
Abstract
Intervertebral disc (IVD) degeneration results in segmental instability and irritates neural compressive symptoms, such as low back pain and motor deficiency. The transplanting of stem cell into degenerative discs has attracted increasing clinical attention, as a new and proven approach to alleviating disc degeneration and to relieving discogenic pains. Aside from supplementation with stem cells, the IVD itself already contains a pool of stem and progenitor cells. Since the resident disc stem cells are incapable of reversing the pathologic changes that occur during aging and disc degeneration, it has been debated as to whether transplanted stem cells are capable of providing an efficient and durable therapeutic effect, even though there have been positive outcomes in both animal models and in clinical trials. This review aims to decipher the interactions between the stem cell and the disc microenvironment. Within their new niches in the IVD, the exogenous stem cell shows metabolic adaptation to the low-glucose supply, hypoxia, and compressive loadings, but demonstrates little tolerance to the disc-like acidity and hypertonicity. Similarly, the survival of endogenous stem cells is threatened as well by the harsh disc microenvironment, which may exhaust the stem cell resources and restrict the self-repair capacity of a degenerating IVD. To eliminate the intrinsic obstacles within the stressful disc niches, stem cells should be delivered with an injectable scaffold that provides both survival and mechanical support. Quick healing or concretion of the injection injuries, which minimizes stem cell leakage and disturbance to disc homeostasis, is of equal importance toward achieving efficient stem cell-based disc regeneration.
Introduction
T
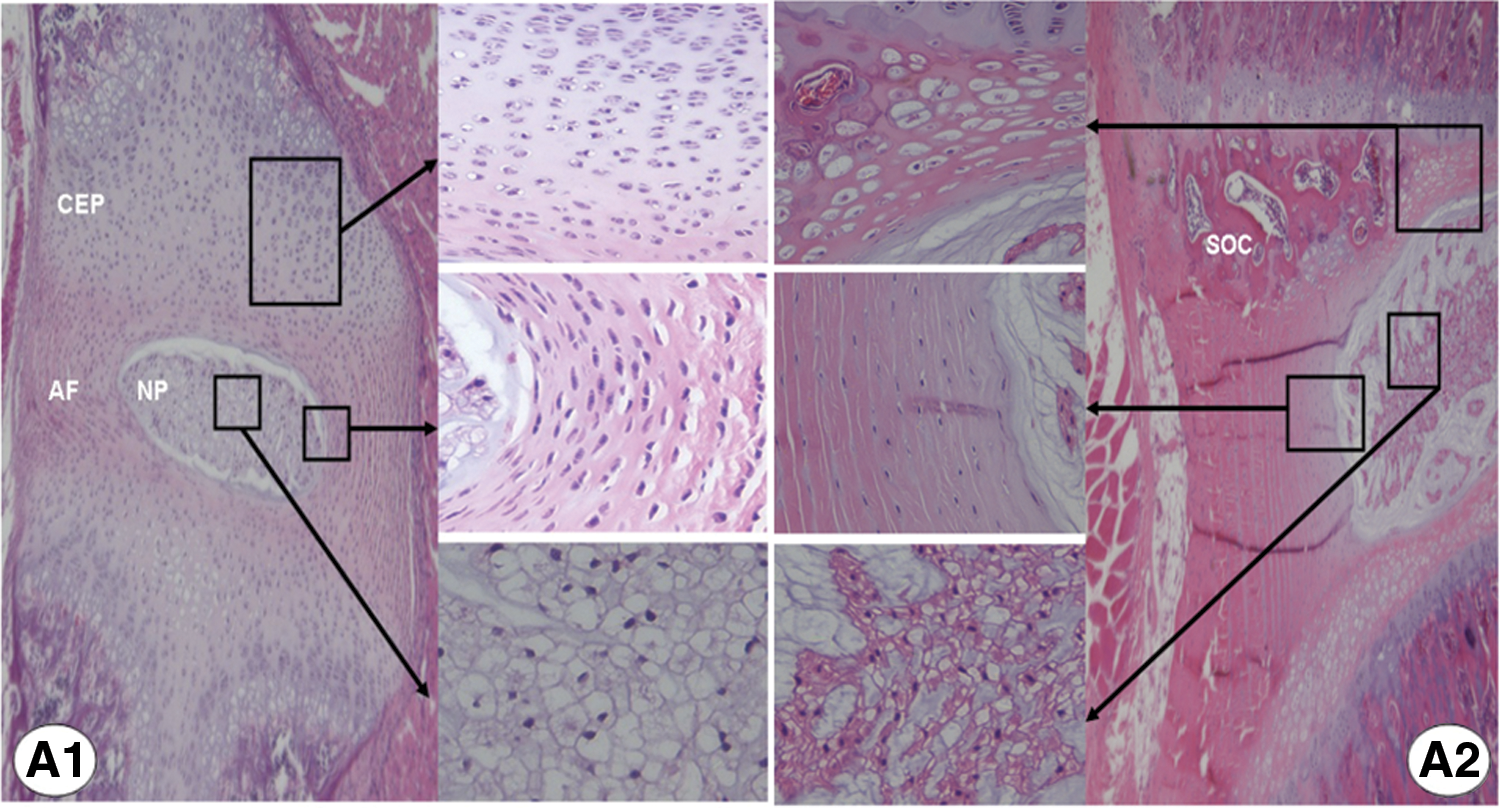
Development and age-related change of intervertebral disc (IVD). Hematoxylin–eosin staining of rat IVD. A disc from a young rat

Surgical approach to disc repair. Magnetic resonance imaging (MRI) of a 36-year-old man with symptomatic disc degeneration at lumber 4/5. The preoperation MRI
IVD degeneration is a complicated process that involves both age-related changes and tissue damage caused by multiple stresses and genetic disposition [6 –8]. Despite unknown pathogenetic details, there is increasing consensus that loss of disc cell viability and functionality is critical in disturbing disc homeostasis [6,9]. Therapies to halt this trend have included biological repairs with growth factor supply [10], gene therapy [11], and delivery of functional cells, the latter of which has been studied extensively over the last decade [5]. Owing to the wide availability and multilineage differentiation potential, stem cell transplantation has attracted most research interest and yielded positive outcomes in animal models and in clinical trials [12,13]. Interestingly, evidence is mounting that the IVD itself is already populated with resident stem cells [14,15]. Since IVD has very limited self-repair capacity [6,8,16], it is not unreasonable to presume these endogenous multipotent cell resources on their own are unable to decrease the pathophysiological process that occurs during disc aging and degeneration. We hypothesize that deficient intrinsic self-regeneration is related to the unique disc microenvironment, which is characterized by avascularity [17], nutrient deficiency [4], hypoxia [18,19], increased acidity [20,21], hypertonicity [22 –24], and mechanical loading [25,26]. Thus, the harsh survival conditions may also compromise the regenerative effects of those transplanted stem cells.
This review aims to decipher the stem cell behaviors during IVD degeneration and regeneration. By exploring the interactions between stem cells and their new niche in degenerated IVD, we will focus on the obstacles and limitations of stem cell therapy with particular focus on those nonregenerative and detrimental effects complicating stem cell transplantation. Herein, stem cells are defined according to the criteria of multipotent mesenchymal stromal cells [27] as cells (1) capable of plastic adherence in standard culture conditions; (2) positive for CD105, CD73, CD90 and negative for CD45, CD34, CD14 or CD11b, CD79a or CD19, HLA-DR; and (3) capable to differentiate into osteoblasts, adipocytes, and chondroblasts in vitro. Publications discussing progenitor cells or induced pluripotent cells are also included in our review.
Aging and Degeneration of IVD
Age-related change of IVD
Studies in embryogenesis have revealed that the NP originates from the axial notochord, while the AF and CEP are formed from the paraxial mesoderm [28 –30]. During growth and maturation, IVD undergoes significant histocytological changes that include transition of the notochord cells to the chondrocyte-like NP cells, thinning of the cartilaginous layers in the end plate, and fibrocartilaginous metaplasia of the organized AF (Fig. 1). Accompanying this, the gel-like NP reduces the content of aggrecan and collagen type II in the hydrophilic extracellular matrix (ECM) [31], thus weakening its capacity to withstand axial compressive loadings. Disorganization of AF often starts at the transition area between NP and inner AF, where the cartilaginous matrix accumulates and compromises tensile strength of the fibrotic AF (Fig. 1). Since IVD is avascular and its nutrient supply is limited by the inefficient permeation of the end plate [17,32], the thinning and calcification of CEP is supposed to cause a further nutritional deficiency within aged NP.
Although a widely accepted definition of IVD degeneration is not available as yet, this process is currently known to mimic many of those age-related changes and is substantially accelerated and/or exacerbated by multiple degeneration-inducing stressors [6,8,16]. To date, promoters of IVD degeneration have extended from aging to excessive mechanical loading [33,34], disc injury [35,36], oxidative stress [37,38], diabetes [39,40], chronic tobacco smoking [41,42], obesity [43,44], and genetic inheritance [7]. These etiological factors are overlapping and may work in a coordinated manner to disturb disc homeostasis.
Molecular basis of IVD degeneration
In contrast to other musculoskeletal tissues, adult IVD contains a small cell population that occupies <1% of the total disc volume [45]. These cells are sparsely located in the ECM and function to maintain a balance between anabolism and catabolism. During aging and degeneration, excessive apoptosis [46] and cellular senescence [8,16] contribute primarily to a reduction in viable disc cells. A deficiency in anabolic factors, such as transforming growth factor-β (TGF-β) [47] and insulin-like growth factor-1 (IGF-1) [48], may cause further reduction in cellular viability and ECM synthesis. Meanwhile, increased expression of proinflammatory cytokines [1,49], aggrecanases [50], and matrix metalloproteinases [51,52] promotes substantial matrix degradation, thereby enhancing the imbalance between anabolism and catabolism. By generating reactive oxygen species (ROS) and catabolic cytokines [16,37,38], the accumulated senescent disc cells may further deteriorate the microenvironment within aged and degenerated IVD. Based on these observations, an ideal biological regeneration should not only provide disc with sufficient functional cells but also be dedicated to improving the survival conditions in a degenerating IVD. Additionally, the safety concern, user-friendliness, and durability of therapeutic effects are of equal importance when translating stem cell-based disc regenerations from basic research into clinical practice.
Exogenous Stem Cell and IVD Regeneration
The idea of transplanting stem cells into IVD was introduced initially by Sakai et al. in 2003 [12]. Using autologous bone marrow mesenchymal stem cells (MSC) embedded in collagen gel, Sakai and colleagues found that stem cell injection could decelerate degeneration in rabbit IVD. Since then, the stem cell-based regenerations have been extensively studied across species in rat [53,54], canine [55,56], and porcine IVD [57,58]. The stem cell resources used for disc repair have also extended beyond the bone marrow to adipose [56,59 –61], muscle [62], synovium [63 –65], olfactory mucosa [66], placenta [67], and the umbilical cord [68 –70].
Regenerative effects of stem cells
In the disc treated with stem cell supplement, the content of aggrecan and type II collagen was reserved in most cases [12,35,57,61,71 –73]. Accompanying this was the preserved disc height and the magnetic resonance imaging (MRI) T2 signal, which in our observation correlated well with improved histopathological evaluations [35]. The results of one study suggested that the restored ECM content of NP also contributes to maintain perfusion and permeability of the vertebral end plate [58].
In addition to matrix restoration, significant pain relief has been reported in clinical observations [13,74 –76]. However, increased disc height was seldom evidenced in those pain-relieved human discs, suggesting the nerve root decompression might not originate directly from the restored matrix content. The longest observation of stem cell transplantation was 6 months in animal discs [73]; however, the pain relief may last longer than 1 year under clinical trial conditions [13,74]; thus, additional case–control studies are required to validate active ingredients and therapeutic significance of stem cell therapy.
Regarding the issue of safety, complications of acute discitis and epidural abscess were reported in a 64-year-old patient who received intradisc injection of bone marrow and adipose tissue [77]. In a 41-year-old man with cervical herniated IVD, a pulmonary embolism occurred after intravenous autologous adipose stem cell therapy [78]. Similar concerns may arise from the back flow of stem cells into epidural space, which compromises disc regeneration and increases the risk of ectopic ossification [79]. More importantly, despite the improved radiological and histochemical markers, no study has reported a disc with stem cell injection that could be equally normal to or more rejuvenated than a normal control disc.
Regenerative mechanisms of stem cells
After transplanted into IVD, stem cells have been found to migrate, relocate, and even home the inner AF [80 –83]. In the NP tissue, the stem cells have been shown to demonstrate chondrocyte-like round morphology, whereas in the inner AF, they were much elongated and fibroblastic, suggesting differentiation into the disc-like cells [80]. In support of this finding, several chondrocytic markers, such as aggrecan, type II collagen, and Sox-9, were induced in the MSC that were transplanted into NP [35,57,72], cocultured with NP cells [64,68,84,85], or maintained in the NP-like hypoxic and mechanical conditions [86 –88]. When cocultured with AF cells [89] or seeded onto structure mimicking AF lamellae [90,91], the MSC tended to produce fibrotic ECM that was rich in type I collagen. Within a degenerating disc, the stem cell-derived disc-like cells may compensate for the cellular loss caused by apoptosis and senescence, enhance anabolism, and therefore contribute to restore ECM content and disc homeostasis.
In addition to differentiation, the stem cell itself is a reservoir of various growth factors and cytokines. Paracrine secretion of these bioactive factors exerts trophic effects onto the neighboring cells [92,93]. A large number of coculture studies have shown that disc cells increase proliferation and activate matrix synthesis after being cocultured with MSC [94,95]. The enhanced disc cell viability might originate from the elevated growth factors in the coculture system, which includes but is not limited to TGF-β, IGF-1, endothelial growth factor, basic fibroblast growth factor (bFGF), and various bone morphogenetic proteins (BMPs) [94,95]. Most of these growth factors have been proven to be not only as anabolic to disc cells [10] but also as antisenescence (TGF-β, IGF-1) [96,97], antiapoptosis (IGF-1, BMP-7) [98,99], anticatabolic (TGF-β, BMP-7) [100,101], and anti-inflammatory (TGF-β, BMP-7) [100 –102]. Given that discogenic pain is closely related to proinflammatory cytokines, such as tumor necrosis factor-alpha (TNF-α), interleukin (IL)-1β, and IL-6 [1,49], the pain relief effects of stem cell therapy might also result from these trophic molecules [60].
Stem cells can affect disc cells through paracrine secretion [95], direct cellular contact [94,103 –106], and even cell-to-cell fusion [107]. Direct contact with disc cells has been known to enhance the biological functions of stem cells [105,106], probably by amplifying the secretion of trophic factors. Although the regenerative process involves both differentiation into disc-like cells and trophic cocktails, it remains elusive whether or not the two mechanisms occur simultaneously and contribute equally to disc regeneration. Our previous study showed that the expression of TGF-β1 and IGF-1 in the GFP-labeled MSC peaked at 2-week postintradisc transplantation, whereas the type II collagen and hypoxia inducible factor-1α (HIF-1α) showed a steady increase for at least 8 weeks [108]. Collectively, it is probable that stem cell-based disc regeneration is a dynamic process that initiates an early trophic effect and then a gradual differentiation into new disc cells.
Endogenous Stem Cells and IVD Self-Repair
Identifying IVD stem cells
Emerging evidence has shown that cells isolated from NP [14,15,109 –111], AF [15,109,112,113], and chondrocytic end plate [114,115] also express most of the phenotype markers that define mesenchymal stromal cells [27], thus supporting the existence of resident stem cells within IVD. To date, endogenous stem cells have been detected in human [15,109,110,116], rhesus macaque [117], porcine [118], murine [14], rat [111], and rabbit IVD [113,119]. These cells maintain the potential of multilineage differentiation, but are diversified in phenotype signatures and biological capacities (Table 1). Compared to bone marrow MSC, the MSC derived from degenerated NP demonstrated much lower adipogenic differentiation ability [110], whereas the end plate MSC exhibited an enhanced potential of osteogenesis and chondrogenesis [114]. When maintained in the disc mimicking microenvironment, the NP MSC exceeded the adipose-derived MSC in tolerating hypoxic [120], hypertonic [121], and acidic culture conditions [111].
AF, annulus fibrosus; CEP, cartilaginous end plate; CK, cytokeratin; CSE, chondroitin sulphate epitopes; Flt-1, vascular endothelial growth factor receptor 1; GD2, disialoganglioside 2; HLA-DR, human leukocyte antigens-DR; IVD, intervertebral disc; Notch-1, Notch homolog 1; NP, nucleus pulposus; NSE, neuron-specific enolase; NTR, low affinity nerve growth factor receptor; Oct-4, octamer-binding transcription factor-4; Sox-2, SRY-related HMG-box-2; SSEA-4, stage-specific embryonic antigen-4; Stro-1, stromal cell antigen-1; Tie2, tyrosine kinase receptor 2.
Since stem cell markers change with disc aging and degeneration [118,119], the altered components of stem cell pools might account for the varied biological behaviors in those MSC isolated from degenerated IVD. This notion is, in part, supported by the discovery of tyrosine kinase receptor (Tie2) and disialoganglioside 2 (GD2), double-positive NP progenitors in the human and mice disc [14]. In contrast to the Tie2-positive but GD2-negative ancestors, the double-positive cells preserve multipotency and are superior to form chondrocytic clones with increased accumulation of aggrecan and type II collagen. Moreover, the survival and functionality of the NP progenitor cells have been revealed to be maintained by interacting with angiopoietin-1, which is the Tie2 ligand and coexpressed in the Tie2-positive cells [14]. Although the identification of disc stem cells has provided new approaches to understand and regenerate IVD degeneration, many questions remain unanswered regarding the origin and spatiotemporal regulation of these multipotent cell resources.
Origin of IVD stem cells
Given that IVD itself is avascular and originates from the axial (notochord) and paraxial mesoderm (somite) [28 –30], it is possible that the endogenous stem cells are remnants of the multipotent mesendoderm cells during embryogenesis. The stem markers are ubiquitously detected across species both in the young and the aged IVD [15,111,113,117,118]. Furthermore, cells isolated from the IVD of fetuses have been shown to maintain multipotency and inherit enhanced chondrogenic differentiation capacity [122,123]. Based on this perspective, the stem cell pools, especially those chondrocytic progenitors, may be stored in the thick cartilaginous layers of the young end plate (Fig. 1). The large and vacuolated notochord cells in the gelatinous NP, which yield offspring of smaller chondrocyte-like cells and giant cells [124], are probably precursors of the NP progenitor cells [14,125,126]. This inference is in accord with the positive staining of notochord makers both in the smaller bovine NP cells [127] and human degenerated NP tissues [128]. Since the end plate chondrocytes can be attracted and can migrate into NP tissues [129 –131], the NP stem cell pools might also contain a proportion of those end plate stem or progenitor cells. A recent study lent strength to this notion by showing that the migration of end plate-derived stem cells is regulated by the macrophage migration inhibitory factor produced by NP cells [132]. Consistent with this, stem cells isolated from the CEP and the NP demonstrated similar mesenchymal phenotypes [110,114].
In addition to the remnants of embryonic multipotent cells, there is increasing evidence supporting the infiltration or migration of exogenous stem cells into the IVD [133 –135]. To date, specific stem cell niches have been identified in the outer AF borders that connect the ligament or perichondrium of adjacent vertebrae [133]. Cells expressing progenitor markers (Notch1, Delta4, Jagged1, C-kit, Ki-67, Stro-1) are detected there and can migrate along certain routes [positive for growth differentiation factor-5 (GDF-5), Sox-9, SNAIL homolog 1 and 2, β1-integrin] toward the inner parts of IVD [134]. Given that similar patterns of cellular migration exist in the rabbit knee joint, recruiting cells from these stem cell niches seems to be a highly conserved self-repair mechanism for cartilage tissues [135].
Despite the avascular nature, MSC from the neighboring vertebra bone marrow might be another resource of disc stem cells. In a new degeneration model induced by imbalanced loading, the intravenous delivered bone marrow-derived cells can be detected in the end plate capillary, inner AF, and NP of the degenerated tail disc [136]. However, the limited number of infiltrated cells suggests that the self-pair of IVD is probably a chronic and inefficient process [136,137]. Although elevated chemokines such as Chemokine (C-C motif) ligand 5 (CCL5) and stromal cell derived factor-1 (SDF-1) contribute to attract stem cells and facilitate migration [138,139], it remains to be determined whether a degenerating disc actually benefits from these multipotent cell resources. Several differentiation studies provide evidence that disc-derived stem cells can also be induced to express neural or endothelial markers [112,140]. By causing revascularization and/or reinnervation [1,45], the multipotent disc cells may contribute as well to disturb IVD homeostasis and to irritate discogenic pains.
Stem Cell Tolerance and IVD Degeneration Microenvironment
Stem cell niches are composed of a tissue-specific microenvironment and supporting cells that regulate self-renewal and differentiation of stem cells [93]. After being transplanted into IVD, the exogenous stem cells are supposed to survive a new microenvironment that is significantly different from their original niche. In the avascular IVD, cells have to sustain nutrient deficiency, low pH, hypoxia, hypertonicity, as well as various mechanical loads (Fig. 3). Some of these microenvironment factors are not only harsh for resident disc cells but also challenging to the viability and functionality of the transplanted cells [111,121,141,142]. To date, the longest survival of exogenous stem cells was reported as 6 months in a degenerated rabbit disc [73]. Even though the supplemented cells could have lived longer, the age-related exhaustion of endogenous progenitors [14,119] raises the concern that the harsh disc niches may not support a stable and long-lasting stem cell-based regeneration.
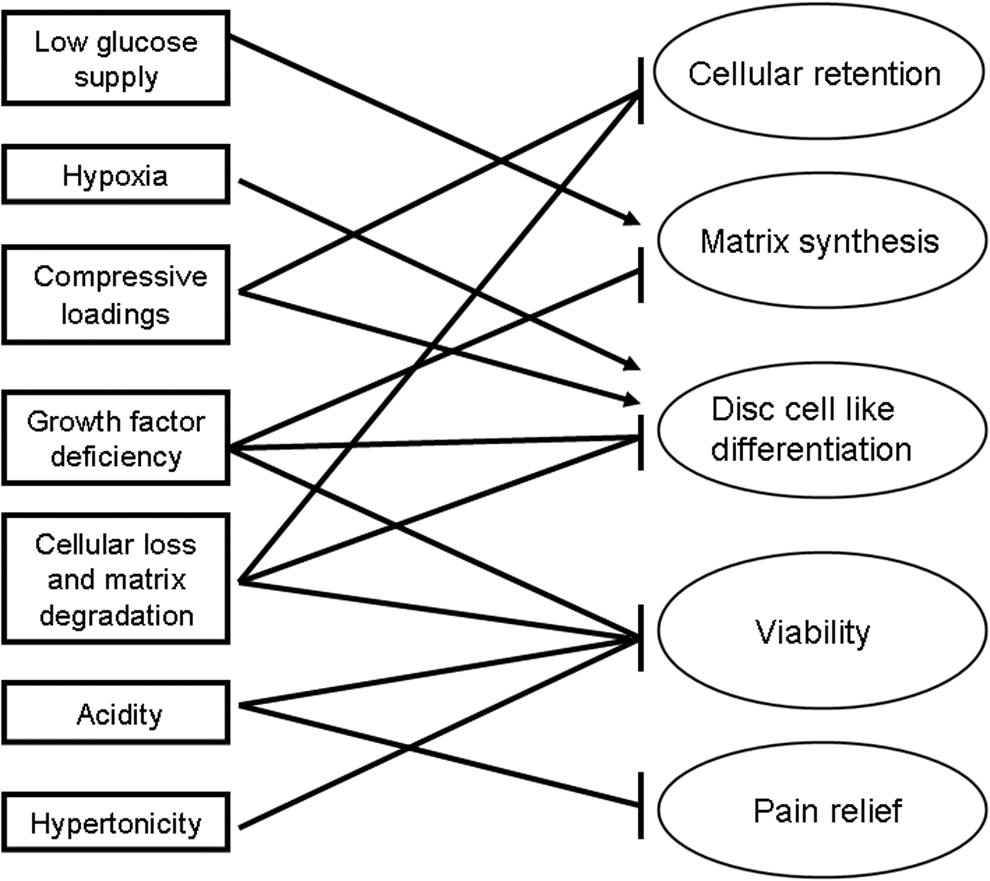
Effects of disc microenvironment on stem cells and stem cell-based disc regeneration. For disc regeneration, both the exogenous and the endogenous stem cells have to survive disc microenvironment first and then generate trophic cocktails and/or differentiate into disc-like cells. Within the unique disc niches, stem cells can metabolically adapt to low-glucose supply, hypoxia, and compressive loadings, but demonstrate little tolerance to the increased acidity and hypertonicity. Moreover, the stem cell-based disc regeneration can be further crippled by low cellularity, matrix degradation, growth factor deficiency, stem cell leakage, as well as the pain irritating acidity. During disc aging and degeneration, the harsh survival conditions may contribute to exhaust the endogenous multipotent cell resources, thus restraining the intrinsic self-repair within a degenerative disc.
Altered cellular and matrix content
In coculture systems, the stem cells contacting directly with disc cells have demonstrated most efficient trophic effects and chondrogenic differentiations [94,105,106]. There are studies suggesting the NP-like differentiation of human MSC can also be enhanced by the conditioned media from notochord cells [143,144]. Unfortunately, these ideal ex vivo conditions cannot be translated directly into the aged or degenerated disc, where few notochord cells are available [125 –127], and the resident disc cells are sparsely located within thick ECM [45].
After implantation into IVD, stem cells are supposed to be interacting primarily with various ECM components. A large number of scaffold studies have revealed that the proliferation and differentiation of stem cells are maintained by macromolecules, such as type II collagen [145 –147], hyaluronan (HA) [147,148], leucine-rich proteoglycans [117], elastin [145], and laminin [69]. In human umbilical cord mesenchymal stromal cells, the laminin-receptor integrin α3 and β4 subunits have been detected [69]. Similarly, the HA receptor CD44 is widely expressed in the disc-derived stem cells [14,113,114,117,140,149]. Although the biological significance of their interactions remains to be determined, breakdown of these matrix contents may contribute to disturb cellular adhesion, thus threatening the survival of both endogenous and transplanted stem cells.
Nutrient deficiency
The IVD is avascular, and the end plate capillaries are responsible for the nutrient supply to the disc cells [17]. As is evidenced by the MRI tracing of intravenous-injected contrast medium, solute diffusion through the cartilaginous layer is much less efficient than the adjacent vascularized tissues [32]. Furthermore, the nutrient transport can be further slowed down by disc aging and excessive mechanical loads [150,151], probably by promoting ossification of CEP. To maintain viability under nutritional deficiency, disc cells may adaptively reduce the demand for nutrient supplies. This is supported by the findings that NP cells enhance proliferation in low-glucose cultures, while AF cells accelerate senescence with glucose abundance [36,152]. The restricted glucose consumption of resident disc cells is in line with their anaerobic glycolysis under hypoxia, which generates energy at a lower cost of oxygen and ROS production [18,19,38].
Interestingly, in the IVD-like low-glucose conditions, MSC derived from bone marrow and adipose can also maintain viability and increase matrix synthesis [141,142]. The metabolic adaptation to glucose deficiency may facilitate the temporary survival of stem cells into their new disc niches. However, for longer maintenance of stem cell viability and functionality, a stable supply of growth factors is also required. It has been revealed that the NP-like differentiation of MSC can be enhanced by supplementation with TGF-β1 [86,153], TGF-β3 [154], GDF-5 [155], GDF-6 [156], BMP-2 [153,154], or cocktails of IGF-1, FGF-2, and platelet-derived growth factor [157]. Although disc cells express most of these factors [158,159], their paracrine actions are likely to be crippled by the low cellularity, thick ECM, as well as the age-related decrease of several anabolic signals [45,47,48]. In light of end plate calcification, diffusion of circulating anabolic factors may be also attenuated in aged or degenerated discs [150,151]. Endeavors are in progress to provide stem cells with scaffolds capable of growth factor release [160]. However, to date, no injectable scaffold has met the requirements for both cellular retention and nourishment support.
Hypoxia
One overriding aspect of disc cell biology is that they are removed from the blood supply and resided under conditions of hypoxia [17 –19]. An increasing number of studies have demonstrated that NP cells enhance proliferation in low oxygen tension and stabilize the expression of HIF-1α even under normoxia [161 –165], indicating a metabolic adaptation to the local hypoxic niches.
With regard to MSC, hypoxia is not only favorable for maintaining viability but also a potent inducer for the NP-like differentiation and matrix synthesis [65,67,86,155,166,167]. Moreover, hypoxia can also antagonize the inhibitory effects of IL-1β on the chondrogenesis of bone marrow-derived MSC [168]. For stem cell-based IVD regeneration, it is suggested that the entire ex vivo process of MSC expansion and modulation be performed under the disc-like hypoxic conditions [166]. However, given that disc injection breaches tissue integrity and promotes revascularization [35], the transplanted stem cells are possibly experiencing fluctuated oxemic tensions within a degenerated IVD. This may initiate oxidative damages since a higher level of mitochondrial-derived superoxide anions is detected in the disc cells under normoxia [38].
By stabilizing HIF-1α independent of oxygen tension [161 –164], the resident NP cells might have metabolically tolerated the oxemic shift during disc aging and degeneration. Although stem cells can be induced to express HIF-1α within disc niches [86,108], it is unknown whether these new NP-like cells also demonstrate an adaptive stabilization of HIF-1α and survive the disturbed oxemic homeostasis. To avoid the revascularization and oxemic fluctuation that complicate annulus puncture injuries, transpedicular cell delivery has been proved technically feasible in ovine and human lumber IVD [169]. Since this approach damages the end plate, more studies are needed to validate its safety and user-friendliness. In the alternative, the side-firing holmium laser in transforaminal endoscopic discectomy may provide quick concretion of annulus lesions [2]. Nevertheless, it remains to be determined whether a disc with a cracked AF and herniated NP is indicated for stem cell therapies [79,170].
Mechanical loadings
Healthy IVDs, especially those at the cervical and the lumber segments, function to absorb and conduct varied mechanical forces, including compression, shear, torsion, and flexion [25,26]. Depending on the loading pattern and intensity, the disc-specific mechanical features have been known to exert a wide range of impacts on both disc cell biology and ECM turnover [25,26]. In scoliotic disc, degeneration is potentially accelerated by the unbalanced mechanical loadings that promote calcification of CEP and deteriorate nutritional deficiency [151]. Transition of the notochord cells to the smaller chondrocyte-like NP cells, which signifies an early onset of disc degeneration, can be significantly enhanced by static compressive loadings or dynamic hydrostatic pressures [34,171].
Taking into account that the overwhelming majority of cells within adult NP, including the NP progenitors, are derived from the embryonic notochord resources [124 –126,172], the accumulated mechanical stress might contribute to exhaust, at least, the NP stem cells during disc aging and degeneration [14,118,119]. Studies in the foreleg-amputated rats suggest that premature senescence can be induced by mechanical injuries [33], thereby potentially accelerating the loss of both disc cells and their precursors. Additionally, the enhanced differentiation driven by mechanical stimuli might also account for the exhaustion of multipotent disc cells. For example, radial compressive loadings promote the AF-like differentiation in bone marrow MSC [173], whereas the NP-like differentiation of adipose-derived MSC can be significantly activated by dynamic compressions [88].
Given that the energy metabolism in resident disc cells responds differently to the static and dynamic compressive stimulations [174], the AF-like or NP-like differentiation of MSC may suggest a mechanical adaptation into the local disc microenvironment. Although this potentially strengthens the differentiation role of stem cell therapy [175], the trophic effects are unlikely to benefit from these compressive loadings, which increase intradisc pressures and promote backflow of stem cells into epidural spaces [79]. To tackle this problem, improved disc regeneration was achieved by combined application of stem cell implantation and spine distraction [176]. In addition, an injectable annulus plug was also designed to block the injection portal that alleviated cell leakage during intradisc cell delivery [177]. Since most symptomatic IVD has lost disc height and yielded segmental instability [1], future studies are needed to provide injectable and expendable scaffolds capable of immediate mechanical supports.
Acidity
Owing to the anaerobic glycolysis, lactic acid is generated and accumulated within the hypoxic disc niches [18,19,163]. It is reported that the pH levels average 7.0–7.2 in normal IVD, but can be as low as to 6.5 in the severely degenerative IVD [178]. The extracellular acidity has been known to be sensed by acid-sensing ion channels (ASICs) that include six subunits: ASIC1a, ASIC1b, ASIC2a, ASIC2b, ASIC3, and ASIC4 [179], and most of which are expressed in human IVD [20,21,180]. Of these, ASIC1a mediates intracellular Ca2+ influx and is proapoptotic to end plate chondrocytes [181]; activated ASIC3 is contributing to the survival of both AF and NP cells under acidic stresses [21].
In contrast to disc cells, the viability and matrix synthesis of MSC are significantly inhibited when cultured in low pH conditions [111,141,142,178]. Since normal disc cells express mainly ASIC2 and ASIC3 [180], while bone marrow MSC are positive for ASIC1a and ASIC3 [182], the varied ASIC signatures might account for their distinct adaptations into the acidic disc niches. A recent comparative study with normal and degenerative human discs shows that the expression of ASIC1, ASIC2, and ASIC3 is increased in the degenerated NP, whereas the ASIC1 and ASIC4 are elevated in the AF tissues [180]. Given that the basal ASIC3 activity of disc cells is maintained by the neural growth factor (NGF)/p75/ERK signals [21], the elevated NGF expression may contribute to tolerate the increased acidity within degenerative IVD [183,184], possibly by enhancing the activity of ASIC3.
However, being a major neurotrophic factor, the activated NGF is generally associated with inflammation [185,186], innervation [187,188], as well as pain induction [1,184]. Similarly, under the neural differentiation conditions, the NP-derived stem cells also upregulate ASIC3 and gain adaptation to the acidic disc niches [140]. These observations indicate that tolerating disc acidity is probably a painful process that promotes neurogenetic differentiation and facilitates nerve ingrowth. This might weaken the pain relief effects of stem cell therapies [13], although the cellular survival within a degenerating disc could be enhanced by upregulating ASIC3 and/or downregulating ASIC1a [21,140,189]. There are studies suggesting that the ASIC3 overexpression in dorsal root ganglions contributes to evoke radicular pains [190,191]. In addition to matrix preservation and trophic cocktails, it remains to be determined whether the stem cell-based pain relief also involves the regulation of ASIC components in those disc-related nerve fibers.
Hypertonicity
Within the avascular IVD, the gelatinous NP maintains hypertonic osmotic pressures to facilitate nutrient diffusions and resist compressive loadings [22 –24]. It has been known that the hypertonicity originates mainly from the glycosoaminoglycan side chains of aggrecan molecules and can be sensed by NP cells to activate the tonicity enhancer-binding protein (TonEBP) [23]. Once activated, the transcription factor TonEBP binds to the TonE motif of tonicity-sensitive genes, transactivating the expression of heat shock protein 70 [22], β1, 3-glucuronosyltransferase-I [192], aquaporin2 [193], as well as the aggrecans [23]. Although most of these TonEBP targets function to synthesize and preserve the water-bounding matrix [194], the hypertonic stimuli have been revealed to irritate DNA damage response and arrest cell proliferation in the bovine NP cells [195]. For the MSC, the cellular viability and matrix synthesis are also reduced by disc-like high osmolarity [121,141,142].
Conversely, by restoring the osmotic balance across the cell membrane, the notochord cells have demonstrated tolerance to hypotonic stresses [196]. This is attributed to the large cytoplasmic vacuoles that release their low-osmolar content to dilute osmotic pressures [196]. However, to date, it is unknown whether these osmoregulatory vacuoles contribute as well to tolerate the hypertonicity. There is a study showing that stem cell transplantation can restore the notochord content in a degenerative IVD [197]. Future experiments are needed to understand the osmotic regulation of TonEBP and its targets in both the notochord cells and the transplanted stem cells.
Injectable Scaffold for Stem Cell Delivery
To minimize the disturbance of disc homeostasis and provide seed cells with survival support, the ideal scaffold for stem cell-based disc regeneration should be injectable or in situ forming, which allows less invasive delivery of stem cells and simple incorporation of various bioactive factors. To date, the main components of injectable scaffolds range from nature biopolymers, such as chitosan [198], alginate [198,199], and collagen [145,147,200 –203], to synthetic polymers like polyethylene glycol [204], poly N-isopropylacrylamide (PNIPAAm) [205,206], and pHEMA-co-APMA grafted with polyamidonamine [207]. Besides, both the nature and the synthetic polymers can be further crosslinked with HA [147,148,200,202 –204,208], aggrecan [200], elastin-like polypeptide [209], as well as chondroitin sulfate (CS) [205,206]. In a degenerating disc with enhanced ECM degradation, these incorporated matrix macromolecules might allow stem cells to contact with a comparatively healthier ECM microenvironment, thus potentiating cellular survivals during and after transplantation [145 –148].
To tolerate growth factor deficiency within degenerated IVD, an injectable scaffold incorporated with a synthetic glycosaminoglycan analog has been revealed to facilitate chondrogenesis of mesenchymal precursor cells, even without an additional supplement of TGF-β [204]. Similar results were also reported in a HA/PNIPAAm scaffold that supported the disc cell differentiation of human MSC in the absence of TGF-β or GDF-5 [148]. In a three-dimensional microgel composed of type II collagen and HA, combined use of nonviral gene transfection has been proved feasible to increase the production of therapeutic proteins in the embedded adipose-derived stem cells [202].
To alleviate implant expulsion under compressive loadings, thermogelling and in situ-forming scaffolds are recommended [205,210]. The temperature-sensitive components in these scaffolds, such as hydroxybutyl chitosan [210] and PNIPAAm [148,205], remain as flowable solution at around 32°C while forming compact hydrogel at physiological temperature (usually 37°C). A recent study reports an in situ-forming scaffold that significantly enhances adhesive strength by incorporating CS and alginate particles to the PNIPAAm polymer [206]. Herein, the alginate forms ionic and/or hydrogen bonds with the disc matrix components, whereas the cross-linked CS provides swelling and mechanical property in the in situ-formed gels [206]. Additionally, local adhesives can also be achieved in fibrin sealants [211 –213] or improved by adding aldehyde groups onto scaffolds [206]. However, the low cohesive strength of the fibrin hydrogel and the cytotoxicity of the reactive aldehyde have restrained their applications into IVD regenerations [206].
Although no injectable scaffold has been reported to decrease disc acidity or hypertonicity, the NP stem cells have been shown to perform better than the adipose-derived stem cells and the NP cells under the acidic [111] or hypertonic microenvironment [121]. As an alternative, the intrinsic obstacles for stem cell-based disc repair might also be alleviated by selecting appropriate seed cells for transplantation. Nevertheless, more in vivo studies are needed to validate whether these injectable scaffolds are capable of protecting the injected stem cells from the harsh disc microenvironment and promoting IVD regeneration.
Conclusions and Outlook
Despite its nascent clinical applications, the stem cell-based disc regeneration has provided new approaches to tackle disc degeneration and relieve discogenic pains. By differentiating into disc-like cells and generating trophic cocktails, the stem cell supplement contributes to restore ECM content and regain balance between anabolism and catabolism. Aside from transplantation, the IVD itself is already inherited with multipotent cell resources or attracts progenitors from the nearby stem cell pools and the circulatory system (Fig. 4). Within their new niches in the IVD, the exogenous stem cells have demonstrated metabolic adaptation to the low-glucose supply, hypoxia, and compressive loadings, but little tolerance to the disc-like acidity and hypertonicity. For the endogenous stem cells, the harsh disc microenvironments contribute not only to their age-related or accelerated exhaustion but also to restrict the self-repair capacity in a degenerating IVD.
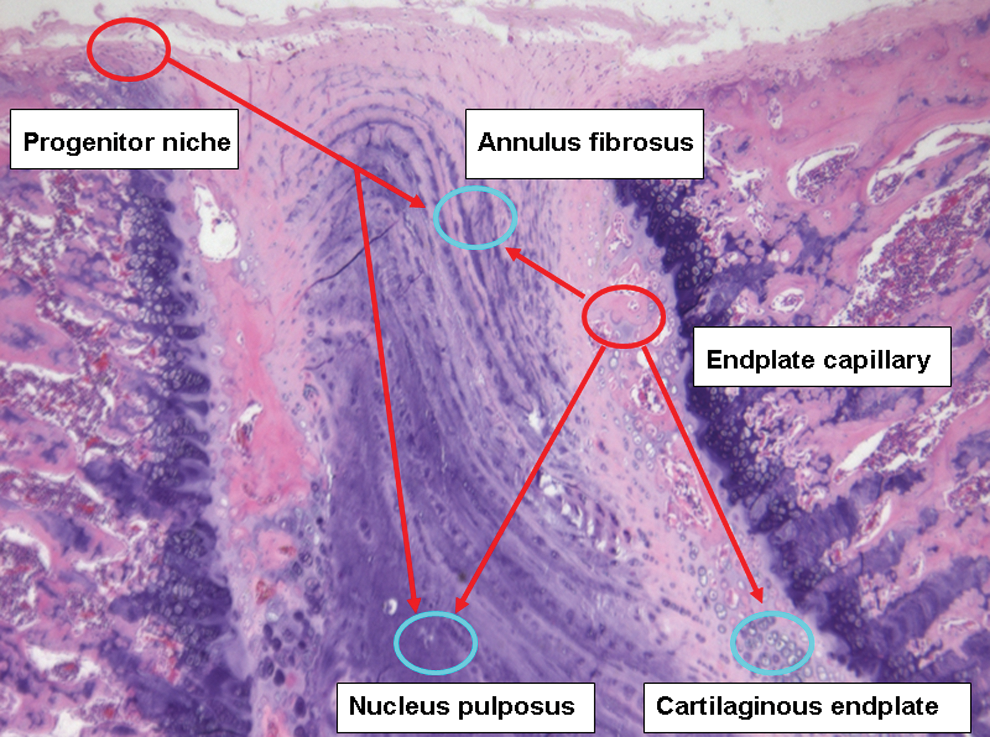
Migration route of stem cells into IVD. Hematoxylin–eosin staining of rat IVD (20 weeks old). Based on the study by Henriksson et al. [133], specific stem cell niches localize in the outer AF borders that connect the ligament or perichondrium of adjacent vertebrae, where cells express progenitor markers that can migrate along certain routes toward the inner parts of IVD. According to the study by Sakai et al. [136], intravenous delivered bone marrow-derived cells can be detected in the end plate capillary, which may be attracted into CEP, inner AF, and NP. Color images available online at
To achieve efficient stem cell-based disc regeneration, future studies should better understand the interactions between stem cells and disc niches and provide an injectable scaffold capable of cellular retention, nourishing supply, and immediate mechanical support. Moreover, quick healing of the injection injuries is also needed to minimize the back flow of stem cells and the disturbance of disc homeostasis. Since the intrinsic obstacles for stem cell-based disc regeneration cannot be solved once for all merely through scaffold renovations, choice of stem cell sources [214], supplement of bioactive factors [215], and activation of resident disc stem cells [216] are also essential for a successful IVD regeneration. For better evaluating the differentiation effects of stem cell therapy, identifying more specific phenotypes of disc cell is needed [172]. To achieve guidelined clinical practice, indications for stem cell transplantation should be defined to benefit the right discs and the right patients [214]. Progresses in these important issues are subjects of several recent reviews that expend our understanding of stem cell approaches to IVD repairs [5,172,214 –217].
Footnotes
Acknowledgments
This study was supported by the National Nature Science Foundation of China (nos. 81201423, 81272035, 81071493, and 31070876) and the Fundamental Research Funds for the Central Universities (no. KYLX_0202).
Author Disclosure Statement
No competing financial interests exist.