
Abstract
Cardiomyocytes (CMs) derived from induced pluripotent stem cells (iPS-CMs) are promising candidates for cell therapy, drug screening, and developmental studies. It is known that iPS-CMs possess immature electrophysiological properties, but an exact characterization of their developmental stage and subtype differentiation is hampered by a lack of knowledge of electrophysiological properties of native CMs from different developmental stages and origins within the heart. Thus, we sought to systematically investigate action potential (AP) properties of native murine CMs and to establish a database that allows classification of stem cell-derived CMs. Hearts from 129S2PasCrl mice were harvested at days 9–10, 12–14, and 16–18 postcoitum, as well as 1 day, 3–4 days, 1–2 weeks, 3–4 weeks, and 6 weeks postpartum. AP recordings in left and right atria and at apical, medial, and basal left and right ventricles were performed with sharp glass microelectrodes. Measurements revealed significant changes in AP morphology during pre- and postnatal murine development and significant differences between atria and ventricles, enabling a classification of developmental stage and subtype differentiation of stem cell-derived CMs based on their AP properties. For iPS-CMs derived from cell line TiB7.4, a typical ventricular phenotype was demonstrated at later developmental stages, while there were electrophysiological differences from atrial as well as ventricular native CMs at earlier stages. This finding supports that iPS-CMs can develop AP properties similar to native CMs, but points to differences in the maturation process between iPS-CMs and native CMs, which may be explained by dissimilar conditions during in vitro differentiation and in vivo development.
Introduction
O
It is a precondition for all applications that iPS-CMs have defined properties matching those of their native counterparts. It has been described that differentiated murine and human iPS-CMs tend to be immature compared with native CMs [8,15], including immature electrophysiological properties [16]. For the electrophysiological characterization of iPS-CMs as well as embryonic stem cell-derived CMs [17], action potential (AP) morphology is often used as an indicator for subtype differentiation and developmental stage classification [18,19]. However, an exact characterization is hampered by insufficient knowledge about electrophysiological properties of native CMs from different developmental stages and regions within the heart, even in mice. Single-cell AP recordings with the patch clamp technique in neonatal and adult atrial [20] as well as embryonic [21], neonatal, and adult ventricular [22] murine CMs showed significant changes in AP morphology during development, especially regarding repolarization. However, these studies have limited value for the characterization of stem cell-derived CMs for several reasons: (I) few time points of the broad spectrum from the embryonic to the adult stage were analyzed, (II) studies focused on atrial or ventricular cells, but did not include cells from both origins, (III) measurements were performed in single-cell preparations, which may have different properties compared with CMs in an intact syncytium due to cell–cell interactions and potential dissociation damage.
Owing to the importance of murine iPS-CMs for preclinical research and the lack of reliable criteria for classification of electrophysiological properties, we sought to systematically assess AP properties in murine atria and ventricles from the early embryonic to the adult stage with preserved in vivo-like tissue structure, with the goal to establish a database that allows the classification of developmental stage and subtype differentiation of murine iPS-CMs and other stem cell-derived CMs.
Materials and Methods
Preparation of atrial and ventricular tissue samples
All experiments conformed to the guidelines of the local animal welfare committee and to the Directive 2010/63/EU of the European Parliament. Mouse strain 129S2/SvPasCrl was used for the experiments. Hearts were resected at embryonic day 9 (E9), E10, E12, E14, E16, and E18 postcoitum and after 0–1/2, 1–2, 3–4, and 6 weeks postpartum (p.p.) as indicated. Atria were separated from the ventricles, stored as a whole in Dulbecco's modified Eagle's medium (DMEM; Life Technologies, Carlsbad, CA) at 37°C, and bubbled with 95% O2 and 5% CO2 for 30 min before AP recordings. Ventricles not older than E12 were also stored as a whole in DMEM at 37°C and bubbled with 95% O2 and 5% CO2 for 30 min before AP recordings. Ventricles older than E12 were not sufficiently supplied by diffusion. Therefore, they were separated from the atria and dissected into 150-μm-thick viable tissue slices as described below before electrophysiological recordings were performed.
Preparation of ventricular heart slices
Ventricular heart slices were prepared as described before [23,24]. Briefly, hearts were resected and perfused with ice-cold Tyrode solution (composition in mM: NaCl 136, KCl 5.4, NaH2PO4 0.33, MgCl2 1, glucose 10, Hepes 5, 2,3-butanedione monoxime 30; pH 7.4 adjusted with NaOH; all chemicals were purchased from Sigma Aldrich, Taufkirchen, Germany). Ventricles were separated from the atria and embedded in 4% low melt agarose (Roth, Karlsruhe, Germany). Short-axis slices (150 μm) were cut with a microtome (Leica VT1000S; Leica Microsystems, Wetzlar, Germany) and stored in Tyrode solution containing 0.9 mM Ca2+ for 30 min on ice. Afterward, slices were transferred to DMEM at 37°C, aerated with 95% O2 and 5% CO2, and allowed to recover for another 30 min before AP recordings.
iPS cell culture and cardiac differentiation
The murine iPS cell line TiB7.4 was used, which was generated from murine tail tip fibroblasts isolated from 129S4/Sv4JaeJ x C57Bl/6 mice [25]. Cells were transfected with the α-PIG plasmid vector containing the PAC- (encoding puromycin N-acetyl-transferase) and IRES (internal ribosomal entry site)-flanked eGFP gene under control of the α-Myosin Heavy Chain promoter (GenBank Accession No. U71441), as described previously [14]. iPS cells were grown on inactivated murine embryonic fibroblasts in DMEM supplemented with 15% fetal calf serum, 1 × nonessential amino acids, 2 mM
For cardiac differentiation, 1 × 106 iPS cells were suspended in 10-cm bacterial dishes in 14 mL Iscove's modified Dulbecco's medium (IMDM) supplemented with 20% fetal calf serum, 100 μM β-mercaptoethanol, 1× nonessential amino acids, and 50 μg/mL ascorbic acid and placed on a horizontal shaker for 2 days to allow embryoid body (EB) formation. After 2 days, 30,000 EBs were transferred to a spinner flask filled with 200 mL of IMDM differentiation medium. At day 9, 8 μg/mL puromycin (InvivoGen Europe, Toulouse, France) was added for cardiomyocyte purification, and medium with fresh puromycin was changed every second day. At day 16, purified iPS-CM clusters were dissociated into single CM with 0.25% trypsin-EDTA (Life Technologies) supplemented with 5 U/mL of DNAse I (Sigma-Aldrich, Taufkirchen, Germany). iPS-CMs were plated on fibronectin-coated (20 μg/mL for 2 h at 37°C; Sigma-Aldrich) 3-cm dishes (650,000 cells/dish) containing IMDM differentiation medium. Medium was changed every second or third day. AP measurements were performed 2 days, 6–7 days, 12–14 days, and 19–23 days after plating (ie, 18 days, 22–23 days, 28–30 days, and 35–39 days after initiation of cardiac differentiation) in oligolayers. The purity of the cell population was assessed by flow cytometry (Attune® Cytometer, Applied Biosystems by Life Technologies, Carlsbad, CA).
AP recordings
Intracellular AP recordings with sharp glass microelectrodes (20–50 MΩ resistance when filled with 3 M KCl; World Precision Instrument, Sarasota, USA) were performed as described before [14,23]. APs of native CMs up to E12 were measured in whole heart preparations in the left and right atria and at three different positions (apical/medial/basal) in the left and right ventricles (Fig. 1). In ventricles older than E12, AP measurements were performed midmyocardially in apical, medial, and basal heart slices. Atrial and ventricular samples were stimulated with a square pulse generator (Grass Technologies, Warwick, RI) as indicated. APs of iPS-CMs were measured in spontaneously beating oligolayers (Fig. 2) without external stimulation.
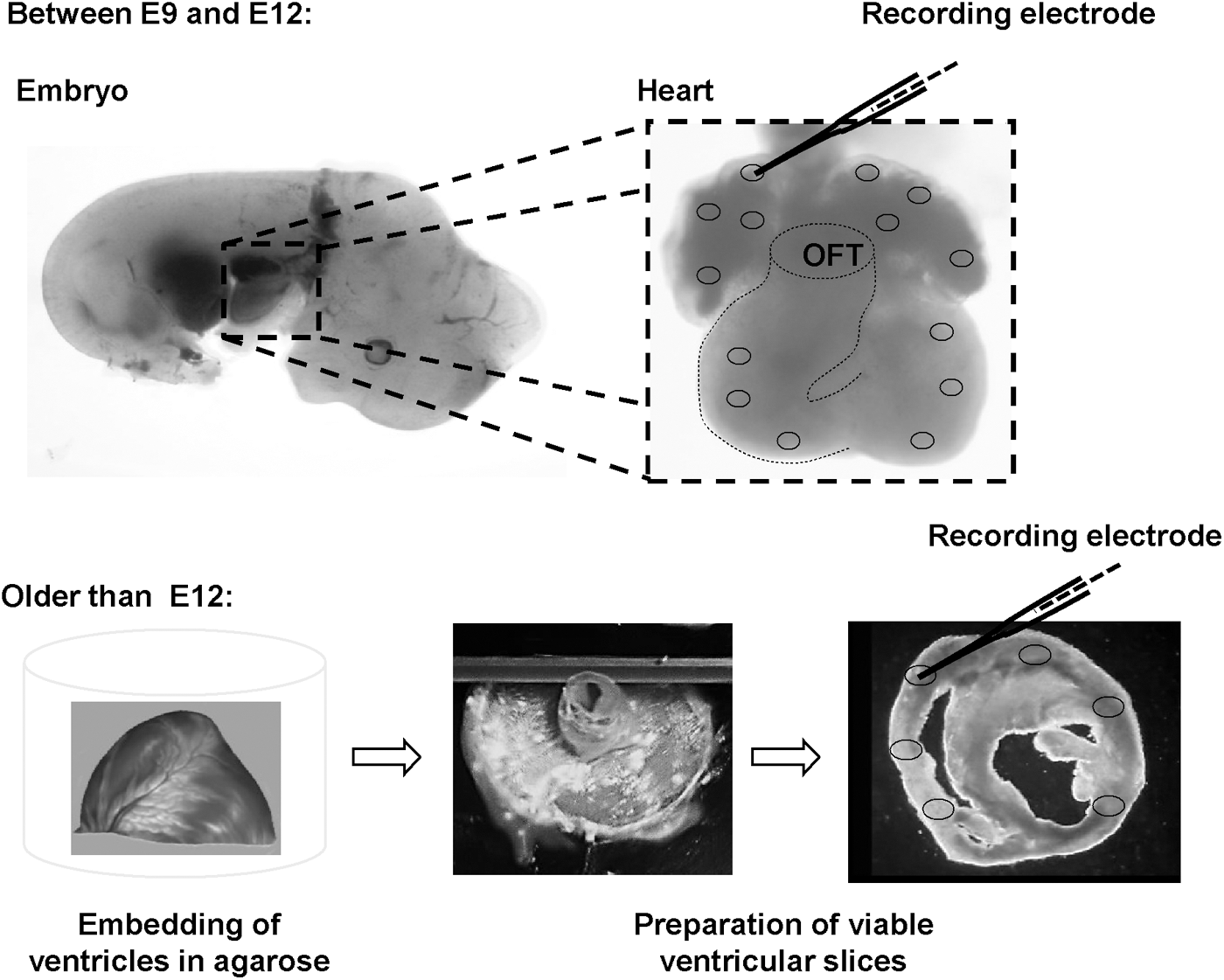
Preparation of native myocardium for AP recordings. Between embryonic day 9 (E9) and day 12 (E12), AP measurements with sharp glass electrodes were performed in whole hearts. Ventricles older than E12 were not sufficiently supplied by diffusion and therefore dissected into 150-μm-thick, viable, short-axis tissue slices before measurements. Afterward, AP recordings were performed in apical, medial, and basal slices. Atria older than E12 were resected from the ventricles, and recordings were performed in whole atria without slicing. AP, action potential; OFT, outflow tract.
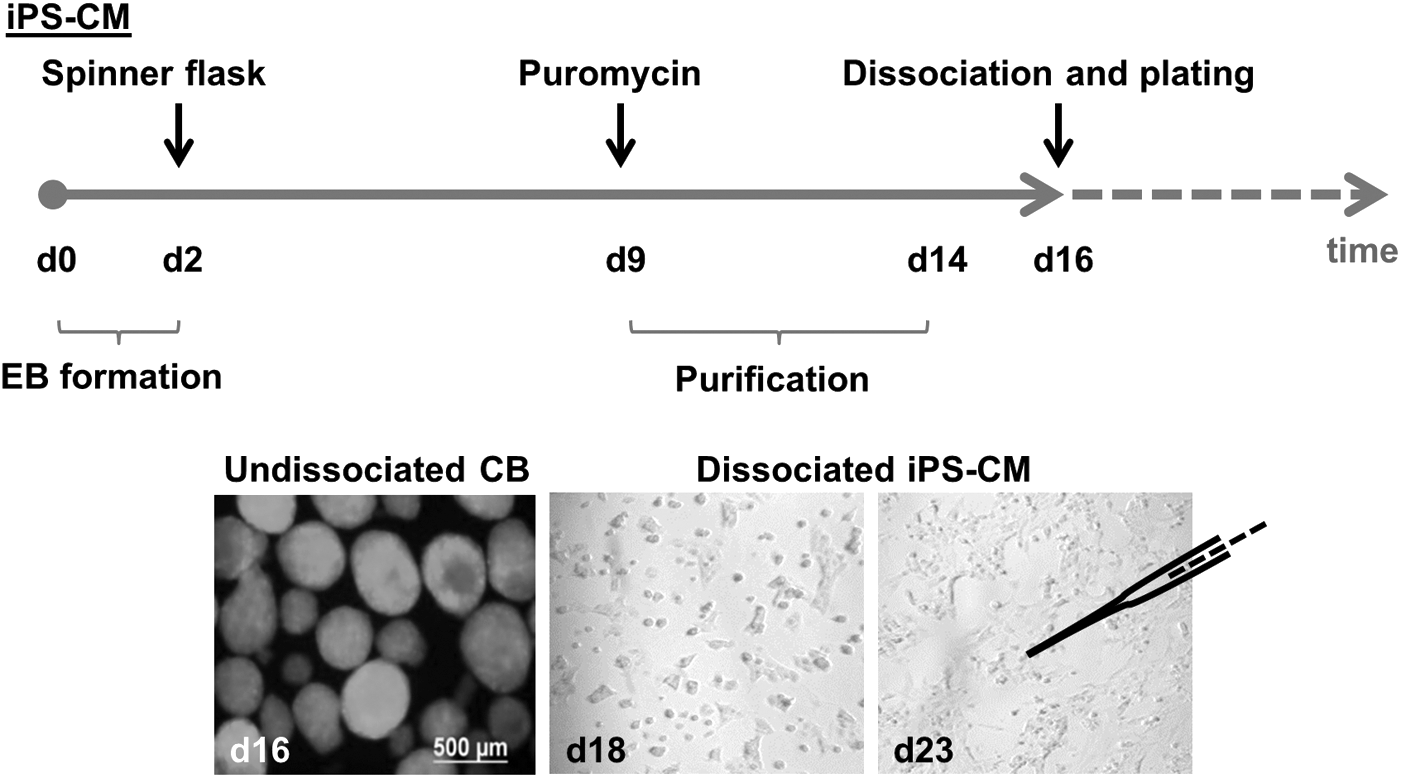
Differentiation and purification of iPS-CMs. iPS-CMs expressing eGFP and puromycin N-acetyl-transferase under control of the α–MHC promoter were differentiated in a spinner mass culture system and purified by addition of puromycin starting at day 9 of differentiation. Cells were dissociated and plated at d16 of differentiation. Afterward, AP measurements were performed at different time points. EBs, embryoid bodies; CBs, cardiac bodies; iPS-CMs, cardiomyocytes derived from induced pluripotent stem cells.
The recording electrode was connected to an SEC-10LX amplifier (npi electronic, Tamm, Germany), and the signal was acquired with the Pulse software (HEKA, Lambrecht/Pfalz, Germany). Data were analyzed with Mini Analysis (Synaptosoft, Fort Lee, NJ).
Statistics
All data are presented as mean ± SEM. Differences of AP parameters were tested for statistical significance by one-way ANOVA with post-test. All other data were tested for statistical significance by Student's t-test. Correction of P-values for multiple comparisons was performed using the Benjamini and Hochberg procedure for controlling the false discovery rate of a family of tests. A two-sided P-value <0.05 was considered statistically significant. SigmaStat (Systat, Erkrath, Germany), GraphPad Prism (GraphPad Software, San Diego, CA), and SPSS Statistics version 23 (IBM, Armok, NY) were used for all calculations.
Results
AP parameters at different regions within the heart
Statistical analyses of AP recordings showed no significant differences between evaluated left and right atrial AP parameters (Supplementary Tables S1 and S2; Supplementary Data are available online at
Developmental changes of atrial AP properties
AP characteristics of the atria changed significantly during development. At a stimulation frequency of 2 Hz, the maximal diastolic potential (MDP) decreased from −61.9 ± 2.2 mV at E9-10 (n = 12) to −76.9 ± 0.9 mV at 3–4 weeks p.p. (n = 31, P < 0.05). AP duration at 50% of repolarization (APD50) decreased from 29.0 ± 3.1 ms at E9-10 to 6.0 ± 1.2 ms at 3–4 weeks p.p. and increased slightly to 10.4 ± 1.1 ms at 6 weeks (n = 15, P < 0.05). AP duration at 90% of repolarization (APD90) decreased from 59.0 ± 2.4 ms at E9-10 to 21.3 ± 0.8 ms at 1–2 weeks p.p. (n = 47, P < 0.05) and increased slightly to 34.5 ± 3.5 ms at 6 weeks, resulting in a consistent decrease in APD50/90 ratio from 50.2% ± 5.7% at E9-10 to 30.3% ± 1.4% at 6 weeks after birth (P < 0.05). These results are presented in Figs. 3 and 4 and Supplementary Table S7.
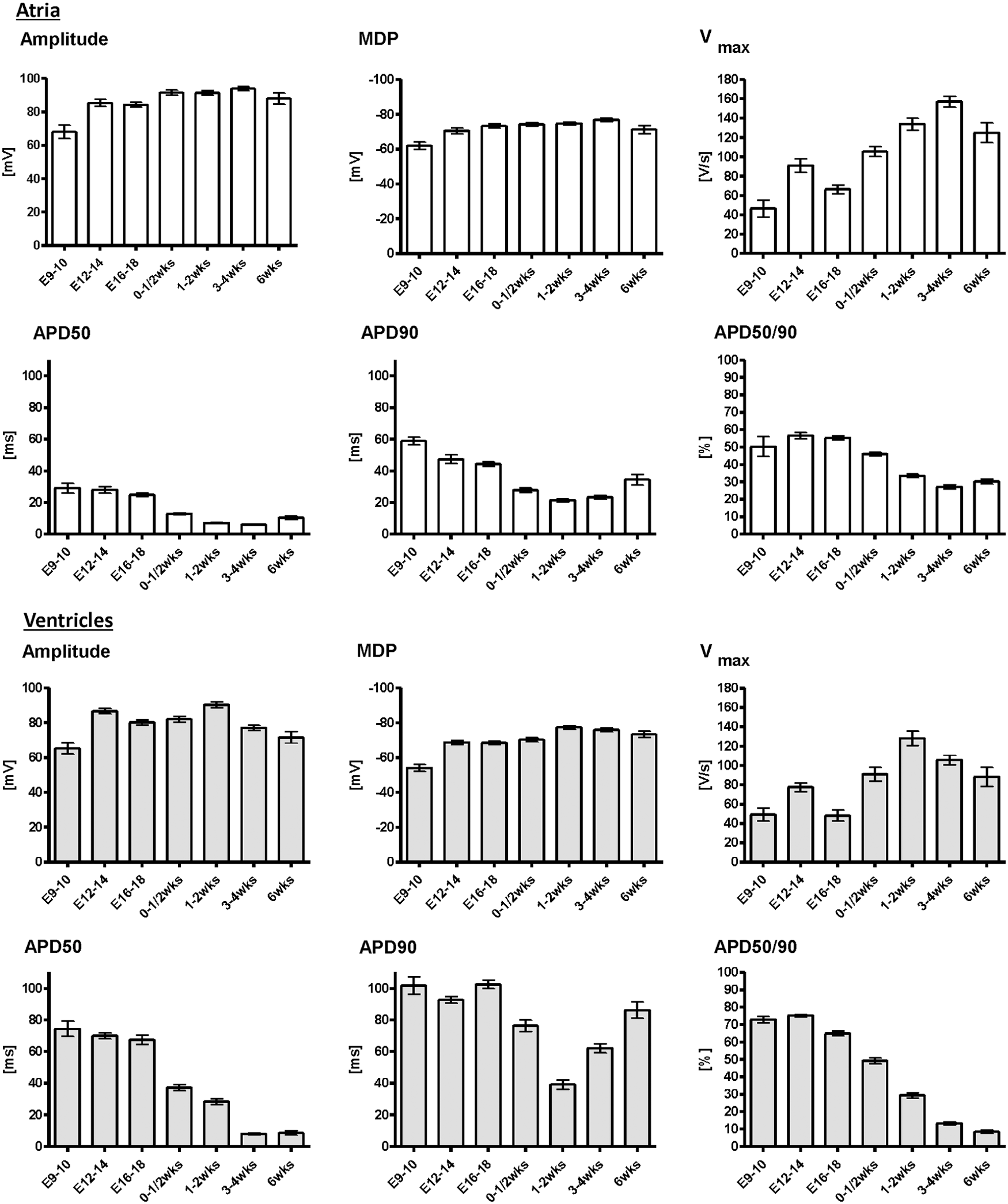
Statistical analysis of AP parameters from native atrial (white columns) and ventricular (gray columns) cardiomyocytes. APs of native CMs underwent significant changes during development. There were significant differences between CMs of atrial and ventricular origin. Data were pooled from left and right atria, as well as apical, medial, and basal left and right ventricles, since there were no significant differences between CMs from these locations (Supplementary Tables S2, S5, and S6). APD 50 and APD 90: action potential duration at 50% and 90% of repolarization, APD50/90: ratio of APD50 and APD90, E9-10: days 9–10 postcoitum, MDP, maximal diastolic potential; Vmax, maximal upstroke velocity; wks, weeks postpartum.
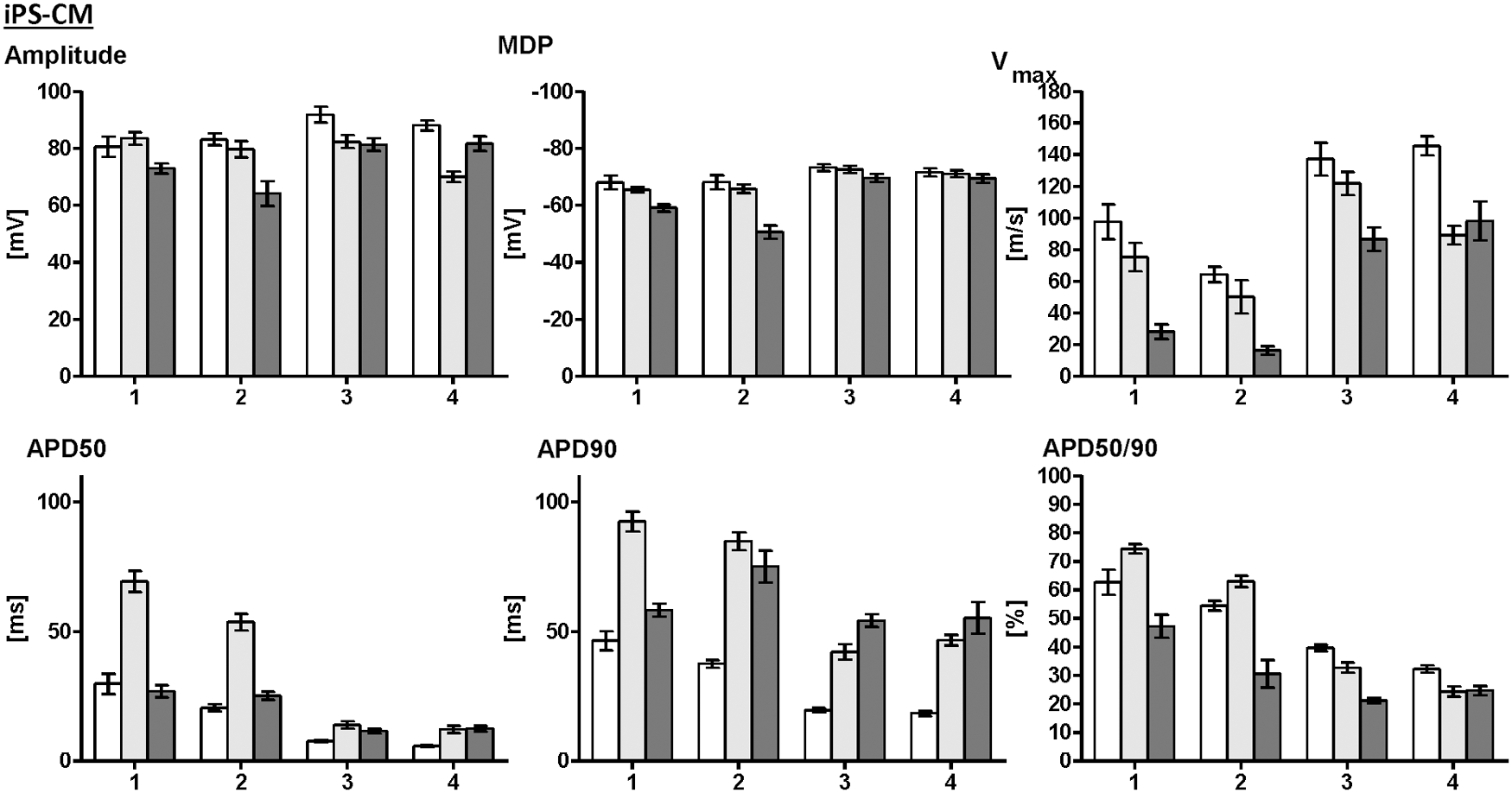
AP parameters of native atrial (white bars) and ventricular (gray bars) CMs in comparison with iPS-CMs (dark gray bars). Four different developmental stages are compared, native atrial and ventricular CMs at E12-14 versus iPS-CMs at d18 (1), native CMs at d16-18 versus iPS-CM at d22-23 (2), native CMs at 1–2 weeks p.p. versus iPS-CMs at d28-30 (3), and native CMs at 3–4 weeks p.p. versus iPS-CMs at d35-39 (4). While iPS-CMs showed significant differences from both atrial and ventricular native CMs at developmental stages 1–3, iPS-CMs possessed AP parameters characteristic for ventricular CMs at developmental stage 4.
After obtaining AP parameters at 2 Hz, the stimulation frequency was increased to 10 Hz without interrupting the AP recording. At a stimulation frequency of 10 Hz, AP parameters of the atria showed similar developmental changes as with a stimulation frequency of 2 Hz (P < 0.05 for all parameters): MDP decreased from E12-14 to 3–4 weeks p.p. The APD50 and APD90 decreased from E12-14 to 3–4 weeks p.p. and increased slightly to 6 weeks, resulting in a consistent decrease in APD50/90 ratio from E12-14 to 6 weeks after birth. AP parameters and P-values at 2 Hz versus 10 Hz are presented in Supplementary Tables S7 and S8.
Developmental changes of ventricular AP properties
Similar to the electrophysiological alterations during atrial development, significant changes in ventricular AP parameters occurred during development. Stimulated with 2 Hz, MDP of ventricular CMs decreased from −54.0 ± 2.0 mV at E9-10 (n = 16) to −77.3 ± 1.1 mV at 1–2 weeks p.p. (n = 31, P < 0.01) and remained unchanged thereafter (Figs. 3 and 4 and Supplementary Table S8). APD50 decreased from 74.4 ± 4.8 ms at E9-10, when it was considerably longer than in atrial CMs, to 8.6 ± 4.5 ms at 6 weeks p.p. (n = 22, P < 0.01). APD90 decreased from 101.8 ± 5.5 ms at E9-10 to a minimum of 39.3 ± 2.5 ms at 1–2 weeks p.p. During further development, an increase of APD90 to 62.1 ± 3.0 ms at 3–4 weeks p.p. and 86.7 ± 4.5 ms at 6 weeks was observed (P < 0.01), resulting in a plateau phase, which was the most obvious morphological difference between late-stage atrial and ventricular CMs. Accordingly, there was a decrease of APD50/90 ratio from 72.9% ± 1.8% at E9-10 to 9.7% ± 1.4% at 6 weeks p.p. (P < 0.01). AP parameters are presented in Figs. 3 and 5 and Supplementary Table S9.
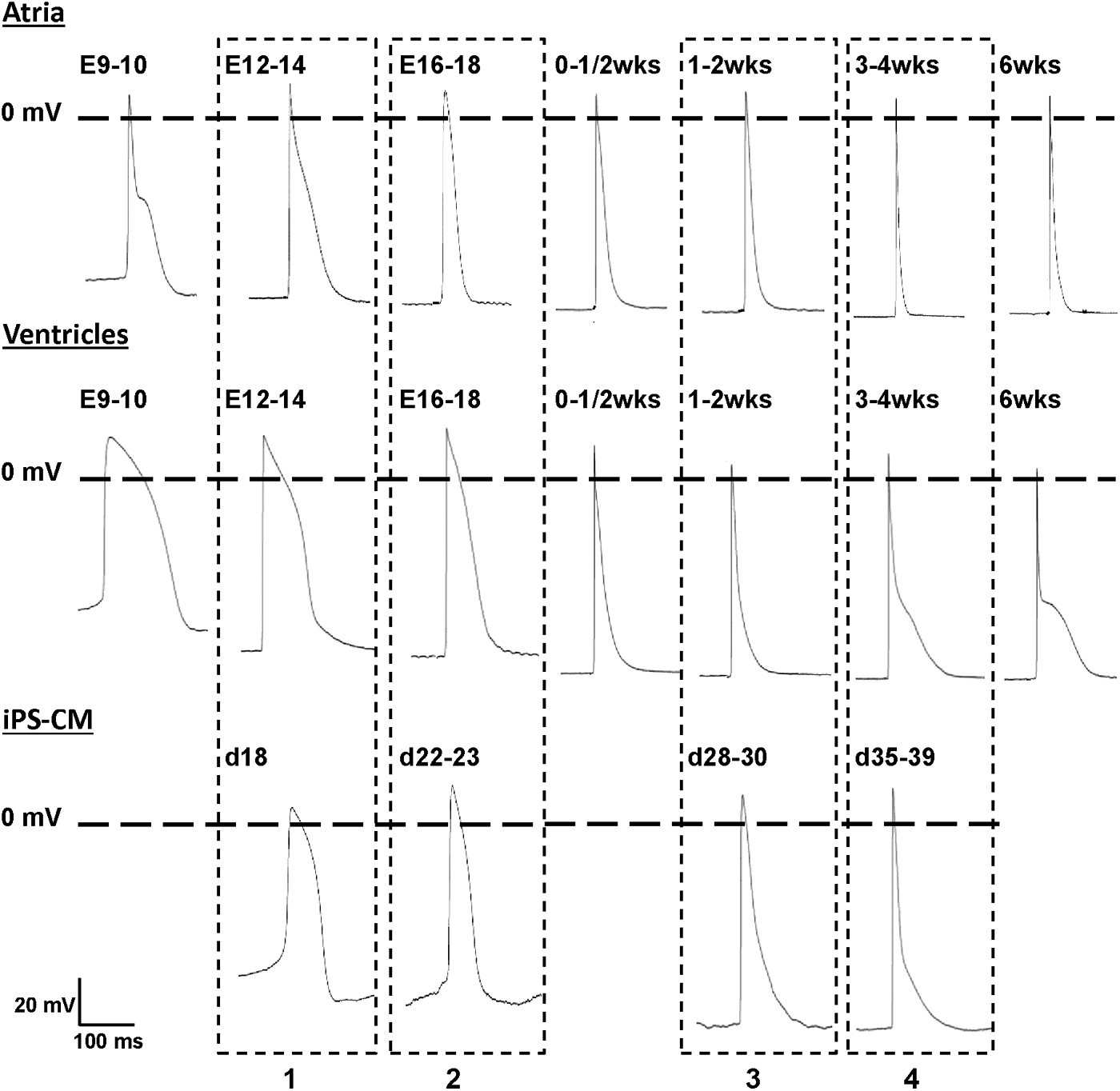
Representative APs of native atrial (upper row), native ventricular (middle row), and iPS-derived (lower row) CMs. Dashed boxes highlight the comparison of AP morphology at four developmental stages of iPS-CMs and corresponding native CMs.
After increasing the stimulation frequency to 10 Hz, AP parameters of the ventricles maintained similar developmental changes as with a stimulation frequency of 2 Hz (P < 0.01 for all parameters). MDP decreased from E12-14 to 1–2 weeks p.p. and remained unchanged thereafter (Supplementary Tables S9 and S10). APD50 decreased from E12-14 to 3–4 weeks p.p., APD90 decreased from E12-14 to a minimum value at 1–2 weeks p.p. and increased afterward until 6 weeks p.p. There was a consistent decrease of APD50/90 ratio between E12-14 and 6 weeks p.p.
Frequency-dependent variations of AP parameters
After increasing the stimulation frequency from 2 Hz to 10 Hz, amplitude, MDP, APD50, and APD90 showed only minor numerical changes in atrial tissue (Supplementary Table S7), and P-values indicated significance just at individual time points, but not consistently throughout development (Supplementary Table S8).
In contrast, changes in ventricular AP parameters were more pronounced. Compared with 2 Hz, there was a numerical decrease of amplitude and MDP at 10 Hz at all developmental stages, which reached statistical significance at the majority of analyzed stages. Increasing the stimulation frequency to 10 Hz led to a decrease of APD50 at early stages, but increased it at late stages. APD90 decreased at 10 Hz at all developmental stages except at 1–2 weeks p.p. when APD90 reached minimal values during development as described above and stayed equal at 2 and 10 Hz. Corresponding to changes in APD50 and APD90, the APD50/90 ratio decreased at early stages, but increased at late stages, when the stimulation frequency was increased. Ventricular AP parameters at 2 Hz and 10 Hz with corresponding P-values are shown in Supplementary Tables S9 and S10.
AP properties of iPS-CMs
iPS-CMs underwent significant changes in AP morphology between d18 and d39 after the initiation of differentiation (Figs. 4 and 5). Spontaneous beating frequency of plated iPS-CMs did not change between d18 and d28-30 (d18: 6.3 ± 0.3 Hz, d22-23: 4.9 ± 0.3 Hz, d28-30: 6.9 ± 0.9 Hz, P = 0.48), but increased to 10.6 ± 1.2 Hz at d35-39 (P < 0.01 vs. all other groups). The MDP of iPS-CMs increased from −59.1 ± 1.3 mV at d18 to −69.4 ± 1.5 at d35-39 (P < 0.01). APD50 decreased from 26.9 ± 2.4 ms at d18 to 12.6 ± 1.1 ms at d35-39 (P < 0.01), while APD90 showed no changes between d18 and d35-39 (58.2 ± 2.5 ms vs. 55.3 ± 6.1 ms, P = 0.52). APD50/90 ratio decreased from 47.3% ± 4.0% at d18 to 21.2% ± 1.1% at d28-30 and 24.7% ± 1.7% at d35-39 (P < 0.01).
Systematic comparison of AP properties of induced pluripotent stem cell-derived and native CMs
Direct comparison of AP parameters between native atrial and ventricular CMs and iPS-CMs required similar beating frequencies since AP parameters depend on beating frequency. Therefore, data used for this comparison were obtained from native CMs with beating frequencies similar to that of spontaneously beating iPS-CMs.
iPS-CMs underwent similar developmental changes as native CMs (Fig. 4). In particular, MDP increased and APD20, APD50, and APD50/90 ratio decreased with time in both, iPS-CMs and native CMs. iPS-CMs at earlier developmental stages (d18 and d22-23) showed significant differences from native atrial and ventricular CMs, especially in MDP, Vmax, APD90, and APD50/90 ratio (Fig. 5). Later, at d28-30 and d35-39, iPS-CMs had typical AP properties of native ventricular CMs, while significant differences from atrial CMs persisted for APD20, APD50, and APD90 (Fig. 5 and Supplementary Table S11).
Discussion
In the present study, we systematically characterized AP properties of native murine atrial and ventricular CMs from the early embryonic to adult stage. Remarkable changes in AP morphology occurred during murine development, especially an increase in MDP, shortening of APD50, J-shaped development of APD90, and decrease of APD50/90 ratio. Knowledge of AP parameters at different developmental stages and at different regions within the native heart allows the classification of subtype differentiation and developmental stage of stem cell-derived CMs based on their functional properties.
AP properties of native murine CMs
A small number of previous studies have investigated AP properties of murine CMs at different developmental stages, but none of those studies has covered the spectrum from the early embryonic to the adult stage [20 –22,26]. Moreover, most previous studies focused on either ventricular or atrial CMs, but did not include CMs from both regions. Furthermore, the influence of beating frequency on AP parameters was frequently not considered. Available data on AP properties of mouse atrial [20] and ventricular [21,22,26] CMs during heart development are based on single-cell patch clamp analyses.
AP recordings in fetal ventricular CMs revealed APD shortening and stable MDP between embryonic days 10.5 and 17.5 [26] and embryonic day 18 and neonatal day 1 [21]. At comparable developmental stages, our study confirmed significant APD shortening, but showed increasing MDP. Studies of postnatal atrial and ventricular CMs described a significant decrease in APD between neonatal days 1, 7, 14, 20, and adult stage, which is consistent with our results on APD20 and APD50. In contrast to our study, no increase in APD90 after 1–2 weeks p.p. was described, thus AP morphology especially in ventricular CMs at the adult stage differed from our findings as there was no distinctive plateau phase in previous studies.
Sharp electrode recordings in ventricular slices of other mouse strains also showed a long APD90, thus the use of different mouse strains cannot explain different AP morphologies [27,28]. Methodical differences in sample preparation and type of recording might explain the differences between previous and present results. We used sharp electrode recordings in multicellular preparations because this technique allows an exact identification of the analyzed region within the heart, which was crucial for the present work. We did not use the patch clamp technique since it is not feasible in multicellular preparations with a high amount of extracellular matrix and would have required enzymatic dissociation of the tissue, which might cause cellular damage and potential electrophysiological alterations and would have made an exact identification of the analyzed region within the heart very difficult. One disadvantage of our approach is that in contrast to the single-cell patch clamp technique, it is not possible to record individual ion currents because membrane potential cannot be clamped in the electrically well-coupled syncytium.
Frequency-dependent variations of AP parameters in native CMs
Increasing the stimulation frequency from 2 Hz to 10 Hz without interrupting the AP recording led to changes in several AP parameters of atrial and especially ventricular CMs. The reduction of amplitude might be explained by the inactivation kinetics reported for cardiac voltage-dependent Na+ channels [29]. A significant decrease of amplitude was just found at early and late developmental stages; this may be explained by the J-shaped development of APD90, that is, a long AP duration in early and late stages, resulting in a shorter phase 4 and insufficient time between successive APs for a complete recovery of fast Na+ channels. The shortening of APD90 at high stimulation frequencies may be due to cumulative activation of delayed rectifier K+ channels, leading to an enhanced repolarizing current at higher frequencies [30].
Classification of subtype and developmental stage of stem cell-derived CMs
Application of iPS-CMs for disease modeling, drug studies, and as a source for cell therapy requires cells with defined properties matching those of their native counterparts. Therefore, a thorough characterization of stem cell-derived CMs is needed. Our database of atrial and ventricular AP properties in the developing mouse heart enables a classification of stem cell-derived CMs regarding developmental stage and subtype differentiation. To serve as an example, we have demonstrated that CMs derived from the iPS cell line TiB7.4 develop typical AP properties of ventricular CMs at later stages, but have properties different from atrial and ventricular CMs at early stages. This result might be cell line specific and does not represent iPS-CM maturation and subtype differentiation in general.
Several studies have classified AP properties of stem cell-derived CMs into nodal-, atrial-, and ventricular-like APs [31 –34]. In these studies, subtype classification is usually not based on comparative data from native CMs, but on assumptive characteristics of CM subtypes, like a distinct plateau phase for ventricular CMs, triangle-like shape for atrial cells, and more positive MDP, diastolic depolarization, and lower Vmax for pacemaker cells [32,34]. However, our data indicate that these morphological criteria may only be present for a limited time period of development, but are not valid throughout the whole developmental process. Moreover, an overlap of AP properties of atrial and ventricular CMs may impede differentiation between subtypes at some developmental stages, for example, between early neonatal atrial and ventricular CMs. Thus, a reliable subtype classification of stem cell-derived CMs based on AP properties should refer to a systematic data set of native AP properties, and ideally the properties of stem cell-derived CMs should be assessed at different stages to demonstrate not only a similarity to a native subtype at one stage (which might be ambiguous) but also a similar development of AP properties.
Kuzmenkin et al. 2009 [31] used the single-cell patch clamp technique for AP recordings in CMs derived from two iPS cell lines and one ES cell line and compared them with fetal atrial and ventricular CMs at two different developmental stages (E10.5–12.5 and E16.5–18.5). Stem cell-derived CMs were harvested at days 10–12 and days 16–19 after starting the differentiation. The study showed a slight increase of MDP and decrease of APD90 in native atrial and ventricular CMs between the two stages, which is in line with our data. However, in this study, the AP parameters were compared at different spontaneous beating frequencies, hampering a reliable comparison of AP parameters at early and later stages. For earlier developmental stages of iPS-CMs and ES-CMs, the authors described a relevant number of atrial-like APs, while predominantly ventricular-like APs were found at later stages of the same cell lines, which is consistent with our findings.
Another earlier study described iPS-CMs at a late stage of differentiation as mainly ventricular-like cells too [35,36]. Changes in AP properties between days 11–16 and days 21–25 included an increase in MDP and shortening of APD50 and APD90, which are consistent with our data. Early-stage AP parameters showed significant differences from both native atrial and native ventricular CMs.
Taken together, the results on AP properties at early stages, which have been found by previous studies and the present work, question that stem cell-derived CMs cultured in vitro undergo the same maturation process like native CMs in vivo, although typical ventricular-like AP properties were found at later stages. Differences in the maturation process may be explained by differences in microenvironment, for example, the presence of nonmyocytes in vivo, but not in vitro, or the composition of the extracellular matrix. An influence of antibiotic selection on the AP properties of stem cell-derived CMs has also been described [37]. In addition, the density of in vitro cultured CMs may influence AP parameters since Du et al. described cell density-dependent characteristics of human iPS-CMs [38]. These authors also suggested that iPS-CMs may have electrophysiological properties of their own kind, meaning that AP morphological similarities to native CMs would be accidental and iPS-CMs may not match their native counterparts in other characteristics than their AP properties.
These findings impact the utilization of stem cell-derived CMs as a model system for drug development and toxicity testing as well as a source for cell therapy. Determination and adequate choice of subtype differentiation and developmental stage are of particular importance. For instance, iPS-CMs with ventricular-like characteristics, which were predominantly found in the present study, may be suitable candidates for regeneration of ventricular myocardium, but not for application as biological pacemaker or model for atrial fibrillation.
Limitations
The aim of our study was to provide a systematic overview of AP properties in different regions of the murine heart and at different developmental stages. Although (in terms of different analyzed stages and regions within the heart) the present study is the most detailed analysis of AP parameters in the murine heart conducted so far, a characterization of CMs from the conduction system is missing. Interestingly, all iPS-CMs analyzed at developmental stages later than d22-23 had stable resting membrane potentials, suggesting a very low differentiation toward pacemaker-like cells. At earlier stages (d18 and d22-23), APs with spontaneous diastolic depolarization and immature characteristics were found, but rather represented immature precursors of ventricular-like CMs than terminally differentiated nodal CMs. This is an important observation since such cells may be misinterpreted as pacemaker cells in studies focusing on stem cell-derived CMs at earlier stages.
Two techniques for tissue preparation needed to be applied for hearts of different stages: at early developmental stages up to E12, whole hearts could be used for experiments without further preparation, enabling optimal preservation of tissue structure and minimizing cellular damage. Since ventricles older than E12 were not sufficiently supplied by diffusion, viable ventricular slices were prepared for electrophysiological characterization of ventricles older than E12, which enable AP recordings in an in vivo-like tissue structure [23,27,39]. Slices of murine ventricles at embryonic, neonatal, and adult stages have been shown to possess physiological AP properties [24,27]. Nevertheless, ventricular slices are only 150 μm thick and could be stunned or damaged by the preparation, which may have influenced AP properties. We tried to minimize bias of preparation-related tissue damages by only including slices in our analyses that fulfilled specific quality criteria, that is, tissue followed a stimulation frequency of 10 Hz, and the structural integrity of the myocardium was confirmed by microscopy.
The use of multicellular native preparations may be advantageous for comparisons with stem cell-derived multicellular preparations such as cardiac bodies [40], engineered heart tissue [41], or transplanted CMs within host tissue [23,42]. On the other side, comparisons with single-cell preparations of stem cell-derived CMs may be biased by the influence of the syncytium. For instance, a previous study has shown a longer APD in single-cell recordings in comparison with tissue preparations [24]. Differences in AP parameters between tissue and single cells may be caused by extracellular K+ accumulation in tissue, which leaks out of damaged cells, or by gap junction disruption during dissociation [43].
In the present study, we used iPS-CMs that were trypsinized, plated on fibronectin covered culture dishes, and reaggregated to multicellular electrically coupled oligolayers. This procedure may also have affected electrophysiological properties, but it was necessary because addition of puromycin led to massive cell death of non-CMs within undissociated EBs, resulting in a high amount of cell debris, which hampered measurements in undissociated EBs later than 20 days after initiation of differentiation.
Conclusions
Significant changes in AP morphology occur during pre- and postnatal murine development. We performed a systematic analysis of AP properties of native murine CMs from all four heart chambers, from fetal to adult stages, and at low and high stimulation frequencies. These data enable a classification of developmental stage and subtype differentiation of pluripotent stem cell-derived CMs based on their AP properties. For CMs derived from the murine genetically modified murine iPS cell line TiB7.4 a typical ventricular phenotype was demonstrated at later developmental stages (days 35–39 after initiation of differentiation), while there were electrophysiological differences from atrial as well as ventricular native CMs at earlier stages. This finding points to differences in the maturation process between iPS-CMs and native CMs, which may be explained by dissimilar conditions during in vitro differentiation and in vivo development.
Footnotes
Acknowledgments
The authors thank Rudolf Jeanisch and Alexander Meissner (Whitehead Institute for Biomedical Research, Cambridge, MA) for providing the murine iPS cell line, Tib7.4. The authors acknowledge the excellent collaboration and support provided by the animal facility of the Institute of Neurophysiology and the Institute of Medical Statistics, Informatics and Epidemiology of the University of Cologne. This work was supported by Köln Fortune, the Hans und Gertie Fischer Stiftung, the Marga und Walter Boll Stiftung, and the Deutsche Stiftung für Herzforschung.
Author Disclosure Statement
All authors declare that they have no conflicts of interest.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.