
Abstract
Hepatocytes constitute the main bulk of the liver and perform several essential functions. After injury, the hepatocytes have a remarkable capacity to regenerate and restore functionality. However, in some cases, the endogenous hepatocytes cannot replicate or restore the function, and liver transplantation, which is not exempt of complications, is required. Stem cells offer in theory the possibility of generating unlimited supply of hepatocytes in vitro due to their capacity to self-renew and differentiate when given the right cues. Stem cells isolated from an array of tissues have been investigated for their capacity to differentiate into hepatocyte-like cells in vitro and are employed in rescue experiments in vivo. Adult stem cells have gained in attractiveness over embryonic stem cells for liver cell therapy due to their origin, multipotentiality, and the possibility of autologous transplantation. This review deals with the promise and limitations of adult stem cells in clinically restoring liver functionality.
Introduction
T
The hepatocytes have a remarkable capacity to regenerate after injury and restore functionality. However, when the inherent regenerative capacity of hepatocytes is impaired, liver transplantation becomes the only efficient treatment. This procedure, however, is highly invasive, is limited by the scarcity of donors, and is not exempt of complications. Following transplantation, the outcome of the patient may change on the basis of hypotension, hypoxia, ischemia and hepatotoxic drugs, surgical-related aspects (intra- or postoperative hemorrhage, vascular or biliary complications, infections), problems posed by prolonged use of immunosuppressive drugs (infections, toxicity, and possible neoplastic risks), or rejection [7].
Transplantation of hepatocytes is an attractive alternative. The first hepatocyte infusion was carried out as early as 1976 in the Gunn rat model, which closely mimics the pathological manifestations in Crigler–Najjar syndrome type I patients [8,9]. Following modeling of hepatocyte transplantation in several animal models, the first preclinical study was performed in 1992 in nonhuman primates to evaluate the safety and efficacy of infusing adult hepatocytes in the portal vein [10,11]. Thereafter, human clinical hepatocyte transplant has been carried out in several liver diseases, mostly as therapy for innate metabolic failures or postresectional liver failure (as a definitive treatment) in an effort to restore functionality [8,12]. More than 30 human cases receiving hepatocyte transplantation for metabolic diseases have been reported hitherto [13].
Human hepatocyte treatment for fulminant or chronic liver failure with acute decompensation (as a bridge to liver transplantation) has extended the life expectancy of patients for years [8,9,14]. Hepatocytes have been isolated from donated human livers considered unsuitable for organ transplantation. Not always good quality hepatocytes are obtained from these livers (due to long, cold ischemic times or presence of steatosis) and, when cultured in vitro in different conditions (oxygenated cocultures or microfabricated culture chips, for instance), these cells lose their metabolic functions after some time [9,15,16].
Recently, a long-term culture and expansion method of primary human hepatocytes has been described [17]. Primary human hepatocytes were transduced with human papillomavirus genes, E6 and E7, which induce the expression of the oncostatin M (OSM) receptor, gp130. Thus, treatment of the cells with OSM promoted proliferation, and hepatocytes could be expanded for up to 40 population doublings, producing bulk quantity of cells in vitro. Removal of OSM enhanced the differentiation of the hepatocytes into cells with transcriptional, metabolic, and toxicity profiles resembling those of primary hepatocytes [17]. This is a major leap forward in the quest of finding conditions to expand primary human hepatocytes, which maintain the capacity to undergo maturation in vitro. At the moment, this is the cell therapy with the best evidence of engraftment and generation of functional liver parenchyma compared with hepatocytes derived from other sources (described below) [13].
Still, much work needs to be done to obtain cell therapy-quality hepatocytes, produced under Good Manufacturing Practice (GMP) conditions, for liver transplant in humans. There are, however, problems with long-term survival of hepatocytes, and importantly, despite the fact that no HLA matching is required for hepatocyte transplant in the liver (hence allowing the use of ample donor-derived stem cells for cell therapy), immunosuppression is still required [9,18]. Thus, the potential advantages of using hepatocyte infusions must be carefully evaluated against any possible side effects resulting from the use of immunosuppressive drugs, sepsis, and embolization of cells into the lungs, for example [19].
Another promising advance in the field of liver cell therapy has been made with the discovery of stem cells and their potentiality. Stem cells offer, in theory, the possibility of generating unlimited supply of hepatocytes due to their capacity to self-renew and differentiate when given the right cues. The perfect stem cell-derived hepatocyte would be one which changes the trajectory of the liver disease and definitively restores functionality. Hepatocyte-like cells fulfilling, at least partially, these criteria have been obtained from both pluripotent embryonic stem (ES) cells and adult stem cells. The clinical application of ES cells, however, has been hampered by ethical concerns raised over the embryo use and the problems of immune incompatibility between donors and recipients.
Another major concern is the risk of tumor formation (teratomas) arising from ES cell injection. It has been shown that teratomas form in vivo not only from undifferentiated human ES cells but also from human ES cells differentiated in vitro for long periods of time [20]. Adult stem cells, on the other hand, despite their lower engraftment and repopulation efficiency compared with mature hepatocytes, have gained in attractiveness for liver cell therapy due to their multipotentiality, ability to differentiate into hepatocyte-like cells, and especially for their use in an autologous way. This review deals with the promise and limitations of adult stem cells in restoring liver functionality.
Methodology
A MEDLINE search of all studies published in English (last performed on 15th June 2016) was conducted. Articles on the use of hepatocytes and stem cell-derived hepatocyte liver diseases were identified and selected using the terms “Liver transplantation AND hepatocytes AND stem cells”.
Stem Cells Differentiated into Hepatocytes In Vitro
Hepatocytes are the main cell type in the liver, accounting for 70% of the mass of the adult organ. Hepatocytes, along with biliary epithelial cells or cholangiocytes, are derived from the embryonic endoderm that emerges at embryonic day (E) 7.5 in mouse and in the third week of human gestation [21]. The activity of two important cytokines, fibroblast growth factor (FGF) secreted by the developing heart and bone morphogenetic protein arriving from the septum transversum mesenchyme, orchestrates the endoderm cells to adopt a hepatic fate [22]. The liver-committed foregut endoderm cells then express transcription factors, Hex and hepatocyte nuclear factor 4α (HNF4α), and the liver-specific genes, α-fetoprotein (AFP) and albumin. These cells migrate as cords into the surrounding septum transversum mesenchyme and become the common progenitor cells (hepatoblasts), which give rise to both hepatocytes and cholangiocytes during liver development [22].
Factors employed for directing hepatocyte specification and maturation to induce hepatic differentiation of stem cells and in vitro differentiation protocols have emerged from the knowledge acquired on fetal liver development in vivo. Several types of adult stem cells have been differentiated in vitro into functional hepatocytes and employed in preclinical studies, as well as in clinical settings (Table 1). Some examples are described below.
Some examples taken from the literature are indicated.
GPSCs, germline cell-derived pluripotent stem cells; HSCs, hematopoietic stem cells; MSCs, mesenchymal stem cells.
Hematopoietic Stem Cells
Hematopoietic stem cells (HSCs) and hepatocytes are entwined since early embryonic development. The mid-gestation fetal liver is the major transitional site for mammalian hematopoiesis [21]. HSCs secrete several cytokines and growth factors that promote hepatocyte maturation. HSCs then migrate to the fetal bone marrow, and the switch from liver to bone marrow hematopoiesis occurs at around E16 in mice and after 70 days' gestation in humans [23,24]. The finding that the multipotent HSCs can participate in liver repair in adult life is therefore not such a surprise. HSCs are the most studied adult stem cells and can be isolated through the expression of c-Kit+Sca-1+Lineage− in rodents and CD34 or CD133 in humans [25]. It has been shown that adult bone marrow stem cells such as HSCs and mesenchymal stem cells (MSCs, discussed below) are a potential source of hepatic progenitor cells, termed oval cells (described below), in rodents and are capable of restoring liver functions, for example, in the lethally irradiated tyrosinemic FAH−/− mice, or following carbon tetrachloride (CCl4)-induced liver fibrosis [23,26].
There are also several human clinical studies reporting the improvement in liver function, for example, following HSC transplant in liver diseases, or in the setting of patients needing liver regeneration before hepatic resection of metastases [27 –29]. The small number of cases reported and lack of defined primary endpoint, as well as absence of controls, render these results difficult to interpret and evaluate [27]. Furst et al. elegantly showed how the infusion of autologous CD133+ cells into the nonoccluded portal vein following portal venous embolization with chemotherapy caused an extensive increase in the residual liver lobe size [29]. This increase in liver volume was significantly higher in the CD133-treated group compared with controls, thus permitting earlier surgery for resection of hepatic metastases.
In a recent study, Schmelzle et al. showed that a subset of murine and human HSCs express high levels of CD39, which directs the chemotaxis and recruitment of these CD39high HSCs from bone marrow to liver [30]. The CD39-positive HSCs significantly increased in the bone marrow after 70% hepatectomy in mice and promoted liver regeneration. In the clinical setting, mobilization of human HSCs was observed 2 days after liver hepatectomy and was associated with the extent of liver resection as well as growth and functionality after hepatic resection. However, the mechanisms by which extrahepatic stem and progenitor cells participate in liver regeneration still need elucidation.
Mesenchymal Stem Cells
MSCs, due to their multipotency, immunoregulatory/immunosuppressive, and migratory properties and quasi-unlimited availability, have extensively been investigated as potential therapeutic options for the treatment of several degenerative diseases [31]. MSCs are derived from adult tissues and are considered safer in terms of tumorigenicity with respect to pluripotent stem cells (described below) [32]. These MSCs have been obtained from several tissues, including bone marrow, cord blood, amniotic fluid, adipose tissue, and most recently, from the liver [33 –38].
In particular, there are several advantages in using adipose tissue-derived MSCs, or adipose-derived stem cells (ASCs), for hepatocyte derivation—their abundance and easy accessibility from minimally invasive procedures such as liposuction surgery and their higher yield of stem cells upon isolation with respect to other tissues [39,40]. This is a crucial point as at least 1 million/kg body weight of MSCs are required for transplantation each time [41]. Meticulously performed studies have provided the evidence that cells with functional characteristics of hepatocytes can be obtained from multiple tissue-derived MSCs in vitro. For example, when MSCs were treated with HGF, nicotinamide, and dexamethasone, these cells exhibited features that closely resembled human adult hepatocytes and expressed hepatic markers such as albumin and AFP [42].
MSCs act at several levels in tissue restoration after injury. These cells provide immunomodulatory, anti-inflammatory, antifibrotic, antiapoptotic, and proproliferative features [43,44]. Several clinical studies using autologous MSCs, sometimes in the presence of cell-maintained biomaterials, have been performed in various fields (Table 1). For instance, Koh et al. have shown that the injection of ASCs and platelet-rich plasma into 18 patients with osteoarthritis or degenerative cartilage effectively reduced pain and improved knee function in patients undergoing treatment for knee osteoarthritis [45]. However, it is their in vivo propensity to stimulate resident cells and matrix remodeling, through expression of trophic factors (growth factors and antifibrotic molecules such as HGF and cytokines) to promote the differentiation of native progenitor cells as well as the recovery of injured cells, which makes MSCs optimal candidates for liver repair [28,41,46]. For example, allogenic bone marrow-derived MSC transplantation has been performed in patients with primary biliary cirrhosis, and the outcome was the improvement in liver function [47].
Other clinical trials investigating the effect of bone marrow-derived stem cells in patients with liver disease have been performed and have established the safety and feasibility of stem cell therapy in patients with liver diseases (extensively reviewed in Houlihan and Newsome [44]). On the other hand, some studies have pointed out the possibility that MSCs, for instance, those derived from bone marrow, may contribute to the hepatic stellate cell populations and myofibroblast pool and thus promote the fibrotic process within the liver [48,49]. Excessive extracellular matrix deposition may occur, overcoming the MSC-induced cytoprotective effect taking place during the initial repair process [41]. Thus, more clinical studies are needed to ensure that MSCs, especially the ones derived from the bone marrow, can be safely employed to generate new hepatocytes to replace damaged ones without leading to fibrosis.
Adult Pluripotent Stem Cells
The pluripotent stem cells par excellence are the ES cells, but due to ethical and immunological concerns, their use in the clinic has been forbidden in most countries. The scientific breakthrough of the group of Yamanaka has offered the possibility of obtaining adult pluripotent alternatives [50]. Induced pluripotent stem (iPS) cells, generated by reprogramming of somatic cells, are very attractive for regenerative therapy for the following reasons. These adult cells can be expanded for a long period in culture without karyotype change; they can be frozen and thawed without loss in viability, as may happen for other types of adult stem cells; and most importantly, they can give rise to derivatives of the three germ layers upon differentiation induction without the concern for immune rejection for potential patient-tailored therapy [50,51].
Functional hepatocytes have been derived in vitro from human iPS cells, both of human and mouse origin, by several groups [52 –54]. HNF4, albumin, cytokeratin 18 (CK-18), glucose 6-phosphate (G-6P), cytochrome P450 3A4 (CYP3A4), and cytochrome P450 7A1 (CYP7A1) were expressed by the iPS cell-derived hepatocytes [54]. Moreover, in functional assays, these cells showed accumulation of glycogen and demonstrated uptake and excretion of indocyanine green, as well as secretion of albumin into the medium [53]. These iPS cell-derived hepatocytes rescued lethal fulminant hepatic failure in mice [53,54]. Thus, patient-specific iPS cell-derived hepatocytes can be obtained for promoting liver regeneration in vivo or for therapeutic compound testing or studying disease mechanisms in vitro.
However, one of the most important concerns about use of iPS cells compared with that of adult stem cells is the possibility of developing some tumoral tissue [55]. Thus, it is important to ensure that full differentiation of ES-like cells has occurred before proceeding to transplantation in humans. Efforts are now being made to improve the mechanism and method for generating tumor-free iPS cells, such as employing the suicide gene strategy or using regulatory miRNAs in iPS cell-based therapy [55,56]. Despite all hurdles and controversies, clinical trials in treatment of spinal cord injury, macular degeneration of retina, type 1 diabetes, and heart failure are under way [57].
Pluripotent stem cells derived from spermatogonial stem cells, termed herein as germline cell-derived pluripotent stem cells (GPSCs), are also adult stem cells with the major features of ES cells. The difference with iPS cells is that mouse GPSCs are obtained spontaneously from long-term cultures of spermatogonial stem cells and do not necessitate the genetic reprogramming by forcing gene expression for their derivation. GPSCs are thus of particular interest for tissue regeneration [58].
The first clue that GPSCs can be directed to differentiate into hepatocyte-like cells came from the study of Guan et al. [59]. GPSCs were cultured as hanging drops for 5 days and plated thereafter. Hepatocyte-like cells were observed in the embryoid bodies, and gene expression analyses revealed that the early (AFP) and the late (CK-18) markers of hepatocyte differentiation were expressed. Successively, GPSCs were induced, using two different hepatic differentiation approaches, to differentiate into hepatocytes in vitro and were compared with other pluripotent stem cells [60]. GPSC-derived hepatocytes showed prominent expression of AFP, albumin, CK-18, and c/ebp-α and acquired a polygonal-shaped hepatic morphology. These cells also secreted albumin in culture supernatant. We confirmed these results and further showed that GPSC-derived hepatocytes were metabolically active as shown by albumin and haptoglobin secretion in the culture supernatant, urea synthesis, glycogen storage, and indocyanine green uptake [51].
Infection of GPSC-derived embryoid bodies with lentivirus expressing GFP under hepatocyte-specific enhanced transthyretin promoter revealed that 82.68% of cells were hepatocytes in long-term culture (on day 35 of differentiation) [51,61]. An OP9 coculture system was also shown to efficiently generate functional and mature hepatocyte cells from GPSCs [62]. Recently, our group demonstrated that the hepatoblast marker, Liv2, -sorted GPSC-derived hepatocytes were functional in vitro and could engraft and colonize Hfe-null mouse livers following monocrotaline treatment and partial hepatectomy [63]. These studies showed that GPSC-derived hepatocytes could undergo long-lasting engraftment in the mouse liver and are promising for liver regeneration.
The starting cell type—spermatogonial stem cells—also show multipotent characteristics when given the right differentiation cues. In this regard, it was demonstrated that both mouse spermatogonial stem cells can directly transdifferentiate in vitro into cells with morphological, phenotypic, and functional attributes of mature hepatocyte-like cells [64]. More recently, mature and functional hepatocytes have been also obtained by direct differentiation from freshly isolated human spermatogonial stem cells [65]. Thus, it may be possible to rapidly and directly differentiate adult spermatogonial stem cells into mature and functional hepatocyte-like cells, without waiting for the pluripotency acquisition process. The field is now open to finding the right culture conditions for the long-term culture of human spermatogonial stem cells (to derive human GPSCs) and to assess the clinical potentiality of both spermatogonial stem cells and GPSCs.
Resident Stem Cells
Mature hepatocytes are usually quiescent and show minimal turnover [66]. However, following injury induced by partial hepatectomy or chemical insult, hepatocytes exit the G0 phase and undergo cell proliferation to compensate for cell loss [42]. Resident stem cells are also thought to be involved in the liver regeneration process. Oval cells, defined as a population of small proliferating cells with oval-shaped nuclei in rodents, surge within the liver following injury [67]. Oval cells are bipotent cells that are capable of generating both hepatocytes and bile duct cells [68]. Some studies have reported the identification of oval cell equivalent in humans in the so-called ductular reaction [68]. Bipotential progenitor epithelial cells resembling rodent oval cells, morhologically and immunohistochemically, have been traced in the regenerating areas in patients with chronic liver injury in subacute hepatic failure [69,70].
Recently, Huch et al. have reported the long-term culture of adult bile duct-derived bipotent stem cells isolated from human liver biopsies [71]. Interestingly, these cells could be induced to differentiate into functional hepatocyte cells in organoids in vitro and upon transplantation in vivo in mice. These data suggest that the human counterpart of oval cells exists and may participate in liver regeneration. Whether these cells originate in the liver itself or in extrahepatic sites such as the bone marrow still remains a matter of doubt [72]. Lineage tracing experiments may help in this direction. Some studies indicate that the activation of progenitor cells (oval cells in rodents) may lead to the histogenesis of hepatocellular carcinoma or in the development of liver fibrosis [73,74].
More studies are thus needed to clarify the mechanisms that in one instance lead to the hepatic differentiation of oval cells in normal liver and, on the other hand, promote their carcinogenicity under pathological conditions.
A population of human liver stem cells (HLSCs) has also been isolated from surgical specimens of patients undergoing hepatectomies or from cryopreserved normal hepatocytes [68,75]. These cells express MSC markers and can be induced to differentiate into mature hepatocytes when cultured in the presence of HGF and FGF4 in rotary cell culture system in vitro. In vivo, HLSCs contributed to liver regeneration when injected in SCID mice with N-acetyl-p-aminophen-induced acute liver injury and improved survival of mice with fulminant liver failure [68,75]. Thus, these preclinical studies are a prelude to assessing the clinical use of HLSCs. Regarding the use of human adult liver stem cells in treating inborn errors of liver metabolism, patients have been included in phase I/II studies, including Crigler–Najjar and urea cycle disorders [76].
In Vitro Systems for Obtaining Functional Hepatocytes from Stem Cells
Stem cell-derived hepatocytes cultured on different extracellular matrix components in monolayers do not undergo full maturation in vitro. Using 3D bioreactors and rotating microgravity conditions, some studies have demonstrated that hepatocyte viability and function can be improved and maintained for several weeks [68,77]. However, there are contrasting data regarding the effect of microgravity on differentiation. Lately, it has been reported that microgravity reduces the differentiation and regenerative properties of stem cells [78]. Thus, other settings have to be devised to promote maturation of stem cells into functional hepatocytes in vitro. It is known that hepatocytes need heterotrophic cell interactions with nonparenchymal cells, which regulate differentiation and proliferation in a 3D microenviroment in vivo [79].
Importantly, stem cell-derived hepatocytes need the optimal microenvironment when transplanted in the case of end-stage chronic liver disease, for instance, in which the liver architecture is extensively destroyed and the liver parenchyma is replaced by fibrosis with reduced blood supply. Generation of acellular liver scaffolds from rat or human liver for repopulating these with stem cell-derived hepatocytes is a great step forward in mimicking the in vivo system (Fig. 1). For instance, rat liver scaffolds were recellularized with HLSCs and maintained for 21 days in different culture conditions to evaluate hepatocyte differentiation [80]. The rat extracellular matrix enhanced HLSC differentiation into functional and metabolically active hepatocytes and promoted the formation of both epithelial-like and endothelial-like cells [80].
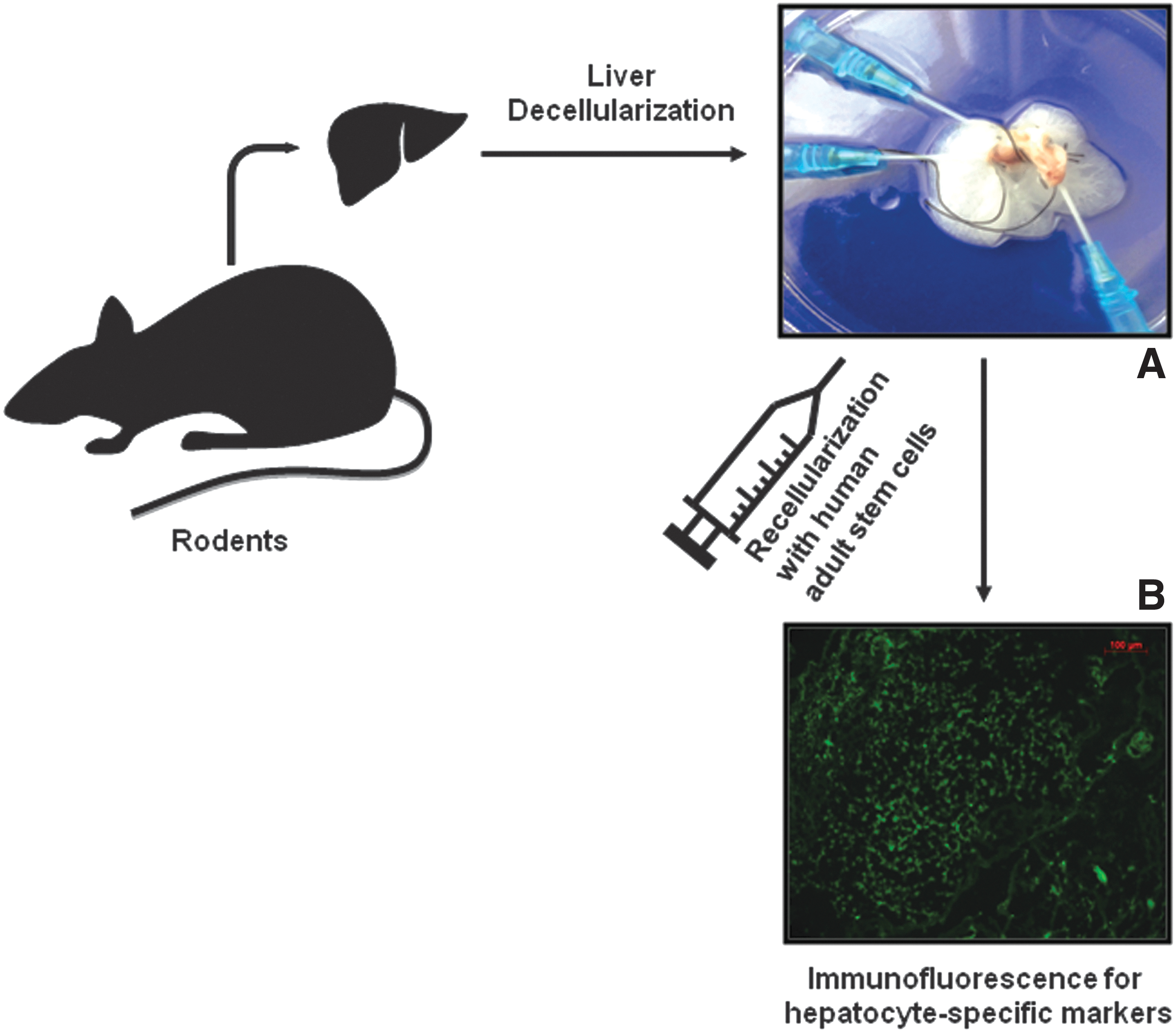
Scaffold of a rodent liver decellularized by perfusion and recellularized with human adult stem cells. The differentiation and functionality of the injected cells
Human segmental lobes or whole livers have been also decellularized and characterized [81]. The scaffold was repopulated with different human hepatic cell lines for up to 21 days and allowed efficient engraftment, homing to correct location, and proliferation of these cells. The human liver scaffold also showed biocompatibility when xenotransplanted into immunocompetent mice [81]. This is major progress in devising bioartificial livers for repopulation with stem cell-derived hepatocytes for the treatment of liver diseases.
Modeling Human Diseases for Assessing Stem Cell-Derived Hepatocyte Function
Stem cell-derived hepatocytes have been transplanted mainly in rodents to test their functionality in vivo. To counteract the rejection of human cells by the rodent immune system, immunodeficient mice have been employed for the transplantation of stem cell-derived hepatocytes. However, often the human cells do not proliferate in normal livers. Thus, several animal models, in which proliferation of endogenous hepatocytes is severely compromised either by genetic causes or by accumulation of toxic products, have been generated. For instance, fumarylacetoacetate hydrolase (FAH)-deficient mice and urokinase (alb-PA) transgenic mice with liver regeneration defects have been transferred into an immunocompromised background to receive human cells [79]. Transplanted stem cell-derived hepatocytes thus have selective growth advantage over endogenous hepatocytes, and homing, engraftment, and proliferation, as well as functionality, can be assessed.
Mice with inherited liver metabolic diseases caused by genetic mutations in key proteins, for example, Crigler–Najjar in which the UDP-glucuronosyltransferase A1 (Ugt1A1) gene is mutated, have been generated, and crossing these mice with immuno-compromised mice will help in assessing whether human stem cell-derived hepatocytes can rescue the phenotype [82]. These mice have a normal liver, and a way to make exogenous hepatocytes home in and proliferate has to be found. The engrafted cells will thus restore the activity of the enzyme, Ugt1A1. Thus, apart from cell therapy, stem cells also serve as a platform for gene therapy. In humans, such an approach can be envisaged in personalized gene/stem cell therapy combination following genome editing.
Several routes of cell delivery to the liver have been exploited. The most convenient mode of stem cell-derived hepatocyte transplantation is the intravenous route [83]. This delivery route takes the injected cells to the lungs and successively to the liver. Liver injury and release of cell-attracting chemokines are required for homing of injected cells. The spleen is also a good site for hepatocyte injection. The hepatocytes infused into the spleen undergo blood flow-mediated translocation to the hepatic sinusoids [84]. However, a large number of cells remain trapped in the spleen. Other routes of infusion include hepatic artery and portal vein [85]. Intraparenchymal hepatocyte transplantation is another way of delivering cells to the liver [63]. Directed injection results in grafted cells that remain localized to the liver.
Most recently, hepatocyte sheets were attached onto the surface of mouse livers following injury and showed high potential for cell engraftment of this sheet transplantation procedure [86]. Thus, the therapeutic values of stem cell-derived hepatocytes can be assessed in vivo using these different routes of cell delivery to liver.
Stem Cell Secretome in Mediating Beneficial Effect on Liver
Liver regeneration occurs not only through transplanted stem cell proliferation and maturation but also by the paracrine factors released by these cells. It was shown that microvesicles, which are shed vesicles ranging from 100 nm to 1 μm in size, can horizontally transfer genetic information to target cells [87]. In vivo administration of microvesicles derived from HLSCs promoted in vivo liver regeneration in 70% hepatectomized rats [88]. The HLSC-derived microvesicles contain ribonucleoproteins involved in RNA biogenesis, microRNAs associated with multiple cell functions such as transcription, translation, and proliferation, as well as proteins [89].
Microvesicles also have other interesting properties. Recently, Fonsato et al. demonstrated that HLSC-derived microvesicles could block the proliferation and survival of hepatocellular carcinoma cells in vitro and induce the regression of ectopic tumors developed in SCID mice [90]. MicroRNAs contained in the microvesicles mediated the antitumor activity. Interestingly, cells can be genetically engineered to produce microvesicles carrying suicide mRNAs/proteins to target tumor cells [91]. MSC-derived extracellular vesicles have also been beneficial in several acute and chronic liver disease models (reviewed in Rani et al. [92]). Recently, comparison of adult-derived human liver stem/progenitor cells (parenchymal) with hepatic stellate cells (nonparenchymal) revealed that the former secreted cytokines with therapeutic and immunomodulatory properties with respect to hepatic stellate cells [46]. Thus, stem cells can be beneficial for liver therapy not only physically but also by effectively delivering therapeutic mRNA/proteins for repair and tumor regression.
The Still Unmet Challenges
Several aspects of liver cell/gene therapy need further investigations before stem cells can be efficiently used in the clinic. In animal models employed hitherto, it has been possible to block endogenous hepatocyte proliferation with alkaloids such as monocrotaline, hence favoring the replication of transplanted stem cells. In patients, however, a similar treatment cannot be applied. Thus, strategies to harness growth advantages of stem cells in human liver, which present a normal phenotype, like in the case of Crigler–Najjar syndrome, should be elaborated. Future studies will determine whether cotransplantation of stem cell-derived hepatoblasts and helper liver-derived stromal cells will engraft and gradually replace morphologically (but not metabolically) normal hepatocytes as was described in the rat [13].
However, fibrosis, which may occur as a side effect of such a procedure, should be taken into consideration. Standardization and protocols should be implemented for isolation, purification, culture conditions and passages, expansion, and hepatocyte-specific induction following GMP guidelines [93]. To which extent a stem cell should be differentiated in vitro to be considered fit for transplantation should be also standardized. In this regard, expression of markers of hepatocyte differentiation should be closely followed to assess the differentiation stage before cell injection, and whether hepatoblasts or fully mature hepatocytes should be injected in the diseased liver also remains a matter of concern, especially because some studies have shown that fetal hepatocytes proliferate, engraft, and repopulate livers better than more mature ones [94].
But then, when considering iPS-derived hepatocytes, for example, the more differentiated they are, the better it is? All these questions are still without an answer. The risk assessments for liver transplantation of each stem cell type should be evaluated in clinical studies due to the fact that the isolation procedures as well as culture conditions can alter clinical efficacy and safety.
Future research should also address the issue of time. The time taken to isolate, expand, and prime patient-specific stem cells into hepatocytes should be greatly reduced so that autologous transplantation can be carried out in a timely manner, especially for acute hepatic diseases. Whether autologous stem cell therapy will be applicable in case of human fulminant hepatic failure, in which rapid deterioration of the liver function occurs and immediate cure is required, remains a matter of concern. Moreover, following transplantation, stem cell-derived hepatocytes encounter a complex and hostile environment in which local hypoxia, oxidative stress, and inflammation may induce cell death. Insufficient retention and survival of transplanted cells may drastically dampen their therapeutic effects.
Thus, tissue engineering approaches (eg, tissue scaffolds) should be ameliorated, and the addition of adjuncts that increase the proliferation as well as differentiation of stem cells into not only hepatocytes but also all the other cell types making up the liver such as sinusoid endothelial, Kupffer, stellate, and biliary epithelial cells may help in this regard [40].
An important issue is the fate of the stem cell-derived hepatocytes once injected in the patients through different routes mentioned above. Do these cells migrate to the liver? Do they complete their differentiation or are eliminated after some permanence in the liver? Another still unanswered question is whether the bone marrow-injected stem cells infused into patients really differentiate into hepatocytes or fuse with pre-existing ones or remain unaltered and release paracrine factors, such as growth factors, angiogenic factors, cytokines, and microvesicles, among others, to enhance regeneration through the stimulation of resident parenchymal or progenitor cells.
Moreover, while in preclinical models, it is possible to assess engraftment and colonization efficiency of transplanted cells by an array of cell tracking methods (fluorescent dyes, magnetic resonance imaging) available, it is often technically difficult to perform on the long term in patients in the clinical setting. Couto et al. employed 99mTc-SnCl2 specific for mononuclear cell labeling to gain knowledge on the biodistribution of peripherally infused bone marrow mononuclear cells in cirrhotic patients [95]. Interestingly, the authors found that ∼41% and 32% of the radioactivity, as revealed by whole-body scintigraphy, was retained in the liver 3 and 24 h after injection, respectively. Cell tracking, in this case, helped in assessing the correct homing of the injected cells to the liver at early time points, but could not be used to assess the colonization efficiency months after cell injection.
Only one infusion of cells is temporarily beneficial. Repeated injections are probably needed to permanently reverse the damage [95]. For instance, in a Phase II study, patients with complex perianal fistulas were randomly assigned to intralesional treatment with fibrin glue or fibrin glue plus 20 million ASCs [96]. In the absence of healing, a second dose of fibrin glue and 40 million ASCs were administered to the patients, and this enhanced the probability of healing by more than four times in patients treated with ASCs plus fibrin glue than in patients treated with fibrin glue alone. A retrospective study performed on these patients showed the long-term safety of the treatment, despite the fact that a low proportion of patients treated with stem cells with closure after the procedure had no recurrence after 3 years' follow-up [97].
Several studies have shown the long-term rescue of liver diseases using primary hepatocyte transplantation [98,99]. That hepatocytes derived from all different adult stem cell types can efficiently engraft and restore liver function in the long-term still need detailed investigations [100]. Large, randomized, controlled clinical trials are required to get robust data on the benefit of adult stem cell transplantation in liver diseases and to tailor treatment to individual patients, thus reducing the risk of rejection [44]. Despite the fact that several studies using stem cells in the treatment of liver diseases have shown beneficial effects, the underlying mechanisms responsible for their therapeutic effects are still unknown. Moreover, the percentage of liver colonization is still very low in spite of arduous efforts to ameliorate cell engraftment. The best application route should be investigated for each adult stem cell type used to maximize engraftment.
Transplanted stem cell-derived hepatocytes secrete factors that contribute to liver repair and regeneration, but which recovery processes are regulated by these soluble factors remain undetermined.
The bone marrow contributes functionally and significantly to liver fibrosis and is a potential therapeutic target in liver fibrosis [49]. However, there are controversies on this subject that still have to be dealt with. MSCs from bone marrow have also been reported to inhibit hepatic stellate cell activation or induce their apoptosis, particularly through paracrine mechanisms, and therefore may be considered in the treatment of liver fibrosis [101]. Clinical studies on bone marrow cell therapy for liver regeneration and repair should thus proceed cautiously while evaluating the risk of hepatic fibrosis development [102].
In case of liver cancer, stem cells may create an immunosuppressive environment by providing antiaging and antioxidative effects, for example, by reducing reactive oxygen species, and hence favoring the escape of tumor cells and promoting metastasis formation [103]. When performing therapy with stem cells, one should not lose sight of the possible implication of these cells in carcinogenesis (reviewed in O'Connor et al. [104]). In the liver, several cell types (hepatocytes, cholangiocytes, progenitor cells) live long enough to be the cell of origin of cancer [105]. In fact, the presence of CK7 and CK19-positive progenitor cells has been found in a substantial number of hepatocellular carcinoma cases [106]. Hepatocellular carcinoma may also originate from the transdifferentiation of bone marrow cells, as also happens in other sites of the digestive tract [107,108]. These results suggest that there may be more than one type of carcinogen target cell in the liver.
Conclusions
Adult stem cells have been isolated from diverse body tissues and their capacity to differentiate into hepatocyte-like cells in vitro and restore liver functions in vivo has been investigated extensively, as depicted in Fig. 2. Now, the major goal is to translate the knowledge gained on animal models into the clinic, which is awaiting the perfect stem cell(s) to reduce death caused by acute and chronic liver diseases, often not curable by pharmacological approach. Hepatocyte infusion and stem cell-derived hepatocyte transplantation performed in humans are till now only bridging therapies to orthotopic liver transplantation, and much improvement is required to fulfill the main aim of cell transplantation—of partially replacing the missing function without having to undergo liver transplantation [13].

Different sources of adult stem cells for differentiation into hepatic-like cells. Mesenchymal stem cells, isolated from different tissues as adipose tissue, bone marrow, and liver, can be differentiated in hepatocyte-like cells. Hepatic stem cells as well as pluripotent stem cells derived from spermatogonial stem cells or reprogrammed somatic cells can also be induced to differentiate into hepatocyte-like cells. These adult stem cell-derived hepatocytes can be used for liver disease treatment as genetic disease, acute liver failure, chronic disease, and liver cancer. iPS, induced pluripotent stem.
Bridging therapy is very important in the case of pediatric liver diseases, especially because it helps in extending the life span of children until liver transplantation can be safely performed. More preclinical studies regarding the long-term engraftment and hepatocyte repopulation are necessary, and more clinical data as well as controlled clinical experimentation are required not only with regard to dosage, application route, and therapeutic mechanisms involved but also on the potential bidirectional role of the stem cells in carcinogenesis before firm conclusions can be drawn on the benefit of stem cells in acute or chronic liver diseases.
Footnotes
Acknowledgments
The authors would like to thank Victor Navarro-Tableros for the photograph of decellularized scaffold and Rinaldo Pellicano for critical reading of the manuscript.
F.A. received grants from Progetti di ricerca di interesse nazionale (PRIN) 2010, Telethon 2014, Prometeo, and Advanced Life Science in Italy (Alisei).
Author Disclosure Statement
No competing financial interests exist.