
Abstract
Cell replacement therapy holds great promise for Parkinson's disease (PD), but residual undifferentiated cells and immature neural progenitors in the therapy may cause tumor formation. Although cell sorting could effectively exclude these proliferative cells, from the viewpoint of clinical application, there exists no adequate coping strategy in the case of their contamination. In this study, we analyzed a component of proliferative cells in the grafts of human-induced pluripotent stem cell-derived neural progenitors and investigated the effect of radiation therapy on tumor formation. In our differentiating protocol, analyses of neural progenitors (day 19) revealed that the proliferating cells expressed early neural markers (SOX1, PAX6) or a dopaminergic neuron progenitor marker (FOXA2). When grafted into the rat striatum, these immature neurons gradually became postmitotic in the brain, and the rosette structures disappeared at 14 weeks. However, at 4–8 weeks, the SOX1+PAX6+ cells formed rosette structures in the grafts, suggesting their tumorigenic potential. Therefore, to develop a fail-safe therapy against tumor formation, we investigated the effect of radiation therapy. At 4 weeks posttransplantation, when KI67+ cells comprised the highest ratio, radiation therapy with 137Cs Gammacell Exactor for tumor-bearing immunodeficient rats showed a significant decrease in graft volume and percentage of SOX1+KI67+ cells in the graft, thus demonstrating the preventive effect of gamma-ray irradiation against tumorigenicity. These results give us critical criteria for the safety of future cell replacement therapy for PD.
Introduction
P
One of the major concerns in cell transplantation is tumor formation. When neural cells are induced from embryonic stem cells (ESCs) or induced pluripotent stem cells (iPSCs), a component of the differentiated cells is heterogeneous, possibly containing undifferentiated stem cells [1]. Tumor formation has been reported in the transplantation of ESC/iPSC-derived neural cells induced on stromal feeder cells [2] or extracellular matrix [3]. Other studies showed that residual undifferentiated ESCs/iPSCs or immature progenitors in the grafted cells form tumors after transplantation [4 –9]. Residual undifferentiated ESCs/iPSCs form nonneural tissues or teratomas in the host brain or they differentiate into neuroectodermal cells [5]. At the same time, another type of tumor is formed by early neural progenitors. Characteristic rosette structures are observed in this type of tumor, and the cells in these rosettes are reported to express neural stem cell (NESTIN) [6] and early neural (PAX6) markers [7]. A comprehensive in vitro gene expression analysis revealed that highly proliferative cells of forebrain origin constructed these rosettes and contributed to tumor formation or neural overgrowth [7].
To prevent tumor formation, several approaches have been attempted: prolonged differentiation [2,6,9], gamma-secretase inhibition [7,10,11], and cell sorting [12 –15]. The first two facilitate the maturation of DA progenitors, while cell sorting is effective at excluding undifferentiated cells and immature progenitors. To enrich DA neuron progenitors and exclude tumorigenic cells, we have developed a cell sorting protocol using a floor plate cell surface marker, CORIN, and proved a suppressive effect against tumorigenesis [15].
Although we have never observed tumor formation with Corin-sorted cells, multilayered measures are needed in the case of clinical application. Therefore, it is important to identify which cellular components may cause tumors and establish an appropriate fail-safe therapy to take measures against them. Radiation therapy is an effective treatment for brain tumors. Previous reports revealed that irradiation induces apoptosis of human neural stem cells and inhibits cell cycle progression through oxidative stress-related DNA damage [16 –18]. In the viewpoint of cell transplantation, however, the preventive effect of irradiation against tumorigenesis has only been investigated for pretreatment before transplantation [19,20], but never for grafted cells.
To identify tumorigenic components in human iPSC (hiPSC)-derived neural cells and determine whether irradiation can prevent tumor formation by these cells, we induced neural cells from hiPSCs and grafted them into the striatum of immunodeficient rats. Immunohistological analyses revealed that the remaining neuroepithelial cells in the graft continued proliferation in the brain. In addition, their proliferation was reduced by gamma-ray irradiation.
Materials and Methods
Human iPS cell culture
Two human iPSC lines (201B7, reprogrammed by retrovirus vectors, shown in Figs. 1
–6, Supplementary Figs. S2 and S5 [Supplementary Data are available online at
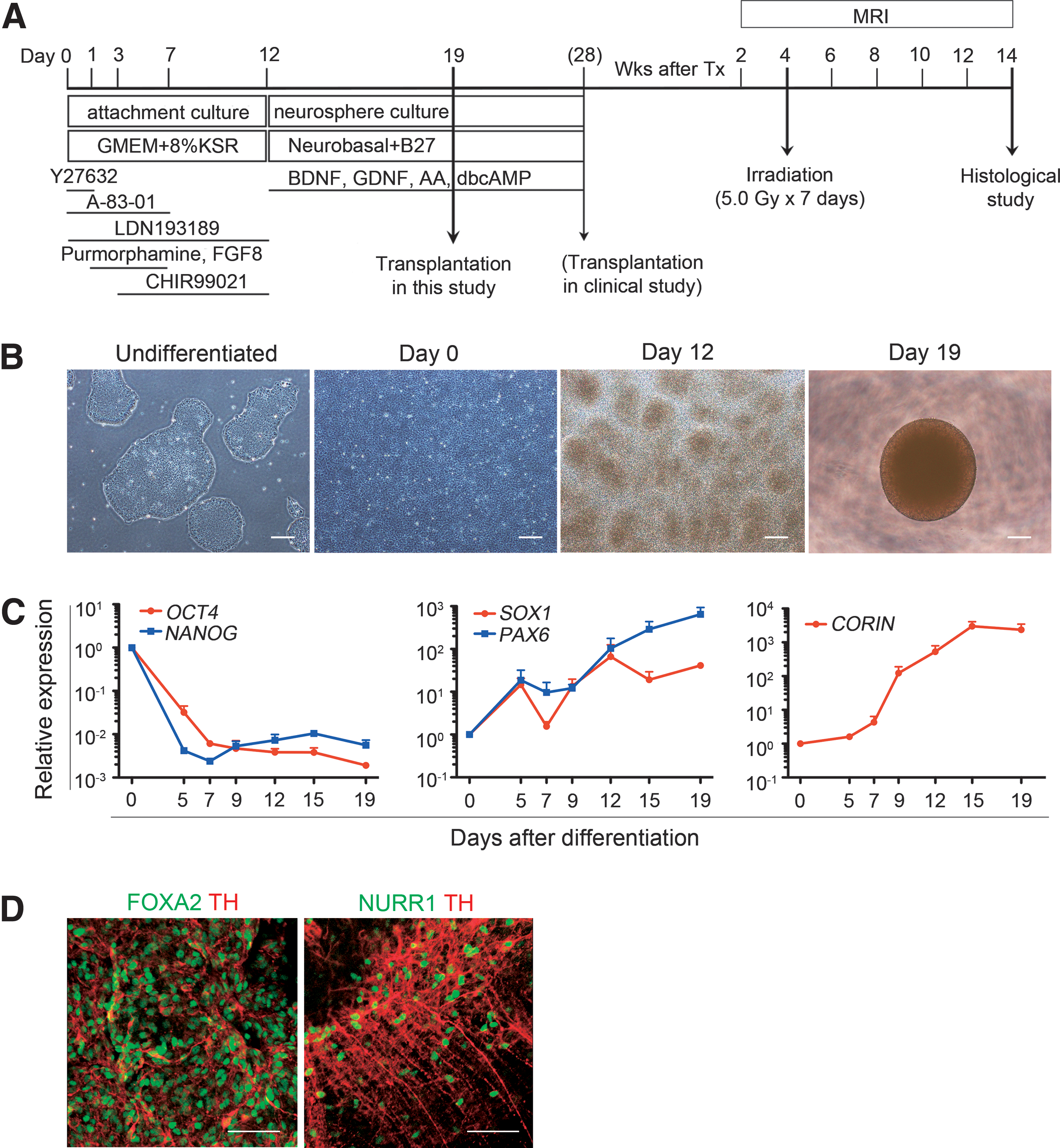
Differentiation of hiPSC-derived midbrain dopaminergic neuron progenitors.
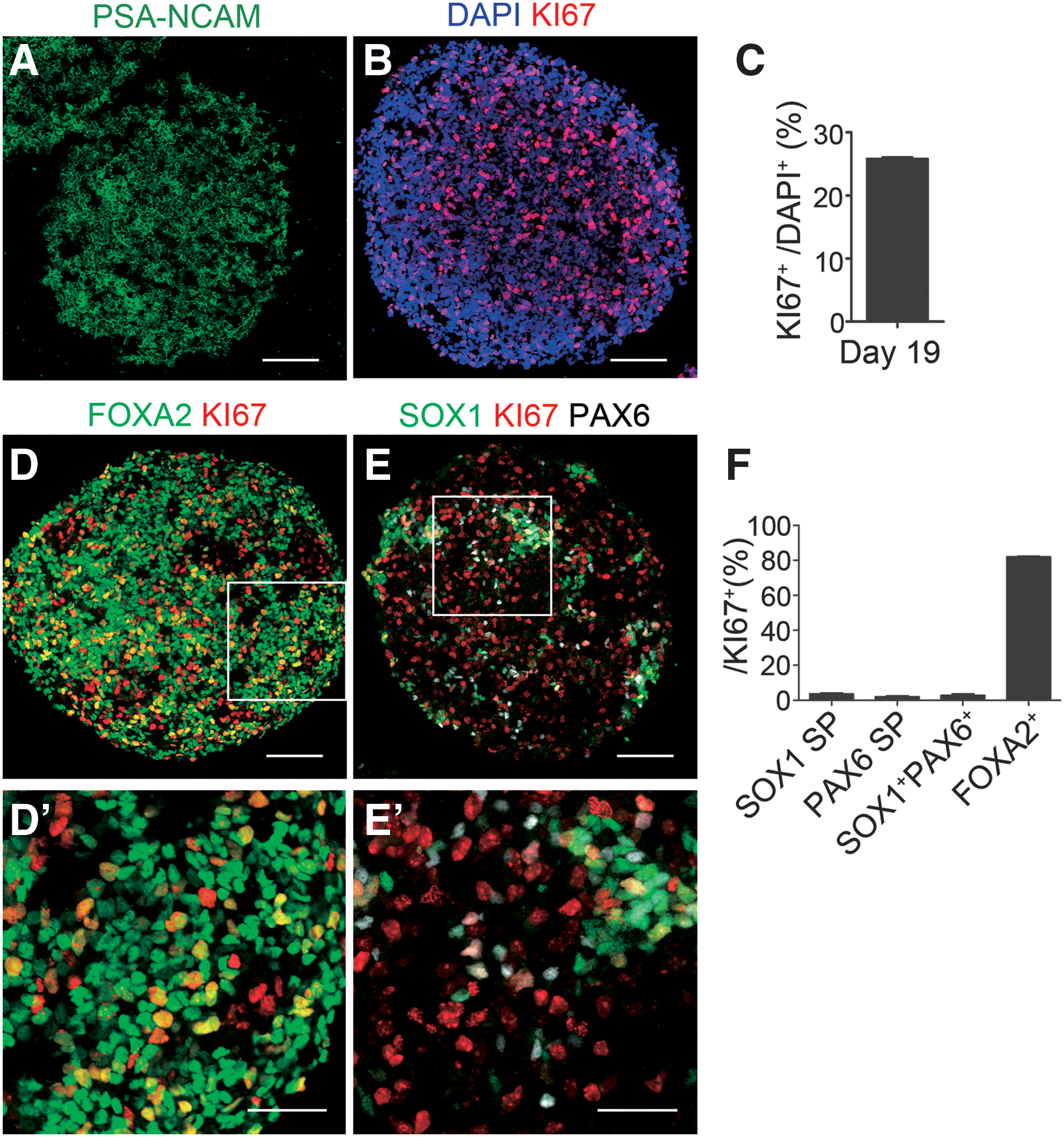
Immunocytochemical analysis of d19 spheres.
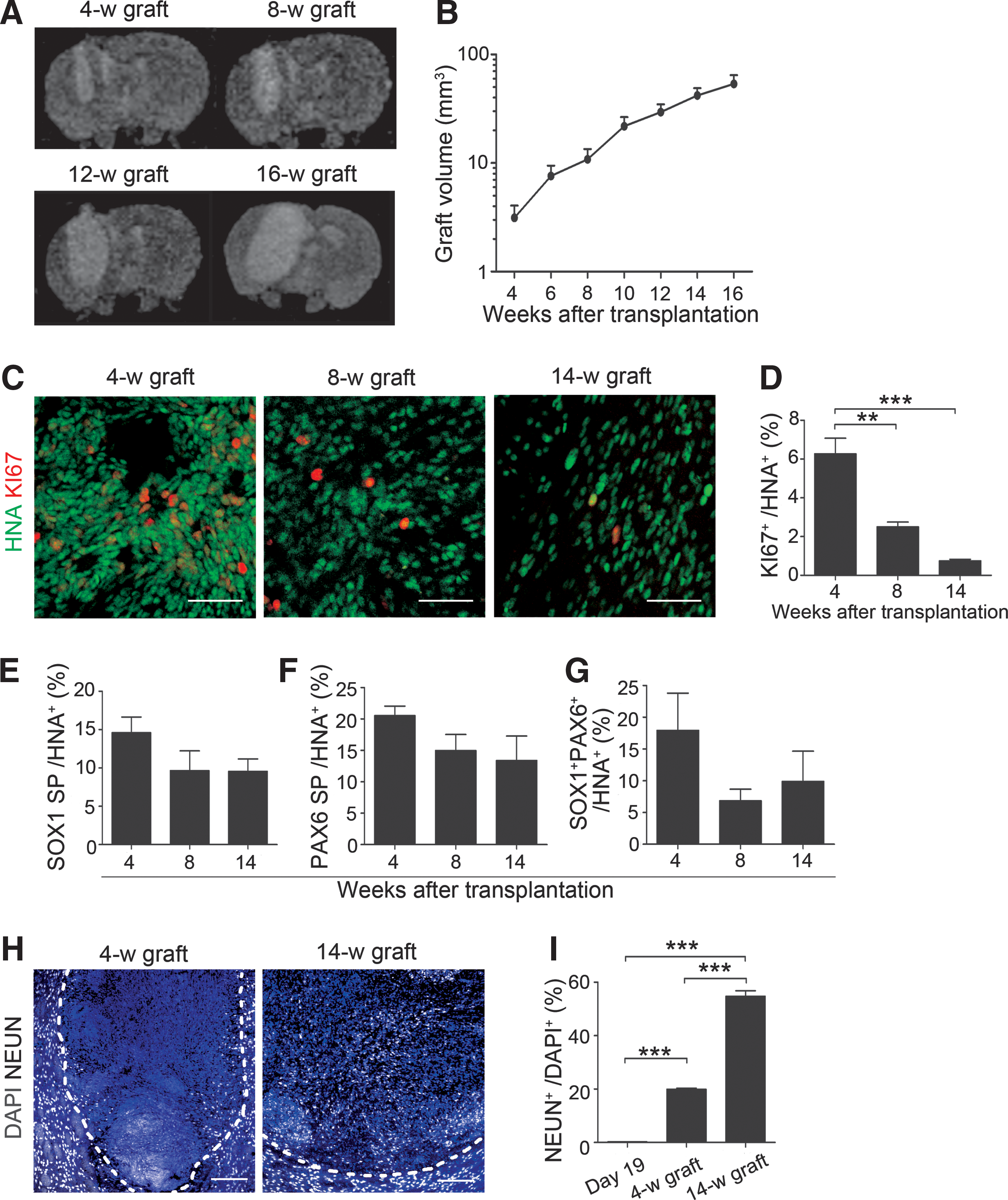
Graft volume and KI67+ cells.
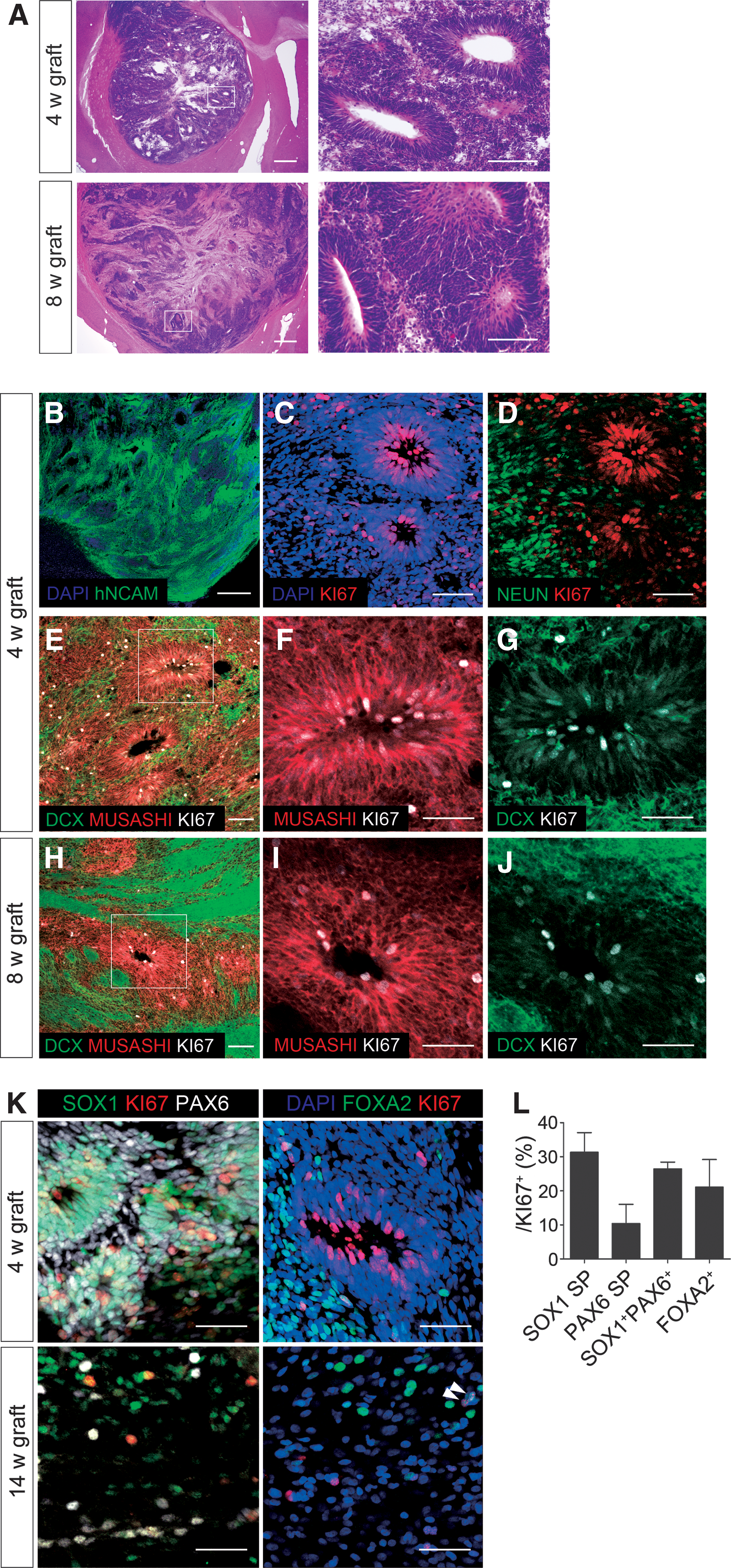
Histological analysis of tumorigenic grafts.
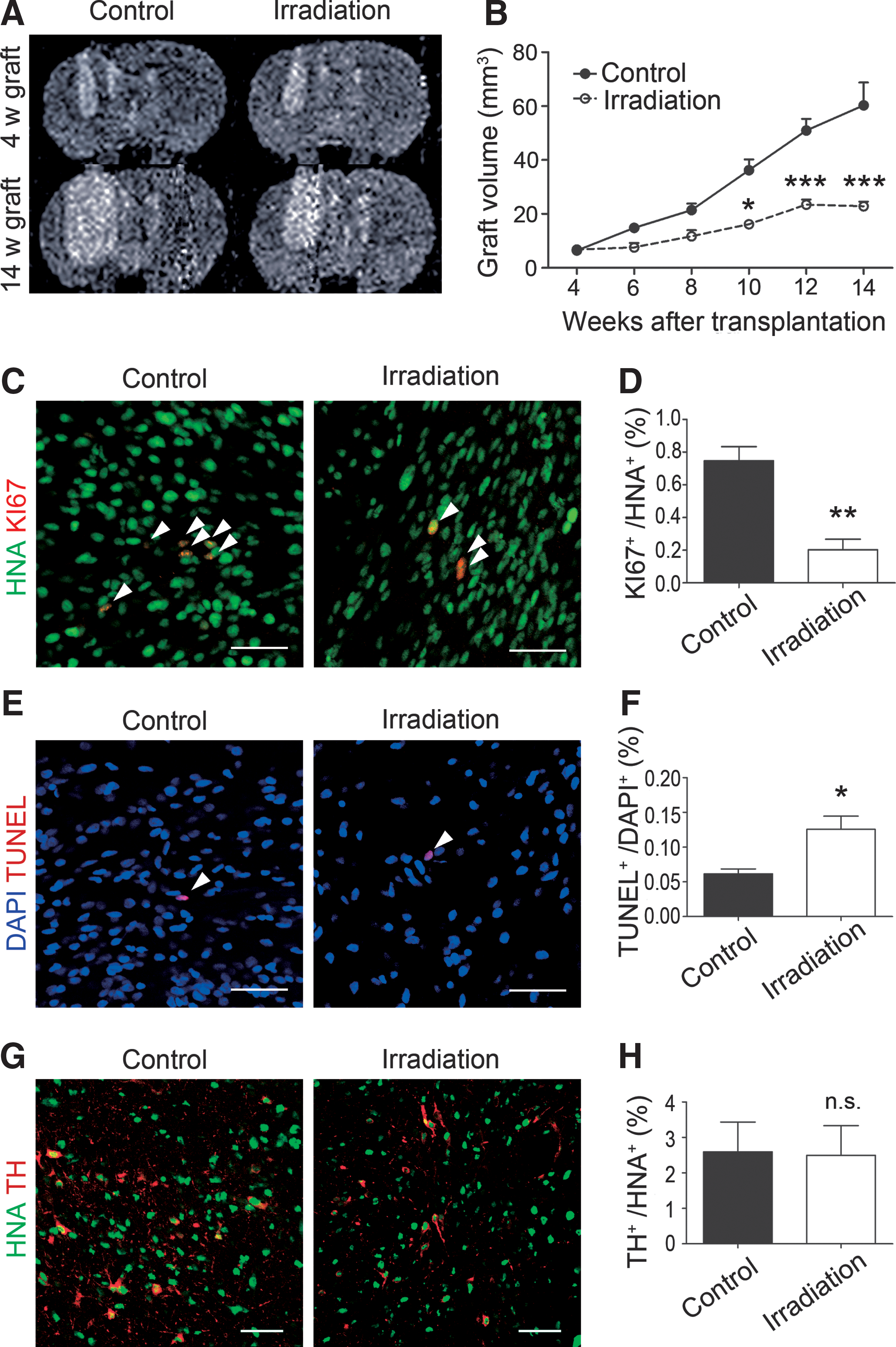
Suppressive effect of gamma-ray radiation against tumor formation.
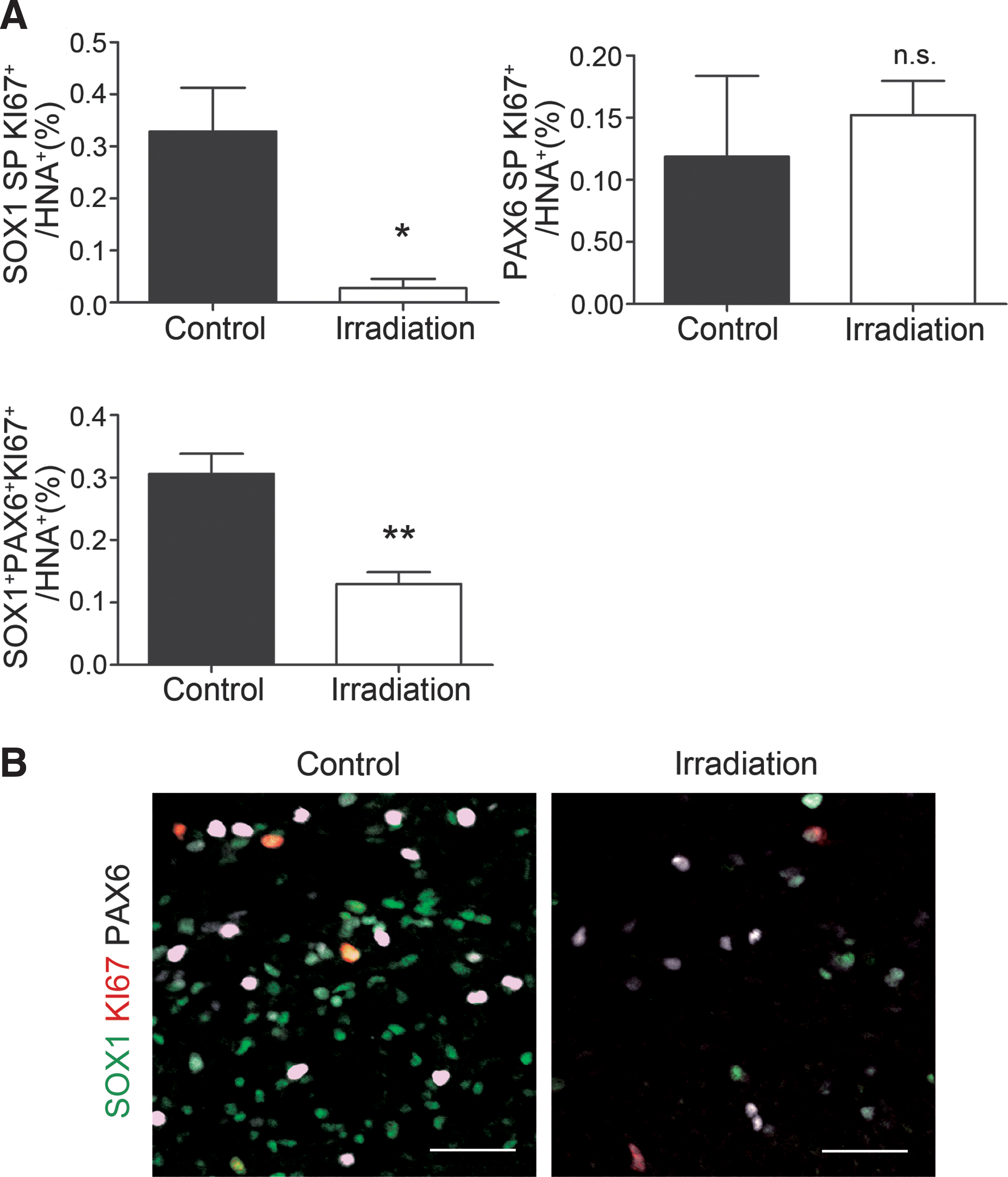
Effects of gamma-ray radiation on the proliferation of neural progenitors.
Neural differentiation
Differentiation was performed according to our previous study [15]. Undifferentiated cells were split to six-well plates at a density of 4 × 105 cells/well. After reaching confluence (day 0), ACiM media were changed to differentiation media containing Glasgow minimum essential medium (GMEM) supplemented with 8% knockout serum replacement (KSR), 0.1 mM MEM nonessential amino acids (all Invitrogen), 0.1 mM sodium pyruvate (Sigma-Aldrich, St. Louis, MO), and 0.1 mM 2-mercaptoethanol (WAKO Pure Chemical Industries). From day 0, 0.1 μM LDN193189 (STEMGENT, Lexington, MA) and 0.5 μM A-83–01, from day 1, 2 μM Purmorphamine and 100 ng/mL FGF 8, and from day 3, 3 μM CHIR99021 were added to the media (all WAKO Pure Chemical Industries). For neurosphere formation, on day 12, neural progenitors were dissociated into single cells with Accumax (Innovative Cell Technologies, San Diego, CA) and 30 μM Y-27632 and replated at a density of 2 × 104 cells/well on low-attachment 96-well plates (Lipidure-Coat Plate A-96U; NOF Corporation, Tokyo, Japan) in Neurobasal medium supplemented with B-27 supplement (Invitrogen), 2 mM
Cell sorting and flow cytometry
Cell sorting and flow cytometry were performed with FACSAria II Cell Sorter (BD Biosciences, San Jose, CA) using FACSDiva software (BD Biosciences) [15]. For cell sorting, on day 12 (d12), cells were dissociated with TrypLE select and stained with PE conjugated mouse anti-CORIN IgG antibody (1:5,000) at 4°C for 20 min. Before sorting, the cell suspension was filtrated through a 35-μm cell strainer (BD Falcon, Franklin Lakes, NJ), and 7-AAD (BD Pharmingen, San Diego, CA) was added to each tube for dead cell exclusion. For flow cytometric analysis, dissociated cells were stained with an Alexa 647-conjugated anti-Oct3/4 antibody (BD Pharmingen) at 4°C for 20 min.
Quantitative PCR
Total RNA was extracted with the RNA Micro Kit (QIAGEN, Venlo, Netherlands), and 1 μg was synthesized into cDNA with Super Script III First-Strand Synthesis System and Oligo(dT)20 primer (both Invitrogen) according to the manufacturers' instruction. Quantitative PCR was performed with SYBR Premix Ex Taq (Takara Bio, Shiga, Japan) and Thermal Cycle Dice Real Time System (Takara Bio). For data analysis, the delta-Ct method normalized by the GAPDH gene was applied for three independent experiments. Primer sequences are shown in Supplementary Table S1.
Animals and cell transplantation
Animals were cared for and handled according to the Guidelines for Animal Experiments of Kyoto University and the Guide for the Care and Use of Laboratory Animals of the Institute of Laboratory Animal Resources (ILAR; Washington, DC). X-linked severe combined immunodeficiency (X-SCID) rats, F344-Il2rgem2Kyo (NBRP-Rat No. 0586), were provided from the National BioResource Project - Rat, Kyoto University (Kyoto, Japan), and inbred in our laboratory [23]. Ten-week-old male rats weighing 180–220 g were chosen for the cell transplantation. d19 or d28 spheres (4 × 105 cells in 2 μL, 1 μL/10 s) were loaded in a 22-gauge needle and stereotactically injected into the right striatum (from the bregma: A +0.5, L + 2.0, V + 3.0 and +2.5, incisor bar 0) under inhalation anesthesia. From 4 to 24 weeks posttransplantation, animals were euthanized with sodium pentobarbital for perfusion fixation with 4% paraformaldehyde. Brains kept in fixation overnight were sequentially transferred into 15–30% sucrose solution before being cut into 30-μm-thick sections on a cryostat microtome. Free floating sections were studied with H&E staining and immunostaining.
Radiation therapy
Whole brain gamma-ray irradiation was performed with 137Cs (Gammacell 40 Exactor, MDS Nordion, Ottawa, ON, Canada). Animals in plastic restraint bags were carefully laid on a chamber to set their head position beneath a collimator hole. For the irradiated group, 5.0 Gy gamma-ray (1.0 Gy/min) was irradiated for 7 days (35.0 Gy in total). The unirradiated group was restrained and laid on a chamber for 5 min.
Magnetic resonance imaging
Magnetic resonance imaging (MRI) was performed every 2 weeks after transplantation. Under inhalation anesthesia, T2 -weighted coronal brain images were acquired using a 1.5-T MR unit (MRmini SA, DS Pharma Biomedical, Osaka, Japan). The parameters of the two-dimensional spin-echo sequence were as follows: repetition time = 2000 ms, echo time = 69 ms, field of view = 30 mm, voxel size = 0.234 × 0.234 × 1.00 mm, and matrix size = 256 × 128 × 20 slices with 1-mm thickness. The graft volume was calculated by 3D DICOM Viewer (INTAGE Realia Professional, Cybernet Systems, Tokyo, Japan).
Immunostaining
After fixation with 4% paraformaldehyde for 10 min at 4°C, in vitro samples were used for immunostaining. d19 spheres were sectioned at 20-μm thickness on the cryostat microtome. Both in vitro and in vivo samples were washed with phosphate-buffered saline and permeabilized with 0.3% TritonX-100 solution for 15 min. Before blocking with 2% skim milk and 0.3% TritonX-100 solution, the antigen retrieval step with a microwave oven for 5 min was needed for some antibodies (SOX1, PAX6, and KI67). Samples were incubated at 4°C overnight with primary antibodies and then incubated with Alexa Fluor 488-, 594-, or 647-conjugated secondary antibodies (1:400 dilution; Invitrogen) for 60 min at room temperature. Nuclei were stained with 4′,6-diamidino-2-phenylindole (DAPI; Invitrogen). Primary antibodies are shown in Supplementary Table S2. Images were obtained with BIOREVO BZ-9000 (Keyence, Osaka, Japan) or Fluoview FV1000D (Olympus, Tokyo, Japan). Five high-magnification fields (approximately 500 cells/field) were analyzed for quantification. To quantify the percentage of KI67+ cells that were SOX1+, PAX6+, or FOXA2+, at least 100 KI67+ cells were counted for each specimen.
Statistical analysis
Statistical analysis was performed with GraphPad Prism5 (GraphPad Software, La Jolla, CA), and the significance level was set to P < 0.05. Nonparametric one-way ANOVA and Tukey's post hoc test were chosen for histological evaluation. Two-way ANOVA and the Bonferroni correction were used for temporal evaluation of the graft volume. All data are shown as mean ± SEM.
Results
Neural differentiation into midbrain DA neuron progenitors from human iPSCs
Human iPSCs (201B7) were differentiated into midbrain DA neurons on human laminin 511-E8 fragment according to our current protocol [15] (Fig. 1A), which is based on dual SMAD inhibition [24,25] and floor plate induction [26]. In this protocol, which has never been observed to cause tumors in the brain, we isolate cells expressing CORIN, a floor plate marker, by fluorescence-activated cell sorting (FACS) on day 12, and incubate the sorted cells until day 28 [15]. The d28 cells, even unsorted, showed less expansion than the d19 cells and did not always reach sufficient size (Ca >20 mm3) to evaluate the effect of gamma-ray irradiation (Supplementary Fig. S1A, B) [15]. Therefore, to observe tumor formation, we used d19 unsorted cells in this study (Fig. 1B).
A comparative temporal gene expression analysis revealed that the expression of pluripotent stem cell markers (OCT4, NANOG) gradually decreased to less than one hundredth their initial expression by day 12 (Fig. 1C). Early neural cell markers (SOX1, PAX6) gradually increased by day 19. The expression of CORIN increased after day 9 and reached a plateau by day 19. The expressions of mesodermal (BRACHYURY), endodermal (SOX17), and neural crest (SOX10, AP2) markers remained low and were equivalent to those of undifferentiated iPSCs (data not shown).
To evaluate the potential of the d19 spheres to become midbrain DA neurons, the floating spheres were replated on poly-L-ornithine/fibronectin/laminin-coated chamber slides and incubated until day 42. An immunofluorescence study revealed the coexpression of markers for postmitotic DA neurons (TH) and midbrain (FOXA2, NURRl) (Fig. 1D).
These results indicated that our protocol induced neural cells, including progenitors of midbrain DA neurons from hiPSCs.
Characterization of proliferating cells in d19 spheres in vitro
Next, we investigated the proliferating cells in the d19 spheres by an immunocytochemical analysis. No cells expressed pluripotent stem cell markers (OCT4, NANOG or TRA-1–60), but all expressed PSA-NCAM, a neural progenitor marker (Fig. 2A). We also confirmed that there were no OCT4+ cells in the d19 spheres by a flow cytometric analysis (Supplementary Fig. S2). The percentage of proliferating (KI67+) cells per total (DAPI+) cells was 25.8% ± 0.2% (Fig. 2B, C), and the majority of KI67+ cells expressed a floor plate marker, FOXA2 (Fig. 2D). Because DA neurons originate in the floor plate of the developing ventral midbrain and FOXA2 is specifically expressed in adult DA neurons and their progenitors in the medial floor plate [27 –30], we assumed KI67+FOXA2+ cells to be DA progenitor cells.
A neural rosette is a typical feature of graft overgrowth. In previous reports, neural rosette-forming cells expressed early neural markers such as SOX1 and PAX6 in vitro [7,31 –33]. Both SOX1 and PAX6 control early neural specification. At E8.0, the expression of SOX1 is restricted to neuroepithelium of early neural plate and neural tube [34,35] and blocks neural differentiation [36]. At E8.5, PAX6 expression appears in neuroepithelium of the forebrain and hindbrain [37] and promotes differentiation toward radial glia [36]. In this study, d19 spheres also contained cells expressing these markers (Fig. 2E). The percentages of neuroepithelium (SOX1 single positive), radial glia (PAX6 single positive), and neural rosettes (SOX1+PAX6+) per DAPI+ cells were 2.4% ± 0.1%, 4.5% ± 0.9%, and 3.3% ± 1.2%, respectively. Among KI67+ cells, the percentages of SOX1 single positive, PAX6 single positive, SOX1+PAX6+, and FOXA2+ were 3.6% ± 0.4%, 2.0% ± 0.5%, 2.7% ± 0.9%, and 81.9% ± 0.4%, respectively (Fig. 2F). These percentages suggested that most of the proliferating cells in the d19 spheres were DA progenitor cells.
Grafts derived from d19 spheres gradually expanded, and early neural cells contributed
To examine proliferative potential in the brain, d19 spheres (4.4 × 105 cells) were transplanted into the right striatum of X-SCID rats. MRI analyses showed that the graft volume gradually increased over 16 weeks after transplantation (Fig. 3A). The largest change in volume was observed between 4 and 6 weeks (2.8 ± 0.3-fold), but became comparatively mild from 10 weeks onward (Fig. 3B). We evaluated the percentage of KI67+ cells in the human nuclear antigen (HNA)+-grafted cells by immunohistochemistry at 4, 8, and 14 weeks posttransplantation and found a significant decrease over the course (Fig. 3C, D). Furthermore, the percentage of early neural cells (SOX1 single positive, PAX6 single positive and SOX1+PAX6+) decreased between 4 and 8 weeks (Fig. 3E–G) and that of neuronal nuclei (NEUN)+-postmitotic neurons increased between 4 and 14 weeks (Fig. 3H, I). These results suggested that maturation of the grafted cells toward postmitotic neurons proceeded in the brain.
Next, to examine the identity of the proliferating cells, we performed histological analyses of the grafts. First, at any time point, we did not observe the expression of pluripotent (OCT4, NANOG), mesodermal (PECAM-1), endodermal (PAN-CYTOKERATIN), neural crest (p75, AP2), or epithelial (PAN-CYTOKERATIN) markers, suggesting that the grafted cells were already committed to neural lineage. Consistently, H&E staining showed rosette formation in the graft (Fig. 4A), and all of the cells in the grafts were positive for human NCAM (Fig. 4B). The rosette-forming cells expressed KI67 and MUSASHI, a neural stem cell marker. As was expected, KI67 staining was not observed in surrounding newborn (DOUBLECORTIN+) or mature (NEUN+) neurons (Fig. 4C–G). Consistent with neural differentiation, the number of KI67+ and MUSASHI+ cells decreased and that of DOUBLECORTIN+ cells increased at 8 weeks (Fig. 4H–J). Moreover, consistent with in vitro results, KI67+ cells in these rosettes expressed SOX1 and PAX6, but were negative for FOXA2 (Fig. 4K and Supplementary Fig. S1C–F). At 14 weeks posttransplantation, we could no longer observe rosettes and found only scattered KI67+ cells, but 26.4% ± 1.9% of KI67+ cells still expressed both SOX1 and PAX6 (Fig. 4K, L). The remaining KI67+ cells expressed FOXA2 (21.0% ± 8.0%), suggesting that they were DA neuron progenitors (Fig. 4L). Taken together, rosette-forming cells (SOX1+PAX6+) mainly contributed to the graft expansion as they continued proliferation in the host brain.
Suppressive effect of gamma-ray irradiation against tumor formation
Currently, no in vivo gamma-ray irradiation study against hiPSC-derived neural progenitors has been reported. Therefore, to examine the effect of radiation therapy, we grafted the d19 spheres (4.4 × 105 cells) into the right striatum of X-SCID rats and performed whole brain gamma-ray irradiation at 4 weeks. In the case of a C6 rat glioma model, radiosurgery at a margin dose of 35 Gy, which is equivalent to a clinical dose, effectively suppressed tumorigenic overgrowth [38]. In addition, an in vitro study reported that a radiation dose of 5 Gy was able to inhibit the proliferation of hiPSC-derived neural stem cells [17]. Based on these findings, we decided to perform radiation therapy with a total dose of 35 Gy in 7 days (5 Gy/day).
An MRI analysis revealed that the increase in graft volume was significantly suppressed in the irradiation group (Fig. 5A, B). In addition, immunofluorescence revealed that the percentage of KI67+ cells per grafted cells also significantly decreased (Fig. 5C, D). On the contrary, a TUNEL staining of 14-week grafts revealed that the percentage of apoptotic cells in the irradiation group was significantly higher than that in the control group (Fig. 5E, F), indicating that proliferating cells underwent apoptosis by the gamma-ray irradiation. The percentage of TH+-postmitotic DA neurons, however, did not differ between the control and irradiation groups (Fig. 5G, H), suggesting that a dose of 35 Gy did not affect survival or differentiation of the grafted DA neuron progenitors. To confirm the effect of gamma-ray irradiation, we transplanted d19 spheres derived from another hiPSC line (1147F1) the same way and observed again that gamma-ray irradiation at both 4 and 8 weeks posttransplantation significantly suppressed the graft overgrowth (Supplementary Fig. S3 and S4A–D).
Next, to determine the cell population which was affected by the gamma-ray irradiation, we examined the expression of proliferation (KI67) and early neural markers (SOX1, PAX6) by immunofluorescence at 14 weeks. Gamma-ray irradiation significantly decreased the percentage of proliferating SOX1 single positive (neuroepithelium) and SOX1+PAX6+ (neural rosettes) cells in the graft, but showed no effect on proliferating PAX6 single positive cells (radial glia) (Fig. 6 and Supplementary Fig. S4E–H).
Taken together, SOX1+PAX6+ early neural cells formed rosettes in the grafts, and most of the proliferating cells were localized in the rosettes, suggesting that SOX1+PAX6+ cells mainly contribute to graft expansion in the brain. Furthermore, SOX1+ cells are radiosensitive, and suppression of their proliferation contributes to the reduction of tumorigenic overgrowth by hiPSC-derived neural progenitors.
Discussion
Previous studies have indicated possible tumor formation by ES/iPSC-derived neural progenitors, in which rosette structures composed of early neural cells were often observed [5 –7]. However, a detailed temporal analysis of the cell components that contribute to the tumor formation has never been reported. In this study, we show that differentiation of hiPSC-derived neural progenitors continues in the brain and the proliferation of early neural cells, which express SOX1 and PAX6, forms neural rosettes and contributes to the tumor formation. In addition, we clearly demonstrated that gamma-ray irradiation reduced the proliferation of SOX1+ cells, which resulted in the reduction of the graft size.
Previous studies about the transplantation of ESC-derived neural progenitors into the brain have reported that the proliferation of the grafted cells decreases over time [2,6,39]. In the case of human ESC-derived neural progenitors in rat PD models, the percentage of proliferating cells decreased from 12% to 2% between 2 and 13 weeks posttransplantation and from 11% to 1% between 4 to 20 weeks posttransplantation [2,39]. Long-term observation of PD model monkeys found hESC-derived DA progenitors had a decreasing growth rate with time after transplantation [6]. Consistent with these previous findings, our data showed that cell proliferation decreased with neuronal maturation in the brain over time (Fig. 3). In addition, neural rosettes disappeared at 14 weeks posttransplantation (Fig. 4K). These results suggest that if appropriately committed to neural lineage, the grafted neural progenitors will differentiate into postmitotic mature neurons intrinsically or in response to external stimuli.
Histology of unsorted d19 spheres in vitro and in vivo showed that proliferating cells are SOX1 single positive, PAX6 single positive, SOX1+PAX6+, or FOXA2+ (Figs. 2 and 4). Considering that all these cells were PSA-NCAM+ (Fig. 2A) and ventral midbrain cells are predominantly induced in our protocol [15], we regarded these cells as neuroepithelial cells, radial glia cells, neural rosette cells, and DA progenitor cells, respectively [36,40]. Importantly, the expression of a pluripotent stem cell marker (OCT4) in the d19 spheres was almost undetectable before transplantation (Supplementary Fig. S2), and we could not confirm the expression of pluripotent, mesodermal, endodermal, neural crest, or epithelial markers. These results suggested that there were no remaining iPSCs in the unsorted d19 spheres. Before transplantation, about 80% of the proliferating cells were FOXA2+ DA progenitors (Fig. 2F). In contrast, about 80% of the proliferating cells in the graft at 14 weeks expressed either SOX1 or PAX6, or both (Fig. 4L). In addition, rosette-forming cells did not express FOXA2, but did express SOX1 and PAX6. All these results indicate that SOX1+PAX6+ neural rosette cells were the main contributors of the tumor formation by the hiPSC-derived grafts.
To investigate the cell components responsible for tumor formation, we used unsorted d19 spheres. For clinical trial, however, we are planning to transplant sorted CORIN+ cells on day 28, as these cells do not form tumors in rat brains [15]. In a developing mouse midbrain, the region and time of the expression of CORIN is apparently different from that of SOX1 [41,42] and PAX6 [43]. CORIN, a floor plate marker, is selectively expressed in ventral midline cells from midbrain to the spinal cord and is first expressed at E9.75 in mouse embryos [28]. In addition, our protocol predominantly induces ventral midbrain cells [15]. Thus, CORIN+ cells in our protocol are assumed to be negative for SOX1 and PAX6. Consistently, we confirmed by immunostaining that the purification of CORIN+ cells by FACS eliminates both SOX1+ and PAX6+ cells from d19 spheres (Supplementary Fig. S5). The remaining proliferating cells are FOXA2+ DA neuron progenitors, which eventually become mature DA neurons and do not contribute to tumor formation in the brain [15].
As a fail-safe therapy, we determined whether the graft expansion could be reduced by gamma-ray irradiation. A suppressive effect of the irradiation on tumorigenesis was clearly obtained, because the graft volume and the percentage of KI67+ cells in the graft were significantly decreased in the irradiated group. Consistently, apoptotic cells were increased in the irradiated group (Fig. 5E, F). An immunohistological analysis revealed that the proliferation of SOX1 single positive and SOX1+PAX6+ cells was significantly reduced by gamma-ray irradiation. On the other hand, the percentage of proliferating PAX6 single positive cells showed no significant decrease (Fig. 6). The percentage of SOX1 single positive cells per KI67+ cells in the graft at 8 weeks was higher than at 4 weeks (Supplementary Fig. S1C–F). Gamma-ray irradiation at 8 weeks posttransplantation was also effective at reducing the proliferation of SOX1 single positive cells, SOX1+PAX6+ cells, and the graft volume (Supplementary Fig. S4). These results suggest that the main factor of possible tumorigenicity was not PAX6+ cells, but rather SOX1+ cells. The reason can be attributed to differences in cell cycle length, which is shorter in neuroepithelium (SOX1 single positive) or neural rosettes (SOX1+PAX6+) under proliferative division than in radial glia (PAX6 single positive) under neuron-generating division [36,44]. In addition, PAX6 single positive cells are proliferating even in the adult brain, especially in neurogenic niches such as hippocampal dentate gyrus [45].
For the gamma-ray irradiation in this study, we employed 35 Gy (5 Gy for 7 days) as a total radiation dose, based on previous studies [17,38]. For whole brain radiation therapy in a clinical situation, 5 Gy per fraction is quite high. However, we are assuming gamma knife radiosurgery that focally delivers higher doses (10–25 Gy) at once [46 –48]. It is noteworthy that the survival and differentiation of the grafted DA neuron progenitors were not affected by 35 Gy (5 Gy for 7 days) gamma-ray irradiation.
In conclusion, this study revealed that SOX1+ and PAX6+ cells form rosette structures in hiPSC-derived neural grafts and possibly contribute to tumor formation. Gamma-ray irradiation was effective at reducing the graft volume by decreasing the proliferation of SOX1+ cells. We also demonstrated the importance of sorting CORIN+ cells not only to enrich DA progenitors but also to eliminate SOX1+ and PAX6+ cells. These results will contribute to the safety of cell replacement therapy using hiPSCs for Parkinson's disease.
Footnotes
Acknowledgments
We would like to thank Dr. Kaneyasu Nishimura for valuable discussions and Dr. Peter Karagiannis for critical reading of the article. We also thank Kei Kubota for breeding of X-SCID rats. This study was supported by a grant from the Network Program for Realization of Regenerative Medicine from the Japan Agency for Medical Research and Development (AMED).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.