
Abstract
Cutaneous stem cells (CSCs) orchestrate the homeostasis and regeneration of mammalian skin. Epithelial CSCs have been isolated and characterized from the skin and hold great potential for tissue engineering and clinical applications. The actin cytoskeleton is known to regulate cell adhesion and motility through its intricate participation in signal transduction and structural modifications. The dynamics of actin cytoskeleton can directly influence CSCs behaviors including tissue morphogenesis, homeostasis, niche maintenance, activation, and wound repair. Various regulators of the actin cytoskeleton including kinases, actin-remodeling proteins, paracrine signals, and micro-RNAs collaborate and contribute to epithelial CSC proliferation, adhesion, and differentiation. This review brings together the latest mechanistic insights into how the actin cytoskeleton participates in the regulation of epithelial CSCs during development, homeostasis, and wound repair.
Introduction
C
Cutaneous homeostasis and wound healing depend on stem cells whose fate and motility are directly influenced by the actin cytoskeleton [8,10]. Regardless of the origin of the CSCs, the actin cytoskeleton is organized into defined structures that determine the polarity, morphology, vesicle trafficking, cytokinesis, proliferation, and motility of cells via interactions with cytoskeletal proteins and the extracellular matrix (ECM) [10 –12]. As an integral component of cellular structures, the actin cytoskeleton of stem cells is under a continual process of remodeling [13]. Assembly and severing of the actin cytoskeleton occurs in response to various niche requirements to generate the mechanical force required for cell motility, cellular contraction, and adhesion [12]. Both the dynamics and assembly of actin cytoskeletal filaments contribute to the transduction of signals from the ECM to the nucleus, which drive changes in a 3-dimensional cell structure [8,14].
The mammalian actin cytoskeleton consists of microfilaments and stress fibers (SFs) [12,13]. The precise organization of microfilaments is governed by actin binding/remodeling proteins, which modify the ratio and localization of polymerized actin filaments (F-actin) and nonpolymerized globular actin polymers (
Identification of Epithelial CSCs
Epidermal homeostasis is maintained by epidermal proliferative unit, which are central stem cells surrounded by transit-amplifying progenitor cells that give rise to the upper spinous layer; emerging evidences, however, suggest that self-renewal and proliferative progenitor-like activities are fulfilled by the interfollicular stem cells (IFSCs) simultaneously [17]. First identified as basal keratinocyte progenitor cells, IFSCs are defined as quiescent and small blast-like cells with high clonogenic capacity that maintain the stratified layers of the epidermis, these cells are also characterized by their positive keratin 14 (K14) and high alpha 6 integrin (ITGA6) expressions [1,18,19]. Recent studies have identified distinct groups of IFSC that can reestablish the epidermis autonomously, including the toll-like receptor 7 IFSCs and axis inhibition protein 2 IFSCs, further supporting the latter epidermal homeostasis theory; however, whether there is any overlapping characters between these two IFSC populations remains to be further explored [2,20].
The HF is the primary niche of CSCs, and they mostly contribute to the maintenance of HF homeostasis, following a pattern of active growth (anagen), recession (catagen), and rest (telogen) [3]. The HFSCs can be further categorized into various lineages depending on their locations in the HF and specific surface markers have been identified to trace the pattern of these HFSCs [4] (Fig. 1). As an appendage to the HF, the sebaceous gland expresses a unique marker, B lymphocyte-induced maturation protein 1 [5]. Found at the top of the HF, the infundibulum region of the HF harbors cells that express stem cell antigen 1 [6]. Continuing down the infundibulum is the junctional zone, where HFSCs express leucine-rich repeats and immunoglobulin-like domains 1 (LRIG1) and placenta-expressed transcript 1. LRIG1-positive HFSC are capable of differentiating into all skin epithelial cells after transplantation [7,21]. Below the junctional zone is the isthmus region, where a distinctive group of HFSC express the leucine-rich repeat-containing
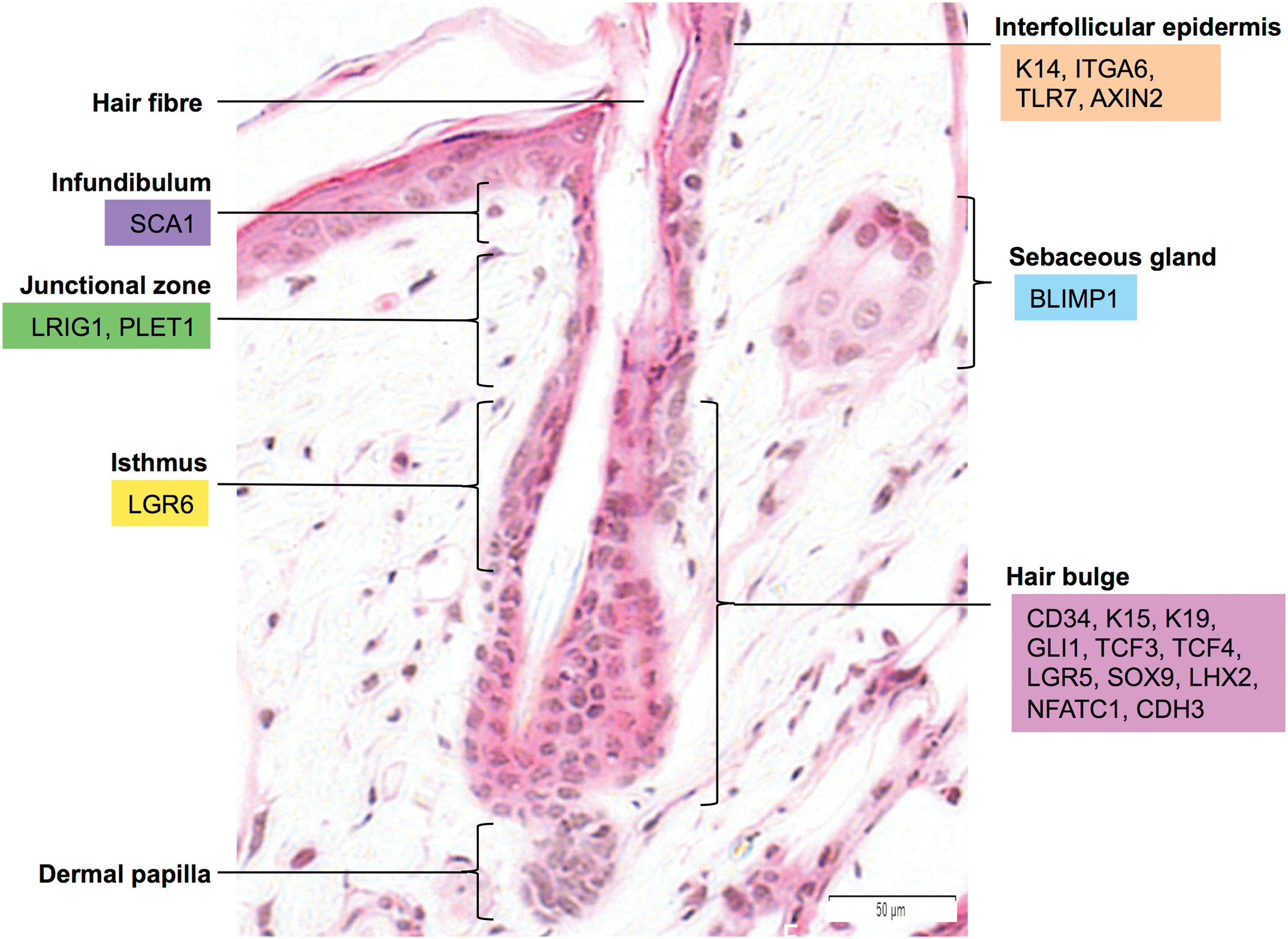
Identification of epithelial cutaneous stem cells (CSCs) with different markers using a hemotoxilin and eosin-stained mouse hair follicle profile. Interfollicular epidermal stem cells are identified using K14, ITGA6, TLR7, and AXIN2 markers, sebaceous gland stem cells are identified using BLIMP1 mark, hair follicle stem cells at the infundibulum are identified using SCA1 marker, stem cells at the junctional zone are identified using LRIG1 and PLET1, stem cells at the isthmus are identified using LGR6 and the stem cells at the hair bulge are identified using CD34, K15, K19, GLI1, TCF3, TCF4, LGR5, SOX9, LHX2, NFATC1, and CDH3. Color images available online at
Dynamics of Actin Cytoskeletal During CSCs Development
Morphogenesis
During embryonic development, the blastocyst forms an endoderm, a mesoderm and an ectoderm, with the majority of cutaneous cells being derived from the mesoderm and ectoderm [14]. Epithelial CSCs differentiate via the epidermal lineage from the surface ectoderm [28]. The actin cytoskeleton plays an essential role in coordinating cellular movements during epithelial morphogenesis. The Wnt/β-catenin-independent pathway is also known to be instrumental in providing embryonic development with instructive signals within a confined part of tissue [29]. Loss of receptor tyrosine kinase-like orphan receptor 2, an alternative tyrosine kinase receptor in Wnt/β-catenin-independent pathway, triggers an increase in the levels of the actin-remodeling kinase Ras homology family member A (RhoA) and enhances the phosphorylation of its downstream signaling partner ezrin/radixin/moesin, which belongs to a family of actin-binding proteins that lead to the disruption of actin cytoskeletal dynamics and subsequently increased branching and differentiation of the mammary epithelium [30]. As epithelial cells develop, their cytoskeletal properties change according to their lineage specification, a process often carried out by the assembly and construction of actomyosin cables [31]. Using embryonic murine eyelid closure as a model for epithelial development, K14 expressing epidermal cells have been shown to act upon local signals to acquire mesenchymal features [17]. Rather than forming a polarized supercellular actin cable, these cells organize and pack their actomyosin mediolaterally and vertically in multiple layers, initiating intercalational movements to tow the surrounding epidermal cells toward the closure point [31].
Homeostasis
Throughout adulthood, homeostasis of the interfollicular epidermis is maintained through proliferation and differentiation of stem cells in its basal layer, where different types of clonogenic IFSCs reside [32]. In terms of proliferation capacity, these IFSCs are categorized into holoclones with 180 doublings, meroclones otherwise called progenitors with less doublings, and paraclones or transient amplifying cells with a maximum of 15 doublings. To maintain stemness, a naturally occurring and irreversible clonal conversion process takes places upon extracellular signals to balance the proliferative capacity of different epidermal CSCs. One of these signals is epidermal growth factor [33], which induces a rapid expansion of holoclones but reduction in paraclones. The clonal conversion of different clones in response to EGF has been shown to be carried out through the reorganization of actin filaments, specifically through the EGFR/ERK/MLCK signaling pathway [34 –36].
Unlike mesenchymal stem cells (MSCs), the differentiation of epithelial CSCs is independent of extracellular composition or density. Instead, changes in the actin cytoskeletal organization triggers shape-induced differentiation by modulating the transcriptional activity of serum response factor (SRF) targeted genes [37]. Monomeric
Niche maintenance
To achieve a dynamic balance between lineage commitment and self-renewal, architectural niche organization needs to be intricately controlled in epithelial CSCs, and this has always been hypothesized to be solely influenced by external signaling pathways [10]. Latest evidence suggests that cytoarchitecture also has an impact on niche maintenance of epidermal CSCs [38]. One of the major decisions made by stem cells of the niche during cell proliferation is the apicobasal polarity as this determines the outcomes of either symmetrical or asymmetrical division, the process whereby a new quiescent progenitor and a terminally committed daughter cell are generated [39]. Using HFSCs as a model system, the actin cytoskeleton has been shown to influence adult CSC polarity and niche choice with the help of internal signaling factors such as Lim-homeodomain TCF (LHX2) [20]. Reduction of LHX2 expression leads to the depletion of F-actin and its regulators, triggering mitotic F-actin spindles to adapt a perpendicular polarity. This process forces the HFSCs to exit the asymmetric division that requires parallel polarity, and hence leads to the loss of HFSC quiescence [38].
Activation of epidermal CSCs
In response to extracellular signals including growth factors and ECM substrates during wound healing, epithelial CSCs go through epithelial-mesenchymal transition to transform into migratory cells [40 –42]. The actin cytoskeletal properties of the stem cells need to be quickly remodeled to generate the necessary mechanical forces required for changes in cell motility and adhesion [10]. Upon activation, epithelial CSCs need to first journey out of their quiescent niches before migration and adhesion can take place [43]. LHX2 expressed in HFSCs upregulates downstream reepithelialization promoting effectors including sex-determining region Y box 9 (SOX 9) and TCF4 while downregulating quiescence factor leucine-rich repeat containing G protein-coupled receptor 5 (LRG5) [44]. This process transiently switches the fate of HFSCs from hair-regeneration to epidermal amplification, and triggers the accelerated actin polymerization and differential remodeling necessary for migration [38,45].
Reepithelialization
To fully restore tissue integrity and function after injury, cutaneous wound healing undergoes a series of complex and dynamic stages including homeostasis, inflammation, granulation, and remodeling [46]. Various cell types collaborate and participate in these overlapping stages of wound healing process to synergize the healing outcome [47]. While endothelial stem cells such as MSCs initiate inflammation and coordinate angiogenesis, epidermal cells including mature keratinocytes, IFE progenitor cells and HFSCs contribute to the reepithelialization process during the granulation phase at different rates and proportions [17,48] (Fig. 2). Upon cytokine signals produced by the surrounding wound matrix and fibroblasts, keratinocytes at the edge of the wound and HFSCs in the adjacent hair bulges are activated to proliferate and produce daughter cells that migrate up the follicular infundibulum toward the center of an acute cutaneous wound, leading to the formation of a neoepidermis [3]. Despite the difference in cell heterogeneity, wound reepithelialization requires the deposition of a provisional basement membrane (BM) and actin cytoskeletal reorganization [49]. To migrate across the wound matrix and form new and stable adhesion sites, the cells first assemble actin at the front of the cell driving the extension of membrane protrusions called lamellipodia and filopodia, which consists of short or long parallel actin filament bundles respectively [50]. At the leading edge of the cell, adhesions are formed with the ECM, anchoring the protrusions to move the cell body. The combination of actomyosin contraction and the disassembly of the adhesion structures at the rear of the cell allows the cell body to move forward [51]. Upon the establishment of the neoepidermis, the majority of newly migrated CSCs switch from an activated state back to their quiescent state to retain their self-renewal ability, while their progenitors continue to support the stratification of epidermis [46,47].
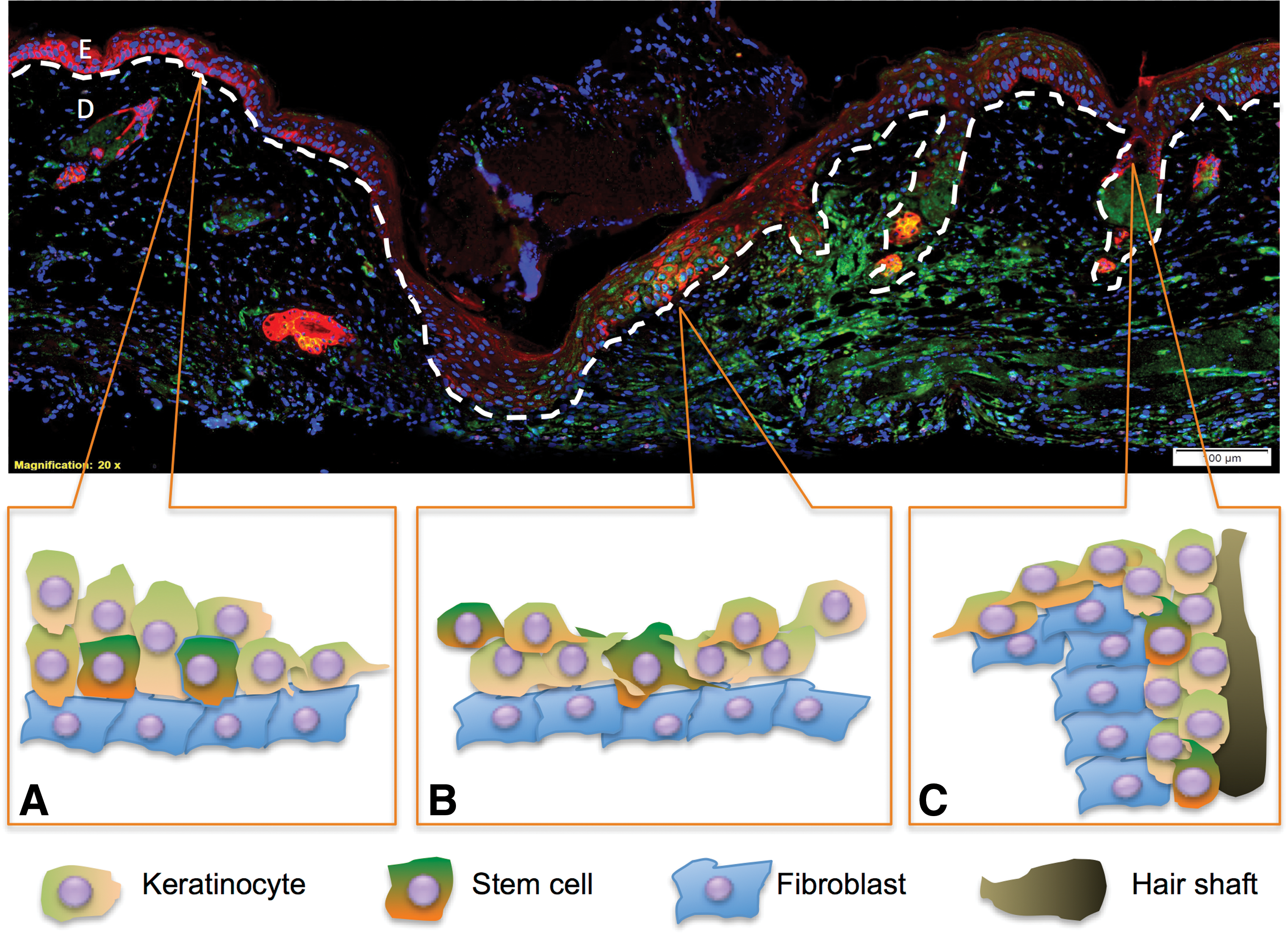
Participation of epithelial CSCs from different locations of the skin during the granulation phase of wound repair illustrated using a day 3 incisional wound marked with CD34 (green), K15 (red), and DAPI (blue).
Regulators of Actin Cytoskeleton in CSCs
Key kinases
The regulation of actin cytoskeleton organization, migration, and adhesion in CSCs relies on the cooperation of Rho GTPases including Rho, Rac1, and Cdc42 along with intracellular protein kinases, which play critical roles in the assembly and disassembly of adhesion junctions and focal adhesions [52,53]. While RhoA is instrumental in the formation of SFs and focal adhesion formation, Rac1 and Cdc42 are required for the formation of lamellipodia and filopodia [12]. Interestingly, keratinocyte-restricted RhoA knockout mice showed no difference in the physiological keratinocyte arrangement including desmosomes, adherens junctions, hemidesmosomes, and BM during normal skin development [54]. This observation led to the discovery that RhoB and RhoC can act as compensational regulators of Rho-kinases, however, not during Rho-associated protein kinase (ROCK)-dependent phosphorylation of myosin light chains (MLC) and cofilin [54]. The same study also showed that RhoA regulates ROCK and MLC independently of Rac1 during cell spreading, however, the impaired directional migration ability of RhoA knockout keratinocytes did not alter wound healing outcomes in vivo [55].
Upon Cdc42 activation through the Rac1/Cdc42/WASP/WAVE signal transduction pathway, the recruited actin-related protein (Arp2/3) nucleates the new filaments from the sides of pre-existing filaments and induces the formation of a branched filament network at the leading edge of the cell [56]. Recent studies have identified Cdc42 as a core signaling partner for both EGF and connective tissue growth factor (CCN2) induced cell migration during wound healing [51,52]. EGF has been shown to induce Cdc42 transducer Toca-1 localization to filopodia and lamellipodia, where it colocalizes with F-actin and the Arp2/3 complex, leading to upregulated migration and invasiveness in epidermoid carcinoma cells [57]. Another study used primary human keratinocytes to show that CCN2, an angiogenesis promoter well known for its ability to stimulate ECM production, was also capable of inducing keratinocyte migration [52]. Upon CCN2 induction, the promotion of F-actin disassembly was found to be triggered indirectly by an increase in Cdc42 and the production of RhoA antagonist p190RhoGAP [52]. Additionally, the effect of CCN2 on cell spreading was found to be dependent on focal adhesion kinase (FAK) but independent of Rac1 [58].
Integrin-mediated cell-matrix adhesions, termed focal complexes, develop underneath lamellipodia and are driven by actin polymerization [55]. A proportion of the stable focal complexes develop into elongated focal adhesions, which provide contractile force required for the effective translocation of a cell body during cellular migration [59]. This is highly dependent on signaling between integrin receptor and a number of actin-associated proteins recruited to the adhesion sites including paxillin, talin, vinculin, FAK, proto-oncogene tyrosine-protein (Src) kinase, integrin-linked kinase (ILK), and phosphatidylinositol 3-kinase (PI3K) [60]. As a central regulator of the integrin-actin linkage, ILK is required to maintain the epithelial CSC niche. Mice lacking ILK in the epidermis display an accumulation of undifferentiated epidermal progenitor cells, while the ILK-knockout keratinocytes exhibit multiple abnormalities in actin cytoskeletal organization and impaired activation of Rac1, resulting in failure to attach and spread efficiently [61,62]. A recent study showed that deletion of ILK altered remodeling of the actin-BM linkage within the HFSC niche and increased Wnt and TGF-β signaling, resulting in aberrant bulge HFSC activation and their subsequent diminution as a result of continuous differentiation [42].
As discussed above, PI3K is another important regulator involved in the actin cytoskeletal organization and motility, through which the proliferation and differentiation of epidermal cells are affected [63]. PI3K activation can be triggered by ligand binding to the integrin receptor, receptor tyrosine kinase, or EGF receptor [34,64,65]. Activated PI3K leads to the downstream activation of protein kinase B [55] and Rac1 [34]. PI3K/Rac1 signaling, but not Akt signaling, has been shown to regulate the organization of actin filaments and dynamics of epidermal stem cell colonies during EGF-induced activation [34]. Interestingly, Akt was also found to inhibit Rac1 independently of PI3K, acting as another upstream regulator of actin organization through the Rac1 pathway [34].
Actin modifiers
Following kinase signaling at new adhesion sites, reorganization of actin filaments is further regulated by different structural modifiers, which establish the appropriate spatial geometry of filaments at the protruding regions of the cell [50]. Some of these actin-binding proteins have been implicated in the regulation of epidermal stem cells. Actin cross-linking factor-7 (ACF7), a microtubule and F-actin linking protein, is particularly abundant in HFSCs [66]. As a direct effector of glycogen synthase kinase 3β (GSK3β), a protein kinase required for cell migration and polarity downstream of PI3K signaling pathway, ACF7 is required to polarize microtubules along actin SFs at focal adhesions [67,68]. Loss of ACF7 results in disrupted microtubule structures in HFSCs, leading to changes in cell polarity and perturbed upward migration of HFSCs in response to skin wounding, and this process is also dynamically governed by GSK3β phosphorylation of ACF7 [68]. Another regulator found to be highly expressed within the HF during the active growth phase of hair cycling, Flightless I (Flii), is an actin-remodeling protein that belongs to the gelsolin family of actin-capping/severing proteins, and it has been shown to be essential during HFSC development [69]. Changes in Flii levels lead to altered actin polymerization, cell migration, and adhesion in keratinocytes and subsequent altered wound healing outcomes [70
–73]. Reduced levels of Flii expression lead to decreased expression of syndecan-1 (CD138) and fibroblast growth factor receptor 2 expression in hair bugle cells, resulting in a delay in HF development [69]. Additionally, another actin-severing protein, cofilin, has been shown to be a crucial effector during SRF-mediated actin disassembly for efficient cell spreading. The phosphorylation of cofilin, activated by SRF, triggers an accumulation of F-actin and a reduction in
Paracrine signaling
A variety of paracrine signals including growth factors and cytokines participate in the regulation of actin organization in epithelial CSCs. Noticeably, ligand activation of the EGF receptor [33] signals both stem cell-specific partners and actin dynamic modulators [36]. Upon activation through the EGFR/ErbB/c-CBl pathway, levels of leucine-rich repeats immunoglobulin-like domain protein (Lrig1), a stem cell quiescence regulator commonly expressed by junctional-zone CSCs, are elevated to counteract the proliferation-stimulatory effect of EGF [74]. Meanwhile, EGFR/Rac1 activation triggers actin reorganization and clonal growth of epithelial CSCs [34]. During HF development, bone morphogenic protein (BMP) controls the hair cycle transition by maintaining HFSCs at a quiescent state [75]. Low levels of BMP receptor 1A is observed in the HFs directly adjacent to the wound [76]. Upon the activation of BMP/SMAD1, the expression of genes including actin-binding myosin 5a (Myo5a), actin-binding LIM protein 2 (Ablim2) and tubulin β6 (Tubb6) responsible for actin-remodeling and motility are significantly reduced, leading to slowed migration of K14-positive primary keratinocytes in vitro and delayed wound healing in mice skin [54]. Additionally, small molecules exerting a paracrine effect, for example cyclic adenosine monophosphate (cAMP), have also been identified as mediators during Rac1/Cdc42 activation. cAMP induction increases Arp3 expression yet decreases cofilin phosphorylation levels, leading to actin cytoskeleton remodeling and improved migration in stem cells during cutaneous wound healing [77].
Micro-RNA
MicroRNAs (miRNAs) are a novel class of endogenous single-stranded RNAs, 19–24 nucleotides in length, which regulate development (stem cell proliferation, cardiac and skeletal muscle development, neurogenesis, and hematopoiesis), cell differentiation, proliferation, and apoptosis and are involved in several other processes including insulin secretion, cholesterol metabolism, immune response, and heart disease [78]. MiRNAs control gene expression on a variety of levels and through several mechanisms including altering chromatin structure, epigenetic memory, transcription, RNA splicing, editing, translation, and turnover[79]. CSCs preferentially express specific miRNAs to direct self-renewal and lineage differentiation, some of these including miR-203, miR-24, miR-574, miR-720, and miR-34a promotes epidermal superbasal differentiation [80], whereas others including miR-205 and miR-125b conversely promotes self-renewal and repress HF differentiation [81 –83]. While majority of these miRNAs are working through transcription processes such as p63 pathway to regulate epithelial stemness, emerging evidences have also revealed miRNAs as important regulators of actin cytoskeleton dynamics in epidermal CSC migration [80,83,84]. Expressed in the basal epidermis and previously shown to be an important regulator of epithelial to mesenchymal transition in cancer studies, miR-205 has been shown to promote the migration of primary keratinocytes by targeting multiple actin remodeling signaling pathways [85]. miR-205 overexpression increases levels of F-actin while the downregulation of miR-205 enhances cell adhesion and focal contacts, leading to improved cell survival and keratinocyte migration. These changes are shown to be triggered by miR-205 post-transcriptional repression of SHIP2, a crucial ubiquitous lipid phosphatase in the PIP3-Akt pathway; and also through Rho-ROCK activation, which inactivates the actin-severing protein cofilin via phosporylation. The same study also found that miR-184, an antagonist of miR-205, was able to reverse the repression on SHIP2 and coordinately regulate actin dynamics via Akt survival signaling in keratinocytes [85]. Found to be expressed during terminal differentiation in various tissues, miR-24 is expressed concomitantly during epidermal stratification and differentiation, and it has been shown to regulate actin dynamics through the repression of three actin cytoskeletal modulators: PAK4, Tsk5, and ArhGAP19 [84]. miR-24 overexpression diminishes the formation of actin SFs and formation of mature adhesion junctions, while apical-basal polarization of actin filaments is promoted [84]. Not only does this lead to decreased proliferative and migratory ability of basal keratinocytes, but it switches the cells to a differentiating state [84]. Table 1 summarizes epithelial CSC dynamics through the key molecular pathways in the regulation of actin cytoskeleton.
ACF7, actin cross-linking factor-7; Arp2/3, actin-related protein; BMP, bone morphogenic protein; cAMP, cyclic adenosine monophosphate; CSC, cutaneous stem cells; ERM, ezrin/radixin/moesin; FAK, focal adhesion kinase; FGFR2, fibroblast growth factor receptor 2;
Stem Cell Therapies
The use of stem cells from skin origin has been greatly investigated particularly in wound healing, burns, and genetic skin diseases [86]. Transplantation of cultured-keratinocytes-autografts (CEA) that contain CSCs in patients with large burns provides an early and efficient wound coverage while accelerating reepithelialization, yet the low percentage of the CSCs in the CEA limits the long-term durability of this therapy [87]. The ineffectiveness of available skin substitutes has driven medical professionals to consider stem cell therapies, as the multipotency and self-renewal ability of stem cells could provide better treatments and potentially bypass existing obstacles [86]. Various stem cell therapies including MSCs, hematopoietic stem cells, endothelial progenitor cells, multipotent progenitor cells (MPCs), and human induced pluripotent stem cells (iPSC) have been trialed clinically for cutaneous wound repair [88]. iPSC derivation and directed differentiation have been well established in recent years and their proliferative capacity provides a theoretically unlimited cell source for regenerating skin tissue [86,89]. The use of iPSC developed from skin fibroblasts of recessive dystrophic epidermolysis bullosa (RDEB) patients has been shown to successfully restore skin integrity in RDEB patients [89 –91]. However, the risk of genetic reversion of the programmed iPSC back to the diseased phenotype due to instability in DNA repair remains uncertain [90,91].
Stem cells directly participate in the granulation process of wounding healing by migrating to the wound site and differentiating into mature tissues; they also exert paracrine effects through the production of growth factors [92]. This phenomenon is observed in both MSCs and non-MSCs including skin papilloma stem cells and melanoma stem cells [92]. Noticeably, cell therapy using progenitor cells such as MPCs has a safety advantage over others, since the limited lifespan of MPCs lowers the potential risk for malignancy development [88]. While most MPCs are sourced from either adipose tissue or the bone marrow, recent success in isolation and culturing of skin-derived progenitor cells (SKPs) has enabled the skin to become a novel and abundant reservoir for stem cell isolation [93]. SKPs were first derived as multipotent stem cells showing neural and mesodermal lineage differentiation potential [94]. Later on, SKPs were successfully derived from human and other animals [95,96]. SKPs are different from epithelial stem cells, as they form spheres structure rather than aggregated colonies in vitro. SKPs are considered to share the features of embryonic neural crest stem cells in the peripheral system [94 –96]. Skin-derived MSCs have also been shown to restore the function of damaged ovaries in chemotherapy-treated females [97], underpinning the wound-healing ability of stem cells [98]. Epithelial stem cells from the skin have also been successfully isolated and characterized, including HFSCs marked by cytokeratin 15 and CD34 antigen and IFSCs marked by integrin α6 and LRIG1 [7,99]. It may become possible to apply these epithelial stem cells in different therapies in the near future.
Stem cell-based tissue engineering requires precise control of cell differentiation during tissue culturing. As an important mechanosensor, the actin cytoskeleton needs to be maintained accordingly to achieve a self-renewing and undifferentiated pool of epithelial CSCs [99]. A stem cell culture related to the actin cytoskeleton remodeling including feeder cells, growth factor concentration, serum percentage, and calcium levels requires consistent monitoring to enhance proliferation and suppress differentiation [100,101]. An alternative to using feeder cells, which mimics the integrin-rich BM while providing growth factors is the use of surfaces coated with one or a combination of biological materials including laminin-323, collagen-1, and fibronectin. This technique has also been used for the initial establishment of adherent-type stem cell cultures [102,103]. Recent advances in surface technology have further provided skin stem cell culturing with novel strategies, including the use of collagen and poly
Conclusion
The ability of the cutaneous epithelium to sustain homeostasis and repair wounds relies on its reservoir of various multipotent progenitors and stem cells. Although it has been known for decades that the actin cytoskeleton plays a key role in cell motility during development and regeneration, recent innovations in the analysis and isolation of epithelial stem cells have elucidated pathways by which the actin cytoskeleton regulates epithelial CSCs behavior. As skin is an unconventional biological system to explore stem cell dynamics; limited data have been published using epithelial CSCs as a direct model system. However, the knowledge gained from other stem cell and epidermal studies can predict the behavior of epithelial CSCs. Different types of regulatory mechanisms, ranging from gene expression to protein interactions, participate in the fate decision of stem cells, and many of these mechanisms rely on the actin cytoskeleton as an intermediate partner or as the final downstream effector. Understanding actin cytoskeleton dynamics and regulation in various epithelial CSCs enables us to use this knowledge to influence tissue engineering and wound healing outcomes.
With growing clinical use of stem cell therapy, future studies need to provide the medical practice with up-to-date evidence of how they behave and what factors they influence during homeostasis and wound repair. As a major governor of cell motility, the dynamics of the actin cytoskeleton in response to environmental factors can be used as an indicator of cell behavior during wound healing. By combining expertise in actin cytoskeletal regulation, somatic stem cell biology and tissue culturing, it may be possible to enhance and expand the clinical applications of bioengineered CSC therapy in the future.
Footnotes
Author Disclosure Statement
No competing financial interests exist.