
Abstract
Mesenchymal stem cells (MSCs), which can be obtained from various organs and easily propagated in vitro, are one of the most extensively used types of stem cells and have been shown to be efficacious in a broad set of diseases. The unique and highly desirable properties of MSCs include high migratory capacities toward injured areas, immunomodulatory features, and the natural ability to differentiate into connective tissue phenotypes. These phenotypes include bone and cartilage, and these properties predispose MSCs to be therapeutically useful. In addition, MSCs elicit their therapeutic effects by paracrine actions, in which the metabolism of target tissues is modulated. Genetic engineering methods can greatly amplify these properties and broaden the therapeutic capabilities of MSCs, including transdifferentiation toward diverse cell lineages. However, cell engineering can also affect safety and increase the cost of therapy based on MSCs; thus, the advantages and disadvantages of these procedures should be discussed. In this review, the latest applications of genetic engineering methods for MSCs with regenerative medicine purposes are presented.
Introduction
S
The simplicity of obtaining these cells from different organs and the subsequent ease of propagation in vitro [4], as well as the potential for autologous transplantation, make them a tempting possibility for clinical use. However, special emphasis must be placed on the correct characterization of the obtained cells from different sources and species to be able to properly compare and make right conclusions on the grounds of multiple studies [5,6].
Independently of their original tissue location, MSCs have a natural ability to differentiate into mature mesenchymal phenotypes and form bone or cartilage, so they can be used in the treatment of injuries where these types of tissues are in need of repair [7,8]. What is more, MSCs act through paracrine effects, releasing a plethora of beneficial compounds [9]. While Prochymal (Mesoblast Ltd.), a preparation of allogeneic MSCs, has recently received conditional approval from the FDA, the engineered MSCs did not reach the level of clinical trials.
However, genetically engineered neural stem cells (NSCs) are being tested in a clinical trial sponsored by ReNeuron (NCT01151124 and NCT02117635, Clinicaltrials.gov). However, the diverse biological improvements offered by genetic engineering have the potential to greatly increase the therapeutically useful qualities of MSCs and tailor them to specific diseases, so they are more probable to be used clinically in the future. A variety of studies have investigated the engineering of MSCs, aiming at stimulation of their direct differentiation toward endothelial cells and angiogenesis [10 –16]. In addition, in in vitro (Fig. 1) culture conditions and in vivo studies (Fig. 2), MSCs can be differentiated into lineage-specific cells or lineage-specific-like cells, such as hepatocytes [17], cardiomyocytes [18], pacemaking cells [19], and neuronal cells [20]. Genetically modified MSCs can also be used to ameliorate several neurological disorders by exploiting their paracrine characteristics [21,22]. Moreover, MSCs could be modified to gain antifibrotic properties [23].

Differentiation potential of engineered mesenchymal stem cells (MSCs) in vitro. Although MSCs have the natural ability to differentiate into osteocytes and chondrocytes, genetic engineering methods may intensify this natural behavior of MSCs. In addition, genetic engineering techniques have been used in MSC differentiation in vitro into many cell types, such as hepatocytes, cardiomyocytes, pacemaking cells, endothelial cells, pancreatic cells, and neurons. Color images available online at
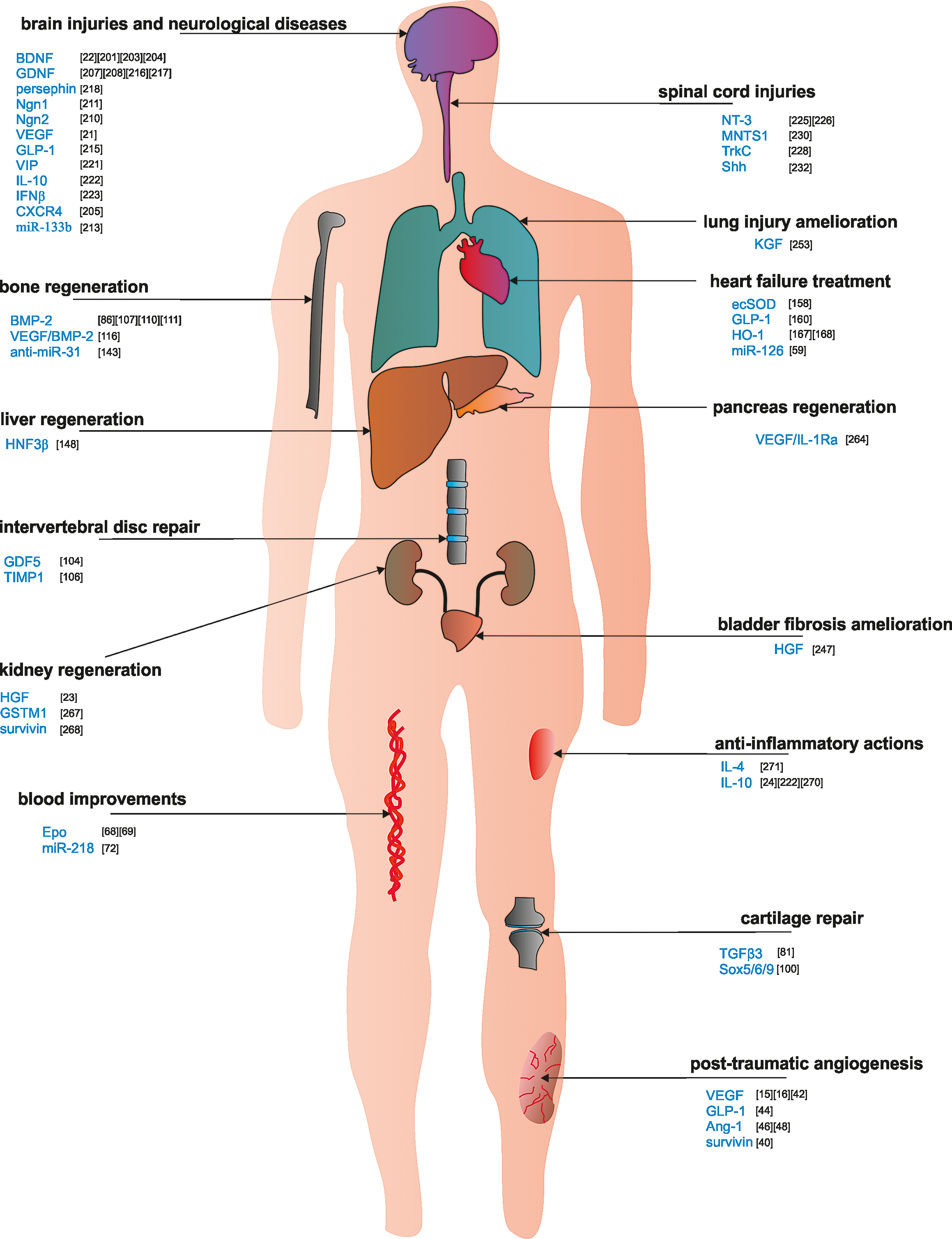
Potential therapeutic applications of engineered MSCs in vivo. The genetic engineering of MSCs converts MSCs into a multifunctional therapeutic tool that may be used in numerous applications in vivo, such as in different organ repair, antifibrotic actions, and anti-inflammatory purposes. Color images available online at
Finally, MSCs can be committed to overproduce anti-inflammatory cytokines that alleviate local tissue inflammatory states [24]. The abundance of genetic engineering modifications in MSCs quoted in this review is summarized in Table 1.
ACE2, angiotensin-converting enzyme 2; Ang-1, angiopoietin-1; BDNF, brain-derived neurotrophic factor; bFGF, basic fibroblast growth factor; BMP-2, bone morphogenetic protein 2; Cbfa-1, core binding factor alpha l; CDMP1, cartilage-derived morphogenetic protein 1; ecSOD, extracellular superoxide dismutase; Epo, erythropoietin; Foxc2, forkhead box protein C2; GDNF, glial cell line-derived neurotrophic factor; GLP-1, glucagon-like peptide-1; GM-CSF, granulocyte-macrophage colony-stimulating factor; GSTM1, glutathione S-transferase M1; HGF, hepatocyte growth factor; HO-1, heme oxygenase-1; HNF4α, hepatocyte nuclear factor 4α; IGF-1, insulin-like growth factor-1; Mash1, mammalian achaete-scute homolog-1; NGF, nerve growth factor; Ngn2, neurogenin2; NT-3, neurotrophin-3; OGP, osteogenic growth peptide; PDGF, platelet-derived growth factor; RAR-β, retinoic acid receptor β; SCD-1, stearoyl-CoA desaturase 1; Shh, sonic hedgehog; TAZ, transcriptional coactivator with a PDZ-binding motif; TIMP-3, tissue inhibitor of matrix metalloproteinase-3; VEGF, vascular endothelial growth factor; VIP, vasoactive intestinal peptide.
This review focuses on a set of examples of the various genetic modifications of MSCs to enhance and/or expand MSC differentiation potential toward multiple cell types for therapeutic purposes, which have been published in recent years, revealing the growing interest directed at these cells and the diversity of strategies (Fig. 3).
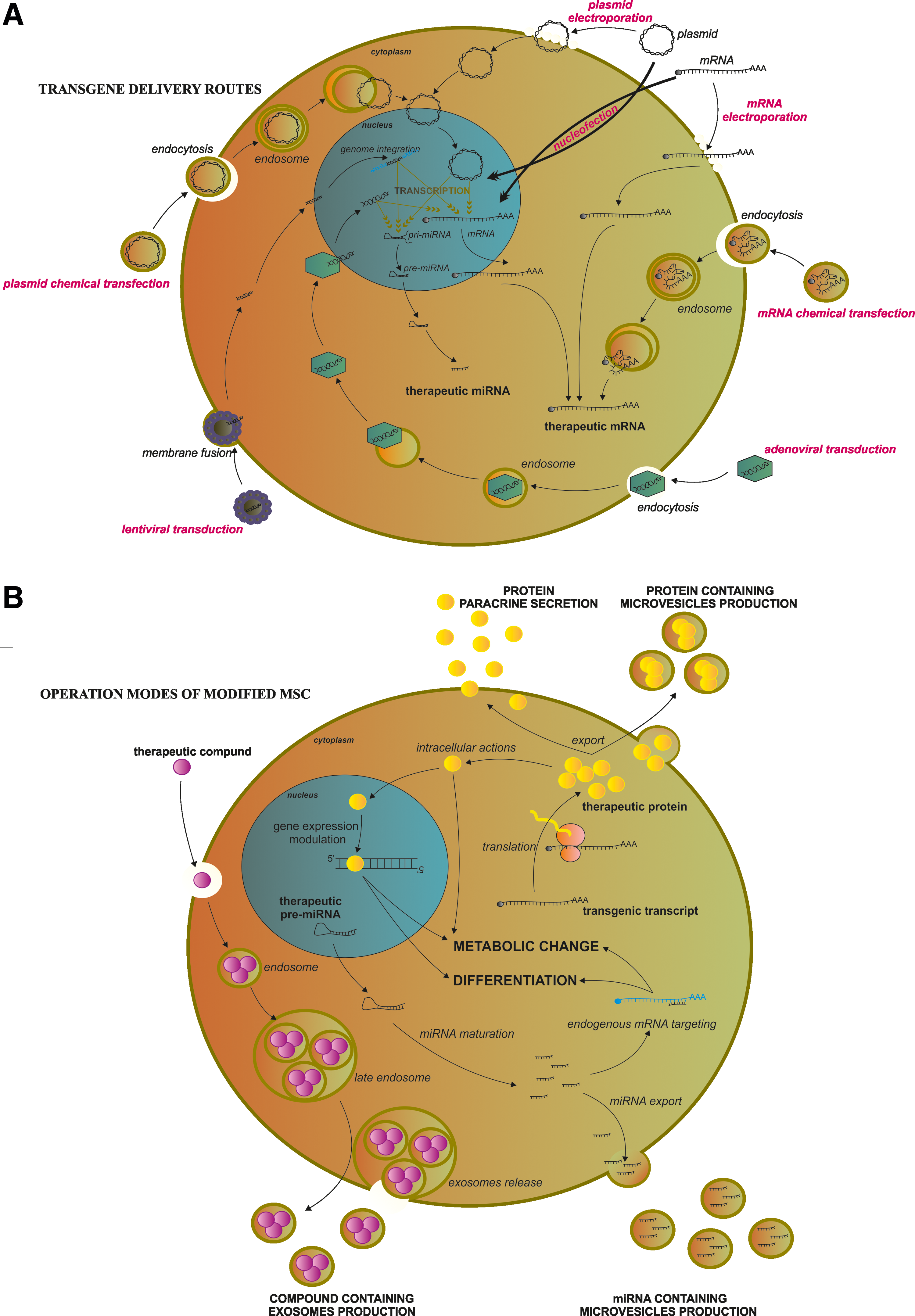
Diversity of transgene delivery routes and operation modes of modified MSCs with therapeutic purposes. (
Vectors, Methods, and Approaches to MSC Engineering
There are various vectors used for delivery of genetic materials to the cells, such as viruses [25], DNA plasmids [26,27], constructs of minimalistic, immunologically defined gene expression [28], and transposon constructs such as the Sleeping Beauty System [29], mRNA [30], miRNA [31], and siRNA [32]. While viruses directly transduce cells, nonviral methods require specific methods to cross the cell membrane, such as lipofection or electroporation. The vectors and methods for crossing the cell membrane by genetic material have been recently extensively reviewed [33].
From the biological point of view, there are also various approaches through which genetically engineered modification could be accomplished. The most basic and widespread approaches are related to the overexpression or knockdown of genes that encode extracellular exported proteins [34], specific membrane anchored [35], or cytoplasmic proteins [36]. The more powerful approach is to induce the expression of transcription factor genes, which allows for cell reprogramming and, in turn, leads to a complete change of phenotype [37]. Recently, the use of miRNAs is garnering growing interest, and permits the control of entire intracellular regulatory pathways, due to the relationship of one miRNA with several genes [38].
Angiogenesis Stimulation Approaches
To start with, one of the applications for MSCs in stem cell therapies is the field of induced post-traumatic angiogenesis. The problem of new blood vessel formation after injury is particularly important in ischemia, where the original arrangement of the blood vessels is destroyed, leading to a lack of oxygen and nutrient supply to the affected area. The process of angiogenesis naturally occurs after injury, but there is still a therapeutic interest in the enhancement of this physiological process. The MSC-based cell therapy may address some of these issues.
One of the most popular approaches in this field is an overexpression of membrane protein: vascular endothelial growth factor (VEGF). VEGF properties have already been reviewed, revealing the important role of VEGF in angiogenesis during tissue injuries, such as in myocardial infarction, stroke, ischemia, and wound healing [39].
There are several examples of studies related to this kind of MSC modification, confirming that VEGF-engineered bone marrow-derived MSCs (BM-MSCs) possessed improved capabilities for tissue repair through the increased rate of angiogenesis [11 –13,15,16]. An increased capillary density, a reduction in the infarct size, and elevated VEGF expression could be reached also by the use of survivin-producing MSCs [40]. Survivin was found to stimulate VEGF gene expression by indirectly upregulating its promoter [41].
On the other hand, the VEGF-modified umbilical cord-derived MSCs (UC-MSCs) found an application in the tissue-engineered dermis for the treatment of skin defect wounds. In that case, the acellular dermal matrix scaffolds were used as platforms for the microencapsulated VEGF gene-transduced UC-MSCs, which effectively improved the vascularization of the graft and participated in the improved quality of wound healing [42].
The VEGF engineering can be accompanied by cotransfections with another factor that could enhance the proangiogenic effect of the VEGF-engineered BM-MSCs. The dual transfection of the TERT and VEGF genes was found to bring prolonged, beneficial proangiogenic effects in BM-MSCs derived from aged donors. An extended life span of the exogenous cells and improved angiogenic capacity were observed in those cells [43].
In another example of encapsulated BM-MSCs with angiogenic purposes, in a mouse hind limb ischemia model, BM-MSCs were modified to overexpress the glucagon-like peptide-1 (GLP-1) gene, which is known as an antiapoptotic, proangiogenic, and cardioprotective factor. Perivascular administration of the GLP-1-producing BM-MSCs improved postischemic reperfusion [44].
Angiopoietin-1 (Ang-1) is a potent growth factor that was found to be essential for new vein formation [45]. Based on this characteristic, Ang-1-engineered BM-MSCs were proposed for studies on angiogenesis. In a rat ischemic limb model, the Ang-1-modified BM-MSCs contributed to improved blood flow recovery as well as to an increase of capillary density in the ischemic limb [46]. The cotransfection of the Ang-1 gene with the bone morphogenetic protein 2 (BMP-2) gene into BM-MSCs was found to significantly improve osteogenic and angiogenic differentiation [47]. Furthermore, the Ang-1-expressing UC-MSCs were delivered intravenously to a rat model of severe acute pancreatitis where these cells were involved in the promotion of pancreatic angiogenesis by reducing pancreatic injury and serum levels of proinflammatory cytokines [48].
The enhanced angiogenic potential of genetically engineered BM-MSCs was found for several additional modifications, such as for granulocyte-macrophage colony-stimulating factor [49]; hepatocyte growth factor (HGF) [50]; human basic fibroblast growth factor [51], a tissue inhibitor of matrix metalloproteinase-3 (TIMP-3) [52]; the Notch homolog 1 [53]; heme oxygenase-1 (HO-1) [54], a member of the family of zinc finger transcription factors; GATA-4 [55]; HIF-1α [56]; and the LIM-homeobox transcription factor islet-1 (Islet-1) [57].
Some studies exist that relate to proangiogenic miRNA and engineered MSCs. miR-126 was featured in neoangiogenesis after myocardial infarction and with the posterior maintenance of vascular integrity [58]. In a mouse model of myocardial infarction, the transplantation of BM-MSCs expressing miR-126 resulted in increased angiogenesis and in the recovery of cardiac function in the infarcted zones of hearts [59,60].
Genetic engineering of MSCs could be applied as well in the field of endothelial cell differentiation. For instance, angiotensin-converting enzyme 2 (ACE2) overexpression in BM-MSCs contributed to the prevention of endothelium-mediated inflammation and improved endothelial repair in in vitro conditions [61]. Physiologically, ACE2 is involved in the degradation of angiotensin II, which, in turn, is involved in vascular injury by contributing to the generation of proinflammatory, prothrombotic, and profibrotic reactive oxygen species [62]. For this reason, ACE2, as a physiological antagonist of angiotensin II, is well suited as an anti-inflammatory factor that protects endothelial cells [63].
In addition to this, the enzymatic activity of stearoyl-CoA desaturase-1 (SCD-1) protects human arterial endothelial cells from lipotoxicity [64], while its overexpression in BM-MSCs was found to stimulate the expression of endothelial lineage marker genes [65].
Another example is the findings on the use of the angiogenesis-related miRNA, miR-126, in BM-MSC differentiation toward endothelial cells [10]. In that case, miR-126-engineered BM-MSCs adopted the typical endothelial cobblestone-like structures in vitro, exhibited the expression of a series of endothelial-specific markers, and increased paracrine abilities.
Engineering of MSCs to Increase Blood Production
Engineered MSCs could have a role in the treatment of hemophilia B since the encapsulated factor IX-engineered umbilical cord blood-derived MSCs (UCB-MSCs) produced sufficient amounts of that therapeutic protein in in vitro three-dimensional cultures [66].
Anemia has been also addressed through the administration of engineered MSCs. There are some examples of erythropoietin (Epo), the main inducer of red blood cell production [67], and its gene overexpression in MSCs to stimulate blood cell formation in host animals after BM-MSC administration. In animals that received either subcutaneously injected [68] or intraperitoneally implanted [69] human Epo-producing BM-MSCs, an increased hemoglobin level was reported. Moreover, HIF-1α-overexpressing BM-MSCs cocultured with hematopoietic stem cells (HSCs) in in vitro conditions were shown to stimulate the growth of the latter cells [70]. A similar phenomenon was observed with the Jagged-1-producing BM-MSCs cocultured with HSCs in vitro [71].
Apart from this, there have been attempts to convert UC-MSCs into hematopoietic cells. For instance, the miR-218 overexpression in UC-MSCs facilitated differentiation toward CD34+ and CD45+ cells if combined with additional stimulation [72].
Cartilage Production Applications
One of the hallmarks of MSCs is their ability to differentiate toward chondrocytes under appropriate in vitro culture conditions, as confirmed for BM-MSCs [73], and the use of scaffolds was reported to enhance this process in UCB-MSCs [74], UC-MSCs, BM-MSCs [75], and dental-derived MSCs [76]. Moreover, multiple studies are underway to increase the chondrogenesis of MSCs by diverse genetic modifications. Previously, it was reported that TGF-β superfamily members were able to induce BM-MSCs to differentiate into chondrocytes under in vitro culture conditions [77].
The TGF-β1 gene could be efficiently delivered to MSCs with low toxicity, converting this biological pathway into a plausible solution for the BM-MSC modifications toward chondrogenic phenotypes [78]. The TGF-β1 gene transfection into BM-MSCs was combined with a calcium alginate gel to create three-dimensional, tissue-engineered artificial cartilage to mimic the cell pattern growth in vivo. Type II collagen synthesis by the TGF-β1-transduced BM-MSCs was detected, indicating that in the experimental conditions, the TGF-β1-modified BM-MSCs gradually differentiated into cartilage [79]. These results are consistent with the results of synovium-derived MSCs transfected with the TGF-β1 gene, where the modified cells were more proliferative and their chondrogenic differentiation potential was increased [80].
Another member of the TGF-β superfamily, TGF-β3, was documented as a forceful inducer of chondrogenesis in engineered BM-MSCs. The transfected BM-MSCs were transplanted into a rat model of osteoarthritis-induced cartilage injury. The administered cells differentiated toward chondrocytes and effectively secreted cartilage-relevant matrix compounds [81]. In in vitro studies, the lentiviral TGF-β3-transfected BM-MSCs were able to differentiate into chondrocytes in silk scaffolds, which was confirmed by the accumulation of collagen type II and cartilage extracellular matrix components [82]. In addition, another member of the TGF-β superfamily, the cartilage-derived morphogenetic protein 1 (CDMP1), was studied as a potential inducer of chondrogenic differentiation for BM-MSCs. The CDMP1-modified BM-MSCs were seeded on scaffolds and used to successfully repair laryngeal cartilage defects [83].
The insulin-like growth factor-1 (IGF-1) is postulated to be another good candidate for the improvement of chondrogenic induction in MSCs since it has been shown to influence MSC chondrogenesis by increasing proliferation and inducing the expression of factors involved in the chondrocyte phenotype [84]. Adipose tissue-derived MSCs (ASCs) were cotransfected with the IGF-1 and BMP-2 genes. The appropriate chondrogenic process was appreciated in these so-transformed cells [85], despite the fact that the overexpression [86] and the treatment of BMP-2 alone are reported to facilitate osteogenesis in MSCs [87]. There is an example of the application of another BMP family member, BMP7, in which the BMP7 gene transfection of BM-MSCs resulted in favorable chondrocyte formation [88].
Interestingly, the IGF-1 overexpression was able to stimulate the expression of the Sox9 gene; however, the expression of osteogenic marker genes was also elevated, suggesting that IGF-1 gene transfections into BM-MSCs with chondrogenic purposes should be considered cautiously [89]. Moreover, the IGF-1 gene transfection, combined with different genes involved in chondrogenesis, resulted in an increase of the chondrogenic differentiation capacities of modified ASCs, especially in the case of IGF-1/fibroblast growth factor-2 double-transfected ASCs [90].
In addition, noncanonical Wnt signaling was reported to be useful for this purpose since Wnt11-modified BM-MSCs had the enhanced expression of chondrogenic genes, such as Sox9, and their chondrogenic differentiation was more profound compared with untransfected cells [91].
Other genetic candidates for the induction of chondrogenesis for MSCs are the Sox trio genes (Sox 5, 6, and 9). They are master regulatory transcription factors known to be engaged in the activation of cartilage-specific genes [92]. The Sox9 gene, the key gene in the early stages of chondrogenesis, has been the most studied in BM-MSCs [93 –95].
In one case, to reinforce the chondrogenesis process, the genetic engineering strategy involved the prochondrogenic stimulation represented by the induction of Sox9 gene overexpression and the simultaneous inhibition of the opposite differentiation route toward osteogenic commitment represented by the application of core binding factor alpha l (Cbfa-1) gene silencing. The Cbfa-1 gene expression was linked to the osteogenic differentiation of BM-MSCs [96]. For this reason, at the same time, BM-MSCs were transfected with the Sox9 gene-containing vector and the small interfering RNA against the Cbfa-1 transcript, both conjugated to poly(ethyleneimine) and coated onto poly(lactide-co-glycolic acid) nanoparticles [94].
In a different study, the Sox9 gene upregulation, and hence increased chondrogenic differentiation of BM-MSCs, was achieved due to zinc finger protein 145 (ZNF145) gene overexpression, which was established as a new upstream regulator of Sox9 gene expression [97]. Indeed, there are some data on the improved results when all three Sox family genes were simultaneously overexpressed in BM-MSCs [98,99] and ASCs [100].
Intervertebral Disc Regeneration
In the area of intervertebral disc regeneration, several modifications were reported to improve BM-MSC differentiation toward intervertebral disc-like cells [101,102]. For instance, the growth and differentiation factor 5 (GDF5) gene engineering was used to obtain GDF5-BM-MSCs. GDF5 stimulated chondrogenic BM-MSC differentiation in vitro [103] and induced the repair of degenerated disc in vivo [104]. The GDF5-producing BM-MSCs differentiated into intervertebral disc-like cells, expressing chondrogenic-specific marker genes, and, when transplanted to the intervertebral disc papain degeneration organ culture model, a partial recovery was detectable [102].
Matrix metalloproteinases (MMPs) are involved in extracellular matrix degradation during intervertebral disc degeneration [105]. For this reason, the tissue inhibitors of metalloproteinases convert into an interesting target for MSC engineering with the TIMP metallopeptidase inhibitor 1 (TIMP1) as an example. In this case, the TIMP1-producing BM-MSCs transplanted into the intervertebral injury site elicited double-positive effects by diminishing the negative effects of the MMP activity and, on the other hand, promoted the synthesis of additional extracellular matrix elements [106].
Worth noting is the case of Bcl-2-overexpressing BM-MSCs that became Sox9-overexpressing cells and were able to accumulate proteoglycans, supporting the notion that antiapoptotic BM-MSC engineering could change these cells toward the cells with a nucleus pulposus-like phenotype in vitro [101].
Bone Formation Attempts Using MSCs
In bone formation approaches based on engineered MSCs, the most popular strategy, with a plethora of examples, is related to BMP-2 gene transfection. BMP-2 is a powerful positive regulator of bone formation in vivo [107] and stimulates osteoblast growth in vitro [108]. The BMP-2-overexpressing BM-MSCs have been used in multiple studies to produce new bone tissues in vitro and in vivo [86,109 –111].
For example, the use of anti-BMP-2 antibodies encapsulated with BM-MSCs to entrap BMP-2 in the vicinity of BM-MSCs enabled the proper induction of osteogenesis [112]. In many cases, the BMP-2 overexpression in MSCs was accompanied by the overexpression of other genes that strengthened the effects of BMP-2. For instance, the SDF-1β overexpression in BM-MSCs potentiated osteogenic differentiation and chemotaxis in in vitro-expanded BMP-2-engineered BM-MSCs [113]. Of interest, the simultaneous proangiogenic and pro-osteogenic strategies applied to BM-MSC engineering were found to substantially improve bone formation potential.
The HIF-1α gene overexpression was reported to have beneficial effects on new bone formation by inducing the proangiogenic genes in the double-HIF-1α/BMP-2-positive BM-MSCs [114]. What is interesting is that the HIF-1α gene overexpression alone enhanced the osteogenic differentiation of BM-MSCs just as well [115]. Similar results were reported in the double Ang-1/BMP-2 and VEGF/BMP-2-overexpressing BM-MSCs [47,116 –118]. However, BM-MSCs engineered to exclusively overexpress the proangiogenic VEGF gene were found to be strong inducers of new bone formation by the promotion of new vessel formation among the scaffolds used as templates [119,120].
Curiously, BM-MSCs were converted into bone tissue after the overexpression of the Epo gene [121]. Epo, apart from its well-known role in hematopoiesis, promoted bone regeneration by stimulation of VEGF and BMP-2 gene expression [122,123]. In a different study, the BMP-2-producing BM-MSCs were seeded together with the endothelial progenitor cells (EPCs) in porous nanocalcium sulfate/alginate scaffolds, and EPCs contributed to the increased new vascular network growth. Meanwhile, the BMP-2-producing BM-MSCs promoted bone regeneration [124].
Interestingly, other BMP family members were reported to effectively contribute to bone formation from the transduced BM-MSCs to overexpress genes, such as BMP-4, -7 [125], and BMP-6, claimed to be even more potent pro-osteogenic factors than BMP-2 in the BMP-2-engineered BM-MSCs [126].
Apart from the BMP-based strategies, other physiologically engaged in osteogenesis and bone regeneration were considered for attempts with BM-MSCs, such as the osteogenic growth peptide (OGP) [127]; the IGF-1 [128,129]; leptin [130]; platelet-derived growth factor [131]; ephrinB2 [132]; the integrin α2 subunit ITGA2 [133]; and NEL-like protein (NELL1) overproduction, reported in ASCs [134].
In addition, the targeting of Wnt signaling proved useful in the bone formation studies on MSCs. BM-MSCs were transformed to either overexpress β-catenin (the pivotal element in the canonical Wnt signaling pathway) or the ROR2 gene (the pivotal molecule in the β-catenin-independent, noncanonical signaling pathway). In both cases, the alkaline phosphatase activity was promoted as well as the expression of the osteogenic-specific genes [135]. In addition, transcription factor Cbfa-1 [136] and a transcriptional coactivator with a PDZ-binding motif (TAZ) were reported to stimulate osteogenesis in BM-MSCs [137].
The forkhead box protein C2 (Foxc2)-engineered MSCs were found to differentiate toward osteoblasts, mediated by the activation of the canonical Wnt/β-catenin signaling [138]. Surprisingly, despite its main commitment to the self-renewal of embryonic stem cells (ESCs), the overexpressed Nanog was reported to effectively stimulate osteogenic differentiation in modified BM-MSCs [139].
Finally, to ensure the high expression of pro-osteogenic genes in MSCs during the differentiation procedure, the regulatory properties of specific miRNAs could be used. During osteoblast differentiation, the miR-29b positively regulates this process by downregulating the inhibitory factors of osteogenic signaling pathways [140]. The osteoblast differentiation was obtained in the miR-29b-overexpressing BM-MSCs [141]; however, in the case of ASCs, Liao et al. reported that more pro-osteogenic potential was attributed to the miR-148b than to the miR-29b [142]. Furthermore, miR-148b-overexpressing ASCs also, cotransfected with the BMP-2 gene, possessed, enhanced, and prolonged BMP-2 gene expression.
A different study reported that the miR-31 suppression obtained in the anti-miR-31-overexpressing BM-MSCs resulted in a significant increase in the expression of osteogenic lineage genes in in vitro studies and bone formation was detected in vivo [143].
Engineered MSCs in Liver Regeneration
In the case of liver regeneration attempts, VEGF was likewise recognized to promote BM-MSC differentiation into hepatocyte-like cells under the specific condition of HGF and EGF stimulation [144]. However, most studies have been focused on attempts to induce hepatic differentiation of engineered MSCs with physiologically committed factors. To this end, a group of transcription factors involved in the physiological control of the hepatic commitment [145] were the subject of interest in the MSC studies. One of them, the hepatocyte nuclear factor 4α (HNF4α), is a specific hepatic lineage nuclear receptor [146]. The HNF4α-producing UC-MSCs differentiated into hepatocyte-like cells under hepatic differentiation conditions and activated various hepatic-specific genes, highlighting the key role of HNF4α in the hepatic lineage commitment [147].
Another HNF family member that regulates liver development, the representative of an HNF3 subfamily, HNF3β, was also investigated in terms of its ability to induce MSC differentiation. An efficient incorporation of the modified BM-MSCs into liver grafts was attributed to the HNF3β effects on the stimulation of the expression of antifibrotic molecules [148]. In the case of ASCs, an interesting approach was proposed by Alizadeh et al. [17], in which the let-7 family miRNA member, let-7b, was targeted, which assumes the physiological role of an inhibitor of hepatic differentiation that silences the expression of HNF4α [149]. In that study, the Let-7b-5p inhibitor was employed for ASC transduction. As a consequence, HNF4α gene expression was enhanced by removing the physiological inhibitor, thus allowing the engineered ASCs to differentiate toward hepatocyte-like cells [17].
What is more, the miR-122 is a key regulator in liver embryonic development and maintenance of hepatic homeostasis, and its overexpression is sufficient for hepatic differentiation of ESCs [150]. In the case of human ASCs, the overexpressed miR-122 was found to induce ASC differentiation into hepatocyte-like cells, confirmed by the increased expression of the specific hepatocyte markers and the production of urea and albumin by the differentiated ASCs [151]. On the contrary, Cui et al. found that the miR-122 was incapable of inducing hepatic differentiation alone in human UC-MSCs [152]. The hepatocyte-like cells were exclusively obtained from UC-MSCs after the concomitant transfection with six different miRNAs—miR-1290, miR-1246, miR-30a, miR-148a, miR-424, and miR-542-5p—previously thought to be overexpressed during hepatic differentiation [153].
Finally, MSCs may serve as therapeutic tools in the personalized medicine treatment of rare, inherited liver disorders. Promising results were obtained from a study on the Wilson's disease (WD) model. The hallmark of WD is the accumulation of elevated levels of copper in the liver due to the failure of the copper transporter, ATP7B, which is responsible for copper homeostasis [154]. The ATP7B-positive BM-MSCs were more resistant to the toxic effects exerted by the copper present in the culture medium than were control cells, indicating that this approach could be a promising solution for cell therapy of patients suffering from WD [155].
MSCs in Cardiac-Targeted Therapies
Native and genetically modified BM-MSCs have found an application in cardiac injury studies [156,157]. In that regard, the experiments have been focused on cardiomyocyte and pacemaker cell differentiation, while other strategies are focused on the improvement of the cardioprotective properties of the therapeutic cells after their administration to the injured myocardium.
BM-MSCs could be efficiently modified to gain cardioprotective properties. For example, extracellular superoxide dismutase-overexpressing MSCs were reported to exert cardiac protection and improve cardiac function by the antioxidative protection achieved by the inactivation of reactive oxygen species in mice with infarcted hearts [158]. GLP-1 is one of the factors that exhibit cardioprotective properties [159]. Therefore, GLP-1 was used for BM-MSC engineering with posterior encapsulation, and a prolonged supply of GLP-1 was maintained after the in vivo study [160].
The Wnt/β-catenin pathway is another subject of studies involving MSC engineering. Wnt proteins could act through two independent pathways: canonical, which includes β-catenin involvement, and β-catenin-independent noncanonical ways of signal transmission [161]. Furthermore, it has been suggested that the Wnt canonical signaling pathway promotes the cardiac commitment of the precursor cells at early developmental stages, but inhibits it later [162]. This dual Wnt/β-catenin signaling feature was used in BM-MSC differentiation attempts, while the BM-MSCs overexpressing the interference sequence for the β-catenin gene were shown to decrease the canonical Wnt pathway in favor of the noncanonical pathway, thus contributing to the myocardial-like cell commitment [163]. However, the overexpression of the Wnt11, which participates in the noncanonical Wnt signaling, resulted in the transdifferentiation of the engineered BM-MSCs toward cells with the cardiac phenotype [164].
HO-1 is an enzyme that degrades heme into biliverdin, free iron, and carbon monoxide [165]. HO-1 also elicits beneficial results during myocardial infarction after exogenous delivery [166]. The cardioprotective properties of HO-1 were evaluated in studies conducted on the HO-1-overexpressing BM-MSCs [167] and ASCs [168] delivered to injured cardiac tissue.
The differentiation of MSCs to generate new cardiomyocytes may occur by inducing the expression of genes physiologically involved in cardiomyocyte precursor differentiation signaling pathways. An example of this is myocardin, a crucial factor in heart development [169], whose overexpression was also accompanied by TERT gene overexpression and together promoted promyogenic gene expression and sustained the growth capacity of the modified ASCs [170]. One other protein involved in myocardial differentiation is a GATA family member, GATA-4. The representatives of the GATA family are transcription factors that bind to the GATA motifs among DNA regulatory sequences.
In particular, GATA-4 is the pivotal controller of cardiac differentiation and regulates cell survival in the adult heart [171]. In the GATA-4-overexpressing BM-MSCs, an increase in other genes committed to myocardial differentiation was detected and cell contraction was observed that might indicate that the abundant presence of GATA-4 paves the way for the BM-MSCs to transdifferentiate into and act as functional myocardial cells [172,173]. However, in the GATA-4-overexpressing BM-MSCs, the members of the miR-15 family were downregulated, which was translated into an increased resistance to the ischemic environment by the stimulation of antiapoptotic Bcl-2 family members [174].
In addition, the GATA-4-engineered BM-MSCs gained the cardiomyocyte protection properties by the production of microvesicles containing elevated amounts of the miR-221 [175]. Previously, it was stated that the miR-221 targeted the 3′-UTR of the p53-upregulated modulator of apoptosis (PUMA), which was a proapoptotic member of the Bcl-2 protein family. Furthermore, the inhibition of PUMA expression contributed significantly to cell survival by inhibition of the apoptosis process [176].
Cardiomyocyte differentiation of MSCs was reported after transfection with the antisense oligonucleotide, AMO-124, targeting miR-124, which, in turn, is involved in neurogenesis [177], but its depletion in BM-MSCs was reported to stimulate myogenic differentiation [178]. Cardiomyocyte commitment was also observed in the miR-133a and miR-499-overproducing BM-MSCs [18,179]. The miR-499 function was assigned to promote cardiac muscle differentiation by provoking the repression of the histone deacetylase 4 or sex-determining region Y-box 6 genes [180], while the miR-133a was shown to be expressed in adult cardiac and skeletal muscle tissues [181]. Functional recovery of the ischemic heart was reported after the cardiac administration of the miR-210 microvesicle-producing BM-MSCs [182], which is concordant with the reported neuroprotective effects of the miR-210 inhibiting apoptosis by silencing proapoptotic gene expression [183].
To produce cardiac pacemaker cells, MSCs were engineered to overexpress the hyperpolarization-activated, cyclic, nucleotide-gated channel (HCNs) genes. HCNs underlie the production of hyperpolarization-acivated cation currents (If), the common feature of the excitable cells [184]. The HCN family consists of four members where each channel type induces a hyperpolarization-activated current with a special activation kinetic [185].
The HCN4-modified BM-MSCs enabled a stable pacemaking function and an appropriate chronotropic response in dog hearts with an atrioventricular block [186]. However, in a different study, the HCN4-producing BM-MSCs were indeed able to produce certain If currents, but were poorly connected with the host cardiomyocytes, since there was a low presence of the gap junctions [187]. Next to this, the HCN1-modified BM-MSCs cultivated in vitro with neonatal rabbit ventricular myocytes were able to produce If currents and compel an increase in the spontaneous beating rate of the cocultured myocytes [19].
There is also a piece of evidence that BM-MSCs could be utilized for engineered tissue heart valves [188]. For this purpose, MSCs were transfected with the fibroblast inducible factor 14 (Fn14) gene, which, among other pathways, is involved in cardiomyocyte proliferation [189].
MSCs in Neural Differentiation and Repair
There has been increasing interest in MSC employment with neurological purposes. This interest stems from the fact that MSCs possess multiple immanent abilities to modulate neural cell growth and survival. It seems that MSCs act through paracrine actions rather than through direct differentiation, by attracting host NSCs to the sites of injury, and by enhancing their proliferation and differentiation in the damaged region [190]. In addition, native MSCs secrete a plethora of biologically active compounds, such as chemokines, anti- and proinflammatory cytokines, angiogenic factors, growth factors, and growth factor-binding proteins [9].
Even more significant for the neurological context, MSCs are able to express a variety of neuroregulatory molecule genes, such as the axon guidance molecules, neural cell adhesion molecules, neurite-inducing factors, neurotransmitter receptors, and neurotrophic factors [191]. Thus, MSCs of different origin have become an attractive pursuit in the studies on animal models of stem cell-based therapies in stroke (ie, BM-MSCs) [192,193], traumatic brain injury (ie, BM-MSCs) [194], and in several models of neurological disorders, such as in Parkinson's disease (PD) (ie, UC-MSCs) [195], amyotrophic lateral sclerosis (ALS) (ie, BM-MSCs) [196], multiple sclerosis (MS) (ie, ASCs) [197], Alzheimer's disease (AD) (ie, ASCs) [198], and Huntington disease (HD) (ie, UC-MSCs) [199]. MSCs could also be considered useful in studies with spinal cord injury models [200].
In addition to the experiments carried out with the administration of native MSCs, many examples exist of the use of genetic engineering methods to enhance the therapeutic effects of MSCs by granting MSCs new characteristics as a result of genetic manipulation.
In this regard, in the case of traumatic brain injury, intraventricularly transplanted BM-MSCs overexpressing one of the neurotrophic factors, brain-derived neurotrophic factor (BDNF), were presented to attenuate neuronal injury due to the elevated concentration of BDNF present in the injured brain [201]. In that case, BDNF exerted its beneficial effects deriving from its role as a member of the neurotrophin family, whose members are involved in neuronal survival during development and injury states [202]. In peripheral nerve regeneration studies, the BDNF-producing ASCs were engaged in nerve regeneration and differentiated into nonmyelinating Schwann cells, fostering significant functional recovery of the examined animals [203]. In addition, the BDNF-releasing BM-MSCs preserved motor function in a mouse model of HD [204].
In the case of studies on stroke, the BDNF-producing BM-MSCs transplanted into ischemic stroke animals protected host cells from apoptosis and accelerated the proliferation and maturation of endogenous NSCs situated in the subventricular zone of the host brains [22].
It is worth noting that the CXCR4-overexpressing BM-MSCs were able to ameliorate stroke effects after femoral vein injection due to the participation in the enhanced growth of capillary vascular volume among the stroke penumbra, a depletion in the volume of the damaged infarction area, and finally by an improvement in neurological functions [205].
Glial cell line-derived neurotrophic factor (GDNF) is critical for the development and protection of the adult central nervous system and peripheral neurons [206]. This feature was exploited to generate the GDNF-overexpressing BM-MSCs that exhibited elevated neuroprotective effects after their transplantation into rats with intracerebral hemorrhage [207]. In another case, genetically modified BM-MSCs that overexpressed GDNF together with neurotrophin-3 (NT-3), another member of the neutrophin family, were able to differentiate into neuronal cells and express nerve markers when induced by fetal gut culture medium. Those results indicate that these cells could be helpful in future therapies for Hirschsprung's disease, characterized by the disrupted migration of neural crest cells during embryogenesis [208].
In stroke models, there are studies describing the application of genetic engineering methods to produce neurons from MSCs. For instance, neurogenin 2 (Ngn2), which belongs to the transcription factor family with helix-loop-helix motifs, demonstrated proneural activities, confirming that it is one of the key factors during corticogenesis [209]. Ngn2 gene transfection to BM-MSCs was adequate to convert these cells into neuron-like cells in vitro, and Ngn2-positive BM-MSCs were able to diminish infarct volume and reduce the number of apoptotic cells in an in vivo stroke model. Furthermore, the functional recovery of the treated animals was appreciated as well [210]. However, BM-MSCs overexpressing another member of the transcription factors with a helix-loop-helix motif family, neurogenin 1 (Ngn1), were able to migrate with high efficiency to the central nervous system and exerted their paracrine functions on the host neural cells in an ALS mice model [211].
Another case reported data on the functional recovery after stroke obtained from animals with a BM-MSC-derived, exosome-mediated miRNA delivery to the injured area of stroke. BM-MSCs were engineered to overexpress miR-133b, which was formerly recognized as a neural cell modulator, since the miR-133b delivery to neural cells resulted in neurite outgrowth in vitro [212]. In an animal study, the miR-133b-containing exosomes were effectively transmitted from the administered miR-133b-overexpressing BM-MSCs to host neurons and astrocytes and influenced gene expressions with a subsequent positive effect on axonal plasticity and neurite remodeling in the ischemic boundary zone [213].
Hippocampal neovascularization was detected after the delivery of the VEGF-overexpressing BM-MSCs to the lateral ventricle in a murine model of AD [21], with the resultant beneficial behavioral effects showing the degree of the vascular damage in neurodegenerative diseases, such as AD. In addition, GLP-1, which was previously noted to exhibit neuroprotective capabilities in in vitro studies [214], was used for BM-MSC engineering, followed by right lateral ventricle administration into an AD animal model. Encapsulated GLP-1-positive BM-MSCs were responsible for a reduction in the accumulated amyloid-beta (Abeta) peptides in a mouse model of AD [215].
Studies carried out with PD rat models show some examples of the substantial, beneficial behavioral results after engineered BM-MSC administration with either GDNF [216,217] or persephin [218], which is another neurotrophic factor that promotes survival, proliferation, and differentiation of neurons [219].
MSC modifications, with the purpose of obtaining cell-based therapies to alleviate the symptoms of MS, have generally been focused on the inhibition of the autoimmune response, in which the myelin sheaths around the axons of the central nervous system are destroyed, causing demyelination and neuronal loss [220]. Vasoactive intestinal peptide (VIP)-producing ASCs [221], IL-10-secreting ASCs [222], and interferon-β [223]-releasing BM-MSCs effectively migrated to the inflamed central nervous system and diminished the autoimmune response of the host by a decrease in proinflammatory cytokine secretion, T-cell activity modulation, and the protection of the blood–brain barrier from new injuries.
In the case of spinal cord injuries, NT-3 was transduced in MSCs because of its action as a neuroprotective agent [224]. NT-3-positive BM-MSC administration to the injured spinal cords facilitated axonal regeneration, promoted neuronal survival [225], and favored remyelination and functional recovery in the demyelinated spinal cords of the studied animals [226]. NT-3 acts through tropomyosin-related kinase receptor tyrosine kinases (Trk) [227], a feature that was used for BM-MSC transduction with Trk type C (TrkC) gene overexpression [228]. TrkC-producing MSCs possessed the ability to prolong their survival after delivery, improve differentiation toward neuron-like cells, and participate in corticospinal tract regeneration in the injured areas, mainly due to the elevated concentration of NT-3.
Another neurotrophic factor that acts through the Trk-mediated signaling pathway is multineurotrophin (MNTS-1). MNTS1 is a mutated form of NT-3 that could bind efficiently all members of the Trk family [229]. After the administration of the MNTS1-producing BM-MSCs to the sites of spinal cord injuries, a noticeable reduction in the cavity volume and intense axonal growth were detected [230].
Sonic hedgehog (Shh) is a pleiotropic factor in the developing central nervous system and spinal cord [231]. Engineered BM-MSCs secreting Shh enhanced VEGF expression in the injured area, thereby causing an improvement in the local microenvironment and leading to enhanced functional recovery [232].
Finally, there is some evidence of positive outcomes after genetically engineered BM-MSC administration in animal models of epilepsy [233], autoimmune encephalomyelitis [234], and vascular dementia [235].
Another broad-range research interest is to establish efficient approaches for MSCs to force them to differentiate toward functional neurons in vitro by means of genetic engineering. For this purpose, several genes were selected to be overexpressed in MSCs. Nerve growth factor (NGF), a member of the neurotrophin family noted to stimulate neuronal survival and neurite outgrowths, was used in MSC engineering. The NGF-producing BM-MSCs were effectively changed into cells with a neuron-like phenotype [236].
GDNF overexpression, combined with retinoid acid (RA) stimulation, resulted in the enhanced ability of transfected BM-MSCs to differentiate into neuron-like cells, which was confirmed by the expression of specific neuronal marker microtubule-associated protein 2 (MAP-2) gene [20]. Caveolin-1 was found to interact with the membrane receptor Notch-1, which, in turn, is involved in neuronal [237] and astroglial differentiation of NSCs [238]. In the case of BM-MSCs, the downregulation of the Caveolin-1 gene expression was achieved with siRNA engineering and modified cells were more sensitive for neuronal differentiation, which was affirmed by the expression of several neural-specific markers [239].
Trans-RA participated in embryonic nervous system development and nerve cell differentiation by acting through the retinoic acid receptor β (RAR-β) [240]. For this reason, the RAR-β gene was overexpressed in BM-MSCs to improve neuronal differentiation [241]. The increased presence of the RAR-β in the nuclei of transduced BM-MSCs made these cells more susceptible to the RA signaling pathway, which, in turn, helped facilitate neuronal differentiation of the RAR-β-presenting BM-MSCs.
Another protein, Nurr1, is a transcription factor that is elemental for the development and maintenance of dopaminergic neurons [242]. BM-MSCs were modified to overexpress the Nurr1 gene with simultaneous stimulation with electrical pulses. As a result, the Nurr1-positive BM-MSCs produced neurites and expressed a series of neurogenic markers, such as Tuj1, nestin, and Map 2 [243]. Yet another protein used, a basic helix-loop-helix transcription factor member, mammalian achaete-scute homolog-1 (Mash1), was reportedly an efficient inducer of BM-MSC neuronal differentiation [244].
The miRNA approaches could be valid as well. The miR-9, with its target, the zinc finger protein 521 (Zfp521), the inhibitor of neuronal differentiation, and miR-125b, important in neuron-like cell differentiation, were implemented for BM-MSC engineering. In both cases, the modified BM-MSCs were shown to become neuron-like cells [245,246].
Other Applications
Ultimately, there are some studies on the use of MSCs in the therapeutic strategy against bladder (ie, fetal BM-MSCs) [247] and hepatic fibrosis by the overexpression of HGF by UC- and BM-MSCs [248 –250]; differentiation of ASCs into hair cell-like cells by Atoh1 overexpression [251]; Achilles allograft replacement through transforming growth factor-1 transfection produced by BM-MSCs [252]; attenuation of intestinal injury by heparin-binding EGF-like growth factor overexpression [27]; and protection against acute lung injury by keratinocyte growth factor expression [253]. MSCs have also been used against acute retinal injury through neurotrophin-4 production [254] and in UC-MSCs with a decrease in excessive inflammatory injury after acute radiation injury by thioredoxin gene overexpression [255]. Furthermore, there is some evidence that the DAZL gene overexpression in BM-MSCs converts these cells into putative germ cells [256].
Several attempts were focused on pancreatic lineage differentiation. The favorable MSC differentiation toward insulin-producing cells was achieved by the overexpression of several genes such as betacellulin reported for BM-MSCs [257], and in the case of BM-MSCs and UC-MSCs, there are some data for pancreatic and duodenal homeobox factor 1 (PDX-1) [258,259], combined with other lineage-specific genes [260,261], and pancreatic characteristic miRNAs, such as the miR-375 in the human placental decidua basalis-derived MSCs [262], or the insulin gene itself in hair follicle-derived MSCs [263].
The VEGF and human IL-1 receptor antagonist (IL-1Ra)-overexpressing BM-MSCs were shown to protect islet cells from cytokines during islet transplantation [264]. Similarly, the immunomodulatory properties of IL1Ra were also shown in the case of IL-1Ra-modified ASCs [265] and BM-MSCs [266] in a cartilage repair model, where the modified MSCs protected the development of new cartilage tissue from the effects of IL-1 within proinflammatory surroundings.
In the studies on nephritis, the glutathione S-transferase M1 (GSTM1)-producing BM-MSCs ameliorated renal pathological damage due to the inhibition of the oxidative stress-induced inflammation by reactive oxygen species removal [267]. The HGF-overexpressing BM-MSCs reduced renal fibrosis after ureteral obstruction [23], and survivin-producing BM-MSCs were found beneficial in renal injury in a mouse model, probably owing to the prolonged survival and increased cytokine production by the engrafted BM-MSCs [268]. In addition to its antifibrotic effects, in the case of the HGF-expressing ASCs, HGF was reported to stimulate thymus regeneration in a rat thymus involution model [269].
One more application of engineered MSCs is the introduction of genetic modifications to gain more prominent anti-inflammatory properties. For instance, the production of anti-inflammatory cytokines such as IL-10 was achieved in BM-MSCs [24] and ASCs [222,270] and IL-4 in BM-MSCs [271]. Curiously, in BM-MSCs transfected with proinflammatory TNF-α, an immunosuppressive activation was observed [272].
Safety and Feasibility Issues Regarding Future Therapies with Engineered MSCs
Fortunately, therapies based on the transplantation of MSCs do not raise significant ethical and legal objections, which are in stark contrast with some other types of stem cells, especially human ESCs [273]. Due to the growing interest and hopes for therapies based on naïve and engineered MSCs, safety and feasibility aspects are extremely important and should be taken into account. These two aspects, if not properly addressed, could pose significant drawbacks to clinical use and, for this reason, should be considered in addition to the aforementioned beneficial effects. Only a comprehensive approach will result in the realistic and practical use of the therapeutic properties of engineered MSCs.
One of the fundamental problems related to genetic cell engineering, when considering clinical trials, is achieving reliable expression and low risks. Viral transduction methods, despite their reported high efficiency of transgene expression, are unlikely to be allowed in clinical trials because of their high risk of insertional mutagenesis [274], chromosomal instability [275], and proto-oncogene activation. Even the use of self-inactivating lentiviral vectors has been shown to be an unsafe solution [276].
A transposon system is a nonviral transfection method that has been used for stem cell engineering [277], including MSCs [278,279]. However, in this case, there is also a risk of insertional mutagenesis [280]. For this reason, the approaches that use putative safe integration sites among target genomes are under consideration for MSC engineering, such as the zinc finger nuclease-based genome-specific locus targeting used for the Epo gene introduction into the MSC genome [281]. What is more, other site-specific genome engineering tools exist, such as transcription activator-like effector nucleases [282] and clustered regulatory interspaced short palindromic repeat/Cas-based RNA-guided DNA endonuclease targeting [283], which could be potentially engaged in MSC modifications.
However, despite the aforementioned solutions, methods that do not lead to genomic integration, such as DNA- and RNA-based transfections, pose a tempting solution for stem cell-based therapies [33]. Unfortunately, in MSCs, DNA-based transfections often result in rather low efficiency; thus, new approaches aimed at improving this strategy are being developed [284]. The goal is to provide an efficient, safe, and yet effective transfection method, which would result in a transduction efficiency comparable with viral methods [285].
It has been shown that viral-free transfection of MSCs is not without risk as it may affect cellular viability and differentiation potential [286]. Therefore, there is still a need to develop new more sophisticated and technically advanced methods, such as plasmid delivery with nanoparticles based on the polysaccharide from Angelica Sinensis [287] or by using microbubble-mediated ultrasound and a polyethylenimine compound [288].
Another attractive method for cell engineering that does not lead to genome integration and provides fairly high transfection efficiency is mRNA-based transfection [30]. This method has been shown to be effective in modulating MSC differentiation [289]; however, it should always be borne in mind that mRNA-based transfection is of a transient nature, so it may pose a problem in applications that require a long operating time, especially in long-term clinical trials.
In addition to biological issues, the financial and legal regulations must be considered when developing putative stem cell therapies [290,291]. Despite the fact that the engineering of MSCs has proven a promising strategy in preclinical research, the application of that technology in the clinic requires a cost-benefit analysis to assess whether the advantages of cell engineering outweigh the cost. The overall costs apply to two main categories: the cost of the engineering procedure itself and the cost of approval by regulatory bodies. Both these can be prohibitive; however, this may differ in various countries, introducing the risk of unequal access to therapeutic procedures in the global healthcare market.
The cost-effectiveness has been already evaluated in the field of tissue engineering [292], showing that the tissue engineering costs are less than those associated with obtaining material from donors for transplantation. This means that tissue engineering may be more accessible to a larger number of patients. A similar financial analysis is expected to be applied to the field of cell engineering once such therapies prove efficacious in clinical trials. If the magnitude of improvement is significantly greater than the costs, it will stimulate the marketing of therapies based on engineered cells [293,294].
Overall, the relatively high financial expenses and complex biological aspects, as well as the wide possibilities offered by genetic engineering, make MSC-based therapies a very interesting research area, but, due to still unresolved issues, this field is in the early phase of development and therapeutic applications are at the preclinical stage.
Conclusion
This review presents an overview of recently published data regarding the spectrum of experiments that have focused on the increased therapeutic relevance of MSCs by means of genetic engineering applicable to regenerative medicine. The presented modifications relate to many aspects of MSC physiology, improve their natural ability to differentiate into bone or cartilage tissues, and introduce other multiple approaches designed to improve the therapeutic potential of MSCs, which subsequently could be applied in various types of pathological incidences, such as cardiovascular and neurological diseases. All of these aspects as well as safety considerations pose an interesting target for future clinical trials with engineered MSCs.
Footnotes
Acknowledgments
The study was supported by a National Centre for Research and Development grant no. 101 in the ERA-NET NEURON project: MEMS-IRBI. The authors thank Mary McAllister for editorial assistance.
Author Disclosure Statement
No competing financial interests exist.