
Abstract
Cardiovascular disease is the leading cause of death in the United States. Heart failure is a common, costly, and potentially fatal condition that is inadequately managed by pharmaceuticals. Cardiac repair therapies are promising alternative options. A potential cardiac repair therapy involves reprogramming human fibroblasts toward an induced cardiac progenitor-like state. We developed a clinically useful and safer reprogramming method by nonintegrative delivery of a cocktail of cardiac transcription factor-encoding mRNAs into autologous human dermal fibroblasts obtained from skin biopsies. Using this method, adult and neonatal dermal fibroblasts were reprogrammed into cardiac progenitor cells (CPCs) that expressed c-kit, Isl-1, and Nkx2.5. Furthermore, these reprogrammed CPCs differentiated into cardiomyocytes (CMs) in vitro as judged by increased expression of cardiac troponin T, α-sarcomeric actinin, RyR2, and SERCA2 and displayed enhanced caffeine-sensitive calcium release. The ability to reprogram patient-derived dermal fibroblasts into c-kit+ CPCs and differentiate them into functional CMs provides clinicians with a potential new source of CPCs for cardiac repair from a renewable source and an alternative therapy in the treatment of heart failure.
Introduction
R
C-kit is a cell surface marker expressed on a subset of CPCs, and these c-kit+ CPCs possess inherent stem cell properties that promote regenerative capacity [8]. In fact, recent studies have demonstrated promising results using autologous c-kit+ CPC therapy on patients with chronic ischemic heart failure [9], and future phase I clinical trials are being planned to use c-kit+ CPCs in combination with MSC transplantation [10]. Current clinical trials, such as CADEAUS and CHART-1 [11], have displayed a positive benefit to the patients suffering from heart failure undergoing cell therapy by demonstrating an improvement in quality of life. This was assessed using two tests: demonstration of an increase in meters walked during the 6-min walk test and improvement in quality-of-life score using the Minnesota Living with Heart Failure Questionnaire (MHLHFQ), pretherapy and post-therapy [10]. Unfortunately, the effective use of autologous c-kit+ CPC therapy has major limitations: (1) CPC isolation from heart is a laborious and time-consuming process with only a 65%–70% success rate and (2) c-kit+ CPCs are limited and represent only ∼3%–5% of cells found in the heart. Therefore, the development of methods to generate a sufficient number of c-kit+ CPCs remains an important clinical need.
To address this unmet clinical need for CPCs, we propose an innovative, nonintegrative RNA-based approach to reprogram fibroblasts into c-kit+ CPCs using an mRNA cocktail of transcription factor-encoding RNAs or isolated RNAs from the CPC transcriptome. Previous studies using transcription factors delivered by either viral vectors [12,13] or by lipid-based delivery systems [14,15] have successfully demonstrated the ability to reprogram somatic cells into a differentiated cell type. Two major concerns in translating viral-based transduction of transcription factors from the laboratory to the clinic are the risk of random genomic integration of the transgenes into the human genome and the inability to control viral overexpression of transgenes. Researchers are currently trying to identify new cell differentiation protocols that utilize nonintegrative strategies [15,16] or small molecules [17]. In addition, lipid-based delivery systems are limited by scale-up for clinical use and transfection agent toxicity. These limitations suggest that using transcription factor-encoding RNA cocktails through electroporation would facilitate the scale-up of induced CPC (iCPC) generation and provide proof-of-principal that primary human somatic cells are amenable to reprogramming/differentiation.
Therefore, the main objective of this study was to determine the reprogramming potential of primary adult human dermal fibroblasts (fibroblastadult) into cardiomyocytes (CMs) using RNA cocktails delivered through electroporation. Our results demonstrate that (1) different mRNA cocktails were able to reprogram primary adult fibroblasts into iCPCs as determined by iCPC expression of c-kit, Isl-1, and Nkx2.5, and (2) iCPCs differentiated into a CM-like state in vitro with functional characteristics following c-kit selection. The ability to use patient-derived dermal fibroblasts and to reprogram them in vitro into functional c-kit+ CPCs would provide clinicians with a renewable source of CPCs for cellular implantation to mediate cardiac repair in the treatment of heart failure.
Materials and Methods
Cells and media
Neonatal human dermal fibroblasts (fibroblastneo) were obtained from Gibco. Anonymized adult human dermal fibroblasts (fibroblastadult) were kindly provided by Dr. Jennifer Bond and Dr. Howard Levinson. Neonatal and adult fibroblast cells were initially cultured in low-glucose Dulbecco's modified Eagle's medium (DMEM). CPCs were obtained commercially from Celprogen. CPCs were grown on fibronectin-coated petri dish (BD Bioscience) in CPC media (Celprogen) at 1 × 106 cells in a 100-mm plate. C-kit+ CPCs from the right atrium were obtained from anonymized human right atrial tissue using protocols established by Dr. Marie-Jose Goumans [18] and Dr. Steven Houser's laboratories using human CD117 MicroBead kit (MACS; Militenyi Biotech) [19]. Right atrium of the heart was digested in a tissue digestion medium composed of collagenase (319 U/mL) and 0.1% Trypsin-EDTA. The human cardiac stem cell (hCSC) medium contained DMEM-F12 (Biowittaker) supplemented with 10% embryonic stem cell-qualified fetal bovine serum (FBS; Gibco), pen/strep/glutamine, 10 ng/mL human fibroblast growth factor-basic (bFGF; Peprotech), 0.2 mM
Construction of in vitro transcription templates
The oligonucleotide sequences used in the construction of in vitro transcription templates are shown in Supplementary Table S1 (Supplementary Data are available online at
Synthesis of mRNAs for mRNA cocktails and RNA extraction
RNA was synthesized with T7 mScript™ Standard mRNA production system (Cellscript, Inc.) with 1 μg of purified linearized plasmid in each 20 μL reaction. For modified mRNA transcription reactions, NTP solution was replaced with a modified NTP solution comprised of ATP, GTP, 5-methylcytidine triphosphate, and pseudouridine triphosphate (5mCTP and ΨTP were both from Trilink Biotechnologies, Inc.). Transcription reactions were incubated for ∼2 h at 37°C. Synthesized mRNAs were capped with cap-1 using ScriptCap™ 2′-O-methyltransferase at 37°C for ∼1 h and 3′-poly(A) tail was added using A-Plus™ Poly (A) Polymerase (Cellscript, Inc.) at 37°C for 30 min. After each synthesis step of mRNA production, the reactions were cleaned with RNA purification columns (Macherey Nagel). RNA was quantified using a Nanodrop 1000 (ThermoScientific). RNA transcriptome of the CPCs (Celprogen) was isolated using the mirVana™ miRNA isolation kit (Ambion) according to the manufacturer's instructions.
RNA transfection and electroporation
RNA transfections were performed using RNAiMAX (Invitrogen), Trans-IT (MirusBio), or JetPEI (Polyplus) as cationic lipid delivery vehicles for mRNA. For Trans-IT, RNAimax, and JetPEI transfections of mRNAs, we followed the manufacturer's recommended protocol based on RNA concentration. For lipid-based transfections, ∼400 ng modified (5mCΨ) mRNA cocktail [0.5:1:1:1:1; GFP:Gata4:Mef2c:Tbx5:Hand2 (G-GMTH)] per well was incubated with the designated transfection reagent in Opti-MEM and was placed onto 20,000 fibroblastneo cells/well in DMEM-LG/10% FBS. GMTH transfection treatments were performed four times (4×) to each well and treatments were performed every 2 days. After a series of four treatments, the cells were stained for cardiac troponin T (cTnT; Abcam) and the RNA was extracted for quantitative real-time PCR (qt-RT-PCR) analysis using the NucleoSpin RNA Clean-up kit (Macherey-Nagel).
Electroporation of RNA was performed with 9–10 μg mRNA cocktail (G-GMTH at 0.5:1:1:1:1) diluted in 25 μL Opti-MEM and fibroblastneo cells (1.0 × 106) diluted in 75 μL Opti-MEM for a total volume of 100 μL in electroporation cuvette (1 mm) for 250 mV, 1 μs. Electroporated fibroblastneo cells were transferred to hCSC media and incubated for 2 days. The treatment was repeated three to five times (5×) for a total of four to six electroporation treatments with mRNA cocktails.
Cardiac differentiation assay
CPCs (Celprogen), mock-treated fibroblastadult, and fibroblastadult electroporated with mRNA cocktail were plated on fibronectin-coated plates (BD) at 10,000 cells/cm2 in hCSC media [22]. Once cells reach >80% confluency, the media were changed to the differentiation media [DMEM-F12, glutamax I, insulin-transferrin-selenium (Life Technologies), 5% horse serum, MEM-nonessential amino acids, and pen/strep]. After cells were incubated for 6–8 h with the differentiation media (DM), the media were changed to fresh media containing 5 μM 5-azacytidine (5-AZ). The media were changed for 2 consecutive days with DM containing fresh 5-AZ at 5 μM. On day 4, the media were changed to fresh media without 5-AZ. Six days after the 5-AZ stimulation began, the following chemicals were added to the DM: 1× ascorbic acid (AA; Gibco, 500× stock) and 1 ng/mL transforming growth factor β (TGF-β; Peprotech) [22]. Further differentiation was conducted by changing the media with the addition of fresh AA to DM every 2 days and adding fresh TGF-β to DM every 3 days. Cardiac differentiation assays were conducted for a total of 25 days.
Quantitative real-time PCR
RNA was extracted from mRNA-treated fibroblasts using the Nucleospin RNA Clean-up (Macherey-Nagel) kit or Trizol (Ambion) [23], and genomic DNA was digested with DNase I (Roche). cDNA was generated from the mRNA using 500 ng of total RNA in a 20 μL reaction volume of the iscript cDNA assay (Biorad) following manufacturer's protocol. cDNAs (2 μL) were examined by qt-RT-PCR using SYBR green assay kit (Biorad) in a final volume of 24 μL in a CFX96 Thermocycler (Biorad). Primers used are described in Supplementary Table S2.
Immunostaining and immunofluorescence
Isolated c-kit+ cells, CPCs, and treated cells were plated on fibronectin-coated glass coverslips in hCSC media overnight. The staining protocol was followed as described by Kubo et al. for c-kit+ immunostaining using CD117 antibody (DAKO A4502) [24]. Immunostaining of transfected fibroblastadult cells was performed on the cells that were fixed with 4% paraformaldehyde at room temperature for 15 min. Fibroblastadult treated cells were blocked with a blocking buffer [1× phosphate-buffered saline (PBS), 5% normal goat serum, 0.30% Triton X-100] at room temperature for 60 min. Cells were incubated with the primary antibody against cTnT (Abcam; 1:400) or sarcomeric α-actinin (Abcam; 1:400) overnight at 4°C. Cells were rinsed with 1 mL PBS, three times at 5-min intervals each. The specimens were incubated with secondary antibody at 1:500 for AF488-rabbit IgG and 1:400 for anti-goat AF594-mouse IgG for 1–2 h at room temperature in the dark. Cells were rinsed with 1 mL PBS, three times at 5-min intervals, and cells were incubated with Fluoromount-G/DAPI at 1:1000. Microscope slides were examined on an inverted Zeiss 510 confocal microscope at Duke Microscopy Core Facility.
Flow cytometry
Control cells (fibroblasts and CPCs) and mRNA-treated fibroblasts were incubated with Tryp-LE Express for 5 min. Approximately 1 × 106 cells were used for each sample. Cells were fixed in 1% paraformaldehyde for 10 min at 37°C in the dark and permeabilized with ice-cold 90% methanol for 30 min on ice. Cells were washed with the FACs buffer (PBS, 5% bovine serum albumin, 0.1% Triton X-100). Next, cells were incubated with primary antibody at 1:100 for cTnT (Abcam) or 1:200 dilution for α-actinin (Abcam) overnight at 4°C. Cells were washed three times with 1 mL FACs buffer and then incubated with secondary antibody of anti-goat AF488 rIgG at 1:200 or anti-goat PE mIgG at 1:200 at 4°C overnight. Cells were finally washed with 1 mL of FACS buffer three times and resuspended in 0.3 mL FACs buffer for analysis.
Calcium release experiments
After the cardiac differentiation assay, treated cells were plated in a black 96-well plate at 10,000 cells/well in triplicate. Cells were washed with Tyrode's solution (140 mM NaCl, 4.7 mM KCl, 1.6 mM MgCl2, 10 mM HEPES, and 5 mM glucose) and incubated with 2 μM Fluo-4-AM (Life Technologies) in 100 μL Tyrode's solution for 45 min at 37°C. Cells were washed twice with Tyrode's solution and left in Tyrode's solution for 15 min. Calcium release was examined in the presence of 10 mM caffeine and the absence of caffeine as a stimulant. Cells were examined kinetically in a SpectraMax i3 (Molecular Devices) at an excitation of 488 nm and emission spectrum of 520 nm with photomultiplier tube (PMT) of 15 flashes/read [25,26].
Results
Electroporation provides optimal mRNA delivery compared to lipid transfection
Initial experiments were conducted to determine the efficiency and effectiveness of nuclear mRNA delivery. mRNA cocktails were delivered into neonatal human dermal fibroblasts (fibroblastneo) through nonintegrative methods of either lipid-based transfection (LT) or electroporation of mRNA (EP) to determine the most optimal method for reprogramming human fibroblasts into either iCPCs or induced CMs (iCMs). mRNA delivery experiments used a series of mRNA cocktails generated from Gata4 (G), Mef2c (M), Tbx5 (T), and Hand2 (H) mRNAs, which have been used in previous reprogramming attempts [27]. In our study, GMTH contained unmodified RNA nucleotides (A,C,G,U) or modified RNA nucleotides (A, 5mC, G, Ψ), substituting 5-methyl cytidine (5mC) for cytidine (C) and pseudouridine (Ψ) for uridine (U). For verification of the efficiency and effectiveness of mRNA cocktail delivery, GMTH and modified GMTH (5mCΨ) in combination with GFP mRNA were first delivered as an mRNA cocktail combination into neonatal fibroblasts through lipid transfection or electroporation to verify mRNA delivery of the cocktails (Supplementary Fig. S1A, S1B). To determine the optimal mRNA delivery method, we examined and confirmed mRNA expression of the transfected neonatal fibroblasts for known cardiac transcription factors Gata4, Tbx5, and Hand2 in the GMTH mRNA cocktail by qt-RT-PCR (Fig. 1A–C). Intracellular staining of cells following lipid transfection or electroporation with mRNA cocktails confirmed protein expression of the transcription factors GMTH in the nucleus of the treated neonatal fibroblasts (Supplementary Fig. S2A, S2B).
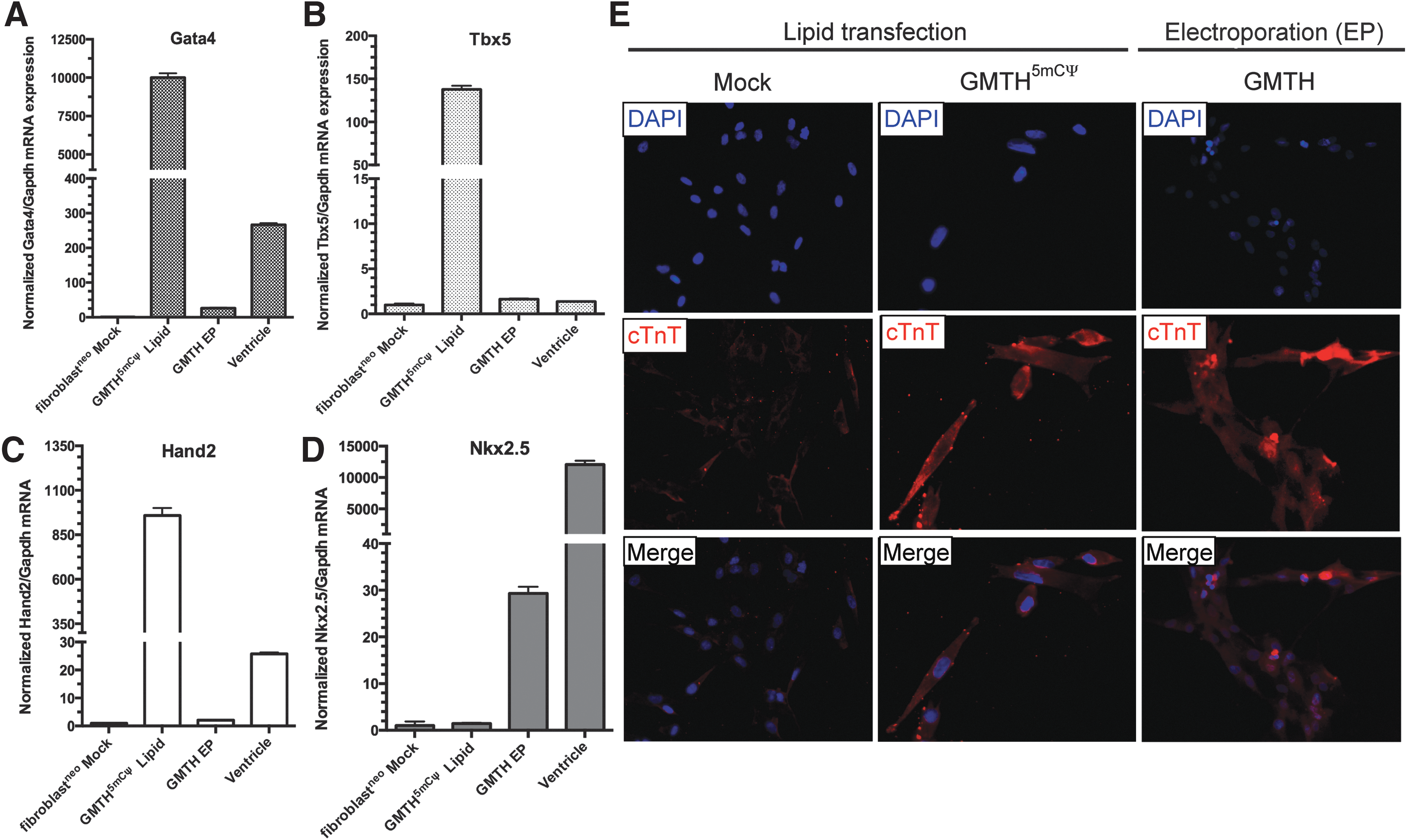
GMTH lipid transfection (LT) versus GMTH electroporation (EP). RNA-mediated reprogramming of modified GMTH (5mCΨ) with lipid-based transfection reagents (Trans-IT) versus electroporation (EP) for GMTH transcription factors. Quantitative real-time polymerase chain reaction (qt-RT-PCR) analysis of cardiac transcription factor expression of Gata4
We assessed the ability of the unmodified GMTH mRNA cocktail to partially reprogram neonatal fibroblasts into iCPCs by examining the upregulation of an endogenous early cardiac transcription factor associated with CPCs known as Nkx2.5, Nkx2.5 mRNA expression increased ∼20-fold after four consecutive electroporation treatments with unmodified GMTH mRNA cocktail compared to four consecutive treatments with modified GMTH (5mCΨ) mRNA cocktail through lipid transfection (Fig. 1D). The use of modified GMTH (5mCΨ) mRNA was required for lipid transfection, since multiple treatments of unmodified GMTH mRNAs delivered to fibroblasts result in increased cell death and inflammation based on Warren et al. [15] and our own experience.
Unmodified RNAs delivered by lipid formulations are recognized by pattern recognition receptors (PRR) in the cytoplasm, such as RIG-I and PKR, which can elicit an interferon response (IFNA) resulting in the inhibition of protein translation and cellular toxicity [28,29]. We examined whether the nonintegrative methods, electroporation or lipid transfection, induces an inflammation response by examining the mRNA expression of IFNA, RIG-I, and PKR. Lipid transfection of neonatal fibroblasts resulted in the upregulation of both RIG-I (Fig. 2B) and PKR (Fig. 2C) even with the incorporation of modified nucleotides (5mCΨ), when compared to both mock-treated cells and neonatal fibroblasts electroporated with unmodified GMTH cocktail. After four successive treatments of lipid-transfected-modified GMTH (5mCΨ) mRNA cocktail or four successive treatments of electroporated GMTH mRNA cocktail, we observed an upregulation of IFNA mRNA and the induction of an inflammation response; however, lipid-transfected neonatal fibroblasts produced ∼2.5-fold more IFNA mRNA (Fig. 2A) than electroporated cells.
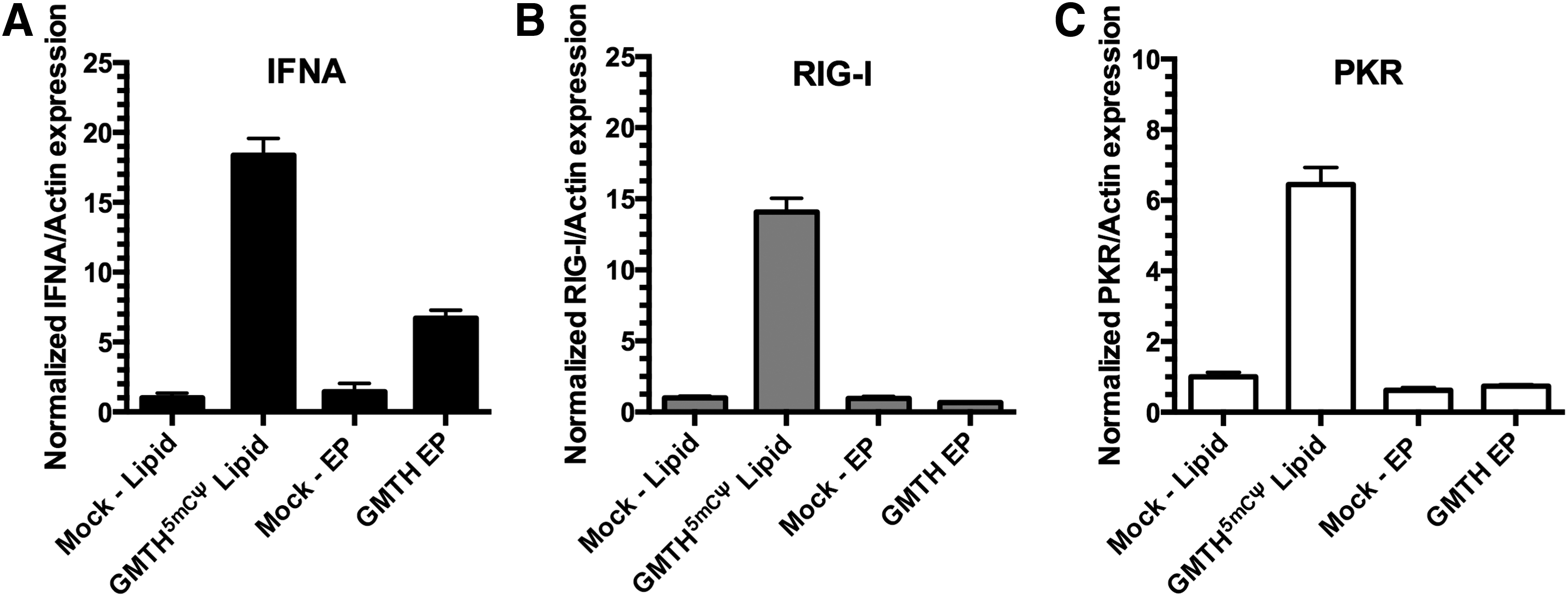
Inflammation-associated genes in neonatal fibroblasts after lipid transfection or electroporation with mRNA. Quantification of inflammation genes [IFNA
When the expression of cardiac marker cTnT was examined by immunofluorescence (Fig. 1E); both lipid-based transfection and electroporation of the GMTH mRNA cocktails resulted in an increased cTnT expression in treated neonatal fibroblasts. Although neonatal fibroblast lipid transfected with GMTH (5mCΨ) were positive for cTnT expression (Fig. 1E), we decided not to pursue this method, because the overall treatment protocol required for the modified mRNA (5mCΨ) cocktail resulted in greater cell death and increased activation of PRRs (Fig. 2), which would not be conducive for future scale-up compared to electroporation. Together, the qt-RT-PCR and immunofluorescence data suggest that relative to lipid-based transfection, electroporation is a superior nonintegrative mRNA transfection method for cellular reprogramming of neonatal fibroblasts.
Adult fibroblasts and GMTH is an optimal combination for reprogramming to iCPCs
After determining that electroporation is the optimal method to deliver mRNAs into the fibroblasts, we examined (1) the best somatic cell type (starting material) for reprogramming and (2) the preferred mRNA cocktail for reprogramming somatic cells into a cardiac progenitor-like cell state. The initial somatic cell types used for comparing reprogramming capabilities were neonatal human dermal fibroblasts (fibroblastneo) and primary adult human dermal fibroblasts (fibroblastadult). Reprogramming experiments were conducted in parallel utilizing the RNA cocktails: unmodified GMTH mRNA, modified GMTH (5mCΨ) mRNA, or RNA derived from the CPC transcriptome (CPC RNA).
Fibroblastsadult required four cycles of electroporation with GMTH mRNA cocktail and six cycles of electroporation with CPC RNA transcriptome to achieve similar morphological characteristics and partial reprogramming toward a cardiac progenitor-like state. Twenty-four hours after the last treatment, the morphological state was evaluated by monitoring expression of the progenitor cell marker c-kit by immunofluorescence staining (Fig. 3A), followed by verification using qt-RT-PCR (Fig. 3B). In addition, the reprogramming of adult fibroblasts into iCPCs was confirmed by examining the mRNA expression of other well-known progenitor markers and early cardiac markers Isl-1, Gata4, and Nkx2.5 (Fig. 3B, C). After treatment with the various mRNA cocktails, the c-kit expression was increased in the adult fibroblast-derived iCPCs to expression levels that were comparable to commercially available CPCs (Fig. 3A). iCPCs derived from fibroblastsadult displayed a qualitatively lower level of c-kit expression on the cell surface compared to commercially available CPCs (Fig. 3A), but exhibited a markedly better c-kit expression compared to CPCs extracted from the right atrium (Supplementary Figs S3B and S4A). To quantitatively assess differentiation efficiency, the c-kit expression on fibroblastadult iCPC's was determined using the MACs c-kit+ cell isolation kit following GMTH mRNA transfection; c-kit+ cells represented ∼4.9% of the total cell population (2 × 106 cells). Furthermore, mock-treated neonatal fibroblasts demonstrated an innate higher expression of progenitor markers when compared to electroporated mRNA-treated neonatal fibroblasts (Supplementary Fig. S4A, S4B). Mock-treated neonatal fibroblasts were exposed to both electroporation and conditional media. The observed higher expression of progenitor markers in mock-treated neonatal fibroblasts made it difficult to determine how effective the mRNA cocktails were at inducing a cardiac progenitor-like state (Supplementary Fig. S4B). RNA cocktail derived from the CPC transcriptome, similar to that previously used to generate murine CMs [16], was electroporated six times into adult fibroblasts, but the c-kit and Isl-1 were expressed at low levels; however, when adult fibroblasts were isolated for the c-kit expression by MACS separation column, an 18-fold increase in the Isl-1 expression and ∼2.63-fold increase in the c-kit expression by qt-RT-PCR were observed (Supplementary Fig. S5). Overall, the electroporation of fibroblastadult was determined to be an optimum method for differentiation because of the significant upregulation of specific progenitor genes associated with iCPCs, namely c-kit and Isl-1 (Fig. 3B), and the increased expression of c-kit on the cell surface (Fig. 3A) compared to mock-treated fibroblastadult and electroporated mRNA-treated fibroblastneo (Supplementary Fig. S4A).
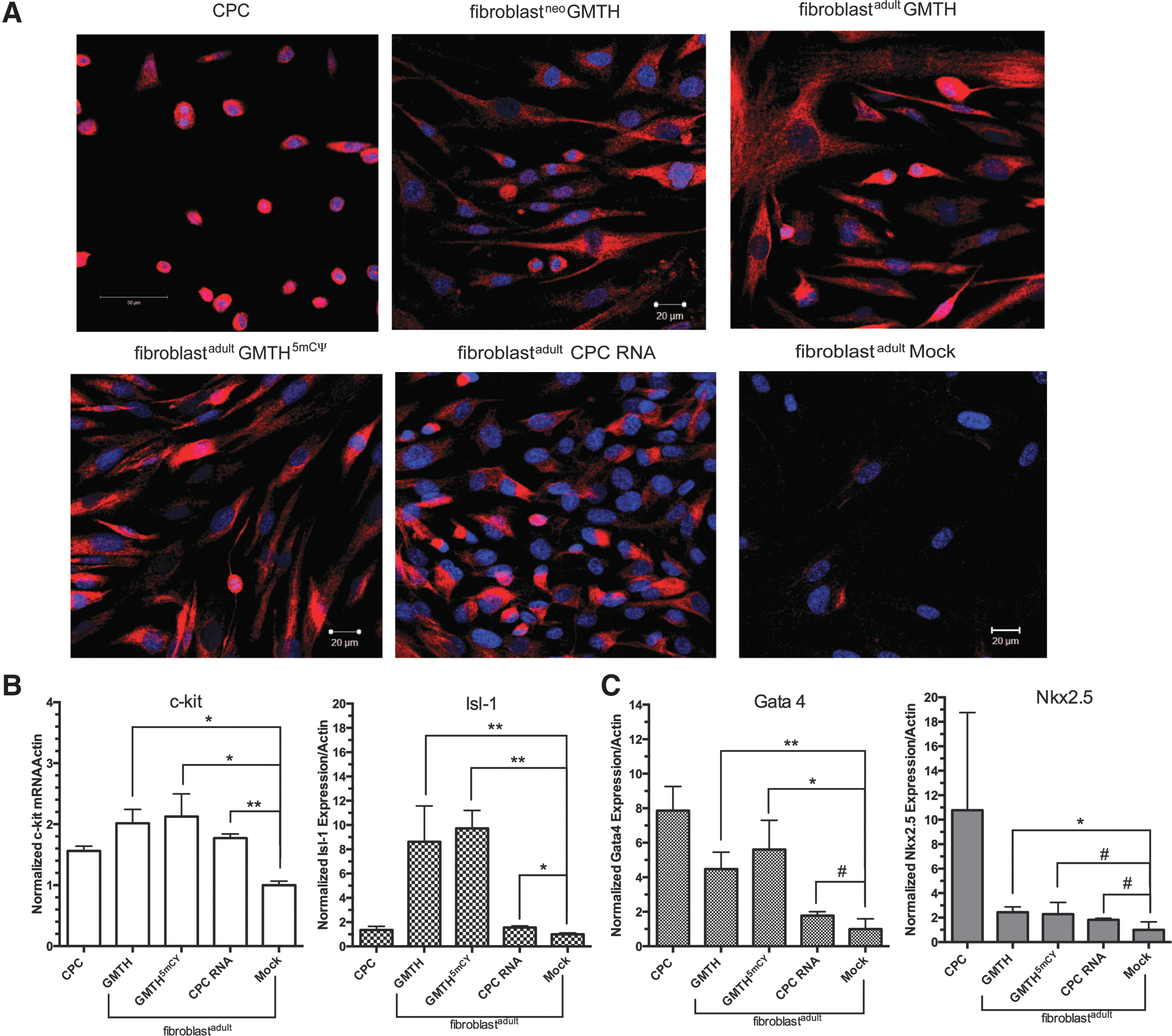
Assessing reprogramming of fibroblasts into induced cardiac progenitor cells (iCPCs) by c-kit analysis.
c-kit+ GMTH-stimulated iCPCs have the capacity to differentiate into iCMs
iCPCs electroporated with the different RNA cocktails (GMTH, GMTH (5mCΨ), and CPC transcriptome) expressed cardiac genes associated with cardiac predisposition–Gata4, Nkx2.5, and cTnT (Figs. 3C, 4A, and 4B). To determine if these iCPCs had the potential to differentiate into a CM-like state and express the CM-specific genes cTnT, α-sarcomeric actinin, and SERCA2, we used an established in vitro protocol for efficiently differentiating CPCs into CMs; CPCs were stimulated with 5-AZ, TGF-β, and AA [22]. Following 5-AZ treatment, all treated adult fibroblast exhibited partial reprogramming and expressed Nkx2.5 and cTnT mRNA compared to mock-treated adult fibroblasts (Fig. 4A, B). Moreover, the GMTH and 5-AZ treatment afforded the greatest increase in cTnT and α-actinin expression, as confirmed by immunofluorescence (Fig. 5A), and the greatest increase in percent differentiation of the cell population as monitored by flow cytometry (Fig. 5B, C). The percentage of total differentiated cells derived from GMTH adult fibroblasts that expressed cTnT or α-actinin was 20% and 7%, respectively. Because commercially available CPCs express 25% cTnT and 12% α-actinin, our data suggest that iCPCs had a similar potential to differentiate into the CM state relative to commercially available CPCs (Fig. 5B, C). CM-specific genes translate into proteins that can localize into structurally distinct striations. We, therefore, examined the presence of striations in iCMs derived with the GMTH mRNA cocktail. Clearly defined and compact striations with distinct sarcolemma edges existed in c-kit+ GMTH-derived CMs compared to the mock-treated fibroblasts or commercial CPCs. Furthermore, we observed a rod-like morphology (Fig. 6A), which is consistent with 100-μm length and 25-μm width typical for adult CMs based on their gross morphology.
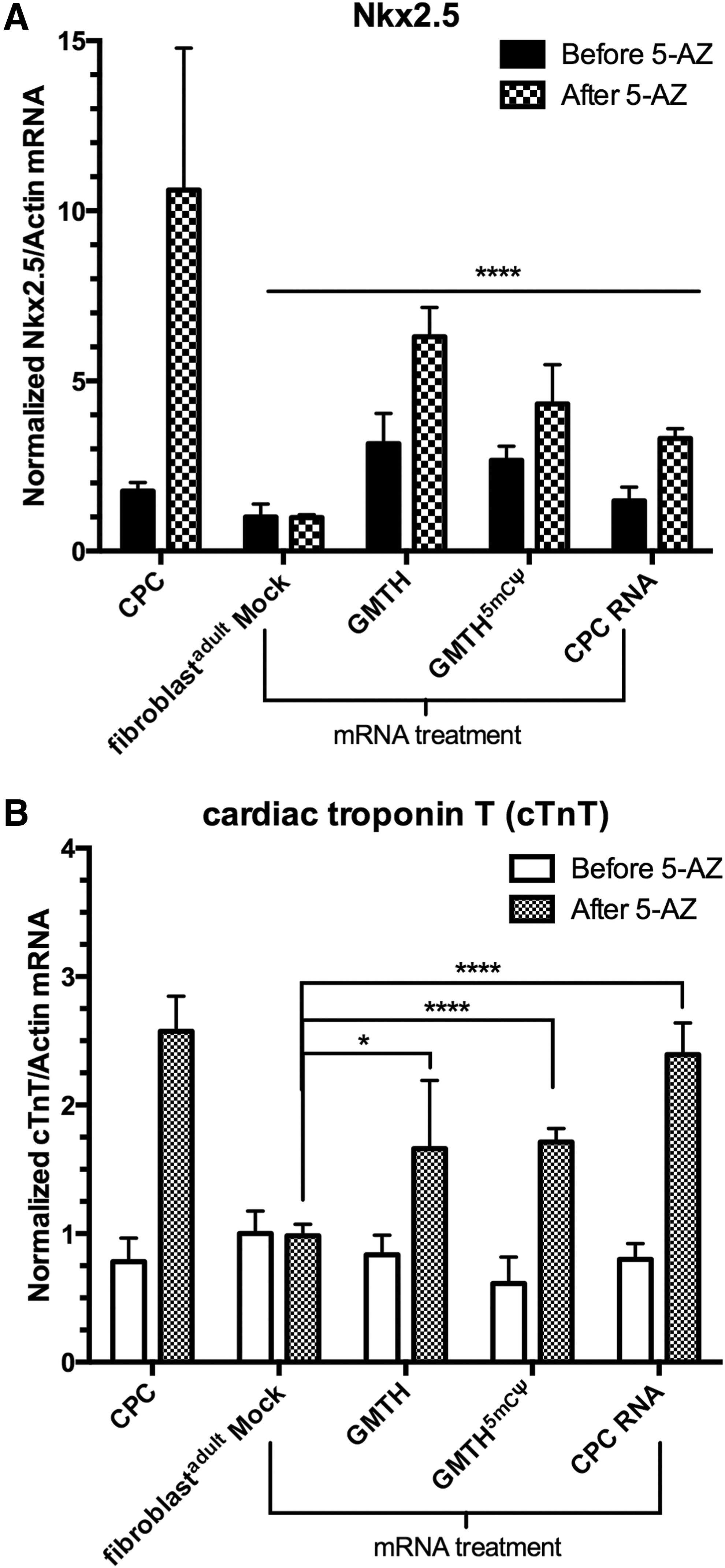
qt-RT-PCR analysis of mRNA-treated fibroblastadult cells before and after treatment with 5-azacytidine (5-AZ). Intermediate and late cardiac markers,
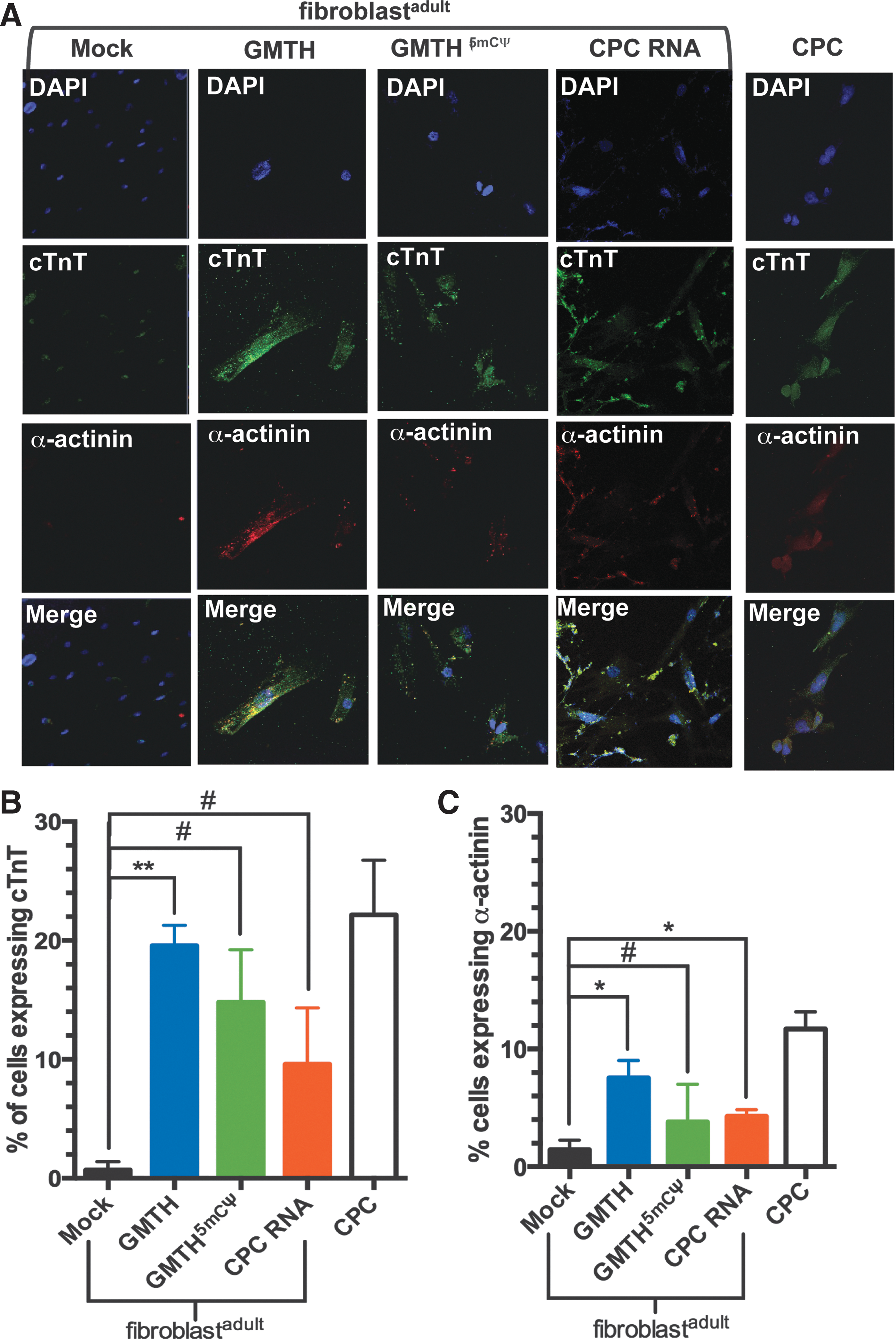
Cardiomyocyte (CM) differentiation of iCPCs and analysis of cardiac markers on day 25 post-mRNA treatment.
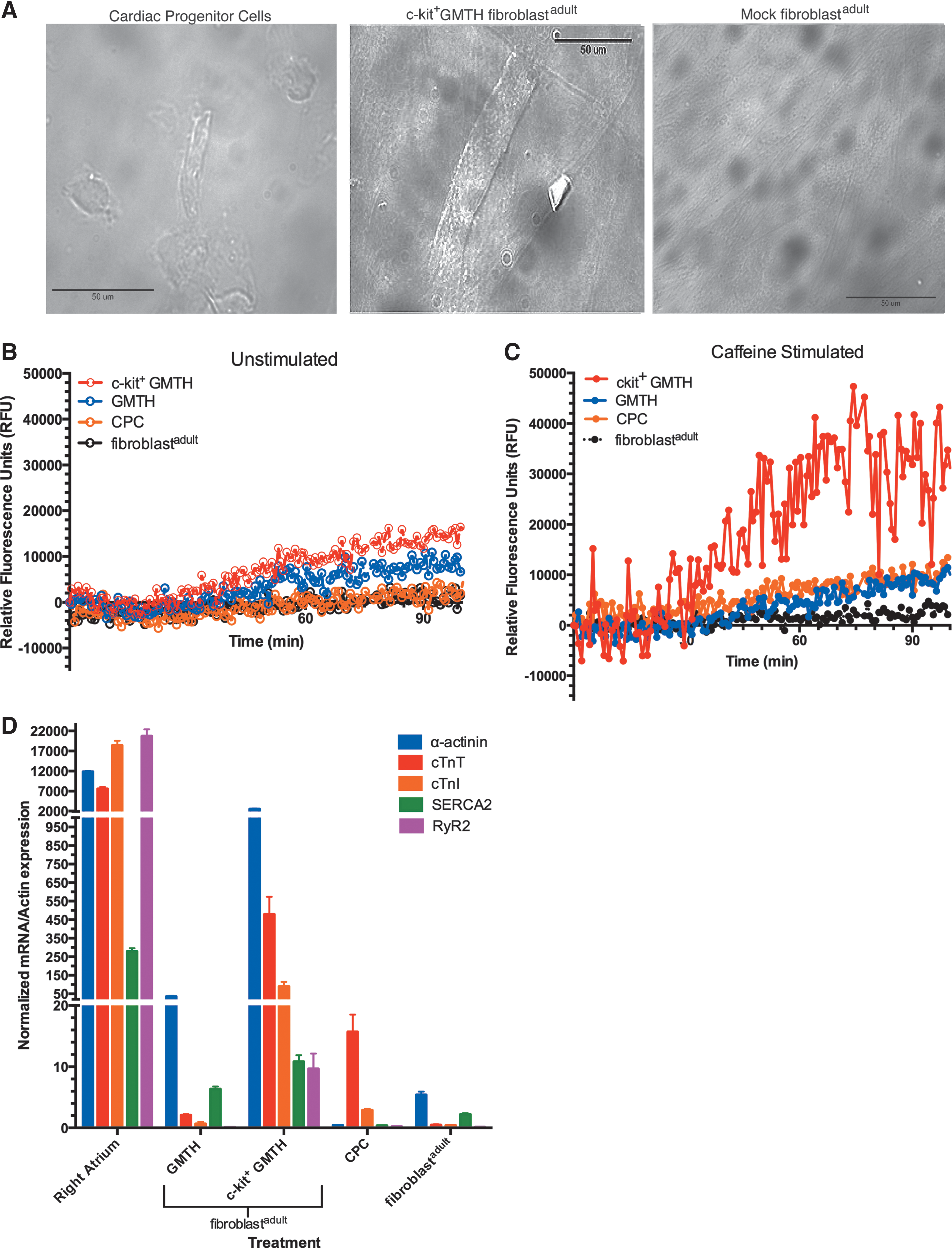
Assessing the structure and cardiac function of induced CMs (iCMs).
In addition to CM-specific gene expression, we evaluated the functional capacity of the iCMs to cycle calcium. Fibroblastadult selected for the c-kit expression after being treated with unmodified GMTH mRNA cocktail displayed the greatest activity in releasing calcium and was the only treatment to respond to caffeine stimulation (Fig. 6B, C). In fact, caffeine enhanced the release of calcium in these c-kit+ GMTH-treated cells to levels higher than those observed in unselected GMTH-treated cells, cardiac precursor cells, and mock fibroblast control (Fig. 6C). Interestingly, the functional activity of the c-kit+ GMTH adult fibroblasts corresponded with >10-fold-increased expression of SERCA2 and cardiac ryanodine receptor 2 (RyR2) when compared to unmodified GMTH adult fibroblasts and adult fibroblasts alone (Fig. 6D). Upregulation of SERCA2 and RyR2 expression in these cells correlates with the enhanced ability of the treated cells to release Ca2+ during stimulation [30,31]. These data demonstrate that fibroblastadult electroporated with GMTH mRNA can be reprogrammed to a CPC-like state, and these cells selected for c-kit expression are able to differentiate into a CM-like state after treatment with 5-AZ. Future studies are needed to evaluate the full functional potential of the iCMs derived from c-kit+ GMTH-treatment, for example patch clamp, assessment of paracrine effects, and in vivo models to monitor the iCPC development.
Discussion
The reprogramming capacity of murine dermal fibroblasts to differentiate into functional CMs both in vitro and in vivo has been elegantly demonstrated by Ieda et al. [32] and Qian et al. [33]. This evidence has provided support for dermal fibroblasts as a fruitful source for CM induction; however, minimal evidence has supported the use of human dermal fibroblasts over cardiac fibroblasts, and the optimal method for reprogramming and differentiation of dermal fibroblasts has not yet been fully explored. The main objective of our study was to determine the reprogramming potential of primary adult human dermal fibroblasts (fibroblastsadult) to differentiate into CMs using nonintegrative delivery methods for RNA cocktail delivery. The major findings of our study are that primary adult human dermal fibroblasts electroporated using unmodified mRNA were reprogrammable into a cardiac lineage, and these induced cardiac precursor cells were able to differentiate into CM-like state in vitro with structural and functional characteristics.
Human dermal fibroblasts have been reprogrammed into stem cells containing cardiac-specific genes following radiofrequency [34], retroviral delivery of KMOS transcription factors [35], and retroviral transduction of cardiac-based transcription factors [13]. Given the concern of insertional mutagenesis from retroviral delivery, we wanted to determine if nonintegrative delivery methods of mRNA would have the ability to differentiate a somatic cell into an iCPC without the risk of insertion, since in vitro transcribed mRNA will be degraded after 24 h. Previous work performed by Boissel et al. [36] and Djurovic et al. [37] have separately demonstrated the effectiveness of the viral delivery of genes compared to nonintegrative methods of electroporation [36] or lipid transfection [37]. However, a direct comparison between the two nonintegrative methods has not been performed. In the context of these studies, we wanted to determine the most effective method for nonintegrative delivery of cardiac transcription factors for potential future clinical settings. We compared the effectiveness of electroporation and lipid transfection to upregulate cardiac-specific genes using unmodified and modified mRNA GMTH cocktails or transcriptome derived from CPCs. The use of both transfection methods revealed that GMTH mRNA effectively reached the nucleus of the human dermal fibroblasts based on increased GTH expression in the nucleus (Supplementary Fig. S2), but delivery of mRNA through electroporation increased the cardiac-specific genes Nkx2.5 and cTnT to a greater extent than lipid transfection (Fig. 1).
As previously observed [38,39], cell death can be induced by hyperactivation of the innate immune system in human fibroblasts following RNA delivery, whereas hypoactivation can lead to inefficient or partial reprogramming; thus, an optimal level of activation of the innate immune system is necessary for efficient and effective reprogramming. Reprogramming induction of the innate immune system happens through the PKR and RIG-I pathways; previous experiments conducted in our laboratory (unpublished data) and others [38] suggested that the delivery of unmodified RNA cocktail through lipid-based transfection multiple times induced the activation of the PKR and RIG-I pathways leading to cell death. Activation of the innate inflammatory and TLR3 pathways has been hypothesized to mediate the viral-based reprogramming efficiency of fibroblasts, since dsRNA generated by retroviruses induces TLR3 signaling and increases effective nuclear reprogramming [40]. Innate inflammatory pathways may be targeted to a lesser degree after electroporation of unmodified mRNAs compared to lipid transfection of modified (5mCΨ) mRNAs, because electroporated mRNAs do not enter the endosomal pathway and activate TLR3. This may also explain why electroporation is more effective at reprogramming adult fibroblasts compared to lipid-based transfection. Furthermore, electroporation was associated with decreased inflammation and cell death, suggesting that electroporation was more effective and safer than lipid transfection, as previously suggested by Drews et al. [28]. Another study has demonstrated that electroporation is a more effective method for generating pluripotent stem cell-like cells compared to lipid-based transfection and the electroporated cells effectively differentiated toward neuronal lineage [41]. Future research will be directed to decreasing the reprogramming time and number of treatments required, since adult fibroblasts were electroporated four times to achieve results. Finding ways to reduce the number of steps required to achieve a cardiac precursor state will facilitate the clinical application of this proof-of-concept study.
Nonintegrated reprogramming methods were performed with both human neonatal and adult fibroblasts. Neonatal fibroblasts derived from a commercially available source were unable to reprogram into an iCPC state or further into a CM-like state after chemical treatment. Reasons for this inability are unclear, but could be a result of the innate plasticity associated with neonatal fibroblasts due to their immaturity and mesenchymal stem-like characteristics [42], since neonatal fibroblasts have the ability to be partially reprogrammed on their own to a progenitor cell state in the cardiac reprogramming media without the delivery of a specific subset of cardiac genes. Adult, primary dermal fibroblasts (fibroblastadult) have a greater predisposition toward an induced cardiac progenitor state after mRNA electroporation when compared to neonatal fibroblasts. In our studies, we lay the experimental groundwork establishing adult dermal fibroblasts as a desirable starting material for cardiac reprogramming in vitro demonstrated by a higher efficiency of differentiation toward an iCPC state that expresses c-kit.
Previously, cardiac reprogramming of murine fibroblasts demonstrated that the expression of either c-kit or Isl-1 was dispensable for successful CM induction [32]. In contrast, our experiments demonstrated that the c-kit expression was necessary in differentiation of adult human fibroblasts (fibroblastadult) and c-kit expression predicted the cell's ability to be chemically induced into functional CMs. Adult fibroblasts receiving both GMTH and exposed to 5-AZ significantly upregulated CM-specific genes, cTnT, cardiac troponin I (cTnI), α-sarcomeric actinin (α-actinin), and SERCA2 compared to mock treatment. In addition, our data show a dramatic increase in RyR2 in the c-kit+ cells compared to mock fibroblasts, CPCs, and the unselected GMTH fibroblasts. 5-AZ has been shown to upregulate CM-specific genes in progenitor cells [43,44]. However, an important finding in this study is that adult fibroblasts receiving GMTH and selected for the c-kit expression before exposure to 5-AZ displayed an even greater increase of the same CM-specific genes compared to adult fibroblasts not selected for the c-kit expression (Fig. 6D). Furthermore, c-kit+-selected fibroblastadult calcium profile demonstrated an increased physiologic response to caffeine compared to fibroblastadult not selected for the c-kit expression, which supports the greater CM-like potential observed in c-kit+-expressing cells (Fig. 6C).
The striations and rod-like morphology of the iCMs derived from c-kit+ GMTH fibroblastadult, observed in Fig. 6A, coincides with the increase in myocyte-specific genes, α-actinin, cTnT, cTnI, SERCA2, and RyR2, observed in Fig. 6D. These iCM cells have distinctly transitioned away from CPC and fibroblastadult levels toward gene expression levels more consistent with adult CMs. The calcium release (RyR2) and uptake genes (SERCA2) were significantly greater in c-kit+ GMTH cells relative to fibroblastadult and CPC, which further support the transition of c-kit+ GMTH-treated fibroblasts toward a differentiated cardiac lineage. Moreover, the greater levels of all genes in c-kit+ GMTH compared to GMTH suggest that c-kit+-selected iCPCs have a greater disposition for transitioning to a cardiac state (Fig. 6D). Caffeine-stimulated c-kit+ GMTH cells displayed a greater ability to move calcium in response to caffeine, suggesting that their RyR2 developed enough to enhance calcium release from sarcoplasmic reticulum, and the calcium released into the cytosol can then be recycled back into the sarcoplasmic reticulum by SERCA2. The caffeine-induced response of c-kit+ GMTH iCMs is theoretically consistent with a transition toward adult CM kinetics, since adult CMs have enhanced RyR calcium release kinetics compared to neonatal CMs. This c-kit+-selective response for the functional activity is not yet clear and future studies are necessary to determine if isolation of iCPCs based on other known cardiac progenitor markers would produce a similar phenomenon, for example, selection for Isl-1- or Sca-1-expressing cells, but the presented data suggest that c-kit+ selection provides a pool of cells capable of transitioning to a cardiac lineage.
Future experiments are necessary to determine if iCPCs derived from adult fibroblasts electroporated with unmodified GMTH mRNA cocktail and selected for c-kit expression can be expanded for clinical applications, if those c-kit+ iCPCs will have a potential benefit to patients suffering from heart failure, and determine if these cells can differentiate directly into cardiac cells; or will these cells act similar to MSCs by causing a paracrine effect that regenerates innate cardiac tissue [6]. Currently, various cell types used in clinical trials have been implanted into the heart through (1) transplantation during surgical procedure, (2) direct injection into the wall of the myocardium, (3) catheter-based injection of cells, and (4) intravenous injection; unfortunately, the majority of these injected cells disappear from the myocardium within 1 week and all potential benefits of the therapy appear to be paracrine in nature [45]. Future studies will evaluate the role of c-kit and its level of expression to determine whether c-kit+ cells enhance CM development. van Berlo et al. in 2014 [46] demonstrated that c-kit+ CPCs were able to differentiate into new CMs, but it was deemed a rare event in vivo. The new CMs derived from c-kit+ CPCs could not be the sole factor contributing to the beneficial outcomes observed for patients in the SCIPIO clinical trial [46,47], therefore the c-kit+ CPCs may be acting by another mechanism that has not yet been identified. Recently, Keith and Bolli [48] published an in-depth review on the controversy surrounding c-kit+ CPCs and presented their facetiously dubbed theory, “string theory” of c-kit+ cells to explain the conflicting reports from multiple laboratories on the benefits of c-kit+ CPCs on endogenous myocardium. Keith and Bolli [48] postulated that two separate population of cells express c-kit with c-kithigh expression correlating to endogenous c-kit+ cells residing in mature adult hearts that contribute minimally to restoring CM function when delivered to patients ex vivo, but may have a beneficial paracrine effect due to their mesenchymal nature compared to c-kitlow cells derived from the fetal cardiac and progenitor pool that have demonstrated the ability to differentiate into CMs in vivo [49]. Based on this new evidence, further studies are needed to determine the level of c-kit expression we observed in our iCPC population and if these iCPCs will differentiate into a cardiac fate in vivo or will only have a paracrine effect in vivo similar to MSCs.
Furthermore, identification of other transcription factors or small molecules that can be substituted for the chemical treatment of 5-AZ will also be necessary to improve the efficiency of iCPC generation. Optimization of in vivo delivery of iCPCs and cell transplantation will enable the assessment of potential arrhythmic problems that could occur [2,50]. Our results from this proof-of-concept study present a starting cellular material, RNA delivery system, and method of differentiation that may confer optimal effectiveness for those future studies. Also, the ex vivo method of differentiating primary human dermal fibroblasts used herein allows for cellular scale-up of the generated iCPCs and banking of the iCPCs for future use in cell-based therapy for cardiac repair during surgeries.
Footnotes
Acknowledgments
We are grateful to Jennifer Bond PhD and Howard Levinson MD for providing primary, adult dermal fibroblasts for research. The Clarence Gardner, M.D. Award from the Department of Surgery (S.K.N., C.A.M.) and discretionary funds from the Department of Surgery (S.K.N.) supported this work.
Preliminary studies were presented during a poster session at Oligonucleotide Therapeutic Society (OTS) in San Diego, CA on October 12, 2014.
Author Disclosure Statement
Authors have no commercial interests to disclose.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.