
Abstract
Individuals with neurodegenerative disorders or brain injury have few treatment options and it has been proposed that endogenous adult neural stem cells can be harnessed to repopulate dysfunctional nonneurogenic regions of the brain. We have accomplished this through the development of rationally designed hydrogel implants that recruit endogenous cells from the adult subventricular zone to create new relatively long tracts of neuroblasts. These implants are biocompatible and biodegradable cylindrical hydrogels consisting of fibrin and immobilized neurotrophic factors. When implanted into rat brain such that the cylinder intersected the migratory path of endogenous neural progenitors (the rostral migratory stream) and led into the nonneurogenic striatum, we observed a robust neurogenic response in the form of migrating neuroblasts with long (>100 μm) complex neurites. The location of these new neural cells in the striatum was directly coincident with the original track of the fibrin implant, which itself had completely degraded, and covered a significant area and distance (>2.5 mm). We also observed a significant number of neuroblasts in the striatal region between the implant track and the lateral ventricle. When these fibrin cylinders were implanted into hemiparkinson rats, correction of parkinsonian behavior was observed. There were no obvious behavioral, inflammatory or tumorigenic sequelae as a consequence of the implants. In conclusion, we have successfully engineered neural tissue in vivo, using neurogenic biomaterials cast into a unique cylindrical architecture. These results represent a novel approach to efficiently induce neurogenesis in a controlled and targeted manner, which may lead toward a new therapeutic modality for neurological disorders.
Introduction
I
This innovation is derived from our understanding of adult neurogenesis in the mammalian brain, which consists of a coordinated series of overlapping steps, including proliferation, differentiation, migration, neuritogenesis, axon guidance, and integration [10]. Induced adult neurogenesis has been demonstrated to be a natural response to various activities such as exercise [11] or neurological disorders such as stroke [12 –15] or traumatic brain injury [15 –17], and this has led to the belief that endogenous neural stem cells can be harnessed to repopulate dysfunctional or lesioned nonneurogenic regions of the brain [18,19]. Studies using cultured neural stem cells and genetically engineered animals have identified numerous neurotrophins and extracellular matrix (ECM) components that appear to mediate the maturation of endogenous neural progenitor cells (NPCs) into neurons [20 –22]. However, there are few reports that integrate this knowledge into an application that can be used to engineer an effective means to recruit these cells to regenerate neural tissue, especially using an in vivo approach. The full potential of therapeutic endogenous neural stem cells requires efficient cell recruitment, targeting, neural subtype-specific differentiation, and long-term cell survival. The results presented in this study represent an important contribution toward achieving these goals.
A renewable source of endogenous neural stem cells is the adult mammalian subventricular zone (SVZ). Much of what we currently know about neurogenesis has come through studies of the SVZ/rostral migratory stream (RMS)/olfactory bulb (OB) pathway [23,24]. SVZ-derived neural progenitor cells (SVZ-NPCs) are the likely source of nascent neurons, which arise in the typically nonneurogenic striatum following exercise or injury [11 –17]. This supports the notion that SVZ-NPCs are naturally rerouted to typically nonneurogenic regions in response to an insult to the brain. While a vital observation, the number of SVZ-derived cells naturally recruited to lesioned areas is too low to expect significant therapeutic efficacy in the case of most neurodegenerative diseases or brain injury.
Recent efforts to deliberately recruit endogenous NPCs into nonneurogenic areas of the brain have met with some success. This approach was in the form of hyaluronic acid engineered to contain neurotrophic factors, made as an injectable low viscosity polymer [25], which increased neurogenesis in rat cortex and striatum after injection into these regions [26]. These authors suggested that the source of new neural cells may be endogenous quiescent NPCs rather than a neurogenic niche such as the SVZ.
The goal of this report is to present research describing a means to reroute and retask endogenous SVZ-NPCs from the RMS to populate nonneurogenic brain regions with new neural cells. Our approach was to develop an implantable, biocompatible, and biodegradable fibrin matrix, which was readily modified to contain judiciously chosen immobilized factors such as neurotrophins, ECM molecules, and aprotinin. Our goal, upon implantation into brain, was to provide an environment that would create a new migratory path from the renewable neurogenic niche of the SVZ/RMS to a new and distant target region. We chose to include both neurogenic [nerve growth factor (NGF)] and angiogenic [vascular endothelial growth factor (VEGF)] factors in the implantable microenvironment since both developmental processes are inextricably linked to the genesis of neural paths [27 –30].
We believe that fibrin is an ideal biomaterial for these implantable matrices; it is readily chemically modified to covalently incorporate bioactive peptides and proteins [31]; it is biochemically similar to extracellular proteins found in the brain [32,33]; fibrin is biologically compatible and is degraded by well-known and inhibitable proteolytic processes [34,35]; and, it is used clinically in the form of fibrin sealant and has low immunogenicity [36,37]. Furthermore, several groups have successfully used fibrin hydrogel injected into animals to address spinal cord and peripheral nerve injury [31,38 –43]. Finally, it seemed logical to create this implantable biomaterial in the form of a cylinder since the RMS is, in fact, cylindrical.
Materials and Methods
Preparation of implantable cylindrical scaffolds
Fibrinogen (Sigma-Aldrich, St. Louis, MO) was dissolved in phosphate-buffered saline (PBS) at a final concentration of 64 μM, degassed, and stored in aliquots at −80°C. An 8 μL aliquot of fibrinogen was placed on a square of Parafilm® to which was added aprotinin (15 μM final concentration; Sigma-Aldrich), laminin I (100 μg/mL final concentration; Trevigen, Gaithersburg, MD), carrier-free VEGF (10 μg/mL final concentration; R&D Systems, Minneapolis, MN), and carrier-free NGF (10 μg/mL final concentration; R&D Systems). The volume of the sample was brought to 18 μL with PBS. The homobifunctional crosslinker disuccinimidyl suberate (DSS; Thermo Scientific Pierce, Rockford, IL) was dissolved in dry dimethyl sulfoxide (dried over 4 Å, 40 mesh molecular sieves) and 1 μL (500 μM final) was added to the sample with thorough mixing. DSS is a chemical crosslinker that is directed toward amino groups, thus facilitating the covalent incorporation of the peptides and proteins used in this study (i.e., VEGF, NGF, laminins, and aprotinin) into the fibrin hydrogel [44 –48]. The crosslinking reaction was incubated for 10 min at ambient temperature in a humidified chamber. Thrombin (1 μL of 0.1 U/μL; Sigma-Aldrich) was added and the sample quickly mixed and injected through a Gilson P20 Pipetman® into a quartz tube (0.7 mm i.d., 3.5 cm long; Fiber Optic Center, Inc., New Bedford, MA) such that a 7 mm long cylinder was created. The dimensions of the final cylinder were 0.7 mm dia. × 6 mm long following the polymerization and concomitant shrinkage of the fibrin. Cylinders were made within 4 h of implant surgery and stored in a humidified chamber until used.
Implantation of cylinders into rat brain
Female Sprague-Dawley rats (175 g; 7 weeks old) were purchased from Hilltop Lab Animals, Inc. (Scottdale, PA). Animals were acclimated and handled every 1–2 days for 2–3 weeks before experimentation. Cages were provided environmental enrichment in the form of PVC pipe fittings. All animal experiments were carried out in accordance with the National Institutes of Health Guide for the Care and Use of Laboratory animals and performed under an approved protocol from Marshall University's IACUC. The fibrin cylinders were implanted along a predetermined path in the right hemisphere of each animal (Fig. 1). A short (ca. 0.5 mm) length of polyvinylidene difluoride (PVDF) fluorocarbon line (0.56 mm diameter leader material; Berkley, Columbia, SC) was inserted into the quartz tube containing the fibrin cylinder and served as a plunger head and a 20 cm length of stainless steel wire (0.5 mm dia.; AmazonSupply) was used as a plunger (Fig. 1). Animals were anesthetized using isoflurane (5% induction, 2% maintenance) with an oxygen flow of 0.8 L/min.

Diagram of coronal section of rat brain showing location of hydrogel implant. The relative location of the cylinder is at an angle 17° right of midline, implanted through the skull, relative to bregma, and at 0.13 cm anterior and 0.30 cm lateral right of midline, with the distal end of the implant 0.73 cm dorsoventral to the skull. The cylinder passes through the cortex, corpus callosum, the RMS, and into the striatum. The fibrin-based biomaterial degrades to completion after ∼4 weeks. Coronal diagram reprinted with permission, originally published in “The Rat Brain in Stereotaxic Coordinates,” George Paxinos and Charles Watson, figure 13, Copyright Elsevier (1998). Inset: Implant assembly. A solution of fibrinogen/thrombin/immobilized factors was injected into quartz tubes (0.7 mm i.d. × 35 mm length) such that 6 mm long fibrin cylinders were produced. A plunger head made of PVDF was inserted into the cylinder and pushed to the top of the fibrin cylinder using a 20 cm long stainless steel wire (0.5 mm dia.) as a plunger. The plunger was removed and the quartz tube/fibrin cylinder/plunger head assembly was affixed to the right arm of a stereotaxic frame and the plunger wire attached to the left arm. The fibrin cylinders were surgically implanted into rat brain as described in Materials and Methods section. PVDF, polyvinylidene difluoride; RMS, rostral migratory stream.
Animals were positioned into a Kopf stereotaxic frame that was outfitted with two manipulators, one on each arm of the Kopf “U” frame, and both were angled 17° to the right of midline. The fibrin cylinder-containing quartz tube was affixed to the right manipulator and the wire plunger was placed on the left manipulator. A small hole was drilled using a round dental bur at, relative to the bregma, 0.13 cm anterior and 0.30 cm lateral right of midline [49]. The dura was gently pricked with a 25-gauge needle and the quartz tube slowly driven 0.73 cm down the dorsoventral axis on an angle 17° off midline to the right. The plunger was lowered into the fibrin cylinder-containing tube until just touching the PVDF plunger head, which itself was just touching the fibrin cylinder. The plunger was held immobile by the left manipulator arm, while the quartz tube was slowly raised (1/4 turn/45 s) by the right manipulator. This maneuver resulted in the 6 mm cylindrical fibrin matrix being placed (not extruded) in the brain along a predetermined path (Fig. 1) as the quartz tube was removed from the brain with the plunger holding the hydrogel cylinder in place.
The final position of the 6 mm long implant started at the surface of the brain and passed through the cortex, corpus callosum, RMS, and striatum. After complete extraction of the quartz tube, the skull hole was plugged with bone wax and the scalp closed with absorbable sutures. Animals were injected with dexamethasone [0.4 mg/kg subcutaneously (s.c.) in 0.9% sterile saline] once at the time of anesthesia induction and then each day for 4 days postoperatively. For pain management, animals were injected with buprenorphine (0.15 mg/kg s.c.) immediately after surgery and then daily for 2 additional days. Ten animals were used in this series; six received fibrin cylinders containing VEGF, NGF, laminin I, and aprotinin; two were implanted with fibrin cylinders containing only immobilized aprotinin and laminins; and two underwent sham surgeries (empty quartz tubes driven to the same coordinates).
6-hydroxydopamine lesions
Twelve rats were prepared for surgery as described above. The hemiparkinson model was generated by 6-hydroxydopamine (6-OHDA) injection into the right medial forebrain bundle (MFB). Desipramine was injected (i.p., 25 mg/kg in sterile H2O) just after anesthesia induction. The coordinates for MFB injection were, relative to bregma, −0.44 cm anterior; 0.12 cm right of midline; and −0.84 cm dorsoventral, and 3 μL of 6-OHDA (5 mg/mL in freshly made, degassed 0.9% NaCl/0.02% ascorbic acid) was injected at a rate on 0.5 μL/min. Following injection, the cannula was left in place for 5 min and then slowly removed (raised 1/4th of a turn every 45 s) and the site closed as described above.
Two weeks postoperative, animals were injected s.c. with 5 mg/kg
Immunohistochemistry
Animals were sacrificed by bilateral pneumothoracotomy, fixed by transcardial perfusion with 0.9% NaCl containing 2 U/mL heparin followed by 4% freshly prepared paraformaldehyde (PFA), and brains recovered. Following overnight postfixing in 4% PFA, brains were equilibrated in 30% sucrose until no longer buoyant and stored at 4°C until processed into 20 μm sections using a Leica Model 1950 cryostat. Samples were incubated for 15 min in 50 mM NH4Cl, permeabilized in 0.1% Triton X-100, blocked overnight at 4°C in 5% bovine serum albumin (BSA; Jackson Immunoresearch Laboratories, West Grove, PA) and 5% normal goat serum, and subsequently incubated with primary antibodies (1:200 in 0.1% BSA-c™; Aurion (Wageningen, The Netherlands); 0.1% normal goat serum) overnight at 4°C. Sources of antibodies were as follows: Doublecortin (Dcx; a marker for immature neurons and neurogenesis), Millipore (Darmstadt, Germany); Neurofilament medium (NF160; a marker for mature neurons), Abcam (Cambridge, MA); tyrosine hydroxylase (TH; a marker for mature dopaminergic neurons), Abcam; microtubule-associated protein 2a and 2b (MAP2ab; a marker enriched in mature neuronal dendrites), Abcam; and glial fibrillary acidic protein (GFAP; a marker for reactive astrocytes), Abcam. Following extensive washing, sections were incubated overnight at 4°C with Alexa-conjugated fluorescent secondary antibodies (Life Technologies, Carlsbad, CA) diluted 1:500. Following washing, samples were mounted on Superfrost plus glass slides (Fisher Scientific, Pittsburgh, PA) and coverslipped with number 1 glass with Fluoro-Gel II-DAPI (Electron Microscopy Sciences, Hatfield, PA) as the mountant. Slides were sealed with clear nail polish and stored at 4°C until imaged/analyzed using a Zeiss Axio Observer with 10× and 20× LD Plan Neofluar air-interface objectives and AxioVision 4.6.3 software.
Results
Development of fibrin cylinders for in vivo neural engineering
Our goals were threefold; first, we engineered a biocompatible cylindrical matrix that was of sufficient mechanical stability that it could be implanted into the brain in a manner that essentially connected neurogenic to relatively distant nonneurogenic regions. Other types of matrices that have been used to stimulate neurogenesis are relatively fluid and, while straightforward to inject, not likely to generate long engineered pathways targeted to specific regions of the brain. Second, we used an aprotinin-stabilized formulation of fibrin for the implant because we posit that the ideal matrix must be somewhat stable, yet biodegradable such that it will eventually yield newly developed neural paths consisting of complex networks of neural cells and microvasculature. Our unpublished data (article in preparation) and others [50,51] have demonstrated the ability for aprotinin, a peptide serine protease inhibitor, to fine tune the in vitro and in vivo neural cell-mediated degradation rate of fibrin. This information was used in this work to generate a matrix that would be completely degraded within 4–5 weeks. Third, we created an implantable biomaterial with a cylindrical geometry that was readily modified with immobilized factors chosen to recruit endogenous NPCs from the SVZ-RMS and direct them into a typically nonneurogenic region. Since the quintessential neural migratory path in adult mammals is the RMS, and this has a three-dimensional architecture that is relatively cylindrical, we engineered fibrinogen, thrombin, and additional components as small implantable cylinders, cast in quartz tubes.
Implanted fibrin cylinders recruit neuroblasts that migrate along the implant track
Animals were sacrificed 4–5 weeks following surgery, a point at which the cylinders were completely degraded, and we determined the phenotype and anatomical distribution of migrating neuroblasts in coronal sections of the brains. This time frame was chosen to visualize Dcx+ cells utilizing a stable migratory path rather than merely responding to the inherent damage of the implant surgery, which should be resolved before the 4–5 week sacrifice time point [18]. Immunohistochemical analyses of samples from animals implanted with fibrin cylinders containing NGF, VEGF, laminin I, and aprotinin revealed significant numbers of migrating neuroblasts (Dcx+; Figs. 2 and 3). These cells exhibited complex, long, and branching neurites and were most numerous along the original location of the cylinder in the striatum. The new path facilitated by the hydrogel implant varied slightly in length between animals and was between ∼2.0 and 2.8 mm in length. Evaluation of the Dcx+ cells at higher magnification (Figs. 3 and 4) reveals the complexity of the neuroblasts' neurites, many exceed 100 μm in length and exhibit numerous branch points (Fig. 4). There was no residual mechanical damage due to the surgical procedure nor was there significant GFAP+ labeling (data not shown) at the site of the implant, indicating there was not a significant inflammatory response [52] to the matrix.
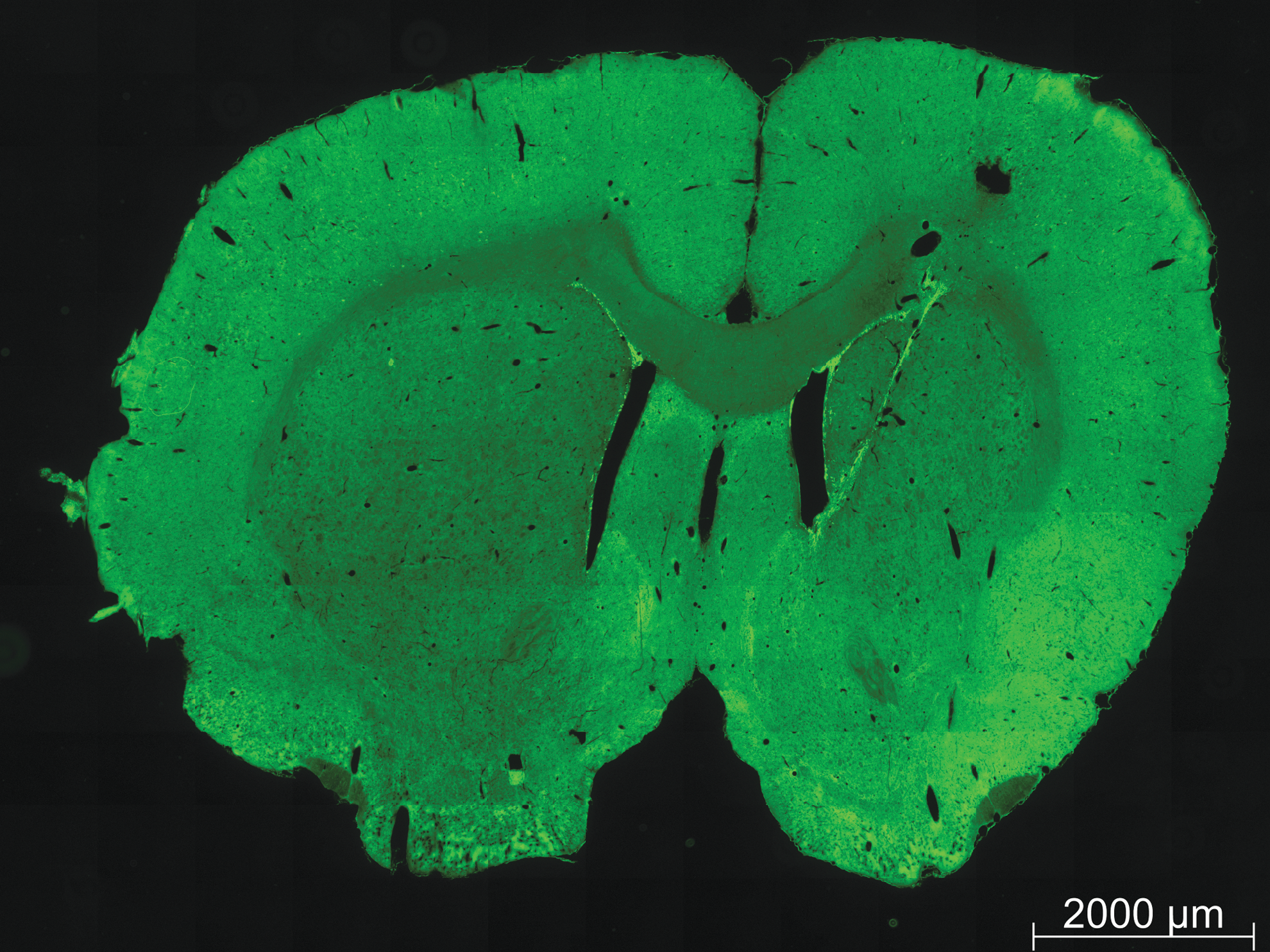
Recruitment of neural precursors by the fibrin implant. Sections were labeled for migrating neuroblasts with anti-Dcx antibodies. Shown is a whole coronal section revealing both the left (contralateral) nonimplanted and right implanted hemispheres. Dcx+ cells (green) were observed in both hemispheres immediately dorsal to the lateral ventricles (indicative of the posterior RMS), while only the implanted hemisphere exhibited significant Dcx+ cells in the striatum, along the implant path. The length of the tract of neuroblasts in this animal is ∼2,600 μm. The fibrin cylinder was itself degraded and the majority of labeling was along the track of the implant; additional labeling was observed between the wall of the lateral ventricle and the implant track (Fig. 5). Dcx, doublecortin.
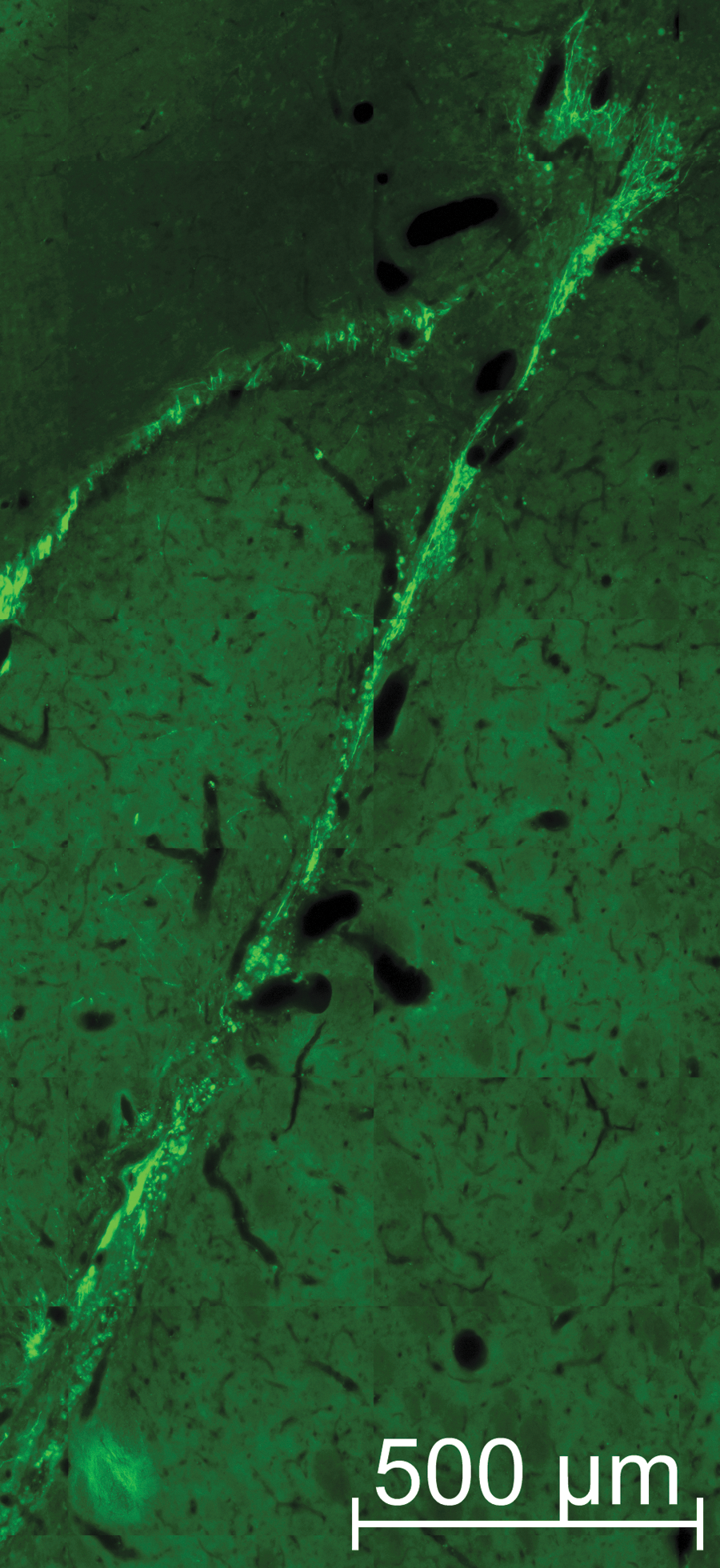
Neuroblasts in the implant track exhibit complex neural morphology. To visualize the neural-like structures of the new cells in the implant region, a high magnification image of the area was generated. Dcx+ cells (green) exhibit long neurites along the implant path and many show complex branching, especially those migrating toward and into the corpus callosum. The length of many of these neurites exceeded 100 μm.
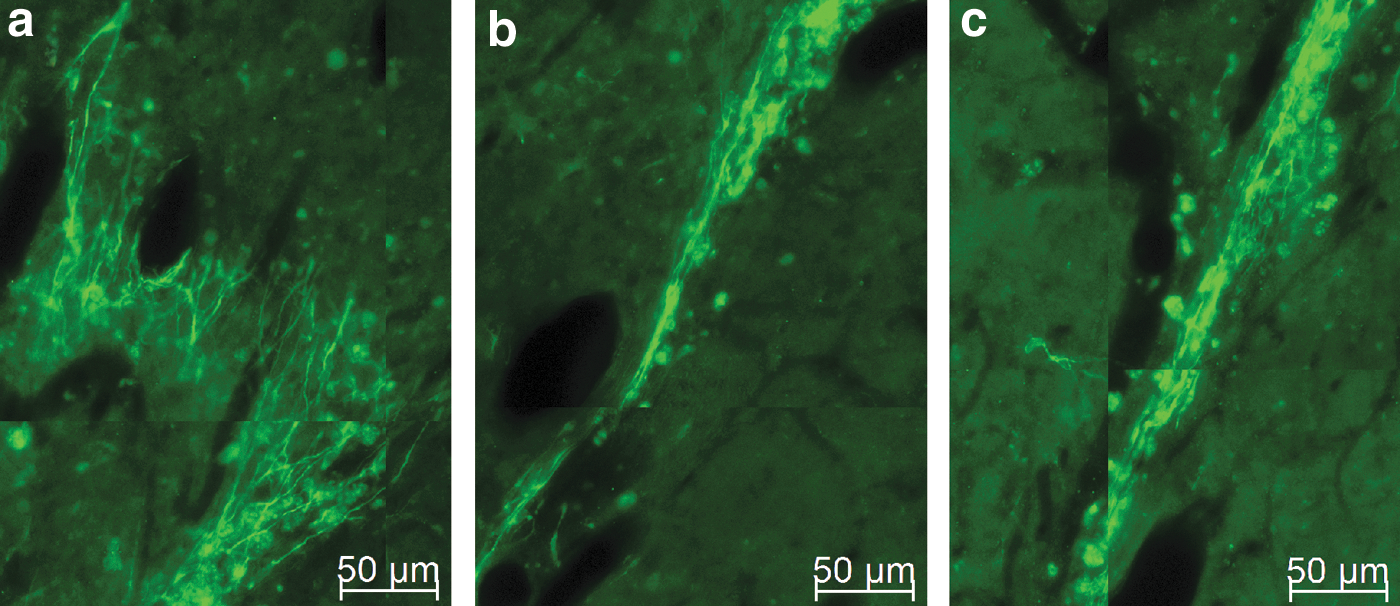
Higher magnification of neuroblasts and neurites in the implant region. Neuroblasts are prevalent along the implant region and their long neurites project parallel to the original, now degraded, cylinder.
Additional Dcx+ cells were observed in implanted striatum in regions lateral to the lateral ventricle, between the implant path and the ventricle wall (Fig. 5b), which were not observed in the corresponding contralateral striatum (Fig. 5a). These new cells may represent occult neural stem cells residing in the striatum that were stimulated to differentiate due to the implanted microenvironment, or they may have arisen from the lining of the ventricle and migrated directly toward the implant. We noted that many Dcx+ cells in this intrastriatal region possess long (>100 μm) branched neurites with morphology consistent with synaptogenesis (visible upon increased magnification), which possibly suggests the beginning of functional integration of the new neural cells into the existing circuitry [18,53,54].

Additional recruitment of neuroblasts between the lateral ventricle and implant site.
The region of the striatum containing the highest density of new cells was a clearly defined path from the RMS to the bottom of the cylindrical implant region in the ventral striatum. It is of interest to note that, although the cylinder implant also led through the cortex to the dorsal-most aspect of the brain, far fewer new cells were found in the cortical region. Although neurites were clearly visible leading into the cortex, they did not extend significantly into this region, despite being in the implant track (Figs. 2 and 3). Finally, the response of the endogenous SVZ-derived cells to the implant was dependent upon the presence of neurogenic factors; no new neural cells, including dopaminergic (TH+) neurons, were observed in factor-free controls (Fig. 6), although there does appear to be a disruption of the NF160-labeled “pencil tracts” [55] in the implanted striatum (Fig. 6a).
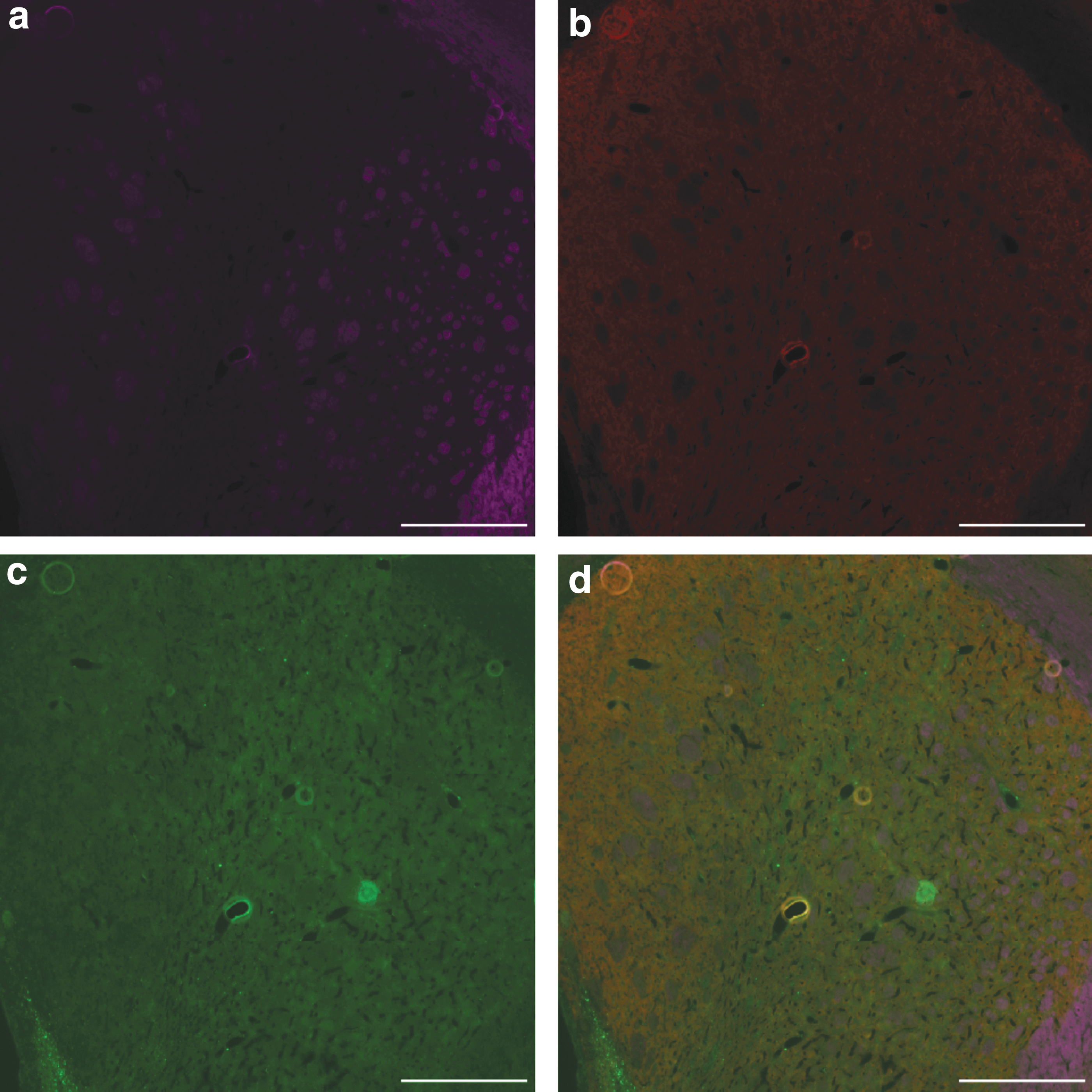
Immunohistochemistry of striatal region of control implant, containing only fibrin, aprotinin, and laminin.
In vivo neural engineering corrects hemiparkinson rat rotational behavior
The robust neurogenic response to these brain implants suggested a therapeutic application for this technology. We predicted that in an animal model of neurodegeneration, the fibrin cylinders would guide endogenous NPCs into damaged/dysfunctional nonneurogenic regions; these cells would then functionally integrate and restore lost function. We chose the 6-OHDA animal model of Parkinson's disease since the anatomical location of the defect is well characterized (striatum, nigrostriatal pathway, and substantia nigra), the specific neural phenotype that is necessary for therapeutic efficacy is readily assessed (labeling for TH), and correction of the behavioral deficiency is quantifiable (rotometer analysis).
Animals underwent unilateral 6-OHDA lesions to generate hemiparkinson rats and subsequently were subjected to in vivo neural engineering by these novel implantable fibrin matrices. This experimental therapeutic approach appeared to correct the asymmetric rotational defect in these animals (Fig. 7a). Immunohistochemical analyses of brains from corrected animals showed cells in the implanted striatum that were decorated with antibodies against TH, suggesting a dopaminergic phenotype (Fig. 7b–d). Control, lesioned animals implanted with fibrin cylinders containing only laminin and aprotinin (and lacking NGF and VEGF) were not corrected by the procedure.

Fibrin implants correct amphetamine-induced ipsilateral rotations in unilateral 6-OHDA-lesioned rats.
Discussion
Endogenous NPCs from the adult mammalian SVZ/RMS are an untapped source of therapeutic neurons. The restorative potential of these adult neural stem cells can be realized if a portion of the migrating neuroblasts can be steered away from their usual destination (the OB) and into nonneurogenic regions of the brain where disease or injury has reduced the number of functional neurons. The long-term goal of our research is to develop a means to retask these endogenous neural stem cells in the adult brain by engineering new, relatively long migratory paths that lead these cells into nonneurogenic regions of the brain.
We describe, in this study, a biocompatible, biodegradable, modifiable, and implantable matrix, in the shape of a cylinder, which can be surgically placed in the brain such that the implant connects the dorsoposterior RMS with the nonneurogenic striatum. These implants were indeed capable of creating long tracts and enriched regions of new neural cells in the striatum. Such in vivo brain tissue engineering has clear clinical applications for a variety of neurological disorders, especially considering that the source of stem cells would be the patient's own brain, without the need to actually implant any cells whatsoever. We believe that these deliberately targeted neurons will be repurposed from an olfactory role to a new function specified by the nature of the neurological dysfunction and the signaling cues within the milieu of the targeted niche [56 –59].
Cell-replacement therapy in the brain has many hurdles that must be overcome and the work described in this study addresses several of these challenges. Engineered fibrin cylinders can provide both the appropriate three-dimensional architecture and biochemically defined microenvironment to create a migratory path with complex neural cells and vasculature. Upon implantation, this serves to recruit endogenous neural stem cells from their usual location and subsequently target them into specific regions of the brain. Harnessing endogenous adult neural stem cells eliminates the problems associated with exogenous cell transplants, which include deleterious immune responses and rejection, unavailability of sufficient numbers of cells for transplantation, low survival and integration rates, and, in the case of embryonic or induced pluripotent stem cells, teratoma formation.
We present three key findings in this report. First, as a result of the fibrin implant containing neurotrophins (NGF and VEGF), ECM (laminin I), and protease inhibitor (aprotinin) in normal rats, significant numbers of neuroblasts were found along an extensive (>2.5 mm long) region of the implant track in the striatum, in the striatal region between the implant track and the lateral ventricle, in the corpus callosum, and, to a much lesser degree, in the cortex (Figs. 1 –5). Fibrin alone was not sufficient to elicit this effect in terms of attracting neural cells into the implant region [compare control animals (Fig. 6) to implanted animals (Figs. 2 –5)] or correcting hemiparkinsonian behavior. Only when VEGF and NGF were incorporated into the cylinders did we observe neuroblasts (Dcx+) along the track of the implant, which itself had essentially completely degraded. Second, there were no obvious deleterious neurological sequelae as a result of the implants. We have not observed necrosis, tumors, reactive astrocytes (GFAP+), or abnormal behavior in more than 80 animals that have been used during the development of this approach. Regarding inflammation, we attempted to minimize such cellular responses (GFAP+ astrocytes) by the administration of an anti-inflammatory agent following the surgery; our unpublished preliminary studies show that GFAP labeling is induced by the surgical procedure and can be subsequently significantly reduced by postoperative dexamethasone administration. Third, 6-OHDA hemiparkinson rats exhibited an attenuation of amphetamine-induced rotations following cylinder implantation (Fig. 7). This provides proof of concept for the idea of creating a new neural migratory stream that connects an abundant source of neural stem cells (the SVZ/RMS) to a dysfunctional brain region.
It is noteworthy to compare the morphology of the Dcx-reactive cells along the implant tract (Fig. 4) with Dcx+-labeled cells along a bona fide migratory path. We observe Dcx+ neuroblasts with long neurites that run parallel to the direction of the implant. Such labeling and morphology are very similar to that observed for Dcx+ cells undergoing chain migration along the rodent RMS [60]. It is reasonable to speculate that the neuroblasts that respond to the fibrin implant are undergoing similar migration. The fact that the animals were sacrificed 4 weeks after surgery and Dcx+ neuroblasts were still present along the length of the implant track suggests that the new migratory path is relatively stable over the postoperative time frame.
The likely source of new cells that were in abundance along the implant region is the RMS, which the cylinder was designed to intersect after implantation. It is not clear to us from where the cells that were seen in the striatal region between the implant and the lateral ventricle are derived. We suggest that they may be either from the wall of the lateral ventricle or are occult neural stem cells resident in the striatum that were activated by the implant. Regardless, in addition to the neural cells along the implant track, the new cells in this intrastriatal region are likely to be therapeutically beneficial.
It is of interest to note that, compared to the robust generation of neuroblasts in the striatal region of the implant, there were no nascent neural cells in the cortex. We speculate that the brain itself provides additional signaling molecules that synergize with the factors in the fibrin cylinder, and differential neurogenic responses are due to the biochemical cues inherent to the adult cortical environment versus those of the striatum. Indeed, these different regions follow different neurogenic mechanisms during brain development [61 –63] and this may account for the differential response (cortex vs. striatum) to the implanted fibrin microenvironment.
In conclusion, we present the development and therapeutic application in an animal model of neurodegeneration of an implantable, biocompatible, biodegradable fibrin-based cylinder, which can be readily modified with immobilized factors to create a bioengineered microenvironment that creates de novo neural migratory paths for in vivo neural tissue engineering. Past efforts to accomplish similar goals have involved the injection of fibrin, usually with factors and exogenous stem cells, into regions of the brain or spinal cord, which then polymerizes in situ [4,39,64,65]. It is difficult to envision the ability to create new, long migratory paths, which redirect endogenous neural stem cells with such approaches. The safety of these implants remains to be characterized in detail, but we have not observed obvious behavioral problems, mortality, tumor-like masses, significant damage to brain tissue, or other gross abnormalities upon the immunohistological analysis of the implanted brains.
Importantly, the main components of the implant described in this study are currently used in both routine and experimental clinical procedures. Fibrin (derived from fibrinogen and thrombin) and aprotinin [36] are common in many surgical procedures, including neurosurgery. NGF, one of the two neurotrophic factors included in these fibrin implants, has been delivered to patient's brains in clinical trials for traumatic brain injury and Alzheimer's disease [66], while VEGF is being considered for stroke therapy [29,67,68]. The recruitment of endogenous neural stem cells into regions of the brain that typically do not have neurogenic potential represents a powerful therapeutic approach for neurological diseases and disorders caused by the loss of specific neurons. The ability to deliberately steer significant numbers of endogenous NPCs from their usual niche into distant nonneurogenic regions of the brain represents a significant advancement for Parkinson's disease, traumatic brain injury, stroke, and other neurological disorders of similar etiology.
Footnotes
Acknowledgments
We thank M. Hobbs and A. Shaver for technical assistance with immunohistochemistry. This work was supported by National Science Foundation grants 1511928 (to E.M.P.) and 1003907; NASA WVSGC Fellowship to A.B.C.; WV SURE Award to A.R.C.; and NASA WVSGC Fellowship to L.E.H.
Author Disclosure Statement
No competing financial interests exist.