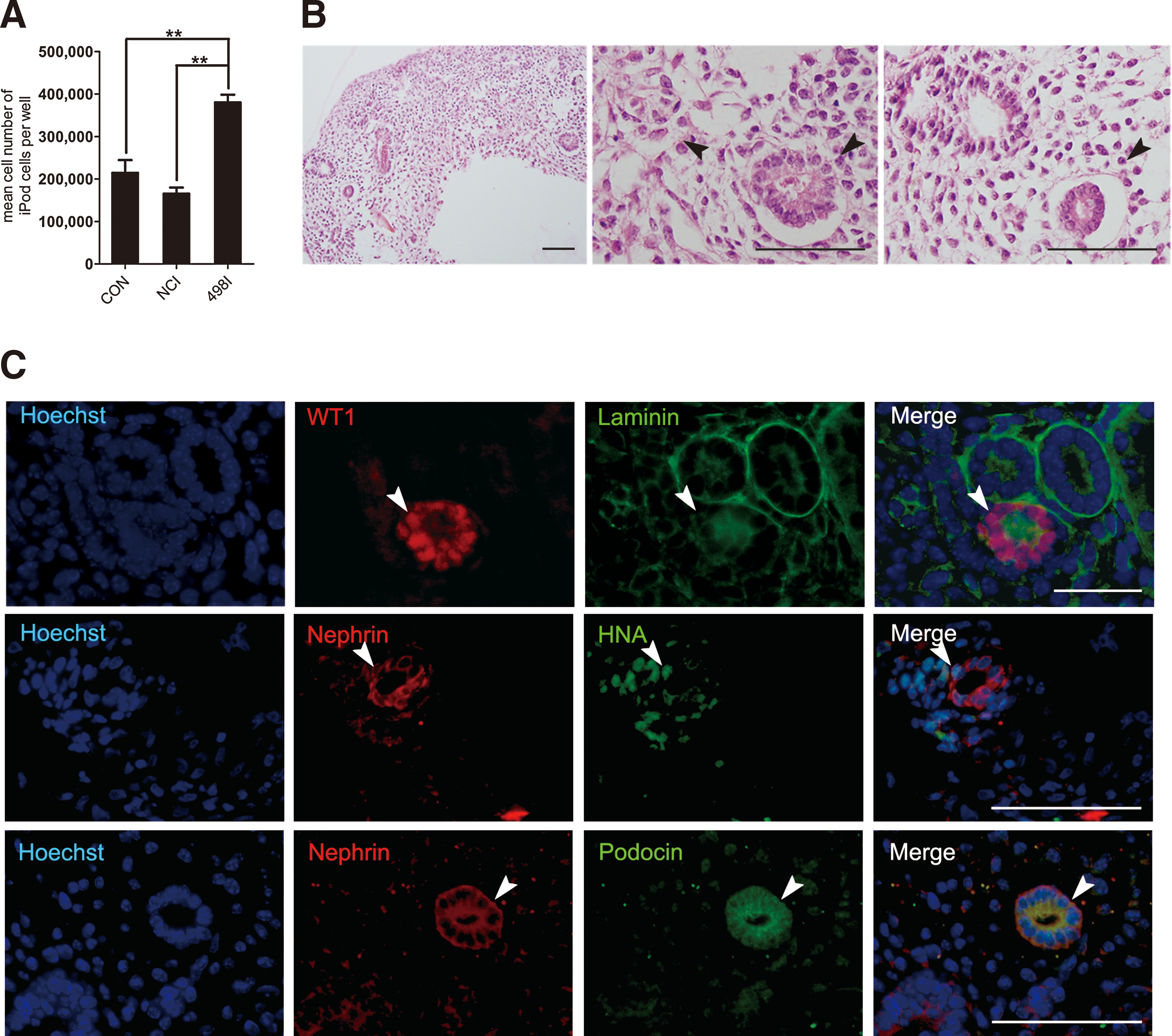
iPod cells differentiated from miR-498 inhibitor-transfected hAD-MSCs retained the integration ability.
To further confirm the integration capacity of iPod cells differentiated from hAD-MSCs transfected with miR-498I, cells were reaggregated with embryonic kidneys. WT1-positive developing nephron and laminin-bounded structures were observed in reaggregations. Also, HNA-positive iPod cells were detected in the glomerulus-like structures and shown to express podocyte-specific protein, Nephrin and Podocin (Fig. 6B, C). We therefore concluded that the inhibition of miR-498 could enhance the generation efficiency of podocyte-like cells from hAD-MSCs, and these cells retained the potential to integrate into developing kidneys.
Discussion
Although the differentiation of MSCs into the hepatic, pancreatic, and neural lineage cells has been widely reported, few studies have attempted to derive cells of the kidney lineage from hAD-MSCs, especially podocytes [38 –40]. In our present study, we initially established a condition that induced hAD-MSCs toward Osr1+ cell population to generate IM-like cells. Subsequently, cells were inducted into podocyte-like cells expressing multiple specific markers of podocytes, such as WT1, Nephrin, and Podocin. To our knowledge, this is the first report of the generation of podocytes from hAD-MSCs that can integrate into the developing nephron of embryonic kidney explant cultures.
Glomerular visceral epithelial cells, also called podocytes, like other kidney lineage cells are derived from the IM [41]. So as the first stage, we tried to induce hAD-MSCs toward IM as the first step toward the induction of podocytes. As one of the earliest markers specific for IM, Osr1 is activated shortly after the migration from the primitive streak and play an important role in both nephron and collecting duct formation during the kidney development [42]. Osr1 mutation may lead to reduction of Wt1 and deficiency of Pax2 and Eya1 in the metanephric region, resulting in failure to form a metanephric mesenchyme (MM) and renal agenesis [43]. Thus, it can be seen that Osr1 acts upstream of Wt1, Eya1, and Pax2 in the nephrogenic process to promote MM formation and survival. Therefore, we selected Osr1 as a specific marker of IM to define the induction of hAD-MSCs.
Previous study showed that both activin A and RA have an effect on the induction of kidney lineage cells from pluripotent cells [44,45]. In our study, treatments with activin A alone did not upregulate Osr1, instead the transcription of other lineage marker genes such as Foxa2 (endoderm) and Pax6 (ectoderm) are increased, which means that activin A itself could not induct hAD-MSCs tending to kidney lineage. While fortunately, we have succeeded in generation of the Osr1+ cells from hAD-MSCs by adding RA in the culture, and in this condition, renal-relative genes were upregulated and genes of other lineages remain unchanged. RA is secreted from the paraxial mesoderm and acts as an important signaling factor for the specification of primitive streak into IM [27,46], and it is also necessary for the nephric duct formation and initial specification of renal progenitor cells [47,48]. The effects of different concentrations of RA were studied, and we found that a combination of activin A and 10 μM RA achieved the highest efficiency of IM differentiation on day 3. The necessity for high concentration of the RA demonstrated that RA is crucial for the induction of IM in the hAD-MSC differentiation model.
During the kidney development, the ureteric bud (UB) and MM derived from IM [49,50]. The metanephric mesenchymal cells differentiate through MET into various types of epithelial cells, including visceral glomerular epithelial cells, podocytes [13]. Numerous transcription factors and signal molecules have been identified to govern these processes. In the following step toward the podocytes, attenuation of the RA concentration and stage-specific addition of growth factors allowed the nephric lineage commitment and podocyte formation. So, we tested the effects of three different growth factors: GDNF, Wnt4, and BMP7. The GDNF is secreted by the MM and is necessary for UB outgrowth and subsequent UB branching morphogenesis through the GDNF/Ret signaling pathway [51 –53]. Wnt4 expression in the MM and Wnt signal is essential for nephron progenitors to undergo a MET and the early phases of nephrogensis [54 –56]. BMP7 is highly expressed in both the UB and cap mesenchyme, which could maintain the survival of metanephric mesenchymal cells. Recent studies have identified that BMP7 also plays an important role in nephrogenesis [57,58]. In our study, compared to GDNF and WNT4, addition of BMP7 to activin A and 0.1 μM RA treatment was most effective for podocyte induction, which suggested that BMP7 signaling is a potent inducer for hAD-MSC differentiation toward podocyte-like cells.
iPod cells differentiated from hAD-MSCs resembled the appearance of glomerular podocytes and coexpressed podocyte-specific makers such as Nephrin and Podocin. And these cells also showed a capacity to integrate into nephric glomerulus-like structure in embryonic kidney explants and ameliorate proteinuria in kidney damage animal model. Unlike benchwork, the systems that can be directly used in clinical applications for regenerative medical often require to get abundant proper cells for transplantation in safety and efficacy. However, an optimizing strategy for differentiation remains a concern. Most efficient methods to generate specific cell types reported so far involve vector-mediated transduction of genetic materials and thus run high risk and costs. Therefore, besides regulatory proteins, we considered employing some novel small molecules to establish a virus-free method for generating podocytes efficiently.
miRNAs are noncoding RNA molecules, regulatory RNAs that act as posttranscriptional repressors. The previous report revealed that specific deletion of dicer in podocytes led to dysplasia in early developing glomerulus and indicated that miRNAs might be essential for the podocyte differentiation [36]. In recent years, researchers have found that several miRNAs involve in the kidney development [59 –61]. It might be a safe and effective approach to increase efficiency of specific cell generation by reducing the miRNAs targeting key differentiation-related genes.
As we know, WT1 is an essential transcription factor with functions in the nephron development, plays a crucial role in the regulation of the process of MET, maturation of podocytes, as well as nephron progenitors survival [62]. So we predicted miRNAs that might modulate podocyte differentiation of hAD-MSCs by targeting WT1 by silico analyses. Fortunately, we found that treatment of hAD-MSCs by miR-498 inhibitor significantly increased the expression of WT1 protein and improved the differentiation of hAD-MSCs into iPod cells. However, to our surprise, we did not find any evidence that supports WT1 as the direct target of miRNA-498 through the detection of a relative luciferase activity (data not shown).
Considering that many transcription factors and signal molecules could regulate the expression of WT1, we then screened the potential target of miRNA-498 with the bioinformatics software again, and found miRNA-498 might target several crucial genes related to the kidney development, including Nephrin and Pax2, etc, while luciferase assay did not show positive results. A tentative inference was that miR-498 probably regulated the expression of WT1 through an indirect effect. We were really fortunate to discover that downregulated miR-498 could get a larger number of iPod cells and improve the differentiation of hAD-MSCs into podocyte-like cells.
In conclusion, we have successfully established a differentiation protocol to generate podocyte-like cells from hAD-MSCs. This induction system, especially the optimizing solution with miRNA inhibitor may advance the development of kidney regenerative medicine and provide an alternative therapy for kidney disorders, including ESKD and kidney failure. On the other hand, this finding may also offer an opportunity to generate human cells that better represent kidney disorders for related researches and drug discoveries.
Footnotes
Acknowledgments
This study was supported by grants from the National Key Scientific Program of China (no. 2011CB964901), the National Collaborative Innovation Program, and the National Science and Technology Major Projects for “Drug Research and Development” (no. 2014ZX09101042).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.