
Abstract
Pluripotent stem cells have the potential to differentiate into all cell types of the body in vitro through embryoid body formation or in vivo through teratoma formation. In this study, we attempted to generate in vivo neural stem cells (NSCs) differentiated through teratoma formation using Olig2-GFP transgenic embryonic stem cells (ESCs). After 4 to 6 weeks of injection with Olig2-GFP transgenic ESCs, Olig2-GFP+ NSCs were identified in teratomas formed in immunodeficient mice. Interestingly, 4-week-old teratomas contained higher percentage of Olig2-GFP+ cells (∼11%) than 6-week-old teratomas (∼3%). These in vivo-derived NSCs expressed common NSC markers (Nestin and Sox2) and differentiated into terminal neuronal and glial lineages. These results suggest that pure NSC populations exhibiting properties similar to those of brain-derived NSCs can be established through teratoma formation.
Introduction
P
One method for testing cell pluripotency is teratoma formation; pluripotent cells should form teratomas containing various cell types, including endodermal, ectodermal, and mesodermal cells. Therefore, teratomas could provide an in vivo PSC differentiation environment. In line with this, recent reports have shown that fully functional and engraftable hematopoietic stem cells could be obtained from induced PSCs (iPSCs) through teratoma formation [14], and in vivo hematopoietic stem cells could be differentiated into functional B- and T-cell lineages [15]. These results suggest that an in vivo environment, even non-niche specific enables pluripotent cells to recapitulate in vivo development into specific cell lineages.
In this study, we developed an in vivo neural stem cell (NSC) differentiation system through teratoma formation using pluripotent embryonic stem cells (ESCs). These in vivo-derived NSCs were similar to brain-derived NSCs in both gene expression profiles and the potential to differentiate into neurons and glial cells. Based on these results, we suggest that teratomas or an in vivo environment, although not niche specific, endow pluripotent cells with the ability to recapitulate the required NSC niche.
Materials and Methods
The methods were carried out in accordance with the approved guidelines, and all experimental protocols were approved by the Institutional Animal Care and Use Committee of Konkuk University.
Cell culture
Olig2-GFP transgenic ESCs (Olig2 ESCs) containing the GFP transgene under the control of the Olig2 promoter [16] were used for our experiments. The GFP-neo cassette was integrated into the Olig2 gene translational start site. Olig2 ESCs were grown on mitomycin C-treated mouse embryo fibroblast (MEF) feeders with standard mouse ESC culture medium: Dulbecco's modified Eagle's medium (DMEM; Gibco) supplemented with 15% fetal bovine serum (FBS; Gibco), 1× penicillin/streptomycin/glutamine, 1 mM nonessential amino acids (NEAA; Gibco), 0.1 mM β-mercaptoethanol (Gibco), and 1,000 U/mL leukemia inhibitory factor (ESGRO; Chemicon).
Directed in vitro differentiation into NSCs
Directed differentiation of Olig2 ESCs into NSCs was carried out using a previously described method [17]. Olig2 ESCs were cultured for 2–3 days with MEF medium supplemented with 15% FBS (Gibco), 1× penicillin/streptomycin/glutamine (Gibco), 0.1 mM NEAA (Gibco), and 1 mM β-mercaptoethanol (Gibco) in DMEM (Gibco). Subsequent to 2–3 days culturing, the ESCs were cultured for 2 days without feeder cells in suspension culture dishes containing N2B27 medium (1:1 mixture of DMEM/F-12 medium; Gibco, Neurobasal medium (Invitrogen) supplemented with 10 ng/mL epidermal growth factor (EGF; Invitrogen), 10 ng/mL basic fibroblast growth factor (bFGF; Invitrogen), N2 supplement, B27 supplement (Gibco), and 1× penicillin/streptomycin/glutamine (Gibco). Then, they were plated onto 0.1% gelatin-coated dishes containing NSC expansion medium: DMEM/F12 (Gibco) supplemented with N2 supplement (Gibco), 10 ng/mL EGF (Invitrogen), 10 ng/mL bFGF (Invitrogen), 50 mg/mL bovine serum albumin (BSA, BSA fraction, Gibco), and 1× penicillin/streptomycin/glutamine (Gibco); this was followed by culture for 3 days.
Teratoma formation
Immunodeficient Balb/c Nude (5 weeks, male) mice were purchased from Orient Bio (Gyeonggi-do, Korea) to carry out experimental procedures. In this study, 1 × 106 cells Olig2 ESCs were injected into the testis capsule of immunodeficient mice. Teratomas were harvested surgically from mice at 4 or 6 weeks postinjection. Dissected teratomas were fixed in 4% paraformaldehyde (Sigma), processed through graded ethanol, and embedded in paraffin, followed by hematoxylin/eosin (Endoderm), PAS (Ectoderm), and Alcian blue (Mesoderm) staining. Immunohistochemistry staining of Nestin and Musashi was performed with an anti-Nestin and anti-Musashi antibody (Nestin, monoclonal, 1:100; Millipore), Musashi (monoclonal, 1:50; Millipore).
Flow cytometry analysis and sorting
Teratomas were enzymatically dissociated in 0.25% trypsin (Gibco) for 15 min at 37°C with mechanical dissociation by pipetting every 5 min. Dissociated cells were passed through a 70 μm nylon mesh (Falcon) to remove large cell clusters. The cells were then centrifuged at 1,500 rpm for 5 min and resuspended in phosphate-buffered saline (PBS; Gibco). Flow cytometry analysis and sorting were performed using a FACSAria (Becton Dickinson).
In vivo NSC derivation
FACS-sorted Olig2-GFP+ cells were plated on suspension dishes containing NSC culture medium [DMEM/F12 (Gibco) supplemented with N2 supplement (Gibco), B27 supplement (Gibco), 1× penicillin/streptomycin/glutamine (Gibco), 1M HEPES (Gibco), 45% Glucose (Sigma), 20 ng/mL EGF (Invitrogen), and 20 ng/mL bFGF (Invitrogen)] and then cultured for 2 to 3 weeks. Neurospheres were then replated on gelatin-coated tissue culture dishes containing NSC expansion medium and cultured for 2–3 days. Expanded neurospheres were dissociated into single cells by 0.25% trypsin (Gibco) treatment and passaged every 2–3 days in NSC expansion medium.
Differentiation of NSCs into neurons and glial cells
NSCs were differentiated into neurons and glial cells by culturing for 3 weeks in neuronal differentiation medium [1:1 mixture of DMEM/F12 medium (Gibco), Neurobasal medium (Gibco) supplemented with 1% FBS (Gibco), N2 supplement (Gibco), B27 supplement (Gibco), 1 × penicillin/streptomycin/glutamine (Gibco)].
Immunocytochemistry
NSCs were stained for markers of the NSCs: Nestin, Sox2, and Musashi. For immunocytochemistry, cells were fixed with 4% paraformaldehyde (Sigma) for 20 min at 4°C. After washing with PBS (Gibco), cells were treated with PBS containing 3% Bovine albumin serum and 0.03% Triton X-100 (Sigma) for 45 min at room temperature. Primary antibodies used were anti-Nestin (Nestin, monoclonal, 1:500; Millipore), anti-Sox2 (Sox2, polyclonal, 1:500; Millipore), and anti-Musashi-1 (Musashi-1, polyclonal, 1:500; Millipore). For detection of primary antibodies, fluorescence-labeled (Alexa fluor 488 or 568; Molecular Probes, Eugene, OR) secondary antibodies were used according to the specifications of the manufacturer.
RNA isolation and real-time quantitative reverse transcription-polymerase chain reaction analysis
Total RNA was isolated using the RNeasy Mini Kit (Qiagen, Venlo, Netherlands,
Statistical analysis
All experiments were performed in triplicate, and data represented as mean ± SD. Significance of differences was assessed by an unpaired t-test at P < 0.05.
Results
Generation of NSCs from ESCs in vitro
Olig2-GFP transgenic ESCs (Olig2 ESCs) containing the GFP transgene under the control of the Olig2 promoter [16] were used to generate NSCs. We first tested whether Olig2 ESCs are efficiently differentiated into NSCs in vitro with activation of the Olig2-GFP marker by examining neurosphere formation; Olig2-GFP was not active in the pluripotent state, but was activated when the ESCs differentiated and formed neurospheres from which monolayered NSCs were established (Fig. 1A). Immunocytochemistry analysis also showed that ESC-derived NSCs expressed NSC markers, including Nestin, Sox2, and Musashi (Fig. 1B). These results indicate that NSCs could be efficiently derived from Olig2 ESCs through the in vitro differentiation system, and Olig2-GFP marker could be a selection marker for NSCs differentiated from ESCs.
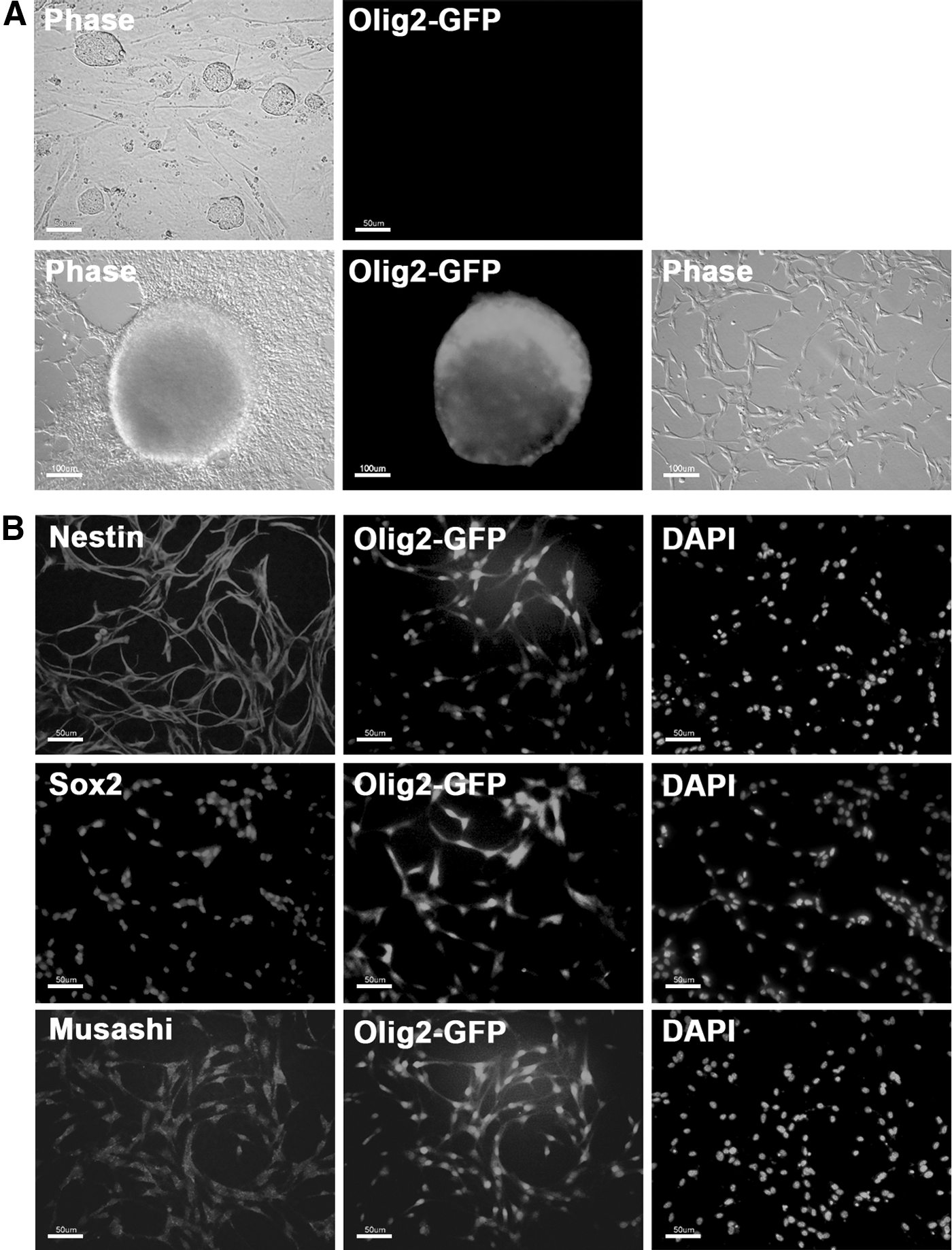
In vitro differentiation of Olig2 ESCs into NSCs.
Generation of NSCs from Olig2 ESCs in vivo
We next set out to generate NSCs from Olig2 ESCs through in vivo environment, or teratoma formation (Fig. 2A). Olig2 ESCs were injected into the testis capsule of BALB c/Nude or NOD/SCID mice, and the testes were harvested 4 to 6 weeks postinjection. Teratoma formation was observed in the testes of mice injected with Olig2 ESC (Fig. 2B). Teratoma sections were stained with tissue-specific antibodies, which showed that all three germ layer tissues were present in the teratoma (Fig. 2C). Staining of teratoma sections with the Nestin or Musashi antibody revealed numerous Nestin+ or Musashi+ cells in the teratomas (Fig. 2D), confirming the presence of NSCs in the teratoma. Next, the teratomas were dissociated into small clumps or single cells, in which many Olig2-GFP+ cells were observed (Fig. 3A). These observations suggest that the teratomas formed from Olig2 ESCs contain a high percentage of Nestin+ and Olig2+ cells.
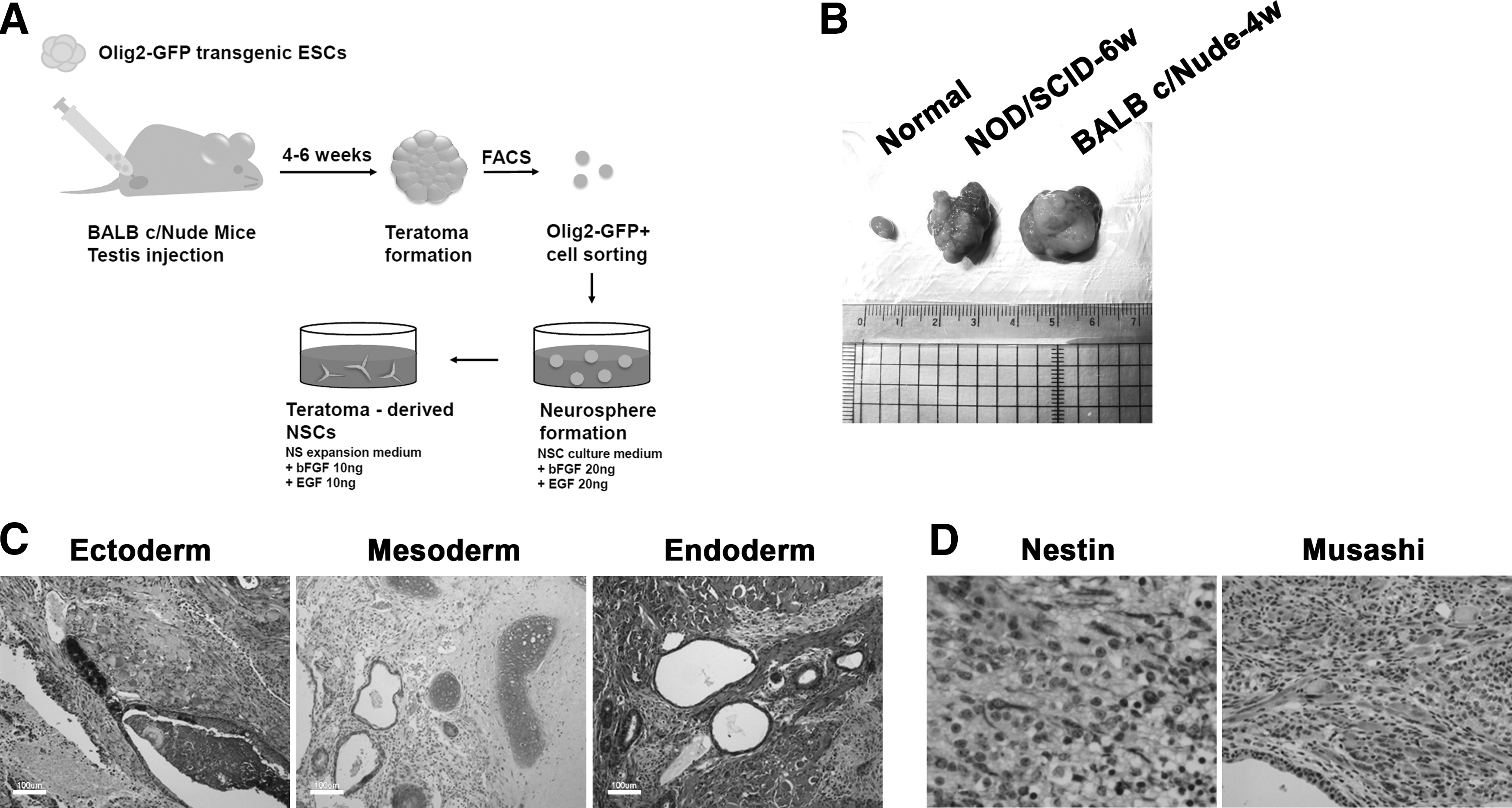
In vivo differentiation of Olig2 ESCs through teratoma formation.
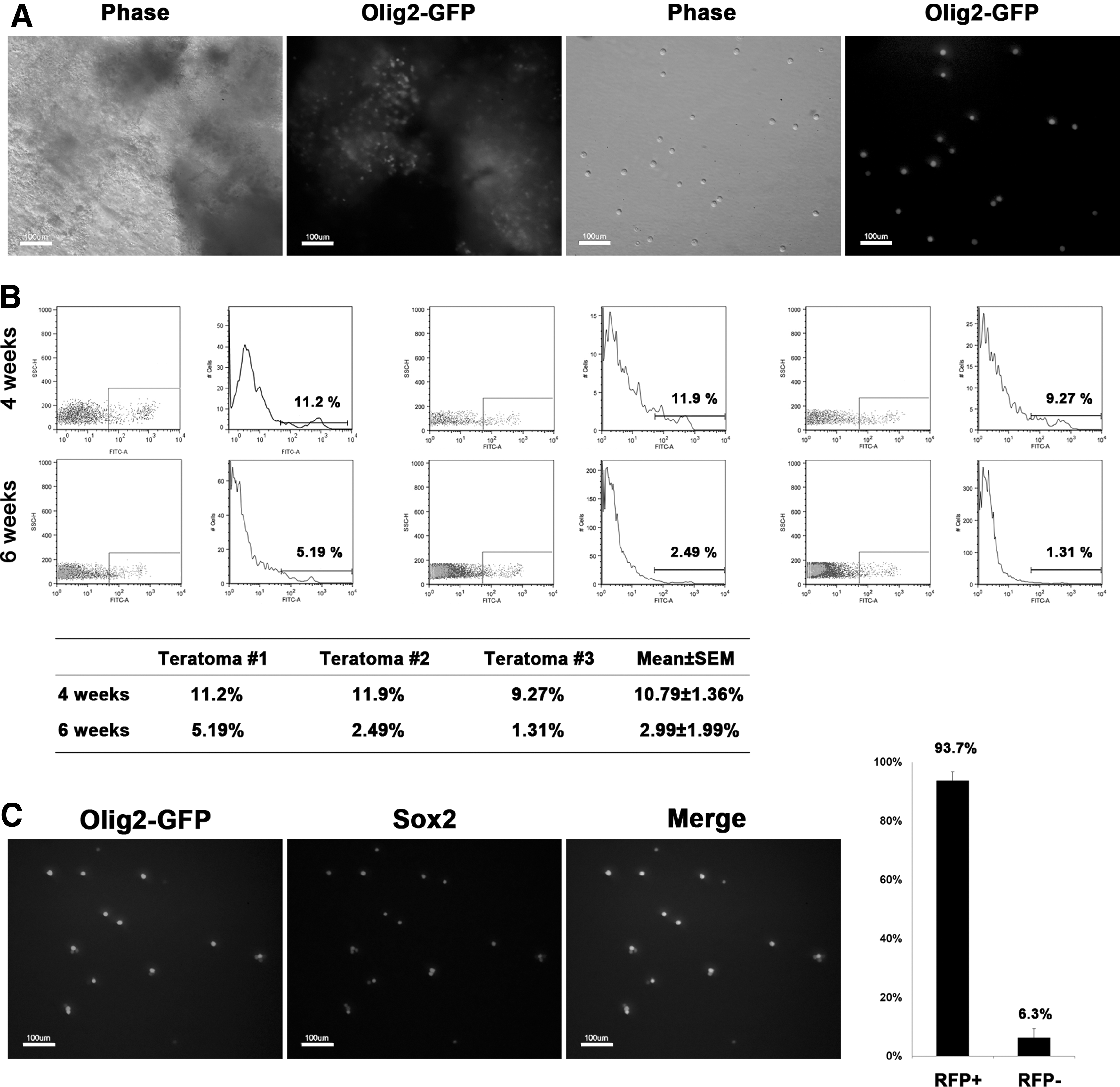
Flow cytometry analysis of teratomas.
Next, Single cells dissociated from teratomas were FACS sorted for Olig2-GFP+ cells. Since the proportion of Olig2-GFP+ cells is different in each teratoma sample, separate cells from each teratoma were independently analyzed by FACS. Interestingly, the proportion of Olig2-GFP+ cells was much higher in 4-week-old teratomas (∼11%) than in 6-week-old teratomas (∼3%; Fig. 3B). This result could be explained by the fact that neural lineage specification is the one of the earliest events during embryo development [19]. Thus, neural lineage might be formed at the earlier stage of differentiation in teratoma environment. Olig2-GFP+ cells from 4-week-old teratomas tended to form neurospheres more efficiently than those from 6-week-old teratomas (data not shown). Therefore, we attempted to obtain NSCs from earlier stage of teratoma, but we could not observe detectable teratomas at 3 weeks after ESC injection. These results indicate that 4-week-old teratomas provide the best option for the derivation of in vivo NSCs through teratomas. To confirm whether the GFP+ cells are NSCs, we stained the GFP+ cells with an NSC marker, Sox2. Nearly all the Olig2-GFP+ cells were also positive for Sox2 (Fig. 3C), indicating that the sorted Olig2-GFP+ cells are mostly a pure population of NSCs.
Sorted GFP-positive cells were subjected to neurosphere formation in EGF and bFGF-containing medium. The Olig2-GFP+ cells began to form small clumps subsequent to one day of culturing and became large neurospheres at day 5 of culturing. In addition, the Olig2-GFP+ neurospheres stained positive for the NSC marker, Sox2 (Fig. 4A). These neurospheres were plated onto gelatin-coated dishes on which adherent bipolar NSCs were propagated (Fig. 4B). These adherent NSCs (in vivo-t-NSCs) expressed the common NSC markers, Nestin, Sox2, and Musashi (Fig. 4C). The NSCs remained Olig2-GFP+ for over 20 passages and their proliferation rate was similar to brain-derived NSCs, which is derived from normal brain tissue of 13.5 embryos (Fig. 4D). Furthermore, adherent NSCs (in vivo-tNSCs) also form neurospheres on nonadherent conditions (Supplementary Fig. S1A; Supplementary Data are available online at
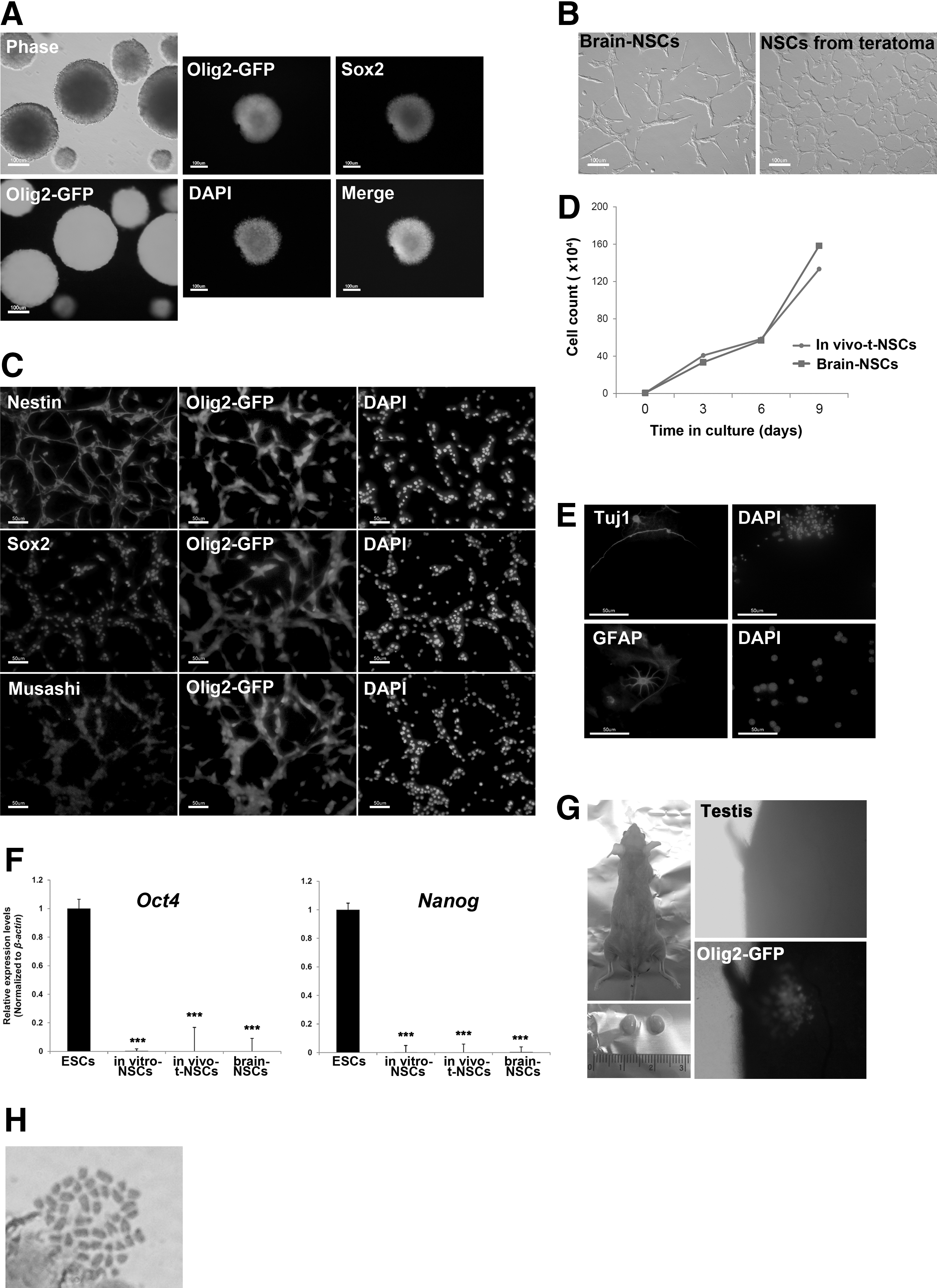
In vivo generation of NSCs by teratoma formation.
Discussion
In this study, we demonstrated that NSCs could be directly derived from teratomas formed subsequent to the injection of ESCs into immunocompromised mice. These results suggest that teratomas could provide a permissive niche for neurogenesis from PSCs. In addition, we found that in vivo NSCs derived from teratomas retain a normal karyotype, suggesting that although teratomas constitute a type of tumor, chromosomal anomalies do not occur in the NSCs grown in teratoma environment.
Directed in vitro differentiation into specialized cell types used for naive pluripotent ESCs is applicable to iPSCs and primed PSCs only by using a modified protocol. For example, different stem cell types require different protocols for NSC derivation. EpiSCs at early passages can be differentiated into NSCs using a medium with a high concentration of bFGF [20]. In vitro differentiation of iPSCs into NSCs is a more time-consuming process compared with ESCs; subsequent to the induction of neural differentiation in vitro, neural tube-like rosettes formed at day 10 from ESCs but at day 30 from iPSCs [17]. In particular, efficiency of NSC derivation was as low as 1% in the in vitro system, but was 11% in the in vivo system (using teratomas harvested 4 weeks post-ESC injection). Thus, in vivo differentiation protocol has an advantage over the in vitro system, at least when it comes to the efficiency of NSC derivation from PSCs.
Teratomas are benign tumors formed by injection of PSCs into immunodeficient mice [14,15,21]. Since teratomas contain many different types of cells derived from all three germ layers, teratoma formation is one of the most stringent assays for confirming the pluripotency of naive and primed PSCs. An in vivo differentiation method was first reported for the generation of hematopoietic lineage cells; Amabile et al. reported the derivation of in vivo human hematopoietic cells from iPSCs through teratomas [14,15,21]. These hematopoietic cells could colonize murine hematopoietic tissues and differentiate into functional B- and T-cell lineages. Recently, Suzuki et al. also derived fully functional and engraftable hematopoietic stem cells from iPSCs through teratoma formation [14]. Although the stem cell lines were not established, Choi et al. initially found that a hepatocyte population of varying maturity was present in the teratomas subsequent to the injection of ESCs [21]. These findings suggest that many other somatic stem cell types in addition to NSCs and hematopoietic stem cells that could also be isolated and established as cell lines may be present in the teratomas. Therefore, the in vivo differentiation approach could contribute to the development of a differentiation protocol for other cell types, not established thus far using in vitro systems.
One of the most interesting findings of the present study is that teratomas harvested 4 weeks post-ESC injection contain approximately a four-fold greater population of Olig2-GFP+ cells than those harvested 6 weeks post-ESC injection (Fig. 3A). It is possible that late-stage (6 week) Olig2-GFP+ cells are more differentiated than early-stage (4 week) NSCs. However, considering the property of NSCs, Olig2-GFP+ cells, which were established in the early stage, might be differentiated afterward and lose Olig2-GFP expression. Determining the exact timeframe of neural specification during teratoma development is essential, since harvesting a pure population of NSCs with high differentiation efficiency is critical for clinical application. The optimal time point for neural specification is as early as 4 weeks; ∼11% of all cells in these teratomas are Olig2+ cells. This is due the fact that neural specification is one of the earliest events to occur during embryonic development [19]. This timeframe may be also useful for deriving NSCs from cells lacking selection markers, such as Olig2-GFP cells. Differentiation using in vivo environments may guarantee efficient derivation of NSCs from teratomas formed 4 weeks postcell injection. The in vivo differentiation method could be combined with specific chemicals or with coinjection of supportive cells to obtain a higher ratio of NSCs in the teratoma. For example, coinjection of OP9 stromal cells with iPSCs enhances hematopoietic specification during teratoma development [15]. This approach requires further improvement for the derivation of human PSCs such as nuclear transfer ESCs and human iPSCs. An efficient in vivo differentiation protocol following cellular reprogramming in combination with gene or epigenetic correction [22] could therefore provide a tool for drug screening and the desirable cell types for tissue replacement therapy for incurable diseases.
We developed an in vivo NSC differentiation system through teratoma formation using ESCs. These in vivo teratoma-derived NSCs were similar to brain-derived NSCs in both NSC marker expression patterns and the potential to differentiate into neurons and glial cells. Moreover, in vivo NSCs maintained normal karyotype and were not tumorigenic, although they were derived through tumor tissue, teratoma. Therefore, this in vivo differentiation system that mimics 3D in vivo environment will be a powerful method for generation of specific cell types.
Footnotes
Acknowledgments
This research was supported by the Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Science, ICT and Future Planning (grant no. 2015R1A2A2A01003604 and 2015R1A5A1009701) and the Next-Generation BioGreen 21 Program (grant no. PJ01133802) funded by the Rural Development Administration, Republic of Korea.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.