
Abstract
Endothelial cells (ECs) that are differentiated from induced pluripotent stem cells (iPSCs) can be used in establishing disease models for personalized drug discovery or developing patient-specific vascularized tissues or organoids. However, a number of technical challenges are often associated with iPSC-ECs in culture, including instability of the endothelial phenotype and limited cell proliferative capacity over time. Early senescence is believed to be the primary mechanism underlying these limitations. Sirtuin1 (SIRT1) is an NAD+-dependent deacetylase involved in the regulation of cell senescence, redox state, and inflammatory status. We hypothesize that overexpression of the SIRT1 gene in iPSC-ECs will maintain EC phenotype, function, and proliferative capacity by overcoming early cell senescence. SIRT1 gene was packaged into a lentiviral vector (LV-SIRT1) and transduced into iPSC-ECs at passage 4. Beginning with passage 5, iPSC-ECs exhibited a fibroblast-like morphology, whereas iPSC-ECs overexpressing SIRT1 maintained EC cobblestone morphology. SIRT1 overexpressing iPSC-ECs also exhibited a higher percentage of canonical markers of endothelia (LV-SIRT1 61.8% CD31+ vs. LV-empty 31.7% CD31+, P < 0.001; LV-SIRT1 46.3% CD144+ vs. LV-empty 20.5% CD144+, P < 0.02), with a higher nitric oxide synthesis, lower β-galactosidase production indicating decreased senescence (3.4% for LV-SIRT1 vs. 38.6% for LV-empty, P < 0.001), enhanced angiogenesis, increased deacetylation activity, and higher proliferation rate. SIRT1 overexpressing iPSC-ECs continued to proliferate through passage 9 with high purity of EC-like characteristics, while iPSC-ECs without SIRT1 overexpression became senescent after passage 5. Taken together, SIRT1 overexpression in iPSC-ECs maintains EC phenotype, improves EC function, and extends cell lifespan, overcoming critical hurdles associated with the use of iPSC-ECs in translational research.
Introduction
I
Sirtuin 1 (SIRT1) is a nicotinamide adenine dinucleotide (NAD+)-dependent histone deacetylase (HDAC) that functions in mammalian cells to promote cell survival [10] and prevent stress-induced senescence [11]. Moreover, SIRT1 plays various roles in maintaining endothelial function, including angiogenesis through deacetylation of the forkhead transcription factor (FOXO1) [12]; nitric oxide (NO) production through deacetylating and activating endothelial nitric oxide synthase (eNOS) [13]; cell proliferation by targeting the LKB1-AMPK pathway[14]; and inhibiting oxidative stress through p53 deacetylation [15,16]. Based on the various roles of SIRT1 in maintaining endothelial homeostasis, we hypothesize that overexpression of SIRT1 in iPSC-ECs would reduce senescence during in vitro culture, thereby maintaining EC phenotype and improving proliferative capacity.
Materials and Methods
Cells
All iPSC-ECs were purchased from Cellular Dynamics International (iCell Endothelial Cells, Madison, WI) and cultured as per the manufacturer's recommended protocol. Specifically, the cells were cultured in the VascuLife VEGF Medium (Lifeline Cell Technologies, Frederick, MD), supplemented with 10% iCell Endothelial Cells Medium Supplement from Cellular Dynamics International. Every 5–7 days, the cells were trypsinized and plated at 10,000 cells/cm2 on fibronectin-coated surfaces.
Lentivirus
All plasmids DNA were purchased from Addgene (Cambridge, MA). Three lentiviruses (LVs) were constructed: empty vector (LV-empty), LV with human SIRT1 (LV-SIRT1), and SIRT1H363Y (LV-SIRT1H363Y ). The SIRT1H363Y protein is a catalytically impaired variant of SIRT1 lacking the deacetylase activity [10]. SIRT1 or SIRT1H363Y was cotransfected with pWPI, pMD2.g, and psPAX2 into HEK-293FT cells [17].
Transduction
LVs (LV-empty, LV-SIRT1, and LV-SIRT1H363Y ) at MOI 5 were added to iPSC-EC at the end of P4 in the iPSC-EC growth medium for 48 h. SIRT1 and SIRT1H363Y transduction efficiency was evaluated by quantifying the percentage of cells that stained positive for human SIRT1 (Santa Cruz, Dallas, TX) using ImageJ.
Flow cytometry
EC markers CD31 (PECAM) and CD144 (VE-Cadherin) were stained with FITC-conjugated CD31 antibody (1:200 dilution; Sigma-Aldrich, St. Louis, MO) and PE-conjugated CD144 antibody (1:200 dilution; Life Technologies, Carlsbad, CA). Flow cytometry was performed using BD LSR II flow cytometer (San Jose, CA) and the data analyzed with FlowJo analytical software (Ashland, OR).
Functional analysis
The cellular senescence assay kit (Cell Signaling Technology, Danvers, MA) was used to stain for β-galactosidase (β-gal) in the presence or absence of Ex-527 (10 μM), a SIRT1 inhibitor [18], and the percentage of β-gal-positive cells was quantified with ImageJ. Tube formation assay was performed on Matrigel™ (Corning, Corning, NY) surfaces for evaluating cell angiogenesis in vitro as previously described [19]. The cells were stained with Calcein AM (Life Technologies) and the tube density quantified with ImageJ angiogenesis analyzer plugin.
The HDAC cell-based activity assay kit (Cayman Chemicals, Ann Arbor, MI) was used to assess the iPSC-EC deacetylase activity due to SIRT1 by measuring the difference in the HDAC activity between normal (0 μM Ex-527) and SIRT1-inhibited (10 μM Ex-527) culture conditions. Cell proliferation was determined by counting the cell number every 7 days after LV transduction using a hemocytometer by excluding dead cells with Trypan blue. Cell mitogenic effect in response to vascular endothelial growth factor (VEGF) (100 ng/mL) was assessed by the MTT assay (Sigma-Aldrich) after treating cells in a serum-free (starvation) medium, starvation medium containing 100 ng/mL VEGF or regular growth medium for 24 h. The NO production was assessed by 4,5-diaminofluorescein diacetate (DAF 2-DA) assay (Life Technologies). All results were normalized to cell number by the Alamar blue assay (Sigma-Aldrich).
Results
Influence of SIRT1 overexpression on EC phenotype
iPSC-ECs exhibit typical EC cobblestone-like morphology between passage 1 and 3, but gradually become fibroblast like with a decreased CD31 and CD144 expression over time (Fig. 1). A transduction efficiency of 60.3% ± 7.3% was measured through immunohistomorphometry of SIRT1-positive cells. Cells overexpressing SIRT1 exhibit a higher degree of EC-like cobblestone morphology compared to the LV-empty and LV-SIRT1H363Y
(Fig. 2). The expression of EC markers, such as CD31, was also significantly elevated in the LV-SIRT1 group (61.8% ± 3.6% CD31+) relative to controls (31.1% ± 4.5% CD31+ for LV-empty and 39.8% ±15.4% CD31+ for LV-SIRT1H363Y
) at the end of passage 5 (Fig. 2G–I). Moreover, as iPSC-ECs overexpressing SIRT1 continued to proliferate, the percentage of CD31+ cells increased from ∼60% at the end of passage 5 to ∼90% at the end of passage 7 to over 95% by the end of passage 9 (Supplementary Fig. S1; Supplementary materials are available online at
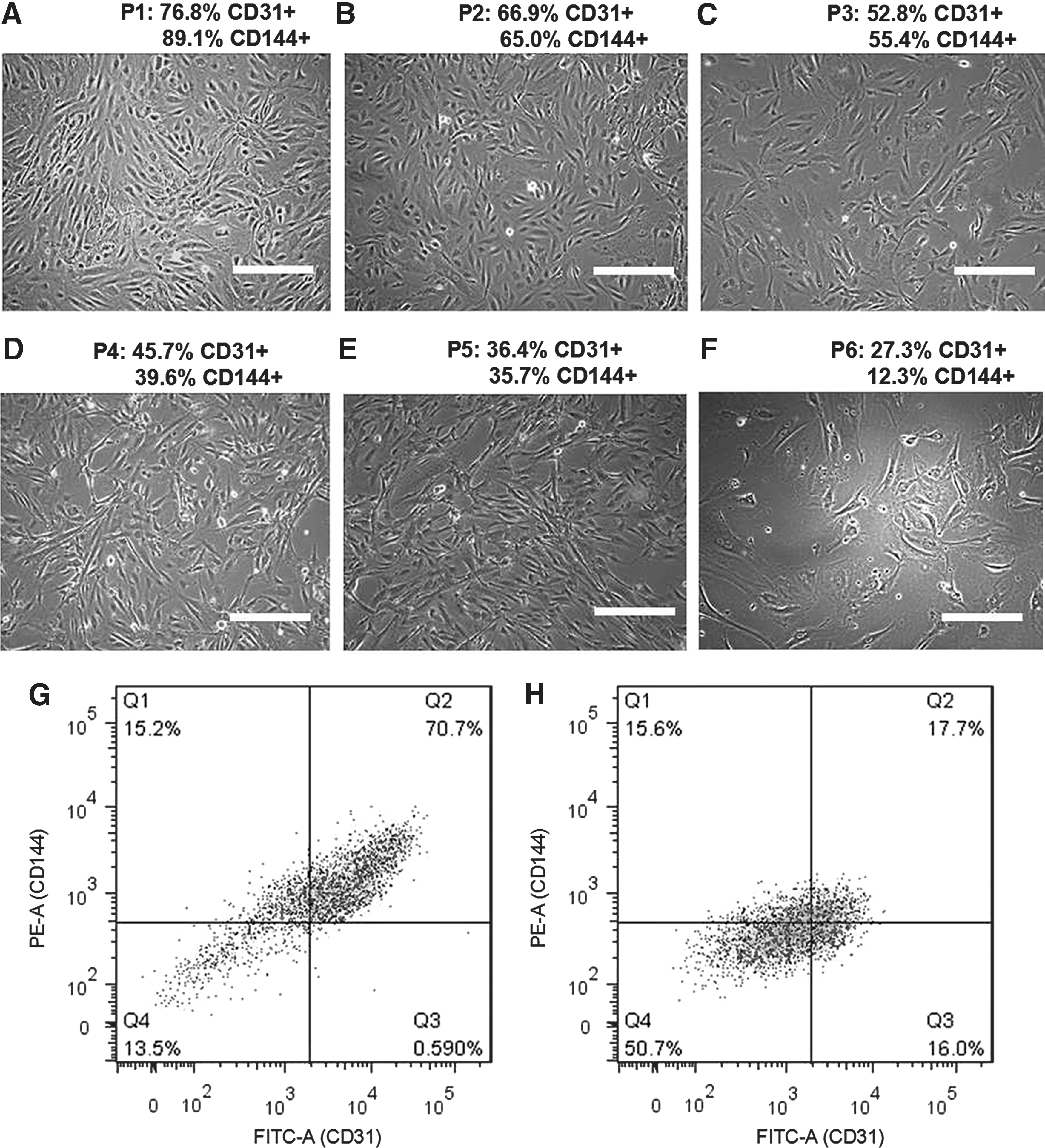
Change in morphology and phenotype of iPSC-ECs from passage 1

Effect of empty
Effect of SIRT1 overexpression on EC function
Overexpression of SIRT1 led to a significant reduction of cells entering senescence when compared to control groups (β-gal+ LV-SIRT1: 3.4% ± 2.7%, LV-empty: 38.6% ± 3.3%, and LV-SIRT1H363Y : 35.7% ± 4.9%) (Fig. 3A–C). Blocking SIRT1 with the inhibitor Ex-527 led to a higher percentage of senescent cells in all groups, suggesting a contribution of endogenous SIRT1 (β-gal+ LV-SIRT1: 49.6% ± 10.0%, LV-empty: 56.0% ± 6.2%, and LV-SIRT1 H363Y: 50.9% ± 6.8%) (Fig. 3D–F). The tube formation assay showed significantly denser and more organized vascular network formation for cells with SIRT1 overexpression (mesh area LV-SIRT1: 44.4% ± 2.3%, LV-empty: 30.9% ± 5.0%, and LV-SIRT1H363Y: 36.8% ± 3.5%) (Supplementary Fig. S2), suggesting an improved angiogenesis potential.

Cellular senescence-associated β-galactosidase (β-gal) staining (blue) of iPSC-EC at passage 6 for empty
The cell HDAC activity was significantly higher in cells with LV-SIRT1 compared to the endogenous SIRT1 HDAC activity in cells with no viral control, LV-empty and LV-SIRT1 H363Y (Fig. 4A). The rate of cell proliferation was significantly increased at two passages following viral transduction with LV-SIRT1, and these cells continued to proliferate throughout passage 10 (Fig. 4B). In contrast, cells from the control groups, including LV-SIRT1 H363Y, remained static after passage 5. VEGF stimulation led to an increased mitogenicity in iPSC-ECs overexpressing SIRT1 (Fig. 4C), and these cells also showed significantly higher NO production compared to controls (Fig. 4D).
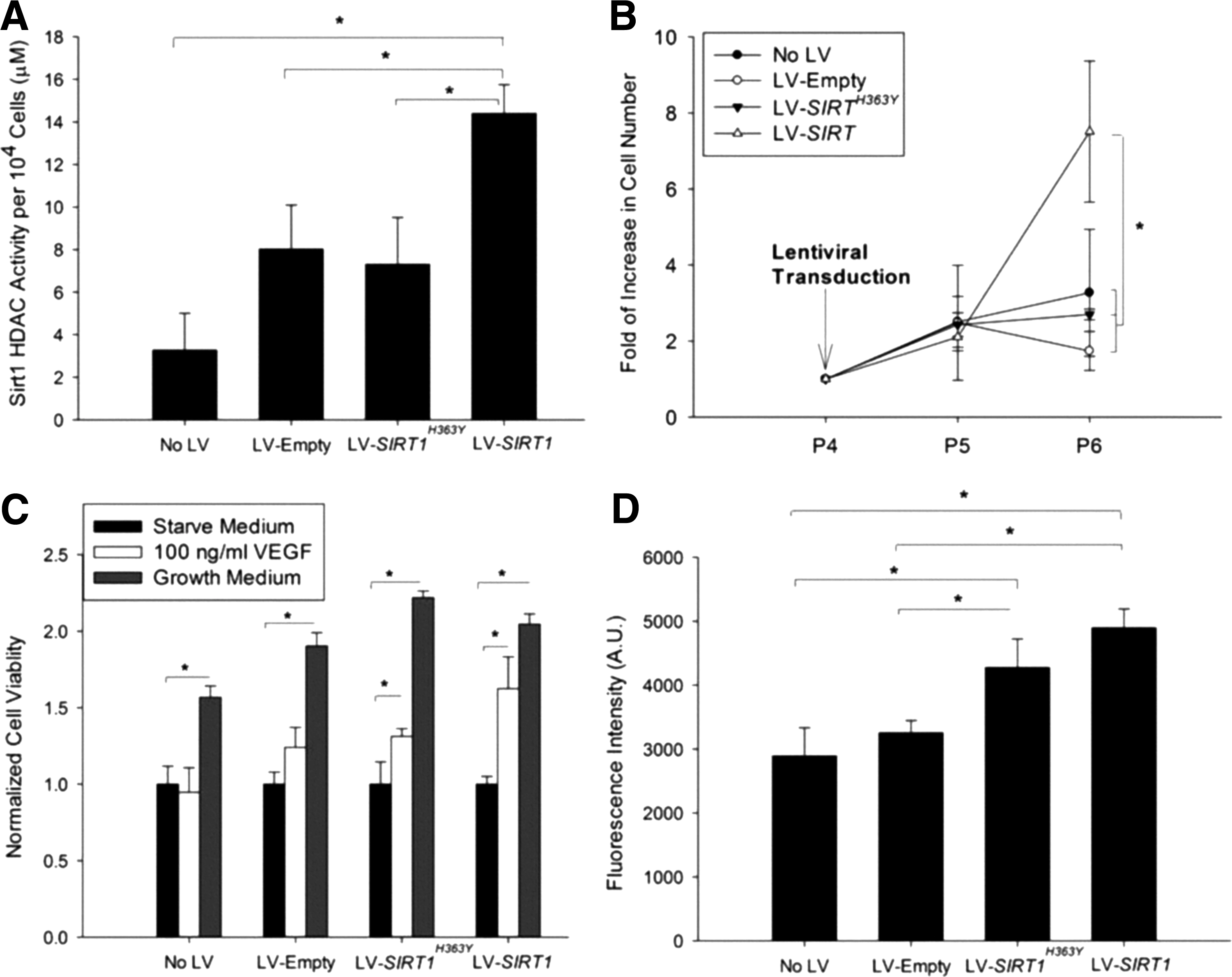
Functional assessment of iPSC-EC with or without viral transduction for
Discussion
The ability of cells derived from iPSCs to maintain a differentiated cell phenotype, function, and proliferative capacity is critical to the use of iPSC technology in disease modeling or tissue regeneration strategies. There has been significant variability regarding the in vitro culture of iPSC-ECs as researchers have reported both satisfactory [7,20] and impaired [8,9] maintenance of EC phenotype. Herein, we report a simple and effective method to prolong EC phenotype, improve cell function, and improve cell proliferation in vitro by overexpressing SIRT1.
The finding that SIRT1 overexpression improves iPSC-EC performance in our investigation agrees with studies by others on primary ECs, which may provide a mechanistic insight to our findings [14,21,22]. A previous study concluded that SIRT1 influences EC proliferation and prevents senescence by promoting the deacetylation, ubiquitination, and proteasome-mediated degradation of LKB1, a serine/threonine kinase and tumor suppressor [14]. In another study, inhibition of SIRT1 activity in ECs resulted in a premature senescent-like phenotype through an increased p53 acetylation in parallel with an increased plasminogen activator inhibitor-1 (PAI-1) expression and decreased eNOS expression [22]. These reports on primary ECs are consistent with our findings using iPSC-ECs demonstrating fewer cells entering senescence upon overexpression of SIRT1 and an increase in senescent cells when SIRT1 was inhibited (Fig. 3). SIRT1 also promotes NO production by activating eNOS through deacetylation [13]. However, how SIRT1 restores or prolongs EC phenotype, including morphology, CD31 and CD144 expression, and response to VEGF in late passage iPSC-ECs, is unclear. Surprisingly, LV-SIRT1H363Y influenced the cellular response to VEGF and NO production, although the effect was not as pronounced as those with LV-SIRT1. In summary, SIRT1 plays critical roles in maintaining iPSC-EC phenotype and function, and its overexpression may be a key step to the large-scale production and wide-scale use of these cells for translational research.
Footnotes
Acknowledgments
This work is supported by the Northwestern Memorial Foundation Dixon Translational Research Grants Initiative, the Robert R. McCormick Foundation, National Institute of Health (5R01EB017129-02 and K08DK101757), American Heart Association (AHA) Midwest Affiliate Postdoctoral Fellowship (14POST20160091), and the Chicago Biomedical Consortium (CBC) Postdoctoral Award (PDR-008).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.