
Abstract
Activins can exert several roles in ovary development. However, little is known about their involvement in early mammalian oogenesis. In this study, we reported that activin receptors (including ActRIA, ActRIB, ActRIIA, and ActRIIB) are expressed throughout the development of the mouse ovaries from 12.5 days postcoitum (dpc) to 21 days postparturition (dpp). Moreover, we found that in vitro, the addition of activin A (ActA) to the culture medium of 12.5 dpc ovarian tissues accelerated the progression of oocytes throughout meiotic prophase I stages. This result was reproduced in vivo following administration of ActA to pregnant mice. The in vitro effect of ActA was associated with increased expression of premeiotic and meiotic genes (including Dazl, Spo11, Stra8, Scp3, and Rec8) in the ovarian tissues. Mechanistically, ActA-dependent SMAD3 signaling modulated the expression of members of the retinoic acid (RA) system, including the RA degradation CYP26B1 enzyme and the RA receptors. Finally, ActA promoted the survival and growth of fetal and early postnatal oocytes and primordial follicle assembly both in vitro and in vivo. In conclusion, the present study identifies new roles of ActA in early oogenesis and suggested that ActA and RA might cooperate in promoting meiosis in female germ cells.
Introduction
T
Several members of the transforming growth factor beta (TGFβ) superfamily are crucial regulators of ovarian functions [1]. Among these, activins are important modulators of ovarian follicle development in many mammalian species, including humans, during the fetal and postnatal periods [1]. Activins are dimers comprising two identical or very similar β subunits. In mammals, two main subunits, called activin βA and activin βB, have been described. Two βA subunits give rise to activin A (ActA) and two βB subunits give rise to activin B (ActB), whereas one βA subunit and one βB subunit give rise to activin AB [1,2]. Inhibin is a closely related dimer comprising one β subunit similar or identical to the activin β subunit and a more distantly related α subunit. Inhibin generally has opposite effects to activins [3]. The TGFβ superfamily signals through membrane-bound serine–threonine kinase receptors [2]. For example, activins bind to a type II receptor (ActRIIA or ActRIIB), which recruits and phosphorylates a type I receptor, leading to the subsequent modulation of gene expression through SMAD protein activation [2].
In the human embryo, activin βA and βB subunits were immunodetected in the gonadal region at very early stages of development [3,4], while mRNAs for the ActA and ActB subunits and the activin receptors, ActRIIA and ActRIIB, were found in ovaries of fetuses at 14–21 weeks of gestation [4]. Moreover, in the fetal ovaries, the production of the βA unit appears to be stimulated by PGE2 [5]. It has been suggested that ActA produced by the human female germ cells promotes the expression of signals, including neurotrophins produced by the companion pregranulosa cells, which stimulate oogonia proliferation and oocyte survival [4]. On the other hand, ActA appears to repress signals that facilitate oocyte maturation and follicle formation [6 –8]. Exogenous ActA affected the number of follicles at puberty, which indirectly affected germ cells through selectively repressing the expression of the membrane-bound isoform of Kit ligand, rather than being an autocrine germ cell effect [6 –8].
In the mouse, βA and βB subunits and ActRIIA or ActRIIB receptors have been reported to be expressed in the gonadal ridges (GRs) at the time of PGC proliferation and GR colonization [around 12.5 days postcoitum (dpc)] [9]. Moreover, transcripts of ActRIIB and ActRIB were expressed in 11.5 dpc PGCs, while by 12.5 dpc, inhibin B was expressed in the ovaries [10,11]. Between days 0 and 4 after birth, βA and βB subunits and ActRIB (or ALK4), ActRIIA, and ActRIIB were present in both pregranulosa cells and oocytes. Contrary to humans, ActA was shown to inhibit the proliferation of mouse PGCs cultured in vitro at 8.5–11.5 dpc [10], while it appeared to stimulate oogonia proliferation and/or promote oocyte survival within cultured 12.5 dpc fetal ovaries or early postnatal ovaries [12 –14]. The effect on these two processes suggests that in the mouse, ActA reduces the germ cell nest breakdown and increases the initial primordial follicle pool [6,12]. This notion was supported by the observation that follistatin, a potent activin antagonist, was expressed in mouse oocytes at birth and ablation of its isoforms resulted in a marked increase of the initial primordial follicle pool [13,15]. Interestingly, in the perinatal mouse ovaries, estradiol, known to suppress germ cell nest breakdown and primordial follicle formation, represses the expression of ActA subunits [16], whereas activin regulates estrogen receptor expression [17]. Like in humans, some of the ActA effects in the prenatal mouse ovaries might be attributed to its ability to stimulate neurotrophin gene expression, namely Ntf5, in pregranulosa cells. Finally, in the fetal mouse ovaries, WNT4/β-catenin pathways maintain the oocyte survival and prevent testis-specific vasculature by inhibiting βB subunit expression [11,18], whereas the loss of activin through gene knockout, activin receptor elimination, or overexpression of dominant-negative cytoplasmic coregulators, results in gonadal dysgenesis or inappropriate follicle function [14,19 –21].
Our previous studies have shown that the presence of ActA during the culture of explants of embryonic ovaries and the subsequent coculture of growing oocytes generated within the explants onto granulosa cell monolayers were crucial to obtain a significant number of fertilizable oocytes able to develop to the morula/blastocyst stages [14]. Based on these results, the objective of the present study was to determine the effects of ActA on the early processes (particularly the entry of meiosis) of oogenesis in the mouse and to identify possible pathways by which ActA achieves its action.
Materials and Methods
Animals
All procedures described in the present study were reviewed and approved by the Ethics Committee of Qingdao Agricultural University. CD-1 mice (Vital River) used for all experiments were maintained on a 12-h light–12-h dark cycle (lights off at 20:00 h) with food and water available ad libitum. Animals were cared for in accordance with all national and institutional guidelines. Mating was timed overnight and the appearance of vaginal plug was considered as 0.5 dpc the next morning.
Ovarian culture
Fetal ovaries were collected and cultured as described previously [14]. Briefly, ovaries dissected from CD-1 embryos at 12.5 dpc were divided into two pieces of equal size and cultured in 600 μL growth medium in 24-well plates (Boyang) in a humidified incubator at 37°C with 5% CO2. The growth medium comprised α-minimal essential medium (Hyclone) supplemented with 10% heat-inactivated fetal bovine serum (FBS, 10099-141; Gibco), 0.23 mM pyruvic acid, 10 mIU/mL follicle-stimulating hormone (Sigma), 100 mIU/mL penicillin G, and 100 mg/mL streptomycin sulfate. Half of the medium was replaced with fresh medium every other day. ActA (Sigma) at a concentration of 100 ng/mL and SB431542 at 5 μM (Sigma) were added to the culture medium since previous studies demonstrated that cells cultured with 10 μM SB431542 for 48 h had no impact on cell viability [22]. At the end of the culturing, the ovarian tissues were collected and washed thrice with phosphate-buffered saline (PBS) for the indicated analyses. Oocytes were isolated from the cultured ovarian tissues as previously described [23 –27].
Immunofluorescence
For immunofluorescence (IF), ovaries were collected and mechanically disaggregated into single cells after 5 min of incubation at room temperature in 0.25% trypsin and 0.02% EDTA (SH30848.01B; Hyclone). After three times of washes with PBS, the cells were fixed in 4% paraformaldehyde (PFA; Beyotime), spread onto poly-
Evaluation of meiotic prophase I stages
The meiotic prophase I stages were evaluated using oocyte cytospreads. Briefly, single cells from digestion of the ovarian tissues were incubated in 1% sodium citrate for 30 min at room temperature, fixed in 4% PFA, and then spread onto poly-
Tissue processing and follicle counting
Ovaries were processed for paraffin inclusion and serial sectioning following standard procedure. Follicles were double-blind classified and counted in every fifth sections. Intact oocytes were collected if there are no surrounding granulosa cells or they were stained positive for MVH. Primordial follicles were classified as an oocyte partially surrounded by either squamous granulosa cells or squamous and cuboidal granulosa cells. Primary follicles contained a small oocyte completely surrounded by a single layer of cuboidal granulosa cells. All primordial and primary follicles and germ cell nests were counted regardless of the presence or absence of the oocyte nucleus [23 –25,27].
Western blot analysis
Briefly, total proteins were extracted from the ovaries after incubation in an RIPA lysis solution (P0013C; Beyotime) for 30 min on ice with frequent vortexing. Sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) sample loading buffer (5 μL/ovary) was then added and boiled for 5 min. The lysates were collected by centrifugation at 14,000 rpm for 5 min at 4°C. The proteins were separated by SDS-PAGE with a 4% stacking gel and a 10% separating gel for 50 min at 100 V and 2.5 h at 120 V, respectively, and then transferred onto a polyvinylidene fluoride membrane by electrophoresis. After being blocked at 4°C overnight in TBST buffer containing 10% BSA, the membranes were incubated with primary antibodies for 5 h at 4°C. Finally, the membranes were washed with TBST thrice, followed by incubation with horseradish peroxidase (HRP)-conjugated goat anti-rabbit or mouse IgG (A0208; Beyotime) at a dilution of 1:2,000 in TBST. PWIN software was employed for the densitometric analyses. All experiments were repeated at least thrice.
Primary antibodies, including rabbit anti-MVH polyclonal antibody (ab13840; Abcam), rabbit anti-DAZL polyclonal antibody (ab34139; Abcam), rabbit anti-STRA8 antibody (ab49602; Abcam), mouse anti-Actin antibody (ab8226; Abcam), activin RIB/ALK-4 Mab (MAB1477; R&D), mouse activin RIIB Mab (3725-RB-050; R&D), mouse IgG1 (MAB3393; R&D), rabbit anti-SMAD3 antibody (ab28379; Abcam), and goat anti-CYP26B1 antibody (EB07481; Everest Biotech), and secondary antibodies, including HRP-conjugated goat anti-mouse IgG (A0216; Beyotime), HRP-conjugated goat anti-rat IgG (A0192; Beyotime), HRP-conjugated goat anti-rabbit IgG (A0239; Beyotime), and HRP-conjugated donkey anti-goat IgG (A0181; Beyotime), were used in this study [23]. The BeyoECL plus Kit (P0018; Beyotime) was used for signal development. The intensity was analyzed by FluorChem FC3 and AlphaView SA software (ProteinSimple).
RNA extraction, cDNA synthesis, and quantitative real-time polymerase chain reaction
Total RNA was extracted using the RNAprep pure Micro Kit (RN07; Aidlab), and cDNA was synthesized by the SYBR® Premix Ex Taq™ II Kit (DRR047A; TaKaRa), as described previously [26]. Primers used in this study for quantitative real-time polymerase chain reaction (RT-qPCR) analysis are shown in Supplementary Tables S1–S3; Supplementary Data are available online at
Construction of eukaryotic expression vector
The primers of Smad3 were forward: 5′- CAG CCA TGT CGT CCA TCCT −3′ and reverse: 5′- AAC CCG CTC CCT TTA CTCC-3′. The PCR products were separated by electrophoresis using 2% agarose gel, and the correct bands were excised from the gel and purified with the Wizard® SV Gel and PCR Clean-Up System (A9285; Promega). The purified DNA was then cloned into a pMD19-T Vector (D101A; TaKaRa) according to the manufacturer's instructions. The positive clones were obtained by antibiotic selection and the insert was sequenced at the union gene. Subsequently, the inserts were ligated to N1 vector [28] or pcDNA3.1(+) vector and the ligation mixture was transformed into the DH5α competent cells on the ampicillin-selective media plate. Recombinant eukaryotic expression vector, pSmad3-cDNA 3.1(+), was obtained. Finally, pSmad3-cDNA 3.1(+) was digested by ScaI (SCA-103; TOYOBO). The linearized vectors were extracted by the gel extraction kit and transfection was performed by electroporation.
Cell transfection
The fetal ovaries from the mice at 12.5 dpc were first digested by 0.25% trypsin for 5 min at 37°C and gently transferred together with the DNA construct into a BTX 4-mm cuvette. Electroporation was carried out at 600 voltage for 2 ms. Electroporated cells were left on ice for 10 min before transferring to a 10-cm culture plate containing DMEM/F12 medium supplemented with 10% FBS, 100 U/mL penicillin, and 100 mg/L streptomycin saturated in 5% CO2 at 37°C. Negative control (without DNA) and positive control (with linearized pEGFP-N1) were treated with same procedures [28].
Statistical analysis
Results from at least three independent experiments are presented as the mean±SEM. Statistical significance was determined with GraphPad Prism analysis software using unpaired Student's t-test with two-tailed distribution of three samples of unequal variance. P<0.05 was considered as significantly different, while P<0.01 was considered as a highly significant difference.
Results
Expression of ActA receptors throughout the ovary development
Before studying the effect of ActA on early oogenesis, we first examined the expression of ActA receptors, including ActRIA (ALK2), ActRIB (ALK4), ActRIIA, and ActRIIB, in ovaries at 12.5 dpc, 0, 7, 14, and 21 days postparturition (dpp) by RT-qPCR. The results showed that all types of activin receptors were expressed throughout the ovary development (Fig. 1A). ActRIA and ActRIB transcripts were expressed at a low level at 12.5 dpc, reached the peak at 7 dpp, underwent a marked reduction at 14 dpp, and increased again at 21 dpp. ActRIIA and ActRIIB mRNA showed a similar expression pattern, but with maximum expression at 0 dpp. The expression of ActA receptors at the protein level was consistent with the mRNA amounts (Fig. 1B). Moreover, IF analysis showed that ActRIB and ActRIIB were present in oocytes and/or follicular cells, depending on the developmental stages. At 12.5 dpc and 0 dpp, ActA receptors were detected mainly in oocytes, while they were progressively confined to the follicular cells at later stages (Supplementary Fig. S1A, B).
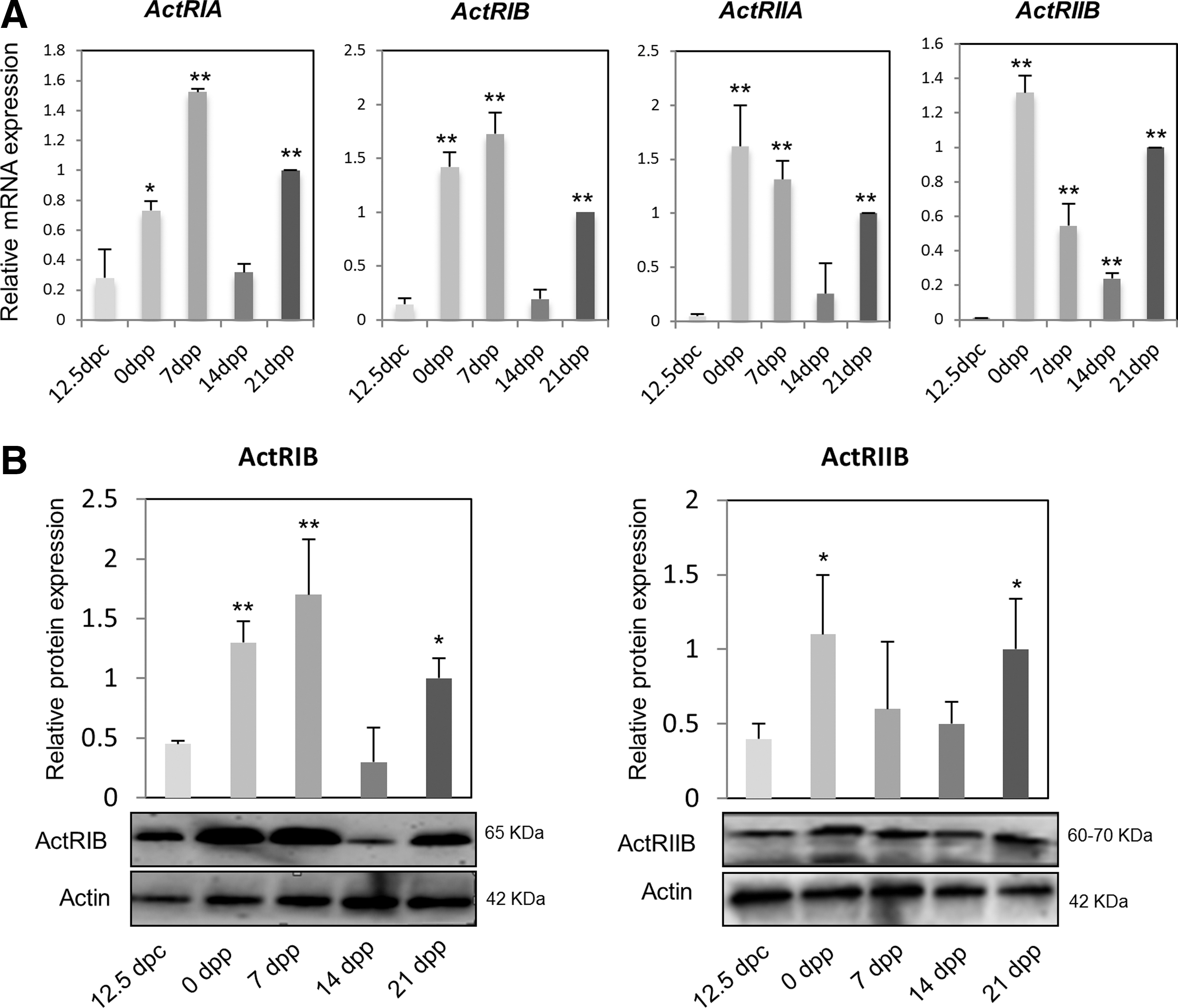
Activin receptors are expressed throughout the process of oogenesis.
ActA accelerates the progression of oocytes through meiotic prophase I
Entering into meiosis and the progression through the prophase I stages are major events of the prenatal oocyte development. Using a culture system of ovarian tissues previously shown to be suitable to recapitulate these events in vitro [14], we examined the effect of ActA on such processes. IF analysis using antibody against synaptonemal complex protein 3 (SCP3) showed that the percentage of oocytes at the leptotene stage was significantly reduced after addition of ActA in the culture medium for 4 days (56.16%±3.42% vs. 39.92%±3.23%, P<0.05) (Fig. 2A, B). In contrast, the percentage of oocytes at the zygotene and diplotene was increased by ActA (zygotene: ActA=40.20%±2.48% vs. control=32.24%±1.82%; pachytene: ActA=16.94%±2.11% vs. control=9%±1.33%; P<0.05) (Fig. 2A, B). Such effect was partially abolished by adding the ActA receptor inhibitor, SB431542, to the culture medium (Fig. 2A, B). These results provided evidence that ActA might promote meiotic progression. Moreover, we found that the number of cells positive for SCP3 and two other meiotic proteins (STRA8 and DAZL) in the ovarian tissues were significantly increased in the presence of ActA in comparison with controls (Fig. 2C). The expression level of these proteins and their coding genes, as evaluated by western blot and real time-PCR, respectively, was higher in ActA-treated ovarian tissues than in the controls (Fig. 2D; P<0.05 or 0.01). The transcripts of two other meiotic genes (Rec8 and Spo11) were also affected in a similar way (Fig. 2E). Interestingly, the addition of 1 μM retinoic acid (RA) to the culture of ovarian tissues at 12.5 dpc caused a marked increase of STRA8 expression and this was further enhanced when RA was added together with ActA (Supplementary Fig. S2A). Collectively, these observations indicated that ActA facilitates oocyte progression throughout the early stages of meiotic prophase I, possibly by promoting the expression of key meiotic genes in such cells.
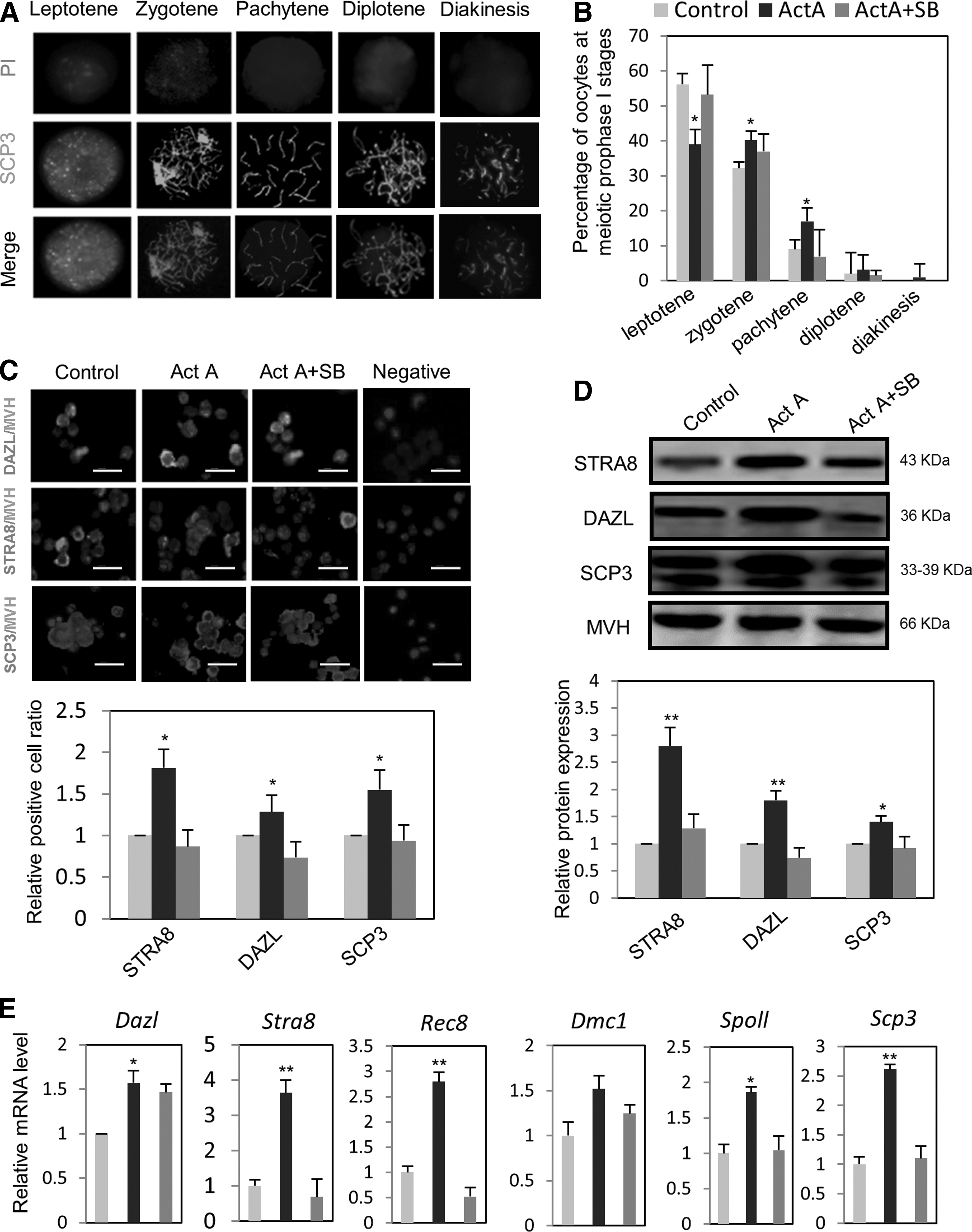
The effect of ActA on meiotic progression of oocytes throughout prophase I stages and the expression of meiosis-specific genes in vitro.
We next examined the effect of ActA on oocyte meiotic progression in vivo. Pregnant mice at 10.5 dpc were treated with a single intraperitoneal injection of ActA (60 or 100 ng/kg) or vehicle dimethyl sulfoxide (DMSO) as control and the ovaries harvested at 16.5 dpc [14]. The results showed that the percentage of oocytes at zygotene stages was decreased, while the percentages of oocytes at diplotene/diakinesis stages were increased by ActA treatment (Fig. 3A). These results confirmed an acceleration of meiotic progression in the oocytes of ActA-treated ovaries. In addition, like in vitro, the expression levels of both mRNA and protein of several meiotic genes, such as Stra8, Scp3, Dazl, and Dmc1, were significantly increased in oocytes obtained from the ovaries of 15.5 dpc fetuses of the ActA-treated mothers (Fig. 3B–D).
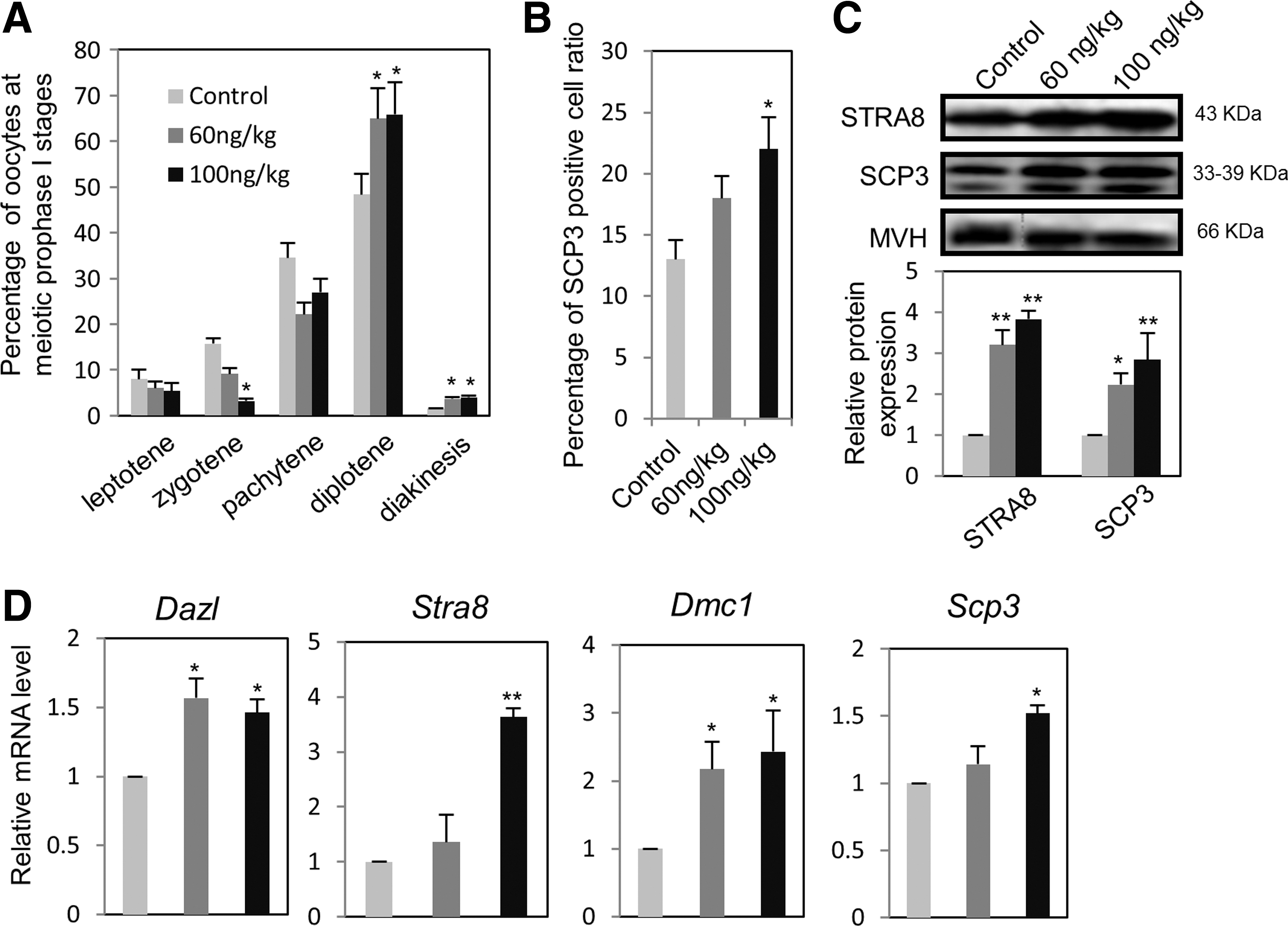
Effects of ActA on oocyte meiotic progression and expression of meiosis-specific genes in vitro.
ActA-dependent SMAD3 signaling modulates the expression of the RA system
To identify possible pathways by which ActA acts in female GRs and fetal ovaries, we determined the effect of ActA on the expression of SMAD3, a common mediator of ActA signaling, and the RA system, including the RA degradation CYP26B1 enzyme and the retinoic acid receptors (RARs). As shown in Figure 4A, CYP26B1 protein was expressed at a high level at 11.5 dpc, decreased at 12.5 dpc, and disappeared thereafter. Conversely, SMAD3 and MVH-positive female germ cells were observed by IF at 12.5 dpc (Supplementary Fig. S2B), while immunoblotting revealed a signal at 13.5 dpc that was progressively increased at later stages (Fig. 4A). After addition of ActA to the culture of ovarian tissues at 12.5 dpc for 3 days, CYP26B1 expression at both protein and mRNA levels was significantly reduced in comparison with the control. Such effect was partially abolished by the contemporary addition of the inhibitor, SB431542 (Fig. 4B, C). Furthermore, the results showed that under the same ovarian tissue culture conditions, ActA caused a significant increase of RAR transcripts, but did not affect the AKT pathway (Supplementary Fig. S2C, D).
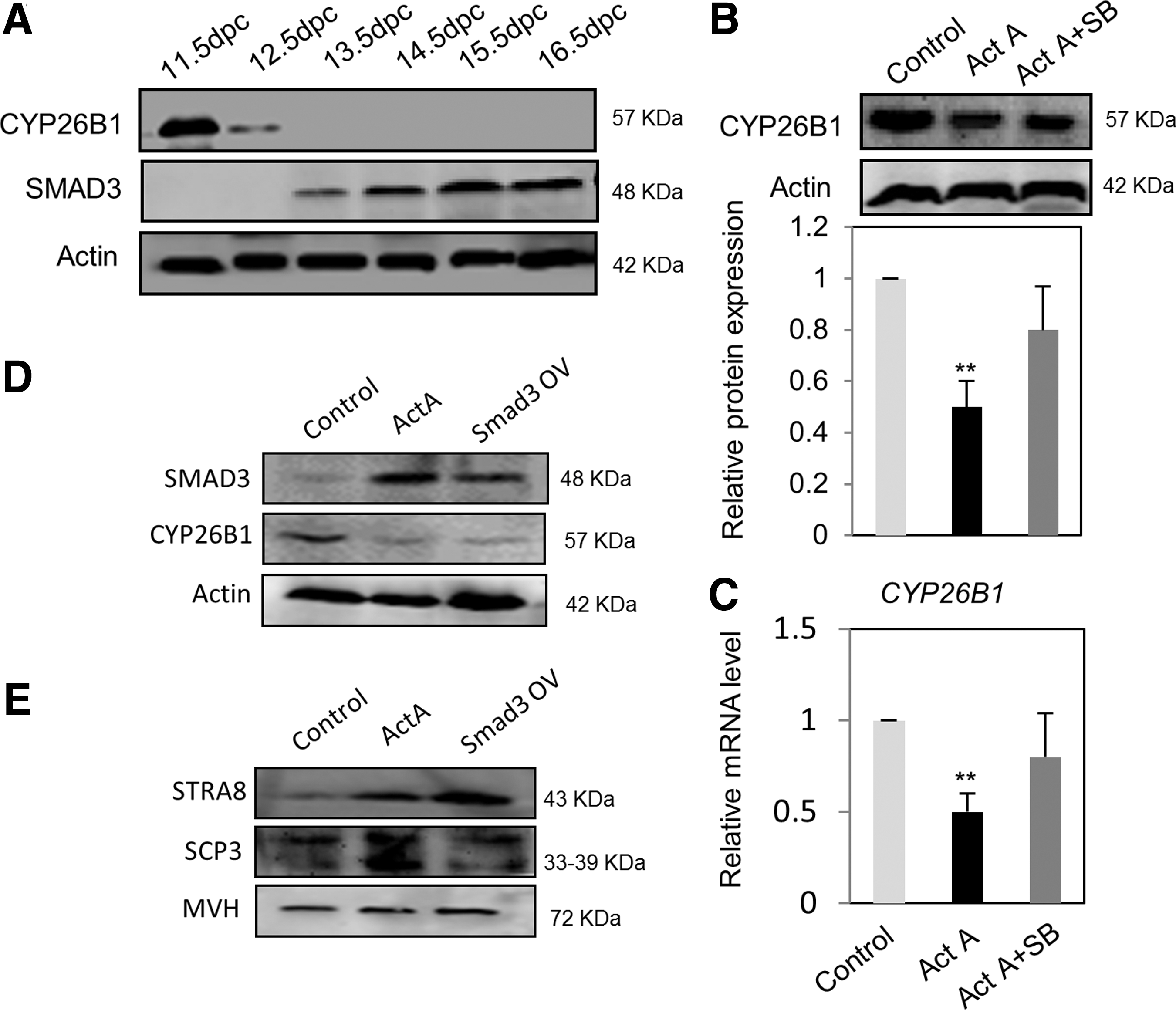
The expression of Cyp26b1 is modulated by ActA-dependent SMAD3 signaling.
On the basis of these results, we hypothesized that there is a correlation between Cyp26b1 downregulation and Smad3 expression possibly mediated by ActA. To test this hypothesis, we transfected pregranulosa cells obtained from ovaries at 12.5 dpc with Smad3-cDNA3.1 constructs. As expected, transfection with Smad3-cDNA3.1 increased SMAD3 and caused a significant decrease of CYP26B1 expression. Under such conditions, a decrease of CYP26B1 by ActA was also observed (Fig. 4D). Similarly, the expression level of STRA8 and SCP3 proteins in 12.5 dpc pregranulosa cells was increased after transfection with Smad3-cDNA3.1 (Fig. 4E). Taken together, these results supported our hypothesis that Cyp26b1 downregulation is driven by ActA-dependent SMAD signaling.
ActA promotes oocyte survival and growth
To further determine the effects of ActA on early oogenesis processes, we cultured ovarian tissues at 12.5 dpc in the presence of 100 ng/mL ActA for different periods up to 28 days according to the protocols shown in Figure 5A [14,19]. In line with our previous results, we observed that the higher number and the best growth of the oocytes were achieved in the continuous presence of 100 ng/mL ActA. In fact, when this compound was omitted from the culture medium for the first or the second 7 days of culture, the number and the diameter of the oocytes were significantly decreased. On the other hand, only minor effects were observed when ActA was absent during the second to the last weeks or only the last week (Fig. 5B, C). The beneficial effects of ActA on the oocyte number and growth were associated with an increase of transcripts of growing oocyte genes, such as Zp3, Cx37, Gdf-9, Kitl, CyclinB, and BMP-15, in oocytes cultured in the continuous presence of the factor compared with those in its absence (Fig. 5D). Moreover, in the presence of ActA, the number of apoptotic oocytes (TUNEL staining) and the ratio of BAX/BCL-2 were decreased (Supplementary Fig. S3A–C).
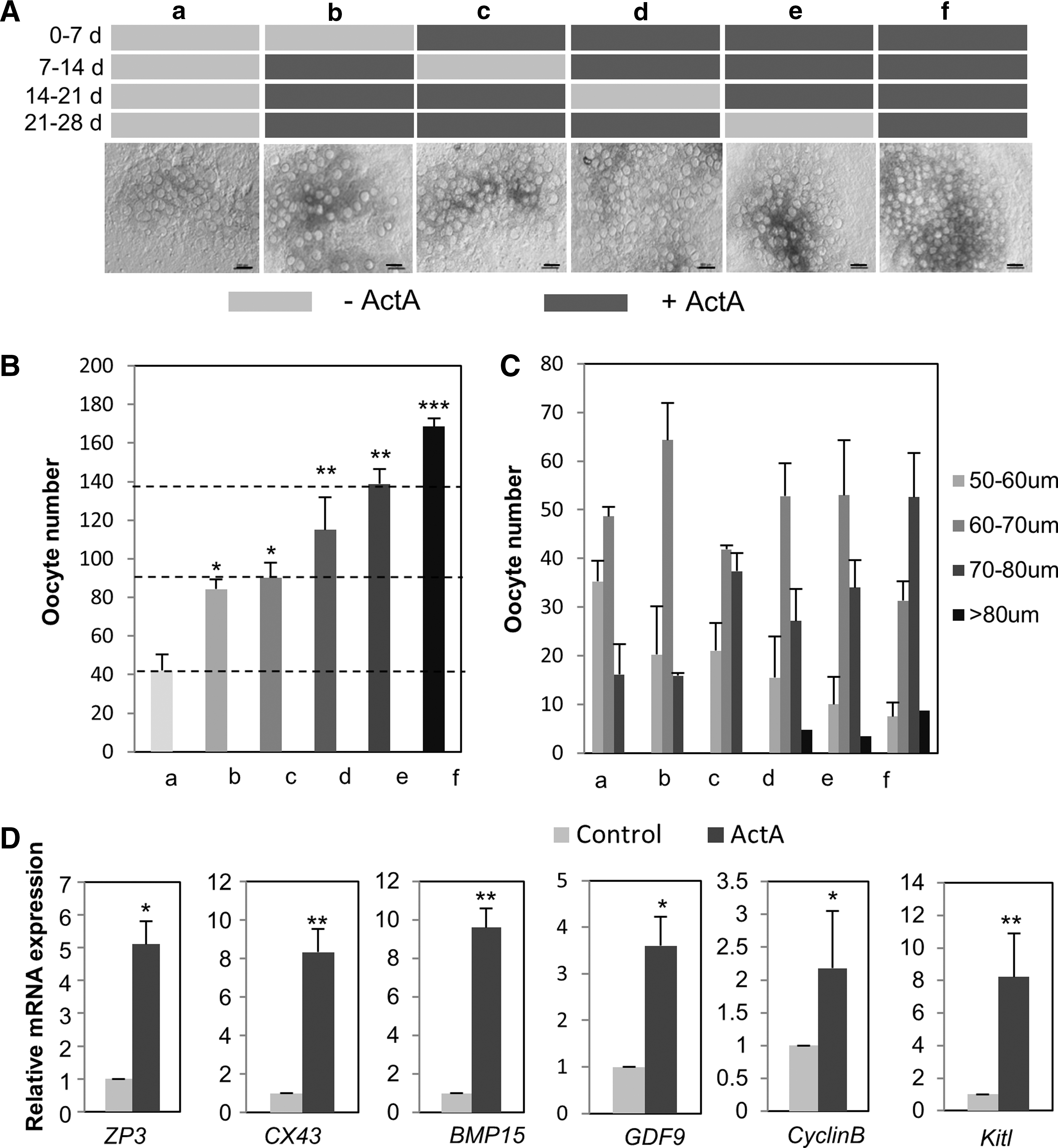
ActA promoted oocyte survival and growth.
ActA promotes primordial follicle assembly in vitro and in vivo
Previous studies have shown that ActA retarded the germ cell nest breakdown and increased the initial primordial follicle pool during the perinatal period in the mouse [6,12]. To determine if ActA also has such effect in our culture system, we maintained the ovarian tissues at 12.5 dpc in the presence of 100 ng/mL ActA for 10 days, a period during which the cultured ovaries should reach developmental stages equivalent to 2–3 days after birth. In tissue sections, ActA-treated ovaries showed a lower number of germ cell nests and higher number of primordial follicles in comparison with the controls (germ cell nests: ActA=27.14%±4.56% vs. control=47.85%±7.97%; primordial follicles: ActA=72.86%±4.56% vs. control=52.15%±7.97%). The ActA effect was abolished by adding the ActA receptor inhibitor, SB431542, to the culture medium (Fig. 6A, B; Supplementary Fig. S4A, B). In ActA-treated ovaries, we also observed some small primary follicles in the center of the ovarian tissues (Supplementary Fig. S4C). The total number of oocytes in the ActA-treated ovarian tissues was higher than that in the control (Fig. 6B). In vivo, a single intraperitoneal injection of ActA (60 and 100 ng/kg) in a female at 10.5 dpc resulted in significant reduction of the germ cell nests harvested from the ovaries at 4 dpp compared with the vehicle control DMSO-treated animals (ActA=36.14%±2.49% vs. control=50.47%±3.51%) and significant increase of primordial follicles (ActA=63.86%±2.49% vs. control=49.53%±3.51%; P<0.05) (Fig. 7).
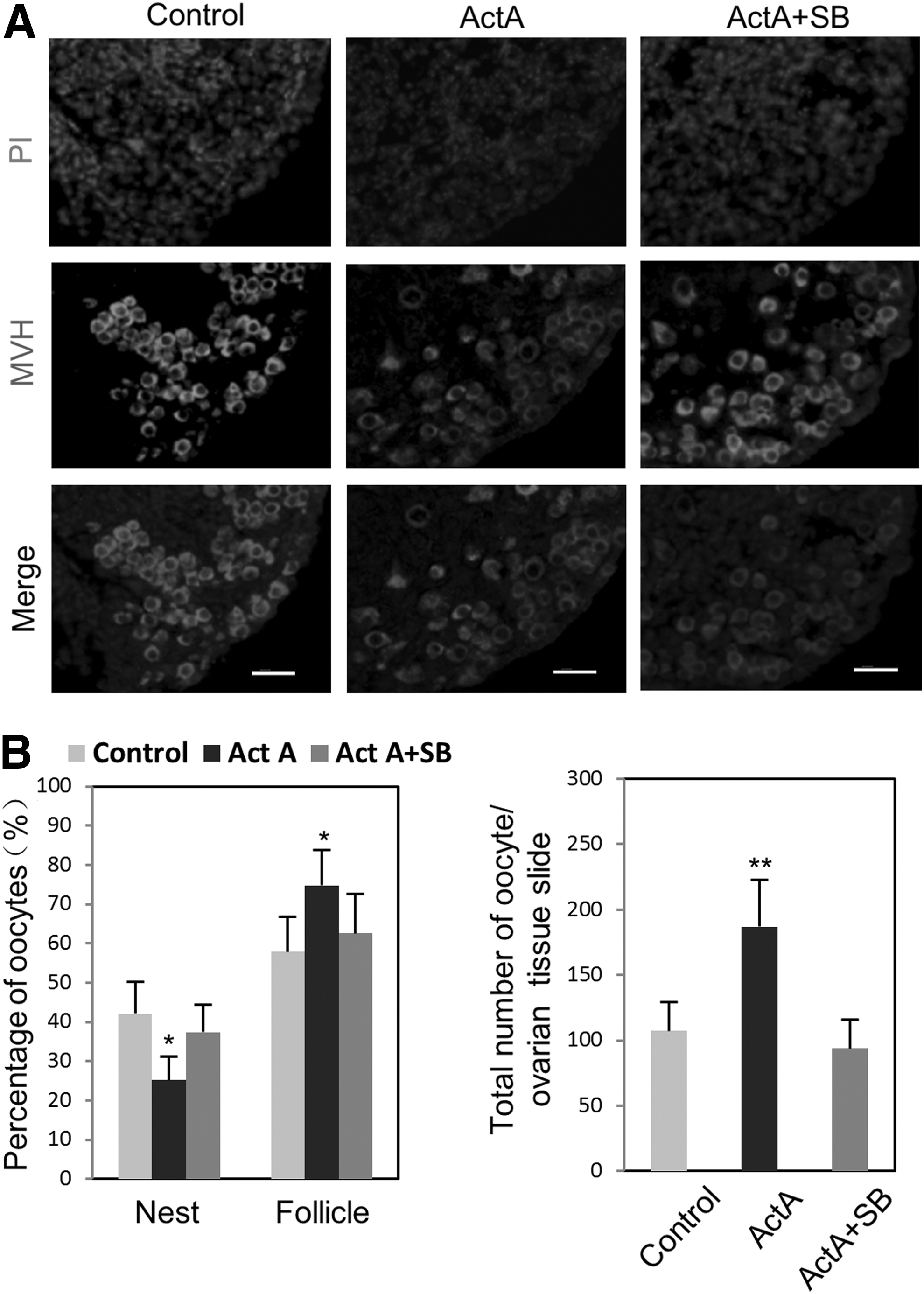
Follicle assembly is promoted by ActA in vitro.
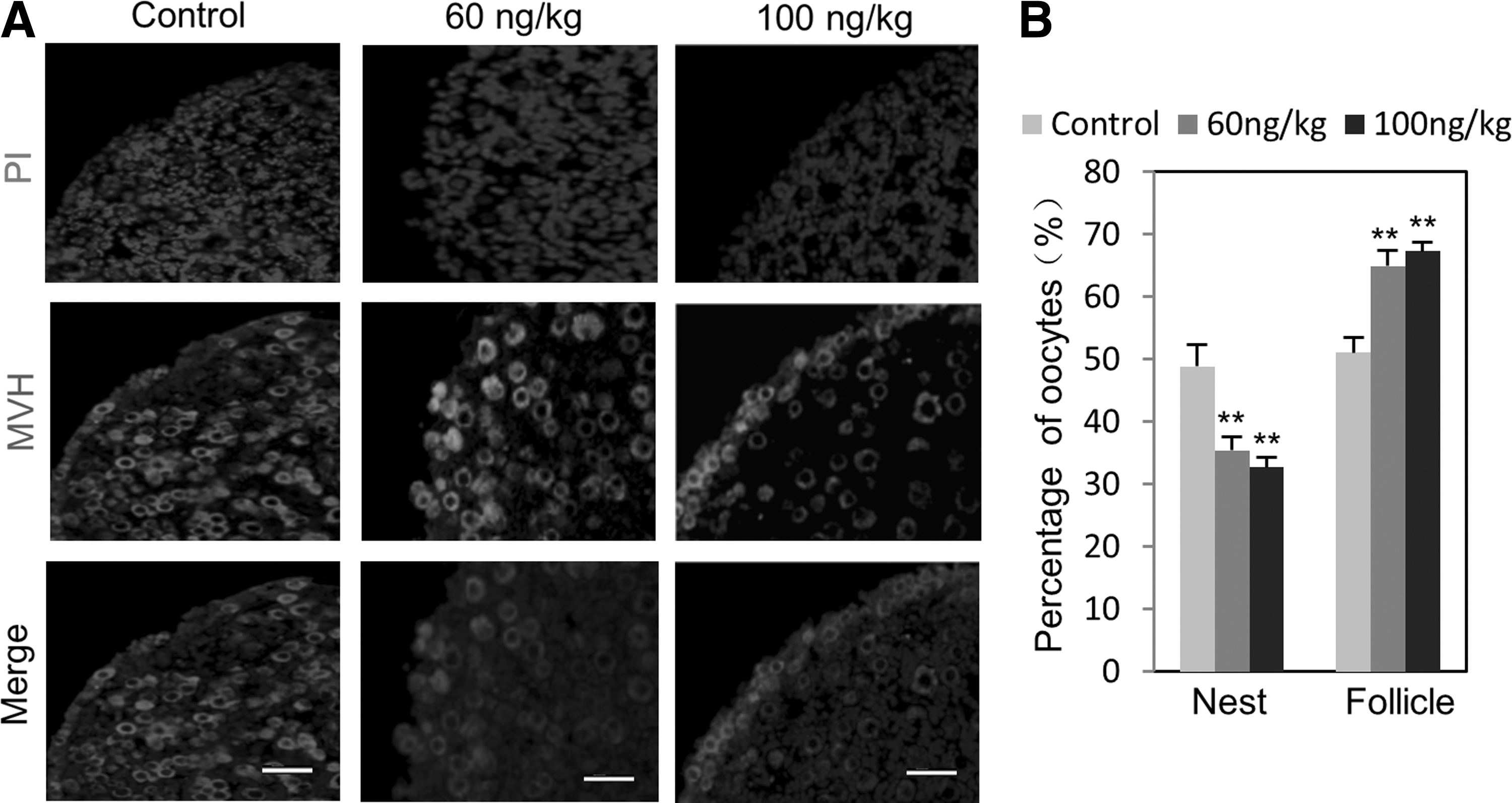
Follicle assembly is promoted by ActA in vivo.
Discussion
In this study, we provided new insights into the roles of ActA during early processes of mammalian oogenesis.
The first evidence about possible ActA actions comes from the findings that both oocytes and somatic cells of the embryonic and fetal mouse gonads express activin receptors. ActA receptors show a significant increase from 12.5 dpc to 7 dpp in mouse ovaries, and all these results coincide with the recognized function of ActA on the meiotic entry of female germ cells, primordial follicle formation. Thus, we paid our attention on the roles of ActA on the meiotic initiation and found that ActA accelerated the progression of the fetal oocytes throughout the meiotic prophase I stages both in vitro and in vivo. Fetal oocytes of mouse ovaries cultured in vitro, progressing throughout the early stages of meiotic prophase I, are slower than that in vivo because in vitro culture conditions still are unable to fully meet the needs of oocyte development. Most interestingly, ActA appears to participate in the control of meiotic progression by modulating the expression of members of the RA system, namely through downregulation of the RA degradation enzyme, CYP26B1, and upregulation of RARs [20,21,29,30]. According to the current more accepted model, female PGCs after arrival at the GRs are induced by the somatic environment, likely through RA, to activate the Stra8 gene [20,31]. STRA8 protein induces the mitotic–meiotic switch and drives female PGCs into meiosis [20,21,29,31]. Other unknown factors are then required for the correct progression throughout meiotic prophase I up to the diplotene stage [29,31].
In the present study, we demonstrated that ActA favors such progression, likely through the activation of SMAD3 expression in pregranulosa cells associated with a parallel decrease of CYP26B1 expression. In line with our results, Kipp et al. suggested that CYP26B1 was a novel target of activin [30]. Future studies are directed to characterize the pathways involved in the SMAD3-associated CYP26B1 downregulation and the mechanisms regulating the oocyte meiotic progression in the fetal ovaries. It should be mentioned that Smad2 and Smad3 expression has been demonstrated in the oocytes and pregranulosa cells of the mid-trimester baboon fetal ovaries [32] and in the pregranulosa cells of the fetal human ovaries [7]. Since a reduction of CYP26B1 likely results in the maintenance of elevated level of RA, our results suggest that RA is necessary not only for the germ cell entering into meiosis but also for its meiotic progression. In a previous article, we showed that Notch signaling also promoted the meiotic progression of the fetal mouse oocytes in culture and that this signaling was involved in the maintenance of an epigenetic state of Stra8 sequence crucial for RA activation [23]. In addition, others have shown that RA is a potent survival factor for mouse fetal oocytes [33] and accelerated the progression into meiosis of rat fetal oocytes [34].
In addition to the effect on oocyte meiosis, ActA also supports the survival of fetal oocytes and the growth of early postnatal oocytes. These results confirmed our previous work [14] and highlighted that these periods are most crucial for such ActA actions. At these stages, ActA may modulate (via autocrine and paracrine pathways) various germ cell activities, including accumulation of GSH in oocytes and the formation of gap junctions and KIT ligand-dependent signals in oocyte/pregranulosa cell complexes [4,6,8,14,35]. Moreover, here and in our previous work [14], we report that ActA may exert the antiapoptotic effect on fetal oocytes through unknown pathways during the primordial follicle assembly.
Finally, our results showing that ActA increased the number of primordial follicles both in vitro and in vivo are consistent with previous studies [6]. However, unlike previous works, we observed that this ActA effect was associated with a reduction of the number of germ cell nests rather than a delay of their breakage. We hypothesize that this apparent discrepancy can be due to the extended period of ovary exposure to ActA performed in the present article. In fact, follicle assembly requires a combination of processes, which partially depends on the genes that are activated during the fetal period. These include genes for growth factors and their receptors [36] and oocyte-specific transcription factors [37] that regulated by ActA might favor meiotic progression and consequently accelerate nest breakage.
In conclusion, the present article identifies new roles of ActA in oogenesis, suggesting that this growth factor and RA might cooperate in promoting meiotic progression in female germ cells and that ActA regulates the expression of genes in fetal oocytes important, besides for their survival and growth, also for their capability to assembly into a follicle.
Footnotes
Acknowledgments
This work was supported by the National Basic Research Program of China (973 Program, 2012CB944401), the National Nature Science Foundation (31171376 and 31471346), the Program for New Century Excellent Talents in University (NCET-12-1026), and the Nature Science Foundation (ZR2013CQ029) the Taishan Scholar Construction Foundation of Shandong Province.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.