
Abstract
Mesenchymal stromal cells (MSCs) are attractive agents for the prophylaxis of acute graft-versus-host disease (aGVHD) after allogeneic hematopoietic stem cell transplantation (allo-HSCT). However, safety concerns remain about their clinical application. In this study, we explored whether extracellular vesicles released from human umbilical cord-derived MSCs (hUC-MSC-EVs) could prevent aGVHD in a mouse model of allo-HSCT. hUC-MSC-EVs were intravenously administered to recipient mice on days 0 and 7 after allo-HSCT, and the prophylactic effects of hUC-MSC-EVs were assessed by observing the in vivo manifestations of aGVHD, histologic changes in target organs, and recipient mouse survival. We evaluated the effects of hUC-MSC-EVs on immune cells and inflammatory cytokines by flow cytometry and ProcartaPlex™ Multiplex Immunoassays, respectively. The in vitro effects of hUC-MSC-EVs were determined by mitogen-induced proliferation assays. hUC-MSC-EVs alleviated the in vivo manifestations of aGVHD and the associated histologic changes and significantly reduced the mortality of the recipient mice. Recipients treated with hUC-MSC-EVs had significantly lower frequencies and absolute numbers of CD3+CD8+ T cells; reduced serum levels of IL-2, TNF-α, and IFN-γ; a higher ratio of CD3+CD4+ and CD3+CD8+ T cells; and higher serum levels of IL-10. An in vitro experiment demonstrated that hUC-MSC-EVs inhibited the mitogen-induced proliferation of splenocytes in a dose-dependent manner, and the cytokine changes were similar to those observed in vivo. This study indicated that hUC-MSC-EVs can prevent life-threatening aGVHD by modulating immune responses. These data provide the first evidence that hUC-MSC-EVs represent an ideal alternative in the prophylaxis of aGVHD after allo-HSCT.
Introduction
C
Currently, a wide variety of drugs and methods are used to prevent aGVHD, and the prophylactic strategies have been greatly refined due to the in-depth understanding of the pathophysiology of aGVHD. However, the majority of allo-HSCT recipients suffer from this potentially life-threatening complication [1]. To date, no optimal aGVHD prophylaxis strategy has been proposed, and more effective prophylaxis for aGVHD is urgently needed.
Mesenchymal stromal cells (MSCs), namely mesenchymal stem cells, may be attractive candidates for cellular therapy during allo-HSCT due to their unique immunoregulatory and regenerative properties [4]. Although inconsistent data exist with respect to their prophylactic and therapeutic effects on aGVHD, several clinical trials have indicated that the adoptive transfer of MSCs prevents life-threatening aGVHD after allo-HSCT [5 –7]. In addition, MSC infusion in addition to conventional immunosuppression was associated with an overall response rate of 47%–75% for steroid-refractory aGVHD patients who have poor survival outcomes [2,8 –10]. MSC treatment resulted in lower chronic GVHD incidence and severity [10]. Moreover, MSCs may promote hematopoietic stem cell engraftment [11 –13], which is a major concern, especially when using umbilical cord (UC) blood as grafts [14]. However, safety concerns remain about the clinical application of MSCs, mainly due to their uncontrolled differentiation and unwanted long-term side effects [15], and the administration of MSCs is associated with an increased risk of pneumonia-related death after allo-HSCT [16].
Emerging evidence indicates that paracrine effects, which are mediated by extracellular vesicles (EVs), play a key role in MSC-based therapy [17], and an increasing number of preclinical and clinical studies have indicated that MSC-derived exosomes and microvesicles are an attractive alternative to MSCs in the fields of tissue regeneration and immunomodulation [18 –26]. Exosomes and microvesicles are two key classes of EVs that play an important role in cell-to-cell communication in a pleiotropic manner [17]. Exosomes are generated upon the fusion of multivesicular bodies with the plasma membrane and range in size from 30 to 120 nm, whereas microvesicles are generated by direct outward budding of the plasma membrane and range in size from 50 to 1,000 nm [17,27].
Recently, Kordelas et al. utilized MSC-derived exosomes to treat one therapy-refractory aGVHD patient, and the clinical manifestations of aGVHD were dramatically alleviated [25]. Given these encouraging results, we aimed to explore whether EVs released from human UC-derived MSCs (hUC-MSC-EVs) can prevent aGVHD after allo-HSCT in a mouse model.
Materials and Methods
Isolation of hUC-MSC-EVs
hUC-MSCs were obtained from the Department of Experimental Hematology, Beijing Institute of Radiation Medicine. The characteristics of hUC-MSCs have been previously reported [28]. Culture supernatants were collected after hUC-MSCs were cultured for 48 h in a fetal bovine serum-free medium. EVs were harvested as previously described, with minor modifications [18]. Briefly, culture supernatants were centrifuged at 2,000 g for 30 min to remove cells and debris. The cell-free supernatants were then ultracentrifuged at 100,000 g using a Beckman Optima XPN-100 Ultracentrifuge with a 45 Ti fixed angle rotor (Beckman Coulter) for 2 h at 4°C to isolate EVs. The pelleted EVs were suspended in sterile phosphate-buffered saline (PBS) and pelleted in a second ultracentrifugation step under the same conditions, and the final EV pellets were suspended in PBS and stored at −80°C until use.
Characterization of hUC-MSC-EVs
The characteristics of hUC-MSC-EVs were detected as previously described, with slight modifications [29,30]. The surface markers of hUC-MSC-EVs were detected by flow cytometric analysis. hUC-MSC-EVs were incubated with 4-μm-diameter aldehyde/sulfate latex beads (Molecular Probes, Life Technologies) overnight at 4°C under agitation. After stopping the reaction by incubation with glycine at room temperature (RT) for 30 min, EV-coated beads were washed thrice and then incubated with anti-CD9, anti-CD63, and anti-CD81 antibodies or corresponding isotype controls (BioLegend) for 40 min at RT. Stained EVs were detected using a flow cytometer (Becton Dickinson), and data were analyzed using FlowJo Software (Tree Star; version 7.6.1). For hUC-MSC-EV morphology, 16 μL of a hUC-MSC-EV suspension was dropped onto a formvar/carbon-coated 300-mesh copper grid, negatively stained with 3% aqueous phosphotungstic acid for 2 min, and observed by transmission electron microscopy (Hitachi) at 80 kV. The protein concentration of the hUC-MSC-EVs was quantified by the bicinchoninic acid (BCA) method.
Mice
Male BALB/c and C57BL/6 mice (8–10 weeks old) were purchased from Charles River Laboratory and were housed in a specific pathogen-free facility. All animal experiments were conducted in agreement with the Guide for the Care and Use of Laboratory Animals and were approved by the Ethics Committee of the Chinese PLA General Hospital.
Mouse model of allogeneic HSCT and hUC-MSC-EV prophylaxis
The allo-HSCT mouse model was established as previously described, with slight modifications [31]. Briefly, recipient BALB/c mice received potentially lethal total body irradiation of 7.5 Gy with an X-ray source (Hitachi); then, 6 × 106 donor C57BL/6 mice bone marrow (BM) cells, together with 3 × 106 splenocytes, were injected through the tail vein within 4–6 h after irradiation to reconstruct the hematopoietic system and induce aGVHD. To investigate the prophylactic effects of hUC-MSC-EVs on aGVHD, 200 μg of hUC-MSC-EVs, suspended in 200 μL of PBS, was injected on days 0 and 7 after transplantation. Survival and in vivo manifestations of aGVHD were monitored and assessed using a previously described scoring system [32].
Mitogen-induced proliferation assay
Mouse splenocytes were prepared and assayed for proliferation as previously described, with slight modifications [31,33]. Briefly, 2 × 105 splenocytes were cultured in 100 μL of RPMI 1640 medium (Gibco, Life Technologies) supplemented with 20% fetal bovine serum (Gibco, Life Technologies) at 37°C in a 5% CO2 atmosphere; concanavalin A (ConA; Santa Cruz Biotechnology) was added at a final concentration of 10 μg/mL to stimulate proliferation. Increasing concentrations of hUC-MSC-EVs or hUC-MSCs were added at the beginning of culture. After incubation for 5 days, cell proliferation was determined using a CCK-8 assay kit (Dojindo) according to the manufacturer's instructions.
T cell subtype analysis
Peripheral blood was collected from recipient mice on days 7 and 14 after allo-HSCT. T cell subpopulations, including CD3+CD4+ and CD3+CD8+ T cells, were stained using the corresponding fluorescently labeled antibodies (eBioscience). Stained cells were detected using a flow cytometer (Becton Dickinson), and data were analyzed using FlowJo Software (Tree Star; version 7.6.1).
Cytokine measurements
Culture supernatants were harvested after 5 days of incubating mitogen-activated splenocytes in the absence or presence of increasing concentrations of hUC-MSC-EVs. Serum samples were collected from recipient BALB/c mice on days 7 and 14 after allo-HSCT. The concentrations of cytokines, including tumor necrosis factor (TNF)-α, interferon (IFN)-γ, interleukin (IL)-2, and IL-10, were detected using the ProcartaPlex™ Multiplex Immunoassay (eBioscience) according to the manufacturer's instructions.
H&E staining
Mice were euthanized 30 days after allo-HSCT. The skin, liver, and large intestine were fixed in 10% phosphate-buffered formalin, dehydrated, paraffin embedded, sectioned, stained with H&E, and observed by light microscopy.
Statistical analysis
Data are presented as the mean ± standard deviation (SD) after analysis using SPSS software (SPSS, Inc., version 17.0). Student's t-test and analysis of variance (ANOVA) were used for comparisons. The log-rank test was used to analyze differences in survival. P values less than 0.05 were considered statistically significant.
Results
The characteristics of hUC-MSC-EVs
MSC-derived exosomes and microvesicles, which both act as effector mediators in MSC-based therapy, can be obtained through sequential ultracentrifugation at 20,000 g and 100,000 g, respectively [34]. In this study, to maximize the beneficial effects of MSC-EVs, we did not separate and distinguish between these two vesicle types. Also, hUC-MSC-EVs were prepared using a special ultracentrifugation step, as mentioned in the Materials and Methods section, and the resulting population included both exosomes and microvesicles. Accordingly, hUC-MSC-EVs appeared as a group of heterogeneous spheroids by transmission electron microscopy, with the size ranging from 30 to 100 nm. The flow cytometric analysis showed that hUC-MSC-EVs were positive for CD9, CD63, and CD81, which are transmembrane proteins that are enriched in EVs [35] (Fig. 1).
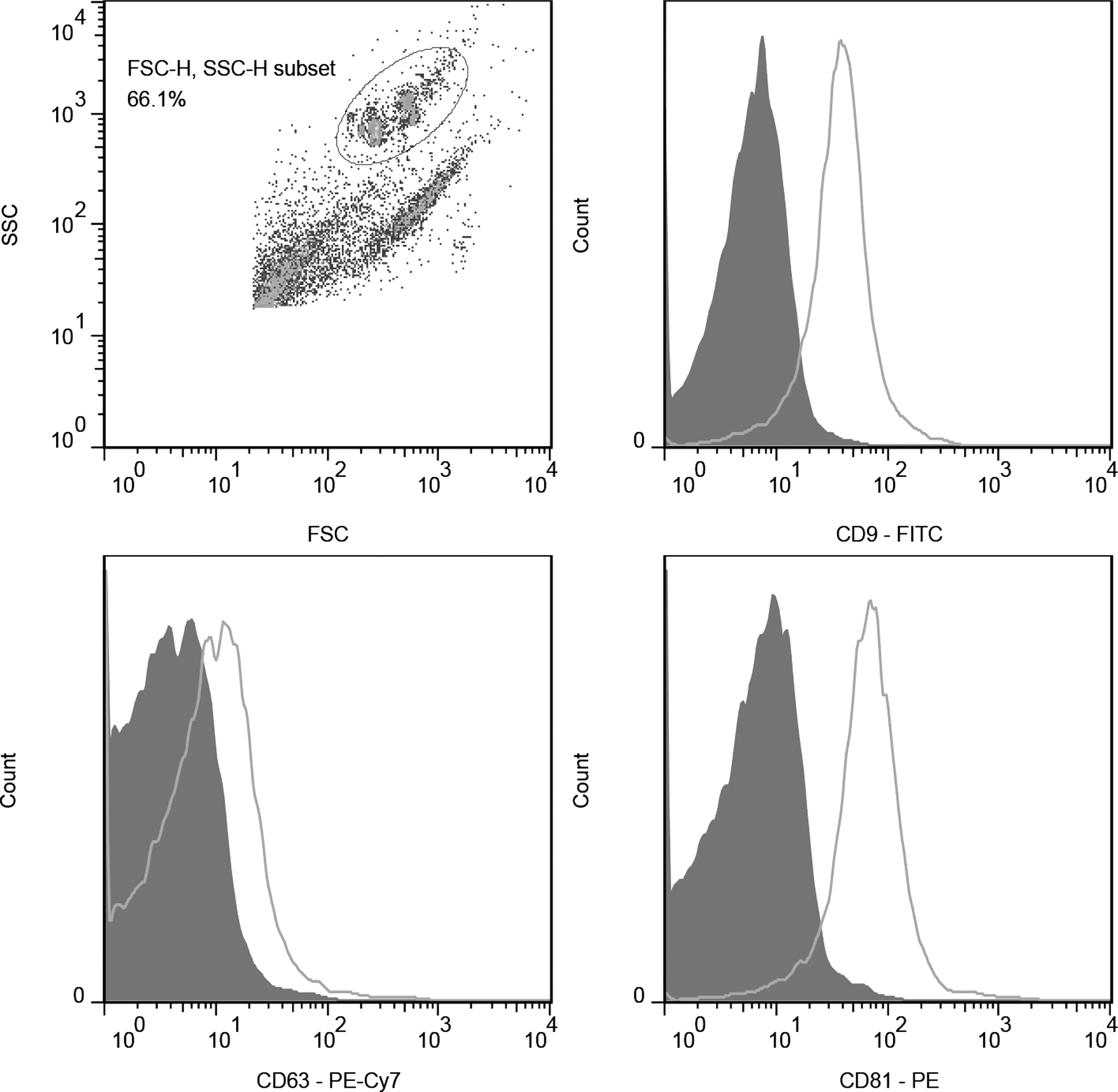
Flow cytometric analysis of hUC-MSC-EV surface markers. hUC-MSC-EVs were positive for CD9, CD63, and CD81. The EV-coated beads were immunostained with anti-CD9, anti-CD63, and anti-CD81 antibodies (open curves) or corresponding isotype controls (filled curves). hUC-MSC-EVs, extracellular vesicles released from human umbilical cord-derived mesenchymal stromal cell.
hUC-MSC-EVs attenuated the in vivo manifestations of aGVHD and the associated histologic changes and prolonged the survival of mice that underwent allo-HSCT
We first explored whether hUC-MSC-EVs had prophylactic effects on aGVHD in a mouse model of allo-HSCT. BALB/c mice that received BM cells and splenocytes showed in vivo manifestations of severe aGVHD approximately 28 days after allo-HSCT (Fig. 2A–C), and the mice died more rapidly: all died from severe aGVHD approximately 2 months after allo-HSCT (Fig. 2D). The skin, gastrointestinal tract, and liver are the main target organs of aGVHD, and, consistent with the obvious in vivo symptoms, the histology of these organs changed profoundly (Fig. 2E). In contrast, mice that received hUC-MSC-EVs exhibited mild in vivo manifestations of aGVHD (Fig. 2A, B), and aGVHD in vivo scores in the hUC-MSC-EV group were significantly lower than those in the GVHD group (P < 0.001, Fig. 2C). Accordingly, hUC-MSC-EV prophylaxis significantly reduced the mortality related to aGVHD (P < 0.01, Fig. 2D) and was associated with fewer histologic changes (Fig. 2E).
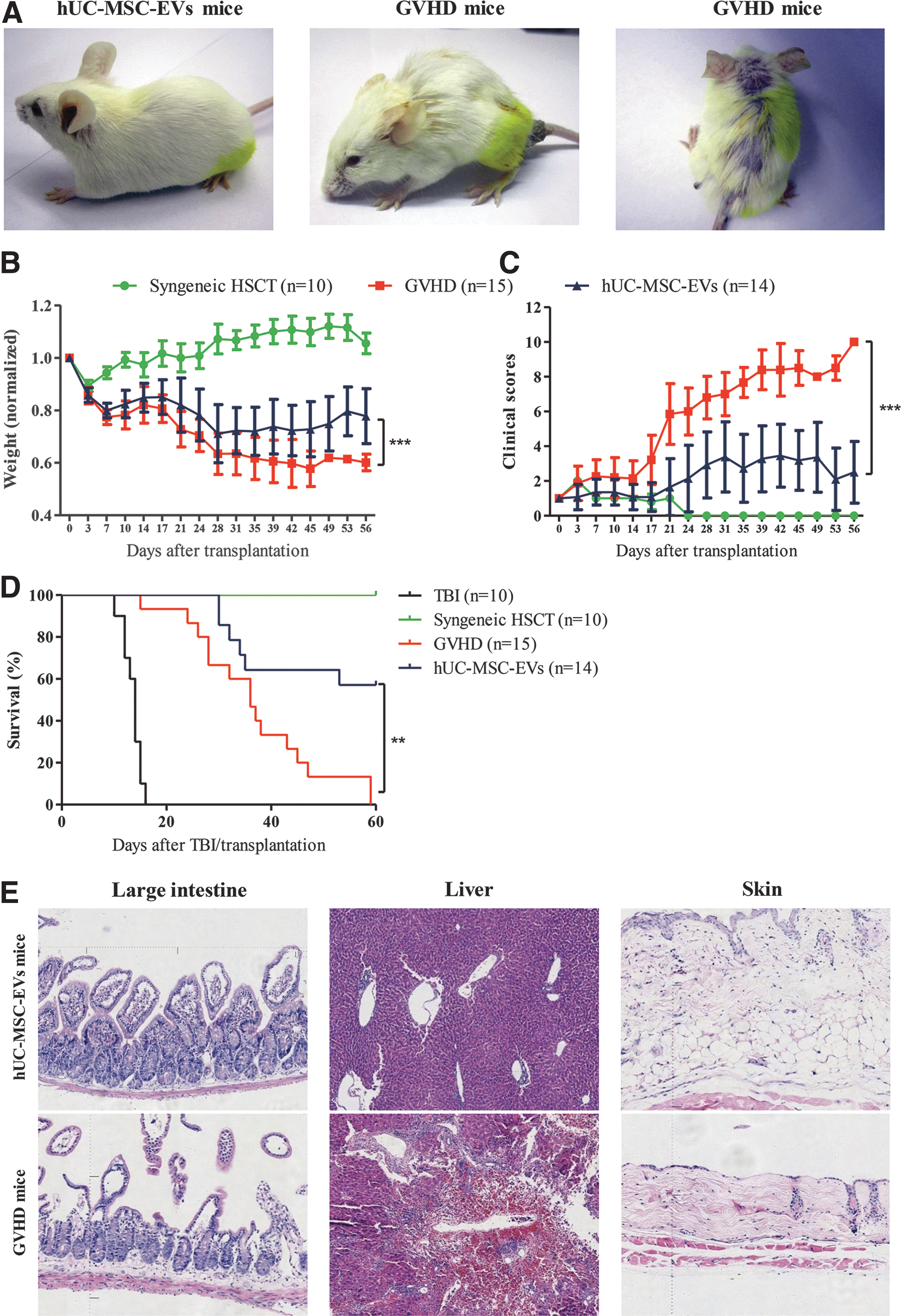
hUC-MSC-EVs prevented life-threatening aGVHD.
hUC-MSC-EVs decreased the frequency and absolute number of CD3+CD8+ T cells and increased the CD3+CD4+/CD3+CD8+ T cell ratio
Donor T lymphocytes are the predominant effector cells that initiate aGVHD. We next explored whether the beneficial effects of hUC-MSC-EVs were due to their impact on immune cells, specifically, CD3+CD8+ and CD3+CD4+ T cells. Consistent with the mild in vivo manifestations of aGVHD, hUC-MSC-EV administration significantly decreased the frequency and absolute number of CD3+CD8+ T cells on days 7 and 14 after allo-HSCT (P < 0.05, Fig. 3A). Although there were decreasing trends in terms of CD3+CD4+ T cells, there were no significant differences between GVHD and hUC-MSC-EV mice (P > 0.05, Fig. 3B). However, the ratio of CD3+CD4+ to CD3+CD8+ T cells was significantly higher in the hUC-MSC-EV group than in the control group (P < 0.05, Fig. 3C). Thus, we hypothesized that the beneficial effects of hUC-MSC-EVs may be due to their inhibitory effects on immune cells, specifically, CD3+CD8+ T cells.
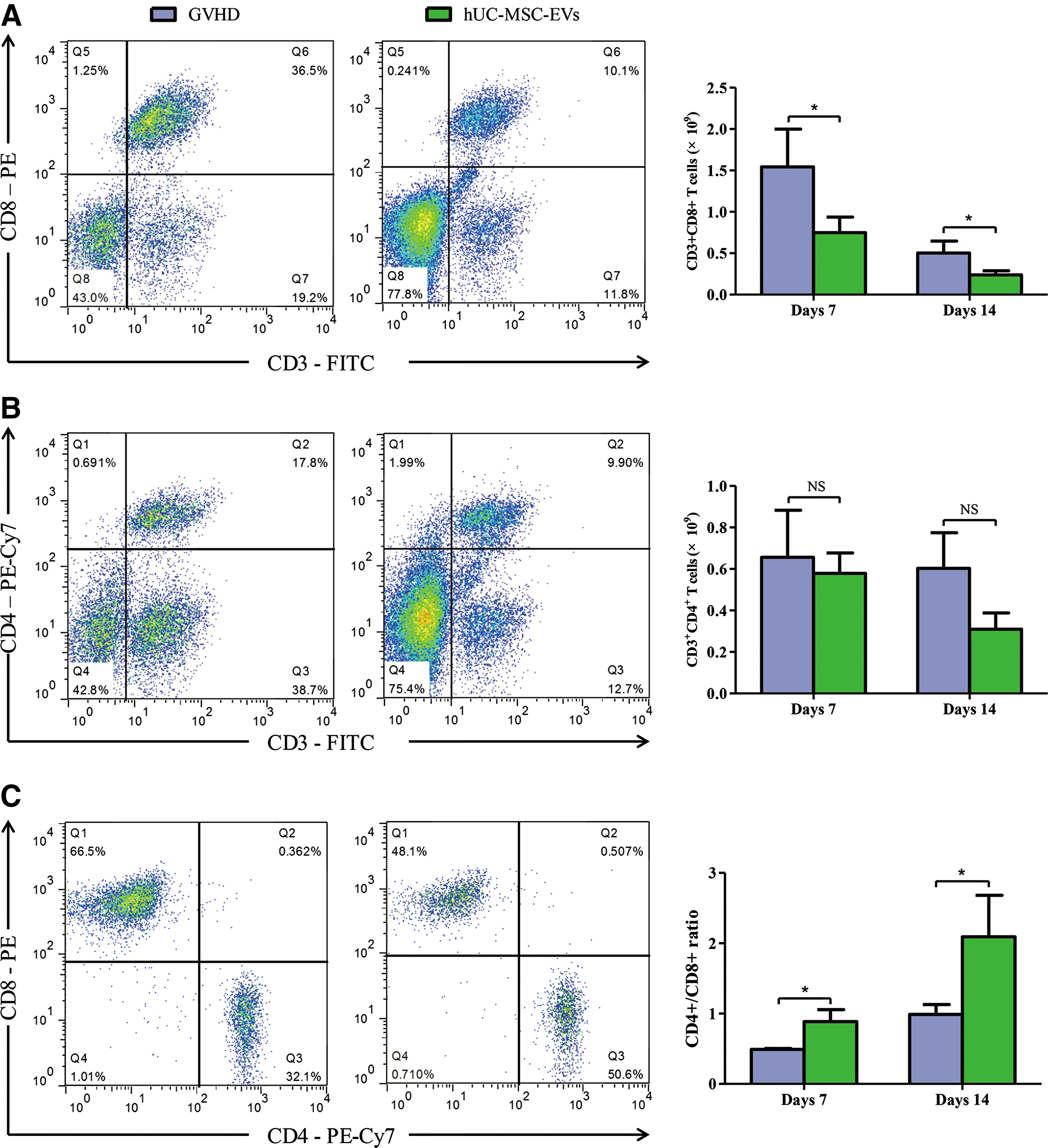
hUC-MSC-EVs suppressed T cells and altered T cell subtypes in recipient mice.
hUC-MSC-EVs exhibited immunosuppressive effects in vitro
To confirm the in vivo results, we examined the immunosuppressive effects of hUC-MSC-EVs using mitogen-induced proliferation assays. Using ConA-stimulated splenocytes as a positive control, hUC-MSC-EVs at concentrations ranging from 25 to 200 μg/mL significantly inhibited ConA-induced proliferation in a dose-dependent manner (P < 0.05, Fig. 4). Although low concentrations (5 and 10 μg/mL) of hUC-MSC-EVs did not significantly inhibit proliferation, there were decreasing trends (P > 0.05, Fig. 4). hUC-MSC-EVs at a concentration of 200 μg/mL exhibited similar inhibitory effects as hUC-MSCs at a ratio of 1:1 to splenocytes (P > 0.05, Fig. 4).

The inhibitory effects of hUC-MSC-EVs on ConA-stimulated splenocytes. Splenocytes cultured without ConA were used as a negative control, and ConA-activated splenocytes served as a positive control. hUC-MSC-EVs and hUC-MSCs inhibited ConA-induced proliferation in a dose-dependent manner. Data are presented as the mean ± SD; the experiments were performed in triplicate, and a representative experiment is shown. **P < 0.01. The hUC-MSC-EV protein concentration was quantified by the BCA method to indicate the concentration of EVs; 250 mL of hUC-MSC culture supernatant produced approximately 1 mg of EVs. BCA, bicinchoninic acid; SPC, splenocytes.
hUC-MSC-EVs altered the release of inflammatory cytokines in vitro and in vivo
The activation and proliferation of allo-reactive donor T lymphocytes and the subsequent release of inflammatory cytokines such as TNF-α, IFN-γ, IL-2, and IL-10 play a pivotal role in the pathophysiology of aGVHD [1,3]. We therefore investigated whether hUC-MSC-EVs influence the release of the aforementioned cytokines. Cytokine levels in the culture supernatants of ConA-activated splenocytes indicated that hUC-MSC-EVs decreased the levels of proinflammatory cytokines, for example, IL-2, TNF-α, and IFN-γ, in a dose-dependent manner (Fig. 5A–C). Levels of the anti-inflammatory cytokine IL-10, which plays an important role in suppressing immune responses, were significantly increased in response to hUC-MSC-EVs (Fig. 5D). Accordingly, hUC-MSC-EVs significantly reduced the levels of IL-2, TNF-α, and IFN-γ in recipient mice (Fig. 5E–G) and were associated with higher levels of IL-10 (Fig. 5H). Together, these in vitro and in vivo data indicated that hUC-MSC-EVs contributed to better outcomes by altering the release of inflammatory cytokines.
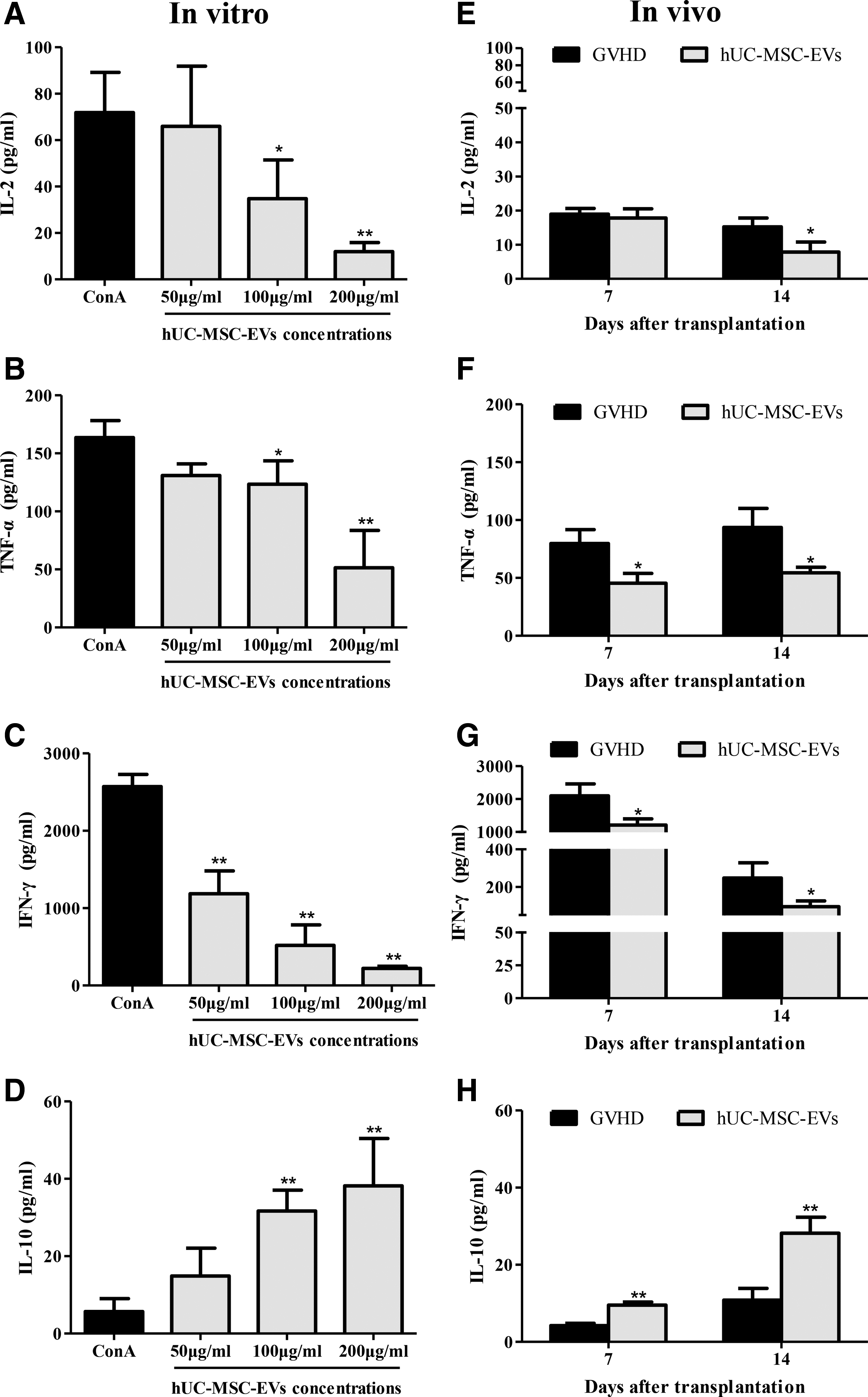
hUC-MSC-EVs altered the release of inflammatory cytokines in vitro and in vivo.
Discussion
To date, MSCs have generated great interest in the field of aGVHD management [2,5 –10]. However, MSCs failed to significantly increase durable complete remission rates in steroid-refractory aGVHD patients in a phase III, multicenter, randomized controlled trial [36]. Several factors, including MSC sources and manufacturing process, may account for the conflicting results [37]. The UC may be an ideal alternative to adult-derived MSC sources such as bone marrow and adipose tissue [38]. First, the UC can be easily, painlessly, and safely collected from discarded tissues following birth. Second, UC-MSCs can be expanded more rapidly in vitro compared with BM-MSCs [39]. Last, UC-MSCs have similar or even superior immunomodulatory properties to MSCs derived from other sources [38,39]. In this study, we investigated the prophylactic effects of hUC-MSC-EVs on aGVHD in a mouse model of allo-HSCT. Our study showed that hUC-MSC-EVs clearly attenuated the in vivo manifestations of aGVHD and the associated histologic changes and significantly prolonged the survival of recipient mice.
A previous study demonstrated that MSC exosomes inhibited mitogen-activated splenocyte proliferation only at relatively high concentrations, for example, 4 μg/mL [40]. In our study, hUC-MSC-EVs exerted obvious inhibitory effects at concentrations ranging from 5 to 200 μg/mL. In line with the in vitro results, hUC-MSC-EVs significantly decreased the frequency and absolute number of CD3+CD8+ T cells in recipient mice. This finding was supported by a previous study, in which MSCs reduced the frequency and absolute number of donor T cells through an exosome-linked adenosine signaling pathway in a xenogeneic GVHD model [41]. However, the ratio of CD3+CD4+ to CD3+CD8+ T cells significantly increased in the hUC-MSC-EV group, which could be partly explained by the fact that MSC exosomes mediate the conversion of T helper type 1 into T helper type 2 cells [42] and the differentiation of T cells into CD4+CD25+FoxP3+ regulatory T cells, although contrary results exist [40,41].
Moreover, our in vitro and in vivo experiments demonstrated that hUC-MSC-EVs significantly decreased the levels of several pro-inflammatory cytokines, including IL-2, TNF-α, and IFN-γ, and increased the levels of anti-inflammatory cytokines, for example, IL-10. These data are supported by previous studies in which MSC exosomes increased IL-10 levels and decreased TNF-α levels in vitro and in a renal ischemia-reperfusion injury rat model, respectively [40,43]. Apart from the aforementioned direct evidence, previous studies have demonstrated that the number of peripheral blood mononuclear cells and natural killer cells that release IFN-γ and TNF-α decreased after MSC exosome treatment [25] and TNF-α- and IFN-γ-producing T cells were reduced by an exosome-involved mechanism [41].
Notably, Conforti et al. showed that BM-MSC-derived microvesicles were inferior to their cellular counterparts in terms of their ability to inhibit mitogen-induced T cell proliferation [44]. This effect is likely due to the involvement of both intercellular contact-dependent and contact-independent processes in MSC-based therapy, both of which are important for the full immunoregulatory effects of MSCs [38]. Moreover, a previous study demonstrated that EVs released from both bone marrow- and adipose tissue-derived MSCs did not exhibit inhibitory effects on CD3-/CD28-stimulated peripheral blood mononuclear cells [45], indicating that the underlying mechanisms of MSC-EVs require further investigation. However, the greater inhibitory effects of hUC-MSC-EVs and the disparate cytokine profiles in this study may be attributable to the following reasons: first, MSCs derived from different tissue sources may exhibit disparate immunomodulatory properties. UC-MSCs may possess a higher immunomodulatory capacity than BM-MSCs [46]. Second, the different ultracentrifugation protocol used in this study resulted in the inclusion of both exosomes and microvesicles released from hUC-MSCs in the final pellet, which may have contributed to the greater immunomodulatory effects. Compared with MSCs, MSC-EVs have several advantages. First, the use of cell-free EVs readily overcomes the aforementioned safety concerns related to MSCs. Second, EVs selectively enrich protein, lipids, and genetic material, including mRNA and microRNA, from their parental cells and shuttle them to specific target cells [27]; meanwhile, their phospholipid bilayer may protect their content from degradation by circulating enzymes [47], which enables a more efficient mode of signal transmission. Finally, EVs are easily prepared and can be obtained using large-scale production protocols, such as has been reported for immortalized MSCs [17,47,48]. In summary, MSC-EVs are an attractive alternative to MSCs.
This study involving EVs was conducted according to the recommendations from the International Society for EVs [35]. However, there are several limitations to our study. First, only a BCA assay, which is an indirect method, was used to quantify the concentration of EVs; the use of nanoparticle tracking analysis to directly determine the EV concentration may increase the accuracy of the quantitative results. Second, the fetal bovine serum used in the mitogen-induced proliferation assay was not EV depleted, which might somewhat influence the measurements [35]. However, the experimental conditions were identical apart from the absence or presence of hUC-MSC-EVs. Therefore, we concluded that the observed immunosuppressive effects were attributed to hUC-MSC-EVs. Last, we did not investigate the molecular content of hUC-MSC-EVs. Further investigations of the content of EVs may improve the understanding of the underlying mechanisms of EVs [35].
Taken together, our study demonstrated that hUC-MSC-EVs attenuated life-threatening aGVHD after allo-HSCT mainly by modulating immune responses, including suppressing the proliferation of allo-reactive T cells, altering the proportion of T cell subtypes, inhibiting the release of several pro-inflammatory cytokines, and promoting the release of anti-inflammatory cytokines. Given the encouraging results and the superiority of MSC-EVs, hUC-MSC-EVs may represent an attractive alternative in the prophylaxis of aGVHD after allo-HSCT.
Footnotes
Acknowledgments
We are grateful to Prof. Lisheng Wang from the Department of Experimental Hematology, Beijing Institute of Radiation Medicine, China, for kindly providing hUC-MSCs. This study was supported by the National Natural Science Foundation of China (81270642; 81070451), the Beijing Natural Science Foundation (7162175), and the Hainan Social Development Special Funding (SF201306). The funders had no role in the study design; the collection, analysis, and interpretation of data; or the preparation and submission of the article.
Author Disclosure Statement
No competing financial interests exist.