
Abstract
We reported previously testis-mediated germline chimera production and characterization of germline stem cell-like cells from chicken testes. The present study aimed to establish an in vitro system for culture of quail spermatogonial stem cells (SSCs) for practical applications in germline preservation and transgenesis. Testicular cells (TCs) from juvenile (4 weeks old) or adult (8 weeks old) quail testis were isolated using sequential enzymatic digestion. The percentages of viability of isolated TCs were 91.00% ± 2.12% and 88.00% ± 1.87% in juvenile and adult testes, respectively, and immunohistochemical evaluation indicated the expression of integrin alpha-6 (ITGA6), GDNF family receptor alpha-1 (GFRA1), and Deleted in azoospermia-like (DAZL) in specific TCs. SSCs were purified by differential plating of TCs and then subjected to quantitative reverse transcription-polymerase chain reaction, which showed differential expression of SSC-specific, and germness and stemness-related genes. Coculture of quail SSCs with mouse embryonic fibroblasts and Sertoli cells as a feeder layer resulted in the generation of stable SSC colonies and short-term cultivation, and the expression of SSC and germ cell markers was maintained during several passages of culture. Collectively, these results demonstrate the efficient isolation and characterization of quail SSCs and the suitability of Sertoli cells as a feeder layer for in vitro culture of quail SSCs. Quail SSCs will facilitate the production of germline chimeras and transgenesis.
Introduction
S
The survival and proliferation of SSCs in the testis microenvironment are firmly controlled and regulated by Sertoli cells [7,8]. In addition, glial cell line-derived neurotrophic factor (GDNF) and basic fibroblast growth factor (bFGF), which are produced by Sertoli cells, are essential growth factors for SSC self-renewal [9,10]. Thus, due to its potential for germline preservation and adult stem cell research, an in vitro culture system for SSC expansion in rodents [11 –14] and various domestic animals—including pig, buffalo, bovine, and goat [15 –18]—has been a focus of research.
In birds, both the expression levels of several molecular markers and studies involving transplantation have suggested conserved functions of germline stem cell-like cells from adult testes. SSCs isolated from adult chicken testis showed expression of chicken germ cell-specific and SSC-related markers, which was maintained during several passages in medium supplemented with leukemia inhibitory factor (LIF), bFGF, and insulin-like growth factor 1 (IGF1) [19]. Moreover, transplantation of chicken SSCs isolated using an anti-GDNF family receptor alpha-1 (GFRA1) antibody and xenogeneic transfer of quail SSCs into chicken recipient suggested that avian SSCs have the capacity to regenerate spermatogenesis [20,21], and so have potential for germline preservation and genome editing [22,23].
The germline transmission capacity of SSCs is largely conserved in avian species, and SSCs can facilitate preservation of endangered birds. However, studies of in vitro culture, molecular analysis, and germline chimera production using SSCs have, to date, focused on chicken species. In this study, we report a simple method for isolation and in vitro culture of Japanese quail (Coturnix japonica) SSCs, and identify molecular markers of quail SSCs during in vitro culture.
Materials and Methods
Experimental animals
Japanese quails (Coturnix japonica) were used as experimental animals. The Institute of Laboratory Animal Resources, Seoul National University (SNU-150827–1) approved the care and experimental use of quails. Quails were maintained according to the standard management program at the University Animal Farm, Seoul National University (Seoul, Korea). All procedures for animal management, reproduction, and embryo manipulation were governed by standard operating protocols.
Enzymatic isolation of testicular cells
Juvenile (4 weeks old) and adult (8 weeks old) testes were surgically removed and washed several times with phosphate-buffered saline (PBS) containing 1× antibiotic–antimycotic (Invitrogen). After removing the tunica albuginea and connective tissue, the testes were minced into small pieces on a Petri dish containing Dulbecco's minimum essential medium (DMEM; HyClone) containing 1× antibiotic–antimycotic. The minced pieces were sequentially treated with 1 mg/mL collagenase IV (Gibco) in DMEM for 30 min, 7 mg/mL DNase I in DMEM for 5 min, and 0.25% trypsin-EDTA (Invitrogen) in PBS for 20 min in a shaking water bath at 37°C [24,25].
The cell suspension resulting from each stage was washed by centrifugation (800 rpm, 5 min) with prewarmed (37°C) PBS. The final cell suspension was filtered through 100 and 40 μm nylon cell strainers (BD Falcon). The cell pellet was finally resuspended in DMEM containing 5% fetal bovine serum (FBS). The number and viability of isolated testicular cells (TCs) were determined by Trypan Blue staining.
Preparation of feeder layer and laminin coating
Quail SSCs were cultured with or without feeders. Sertoli cells and mouse embryonic fibroblasts (MEFs) were used as feeder cells. To prepare Sertoli cells, TCs from juvenile and adult quails in DMEM were added separately to Datura stramonium agglutinin (DSA-lectin, 10 μg/mL)-coated 12-well plates containing DMEM/F12 (Invitrogen) supplemented with 10% FBS, 10 ng/μL GDNF, 1× essential amino acids (EAA), 1× nonessential amino acids (NEAA), and 1× antibiotic–antimycotic solution, and incubated for 3 h at 37°C in a humidified atmosphere of 5% CO2 in air. After discarding nonadherent cells, the adherent Sertoli cells were cultured for 6–8 days. Upon reaching 90% confluency, the Sertoli cells were treated with 10 μg/mL mitomycin C (Sigma-Aldrich) for 3 h. The cells were washed three to four times with DMEM/F12, and Sertoli cells derived from juvenile testes were only used for further experiments as a feeder layer.
MEFs were isolated from ICR mouse embryos at 12.5–13.5 days postcoitum and cultured in DMEM supplemented with 10% FBS on 0.1% gelatin-coated (Sigma-Aldrich) dishes. MEFs at P3 were mitotically inactivated by treatment with mitomycin C for 3 h. For feeder-free culture of quail SSCs, laminin (20 μg/mL) in PBS was added to a 12-well plate and incubated overnight at 37°C. The plate was used after washing three times with PBS.
In vitro culture of SSCs and proliferation assay
The differential plating technique was used to enrich SSCs from TCs [26,27]. Briefly, after isolation, TCs from adult and juvenile quail testes were separately suspended in DMEM/F12 with 10% FBS and 1× antibiotic–antimycotic solution overnight in uncoated six-well plates at ∼3 × 106 cells to enrich SSCs. Then, suspended cells were collected, washed, and cocultured (1 × 105 per well) on laminin-coated plates, mitotically inactivated Sertoli cells, and MEFs in SSC medium at 37°C in a humidified CO2 incubator in 5% CO2. SSC medium consisted of DMEM/F12 supplemented with 1% FBS, 1× EAA, 1× NEAA, 15 ng/mL recombinant human GDNF (PeproTech), 10 ng/mL recombinant human bFGF (PeproTech), 10 ng/mL recombinant mouse LIF (Sigma-Aldrich), 0.55 mM β-mercaptoethanol (Gibco), 2 mM
The medium was replaced on alternate days and quail SSC clusters were passaged every 10–12 days by gently pipetting without disturbing the feeder cells. SSC colonies were counted and their diameters measured. During culture, a colorimetric assay was conducted to quantify proliferative activity of SSCs on Sertoli cell and MEF feeders. The enriched SSCs as single cell were seeded over mitomycin C-treated Sertoli cells and MEFs. Each well was treated with the cell proliferation reagent, WST-1, and cells were then incubated for 0, 0.5, 1, 2, 4, 8, and 12 h, respectively, in 5% CO2 at 37°C. After treatment, the absorbance (450–620 nm) was measured using a microplate reader.
Quail primordial germ cell isolation
Quail primordial germ cells (PGCs) were freshly isolated in accordance with the standard protocols of our laboratory [28]. Quail PGCs were retrieved from the gonads of embryos after 5 days of incubation, when the embryos had reached Hamburger and Hamilton stage 28. Embryos were freed from the yolk by rinsing with PBS and the gonads were then removed by medial section of the abdomen with sharp tweezers under a stereomicroscope. Gonadal tissue was dissociated by gentle pipetting in 0.05% (v/v) trypsin supplemented with 0.53 mM EDTA, and gonadal cells, with which gPGCs were included, were then retrieved by centrifugation at 1,250 rpm for 5 min. Finally, quail PGCs are purified by the magnetic activated cell sorting (MACS), performed according to the manufacturer's instructions (Miltenyi Biotec, Inc.) using a QCR1 antibody.
Quail stage X blastoderm isolation
For stage X blastodermal cell isolation, freshly laid eggshell was washed with 70% ethanol. Then the eggs were opened at the blunt end and the yolk was separated from white. The yolks were transferred onto sterile Petri dishes and cleaned from the remains of white. The stage X blastoderm was separated from the egg using sterilized paper and the shell membrane and albumen were detached from the yolk. A piece of square-type filter paper (Whatman, Maidstone, Kent, UK) with the hole at the center was placed over the germinal disc. After cutting around the paper containing the intrauterine embryo, it was gently turned over and transferred to saline buffer.
Quantitative RT-PCR
Quantitative reverse transcription-polymerase chain reaction (RT-PCR) was performed to assess the expression of markers of SSCs, germness, and stemness in TCs, enriched spermatogonial cells, stage X blastoderms, PGCs, and SSCs cultured in vitro. Total RNA samples were prepared using TRIzol reagent (Invitrogen). Total RNA (1 μg) was reverse transcribed using the Superscript III First-Strand Synthesis System (Invitrogen). The polymerase chain reaction (PCR) mixture was prepared by adding 2 μL PCR buffer, 1.6 μL 2.5 mM dNTP, 10 pmol each forward and reverse primer, 1 μL 20× Eva green, 0.2 μL Taq DNA polymerase, and 2 μL complementary DNA (cDNA) to a final volume of 20 μL. PCR was performed by means of an initial incubation at 94°C for 3 min, followed by 40 cycles at 94°C for 30 s, 60°C for 30 s, and 72°C for 30 s, using a melting curve program (increasing temperature from 55°C to 95°C at a rate of 0.5°C per 10 s) and continuous fluorescence measurement. Sequence-specific products were identified by generating a melting curve.
The Ct value represents the cycle number at which a fluorescent signal increases to a level significantly higher than the background, and gene expression was quantified by the 2−ΔΔCt method [29,30]. Quantitative PCR (qPCR) primers for each target gene, and quail β-actin (ACTB), were designed using the Primer3 software [31], and are listed in Table 1. Gene expression was normalized to that of quail ACTB. qPCR analysis of messenger RNAs (mRNAs) was performed using the StepOnePlus™ Real-Time PCR System (Applied Biosystems).
Immunohistochemistry
TCs, enriched SSCs, in vitro cultured SSCs at P4, and Sertoli cells at P2 were subjected to immunohistochemistry. Antibodies used are shown in Table 2. Cells were fixed in 1× PBS containing 4% paraformaldehyde on poly-
Finally, the cells were stained with a fluorescent nuclear stain, that is, 300 nM 4′6-diamidino-2-phenylindole (DAPI; Invitrogen) for 5 min, followed by rinsing several times with 1× PBS. Images were captured using a confocal microscope (Carl Zeiss) or fluorescence microscope (Leica Microsystems). Negative controls were performed by excluding primary antibodies.
Migration assay
Cultured quail SSCs and TCs were transplanted into the dorsal aortas of recipient 2 days old (Hamburger and Hamilton stage 14–16) quail embryos. Briefly, to transfer into the recipients, the cultured quail SSCs and TCs were agitated by gentle pipetting without trypsinization and harvested from the culture wells. Cultured quail SSCs and TCs were centrifuged at 1,250 rpm for 5 min and then resuspended by gently pipetting in a diluent solution containing PKH26 labeling dye, and the labeled cells were transferred into recipient embryos. A small window was cut into the pointed end of the recipient eggshell, and 2 μL of cell suspension containing ∼1,000 cultured SSCs and TCs were transplanted through this window using a micropipette. The eggshell window was sealed twice with Parafilm, and the egg was positioned with the pointed end down until hatching. Three days after transplantation, the gonads from a sample of the recipient embryos were collected, and the presence of PKH26-labeled quail SSCs and TCs in the gonads was monitored by fluorescence microscopy (Leica Microsystems).
Retinoic acid treatment for induction in quail SSCs
For in vitro induction in stable SSC colonies, 10 nM all-trans retinoic acid (RA; Sigma-Aldrich) was added to the culture medium for 0 and 24 h. Total RNA was extracted and analyzed by quantitative RT-PCR.
Statistical analysis
Differences between groups were analyzed statistically by one-way analysis of variance (ANOVA) or Student's t-test. A P value <0.05 was considered to indicate statistical significance.
Results
Isolation and characterization of quail TCs
In this study, we isolated quail TCs from juvenile (4 weeks old) and adult (8 weeks old) testes using a sequential enzymatic digestion method involving three enzymes—collagenase IV, DNase I, and trypsin. Trypan Blue staining of isolated TCs indicated ∼90% viability in both groups (Fig. 1A). Furthermore, differential plating was used to prevent contamination with Sertoli and myoid cells. To identify SSCs among the isolated quail TCs, immunostaining for Deleted in azoospermia-like (DAZL), integrin alpha-6 (ITGA6), and GDNF family receptor alpha-1 (GFRA1) was conducted. As shown in Fig. 1B, most of the isolated TCs, which were of various sizes, expressed the germ cell marker DAZL. A fraction of the isolated quail TCs expressed the SSC-specific markers ITGA6 and GFRA1 on their surface; these were round- to polygonal-shaped cells with a prominent nucleus and little cytoplasm. Furthermore, the GFRA1+ cells in quail TCs coexpressed with DAZL (Fig. 1B, bottom). Collectively, these results indicate that the ITGA6+ and GFRA1+ quail TCs may be an SSC subset of spermatogonial cells.
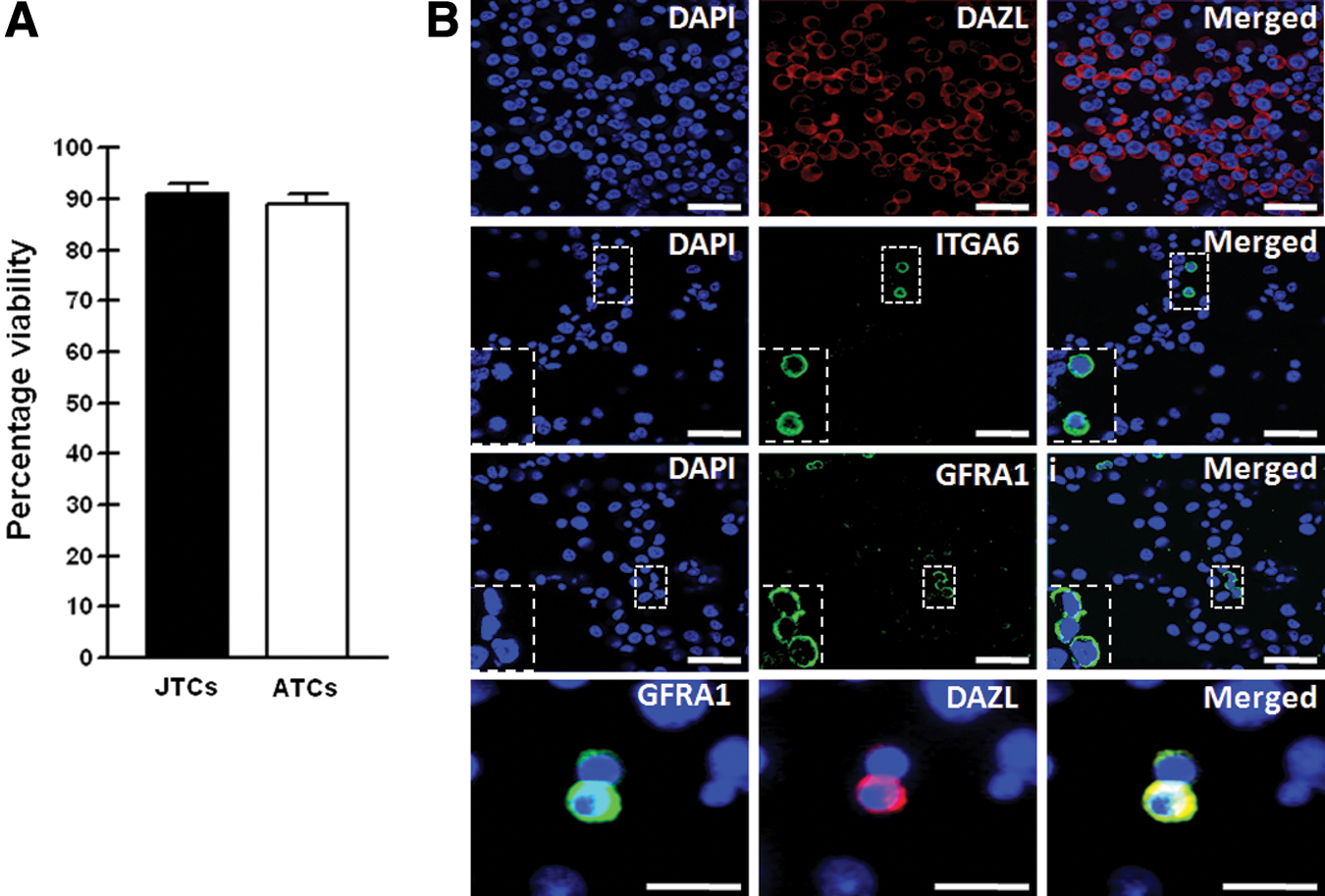
Characterization of isolated TCs from juvenile and adult quail testes.
Characterization of enriched spermatogonial cells
We purified spermatogonial cells from TCs by differential plating followed by attachment to the surface of laminin-coated plates. To examine the enriched population of spermatogonial cells, including SSCs (SPG), we analyzed the expression of genes specific to SSCs and those related to germness and stemness using freshly isolated quail PGC, together with stage X blastoderms (BLs) and TCs from juvenile quail testis as a control. As shown in Fig. 2, expression of the SSC-specific markers, GFRA1 (P < 0.001), ITGA6 (P < 0.001), and integrin β1 (ITGB1; P < 0.001) was significantly higher in spermatogonial cells compared with freshly isolated quail PGCs and stage X blastoderms. However, expression of the germ cell markers DAZL (P < 0.001) and VASA (P < 0.05) was higher in quail PGCs compared with enriched spermatogonial cells, stage X blastoderms, and TCs. In terms of stemness-related genes, expression of POUV (P < 0.001) was significantly higher in enriched spermatogonial cells and stage X blastoderms than in quail PGCs and TCs. There was no significant difference in POUV expression between enriched spermatogonial cells and stage X blastoderms. NANOG (P < 0.05) expression was higher in stage X blastoderms compared with the other cell types. Therefore, based on strong expression of the SSC-specific markers and stemness-related genes in purified spermatogonial cells, these cell populations may be enriched with SSCs.

Characterization of enriched spermatogonial cells, including SSCs, from isolated quail TCs. Quantitative RT-PCR was performed to evaluate enriched spermatogonial cells (SPG) using SSC-specific (GFRA1, ITGA6, and ITGB1), germness-(DAZL and VASA), and stemness-related (POUV and NANOG) genes with quail PGCs (PGC) and stage X blastoderm (BL), and TCs from juvenile quail testis as a control. Gene expression was normalized to that of ACTB and analyzed by the 2−ΔΔCt method. Differences between groups were assessed by one-way ANOVA or Student's t-test. A P value of <0.05 was considered to indicate statistical significance (*P < 0.05, **P < 0.01, ***P < 0.001). ANOVA, analysis of variance; PGC, primordial germ cell; RT-PCR, reverse transcription-polymerase chain reaction.
Testicular localization and Sertoli cell feeder layer preparation
For application of quail Sertoli cells as feeder cells for SSC culture, we first characterized the location of Sertoli cell in quail testis using an anti-vimentin antibody. Vimentin+ Sertoli cells were located in the seminiferous epithelium of seminiferous tubules. Vimentin filaments were localized to the perinuclear region of Sertoli cells and extended in thin cables toward the lumen, generating a spoke-like pattern (Fig. 3A). Next, we prepared Sertoli cells by culturing on lectin-DSA-coated plates after isolation of TCs from juvenile and adult quail testis. The cultured cells from juvenile and adult testes showed a typical epithelioid morphology with a flattened, enlarged body and considerably spread cytoplasm with three to four emanations, generating an irregular polygon appearance (Fig. 3B, C). The cultured cells from juvenile testes were further characterized by evaluating the expression of Sertoli cell markers, including vimentin, anti-Mullerian hormone (AMH), and SOX9, and used for coculturing with SSCs because the differentiation of Sertoli cells derived from adult quail took a long time. As shown in Fig. 3D–F, the majority of cultured cells from juvenile testes strongly expressed these Sertoli cell markers. Therefore, Sertoli cells as feeder cells for SSCs were isolated and cultivated using a DSA lectin method.
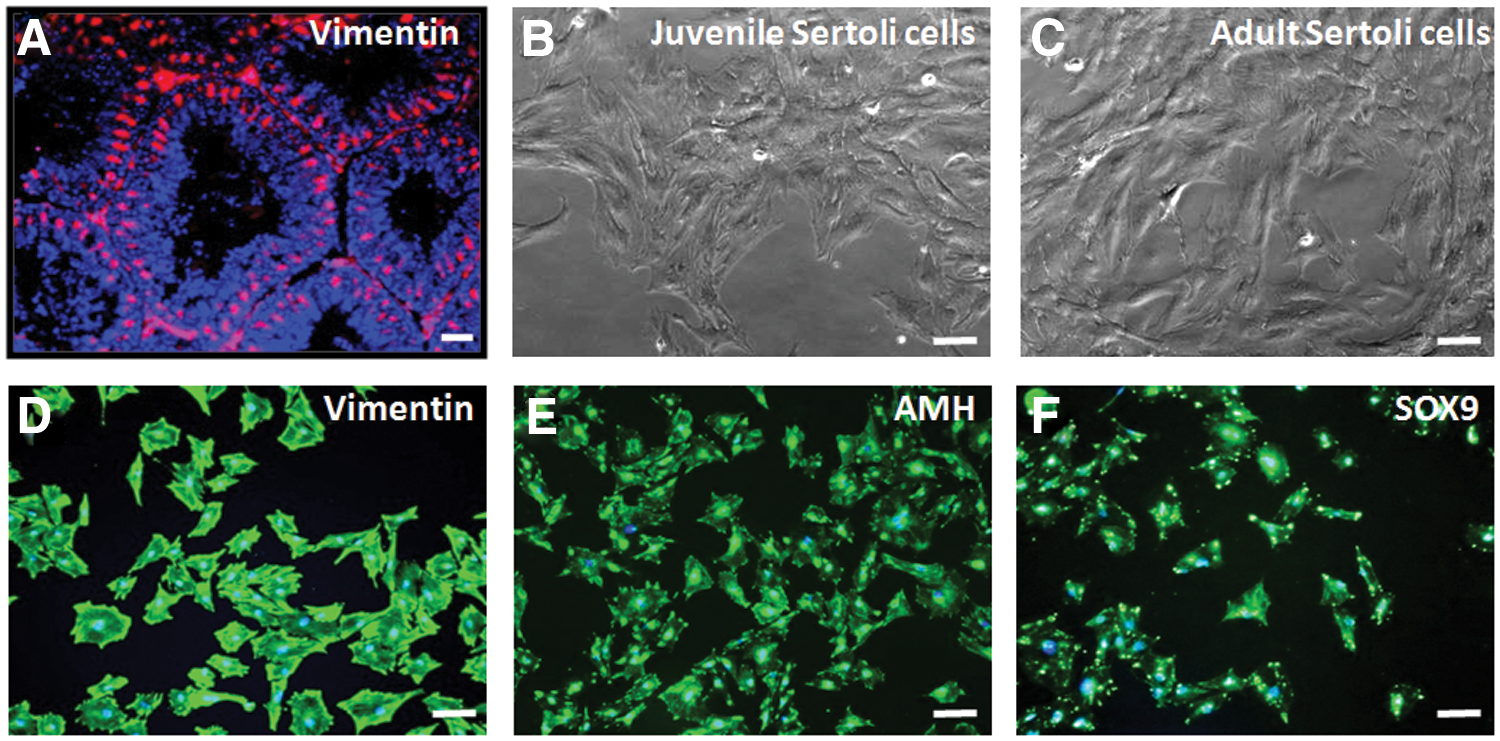
Culture and characterization of quail Sertoli cells.
Effects of feeder layers on colony formation and growth of SSCs derived from juvenile and adult quail testes
To determine the optimum conditions for isolation and culture of quail SSCs, we compared spermatogonial cells derived from juvenile and adult quail testes, and investigated the effects of different feeder layers on the expansion of quail SSCs. The isolated juvenile and adult TCs were seeded on mitomycin C-treated Sertoli cells and MEFs and on a laminin-coated plate (Fig. 4A, B). After 24 h of culture, a large number of adherent SSCs were observed on the feeders. SSCs began to form cell clusters and colonies at 48 h, and the cell clusters increased in number within 3–4 days. Colonies of SSCs cultured on feeder layers appeared earlier (from day 4 of coculturing) than those on laminin (after 6 days of culture). Therefore, juvenile or adult testis-derived SSCs cultured on Sertoli cells or MEFs formed stable colonies, whereas SSCs cultured on laminin generated few colonies and cell clusters. WST-1 value of SSCs was increased significantly during different time periods of culture indicating survival and proliferation of cells on MEF and Sertoli cell feeders. Proliferation rate of SSC was significantly increased from 2 to 4 h on MEF and Sertoli cell feeders, respectively (Fig. 4C). Subsequently, we enumerated juvenile and adult testis-derived SSC colonies and measured their diameters. As shown in Supplementary Table S1 (Supplementary Data are available online at
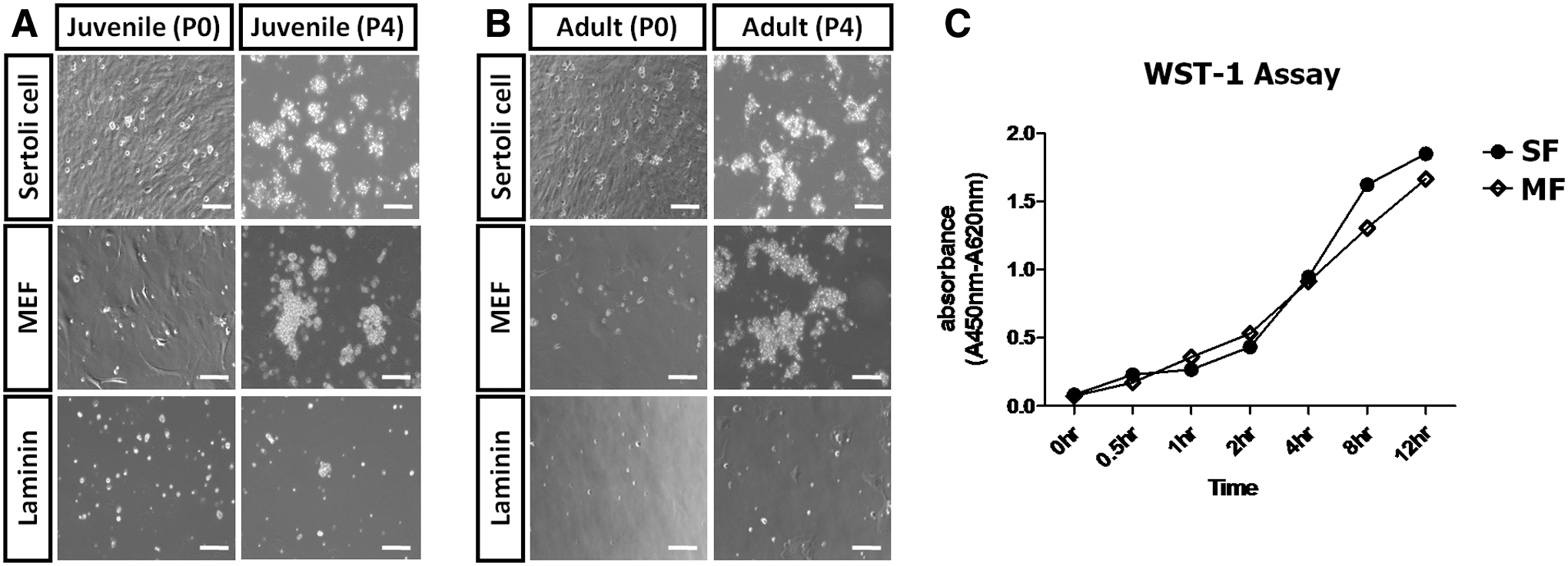
Effect of feeder layers and laminin on quail SSC culture and proliferation assay.
Characterization of juvenile or adult testis-derived SSCs cultured on feeder layers
To characterize the cellular potential of SSCs derived from juvenile or adult testis cultured on feeder layers, we investigated the expression levels of SSC-related genes—ITGA6, GFRA1, ITGB1, c-kit, DAZL, POUV, and VASA—and the Sertoli cell-related genes, AMH and SOX9. TCs were used as controls. As shown in Fig. 5, the SSC-related genes were highly expressed in juvenile or adult testis-derived SSCs cultured on Sertoli cells, as compared with those cultured on MEFs, with the exception of c-kit. In cultures of SSCs from juvenile and adult quail, the expression of GFRA1 (P < 0.001), ITGA6 (P < 0.05), ITGB1 (P < 0.05), POUV (P < 0.001) was significantly higher on Sertoli cells than MEFs. Only the expression of ITGB1 (P < 0.05) was significantly higher in juvenile SSCs than in adult SSCs. We next investigated the expression of Sertoli cell-related genes to determine the purity of SSCs cultured on feeder layers. TCs showed significantly higher expression of AMH and SOX9 than SSCs cultured on Sertoli cells and MEFs. There was no significant difference in AMH and SOX9 expression among the juvenile and adult SSCs cultured on Sertoli cells and MEFs (Fig. 5). AMH and SOX9 expression was lower in SSCs from juvenile quail testis than in those from adult quail testis. Expression of AMH (P < 0.05) was significantly higher in SSCs cultured on Sertoli cells derived from adult testis than from juvenile testis, but that of SOX9 was slightly higher in adult SSCs compared with juvenile SSCs, although not significantly so (Fig. 5). Next, we investigated by immunostaining the expression of the SSC-related markers ITGA6 and GFRA1, and the germ cell markers DAZL and VAZA, in SSCs cultured on Sertoli cells at the first and fourth passages. As shown in Fig. 6, these markers of germ cells and SSCs were expressed by the majority of the cells in SSC colonies after the first passage (Fig. 6A). Moreover, in colonies of SSCs cultured on Sertoli cells, the SSCs formed continuously proliferating, tightly packed clumps after the fourth passage, as compared with the first passage (Fig. 6B). These results indicate that Sertoli cells support the growth of quail SSCs and maintain their potential without loss of characteristics.

Characterization of quail SSC colonies generated by coculturing with MEFs and Sertoli cells as feeder layers from juvenile and adult testes after the fourth passage. Quantitative RT-PCR was performed to examine quail SSC colonies generated by coculturing in MEFs and Sertoli cells from juvenile and adult testes at P4. SSC-specific genes (GFRA1, ITGA6, and ITGB1), germness-related genes (DAZL and VASA), and stemness-related genes (POUV and NANOG), a differentiation gene (c-kit) and Sertoli cell-specific genes (AMH and SOX9) were analyzed. Gene expression was normalized to that of ACTB and analyzed by the 2−ΔΔCt method. Differences between groups were assessed by one-way ANOVA or Student's t-test. A P value of <0.05 was considered to indicate statistical significance (*P < 0.05, **P < 0.01, ***P < 0.001). SF, SSCs cultured on Sertoli cells; MF, SSCs cultured on MEFs; TC, testicular cells.
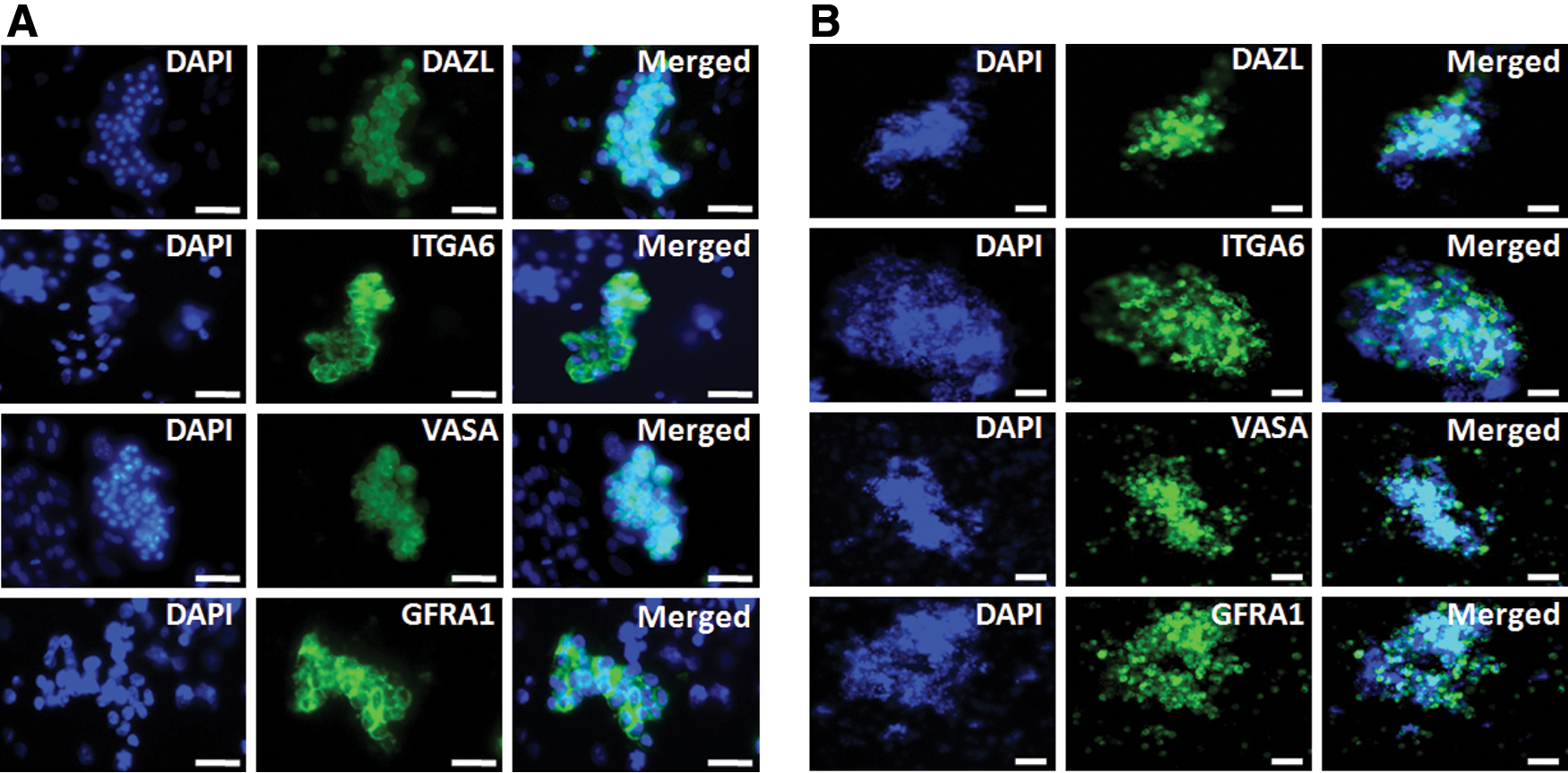
Immunostaining of stable SSC colonies cultured on Sertoli cells for SSC-related markers (ITGA6 and GFRA1) and germness markers (VASA and DAZL) after the first and fourth passages.
Migration activity of stable SSC colonies cultured on Sertoli cells and in vitro induction by RA
To find out migration activity of cultured quail SSCs into embryonic gonad, ∼1,000 cells cultured SSCs and TCs were transplanted into the dorsal aorta of recipient 2 days old (Hamburger and Hamilton stage 14–16) quail embryos after red fluorescent dye labeling with PKH26. As shown in Fig. 7A, PKH26-labeled quail SSCs were exclusively detected in the 5 days old gonads of recipient embryos, whereas PKH26-labeled quail TCs were not detected, indicating that transplanted quail SSCs could localize into the developing embryonic gonads.
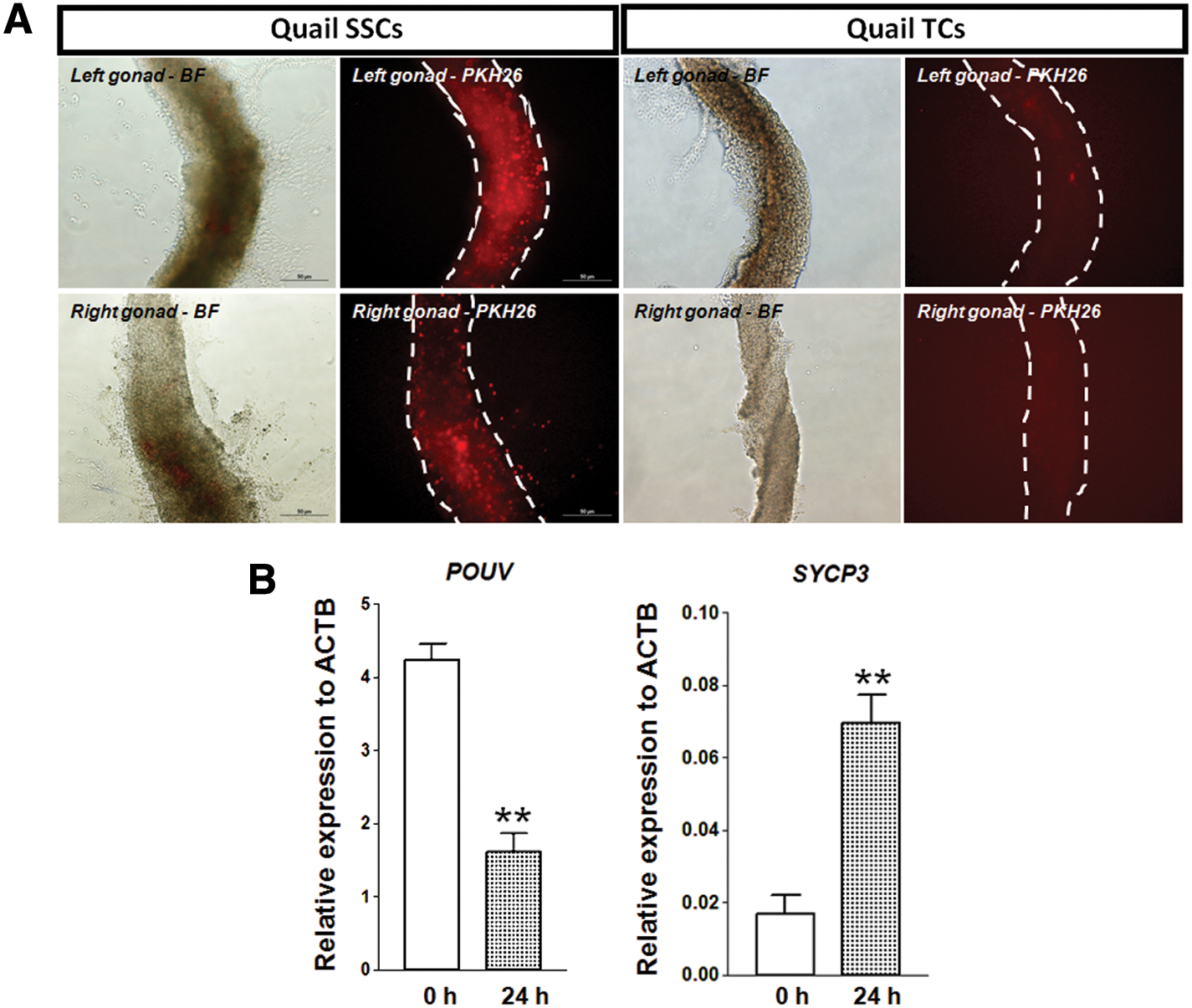
Migration activity and induction of quail SSC colonies cultured on Sertoli cells.
To ensure that the cultured SSCs have stemness, we used all-trans RA in SSC colonies cultured on Sertoli cells to induce meiosis. As shown in Fig. 7B, we found that synaptonemal complex protein 3 (SYCP3) expression, as a meiotic germ cell marker, was significantly increased, whereas POUV expression was significantly decreased after 24 h of RA treatment.
Discussion
Production of germline chimera in quail is valuable for avian conservation and development of transgenic systems due to their short generation interval, disease resistance, and high laying capacity [32 –36]. In our previous studies, in vitro maintenance of germ cells retrieved from embryos in quail had shown successful germline contribution [28,37]. However, germline transmission systems that can be applied after hatching have not been developed in this species. Thus, in this study, we established a method for isolation and cultivation of quail SSCs to overcome the limitations of embryo-mediated germline transmission systems.
First, we investigated the phenotypic characteristics of quail SSCs from isolated TCs using the mammalian SSC markers DAZL, ITGA6, and GFRA1. DAZL is essential for germline development in mammals [38] and is expressed during chicken germline development [39,40]. Immunostaining indicated that DAZL was strongly expressed in the majority of quail TCs (Fig. 1B), suggesting that SSCs are a subset of the total TC population. Subsequently, we applied mammalian SSC-specific antibodies against ITGA6 and GFRA1 to the quail TCs. ITGA6 (also known as CD49f) is the first known surface marker of mouse SSCs and progenitor cells [41] and has also been reported to be a marker of chicken SSCs [19]. Meng et al. demonstrated that GDNF and GDNF-RET-GFRA1 signaling is essential for the self-renewal and maintenance of undifferentiated SSCs [10]. In addition, GFRA1 is a marker of SSCs [10,42]. ITGA6+ and GFRA1+ cells were present among DAZL+ quail TCs (Fig. 1B). Therefore, we confirmed that ITGA6 and GFRA1 are markers of quail SSCs as well as those of mammals. These results indicate that ITGA6+ and GFRA1+ quail TCs may be an SSC subset of spermatogonial cells.
We enriched spermatogonial cells using differential plating followed by the attachment to laminin-coated plates [43], and examined their expression of genes specific to SSCs and markers of germness and stemness. As shown in Fig. 2, comparative analysis with quail PGCs and stage X blastoderms suggested that these enriched spermatogonial cells exhibited the characteristics of SSCs. ITGA6, ITGB1, and GFRA1 are known markers of mouse and human SSCs [41,44] and have been used in genetic analyses. The higher expression of SSC-specific markers—such as ITGA6, ITGB1, and GFRA1—in the enriched spermatogonial cells suggests that SSCs are abundant in the whole TC population. However, the expression of germline-specific genes, such as VASA and DAZL, which are the most prominent markers of germ cells, was lower in spermatogonial cells than in quail PGCs. These results are consistent with our previous report of higher expression of DAZL in PGCs, as compared with testicular and ovarian cells [39]. POUV expression was higher and that of NANOG was lower in enriched spermatogonial cells than in stage X blastoderms. These results suggest that POUV is more important than NANOG in regulation of the self-renewal and pluripotency of quail SSCs. Oct4 expression in mouse gonocytes and undifferentiated SSCs has been reported [45], and is essential for maintenance of the SSCs in vitro and the preservation of their self-renewal property [46].
The maintenance of enriched spermatogonial cells in vitro requires a suitable environment. Culture of mammalian SSCs is challenging primarily due to reduced viability and proliferation, and transformation of cell characteristics [47]. Laminin is secreted by Sertoli cells as an extracellular matrix (ECM) molecule, and binds to α6 and β1-integrin receptors on the surface of spermatogonial cells. Laminin provides conditions suitable for mouse SSC maintenance and proliferation in vitro [43]. Therefore, in the present study, we seeded quail SSCs on laminin-coated plates for expansion. However, SSCs derived from juvenile and adult quail did not proliferate on the laminin-coated plates (Fig. 4A, B), suggesting that more suitable conditions were required. Coculturing of mammalian SSCs on feeder layers (such as MEFs and Sertoli cells) as alternatives to laminin has been proposed [11,13,48,49].
To prepare Sertoli cells as a feeder layer, we characterized their localization in the testis by immunohistochemistry (Fig. 3A) using an anti-vimentin antibody [50]. Additionally, we confirmed the expression of markers of Sertoli cells, such as AMH and SOX9. Stable colonies without differentiation were generated on Sertoli cells and MEFs after 2 months of in vitro culture of quail SSCs from adult and juvenile testes (Fig. 4A, B). WST-1 assay showed that quail SSCs were increased significantly during different time periods of culture indicating survival and proliferation of cells on MEF and Sertoli cell feeders (Fig. 4C). Furthermore, cultivation of juvenile and adult SSCs on Sertoli cells resulted in a greater number of colonies of larger diameter compared with those cultured on MEFs (Supplementary Tables S1 and S2). However, except juvenile quail SSCs on Sertoli cell feeder, others conditions showed a reduction in the number of colonies after fifth passage.
Next, we evaluated the characteristics of stable SSC colonies cultured on Sertoli cells and MEFs. After the fourth passage, juvenile quail SSCs cultured on Sertoli cells showed strong expression of SSC-related genes, including ITGA6, GFRA1, ITGB1, DAZL, POUV, and VASA (Fig. 5). cKIT expression is commonly used to differentiate mouse spermatogonial cells, and the gene product is a member of the tyrosine kinase receptor family expressed in late Aa1 (A-aligned) spermatogonia and differentiated SSCs of mice [51]. Therefore, the lower expression of c-kit in the spermatogonial cells suggests them to be undifferentiated SSCs. Furthermore, quail SSC colonies were devoid of somatic cells, as evidenced by the lower expression of AMH and SOX9.
Immunostaining indicated that ITGA6 and GFRA1 were expressed at the first and fourth passages (Fig. 6). Subsequently, we evaluated migration activity of quail SSCs cultured on Sertoli cell feeder. As shown in Fig. 7A, transplanted quail SSCs could localize into the developing embryonic gonads. To ensure that the cultured SSCs have stemness, we treated all-trans RA in SSC colonies cultured on Sertoli cells to induce meiosis. Based on our result, we found significant increase in the expression of meiotic germ cell marker, SYCP3, whereas POUV expression was decreased after 24 h of treatment (Fig. 7B). Therefore, the stable SSC colonies had stemness and self-renewal properties, suggesting that quail spermatogonial cells and SSCs have phenotypes similar to those of mammals.
In conclusion, we demonstrated for the first time the efficient isolation, characterization, and in vitro culture of quail SSCs. Our results demonstrate that mammalian SSC-specific markers (ITGA6 and GFRA1) can be used to identify quail SSCs, and that quail SSCs cultured on Sertoli cells feeder maintain expression of markers of SSCs and germ cells after several passages. The use of quail SSCs will overcome the current limitation of germ cell availability for avian transgenesis and will increase our understanding of spermatogenesis.
Footnotes
Acknowledgments
This work was supported by a National Research Foundation of Korea (NRF) grant funded by the Korean government (MSIP; no. 2015R1A3A2033826), Bioindustry Technology Development Program (IPET312060-5), Ministry for Food, Agriculture, Forestry, and Fisheries, and “Cooperative Research Program for Agriculture Science & Technology Development (project no. PJ0122362016)” Rural Development Administration (Republic of Korea).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.