
Abstract
Mesenchymal stem cells (MSCs) from rodents and humans have been shown to suppress T cells by distinct primary pathways, with nitric oxide (NO)-dependent pathways dominating in rodents and indoleamine 2,3-deoxygenase (IDO)-dependent pathways dominating in humans. However, the immune suppressive pathways utilized by canine MSC have not been thoroughly studied, nor have bone marrow-derived MSC (BM-MSC) and adipose-derived MSC (Ad-MSC) been directly compared for their immune modulatory potency or pathway utilization. Therefore, canine BM-MSC and Ad-MSC were generated in vitro and their potency in suppressing T cell proliferation and cytokine production was compared, and differential gene expression. Mechanisms of T cells suppression were also investigated for both MSC types. We found that BM-MSC and Ad-MSC were roughly equivalent in terms of their ability to suppress T cell activation. However, the two MSC types used both shared and distinct biochemical pathways to suppress T cell activation. Ad-MSC utilized TGF-β signaling pathways and adenosine signaling to suppress T cell activation, whereas BM-MSC used cyclooxygenase, TGF-β and adenosine signaling pathways to suppress T cell activation. These results indicate that canine MSC are distinct from human and rodent MSC terms of their immune suppressive pathways, relying primarily on cyclooxygenase and TGF-β pathways for T cell suppression, rather than on NO or IDO-mediated pathways.
Introduction
C
MSCs interact with both the innate and adaptive immune systems, generally leading to abatement of ongoing inflammatory responses, though in some cases MSC may also upregulate immune responses [6,7]. The immune modulatory properties of MSC have been employed extensively for suppression of inflammation in a number of different immune-mediated, inflammatory disease models in rodents [8]. For example, MSC derived from adipose tissues or bone marrow have been used to treat experimental allergic encephalitis, inflammatory bowel disease, immune-mediated arthritis, airway inflammation, and graft-versus-host disease in rodent models [1,9 –12]. In addition, human MSC have been administered to rodent models of inflammatory diseases [13 –17]. Thus, it is apparent that the immune modulatory properties of MSC can be utilized therapeutically in a number of different diseases settings.
Numerous studies have investigated the underlying mechanisms that drive the immune modulatory properties of both human and mouse MSC [16,18 –20]. In human MSC, the reported pathways of immune suppression by MSC include the indoleamine 2,3-deoxygenase (IDO) [21,22], cyclooxygenase [23 –25], TGF-β [26], soluble IL-1Ra [27,28], soluble MHC [29,30], and the PD-L1 pathways [31,32]. In general however, the IDO-dependent pathway is considered the primary mechanism of human MSC suppression of activated immune effector cells [21]. For rodent MSC, the nitric oxide (NO)-dependent immune suppressive pathways predominate [33], though other pathways including TGF-β and IL-10 have been reported [34]. It is also important to note that these immune modulatory pathways typically only become operative after the MSC have first been activated immunologically, typically by proinflammatory cytokines such as IFN-γ, TNF-α, IL-1β, or IL-17, or by certain TLR (Toll-like receptor) ligands [35 –37]. Thus, in most assays used to evaluate MSC immune suppressive pathways, activated T cells are cocultured with MSC, and in these assays the activated T cells provide the source of MSC-activating cytokines. In studies with suppression of innate immune effector cells such as dendritic cell (DC), the requirement for addition of MSC-activating cytokines such as IFN-γ is more apparent [38 –40].
MSCs derived from bone marrow or adipose tissues of dogs also exhibit immune modulatory activity. For example, it was reported recently that intrathecal injection of MSC in dogs with idiopathic meningoencephalitis resulted in clinical improvement [41]. Local injection of MSC has also been used in the management of keratoconjunctivitis sicca in dogs, an immune-mediated disorder that results in loss of tear production [42]. Human embryonic stem cell-derived MSC were also reported recently to suppress inflammation associated with furunculosis in dogs, an inflammatory disease of the peri-rectal tissues of canines [43]. MSCs have also been widely used for treatment of osteoarthritis in dogs, a progressive degenerative condition that also has an associated inflammatory component. Studies of canine adipose-derived MSC (Ad-MSC) and bone marrow-derived MSC (BM-MSC) injected into the elbow and hip joints of dogs with osteoarthritis produced positive results in terms of improvement in clinical measures of osteoarthritis reported in most investigations [44 –49].
Previously, it was reported that various different sources of canine MSC modulate immune responses in vitro [12,50 –54]. In the study by Lee et al., which utilized canine bone marrow MSC, PGE2 was identified as the primary MSC factor that suppressed T cell proliferation [12,54]. Kang et al. identified PGE2 as an important T cell suppressive factor secreted by canine Ad-MSC, and also found that IDO production played an important role in T cell suppression [50]. Thus, the cyclooxygenase pathway appears to be a major pathway in canine MSC suppression, though relatively few other pathways have been investigated.
Therefore, we sought in this study to better define the functional immune modulatory properties of canine MSC, and to determine whether there were important differences between Ad-MSC and BM-MSC in terms of either their potency or the pathways utilized for T cell suppression. To address these questions, Ad-MSC and BM-MSC were generated and their immune modulatory properties evaluated using in vitro assays and T cells obtained from unrelated animals. These studies revealed that canine Ad-MSC and BM-MSC were similar in terms of their surface phenotype and overall immune modulatory potency. However, several important differences in the pathways of immune suppression utilized by each cell type were discovered. In addition, our results indicated that canine MSC did not utilize either the IDO or NO pathways as their predominant mechanisms for T cell suppression. Thus, canine MSC appear to be distinct from human and rodent MSC in terms of their utilization of major immune suppression pathways.
Materials and Methods
Culture medium for MSC and T cells
Canine bone marrow and adipose derived MSC were maintained in MSC media. MSC media contained DMEM (Dulbecco's modified Eagle's medium) low glucose supplemented with essential and nonessential amino acids, Glutamax 1% penicillin-streptomycin (all from Life Technologies Corp., Grand Island, NY) and 10% fetal bovine serum (FBS; VWR Life Science, Radnor, PA). Canine T cells were cultured in the same media with the addition of 55 μM 2-mercaptoethanol (Life Technologies Corp.).
Biochemical reagents
Aminoguanidine and meclofenamic acid (MFA) were purchased from Sigma-Aldrich (St. Louis MO), L-NMMA acetate, indomethacin, SB431542, LY364947, 1-MT (1-methyl-
Generation of canine Ad-MSC and BM-MSC
All procedures involving live animals were approved by the Institutional Animal Care and Use Committee at Colorado State University. Adipose and bone marrow were each collected from three unrelated, healthy adult dogs. Canine adipose tissue (0.5 g) was collected from using an 8 mm skin punch biopsy instrument, following sterile prep of the skin surface. The collected tissue was washed with sterile phosphate-buffered saline (PBS) and then cut into small pieces with scalpel blades. The minced tissues were then placed into a 50 mL conical and digested with 1 mg/mL of collagenase type I (Sigma-Aldrich), for 30 min at 37°C. After 30 min of enzymatic digestion, MSC complete growth medium was added to inactivate collagenase. The digested cells were then collected by centrifugation and plated in T75 cell culture flask (Corning, Inc., Corning, NY) and allowed to adhere for 72 h. All nonadherent cells were then removed at the first medium change at 72 h and the adherent cells were refed with media. Media was then changed every 72 h thereafter, and cells were passaged by trypsinization once Ad-MSC reached ∼80% confluence.
Bone marrow aspirates were collected by bone marrow needles from the proximal humerus of dogs that were anesthetized or heavily sedated. The bone marrow samples were further minced in vitro and washed twice with PBS. Bone marrow samples were then placed into 50 mL conical tubes and digested with 1 mg/mL of collagenase type 1 for 45 min at 37°C. The single cell suspensions were pelleted after digestion by centrifugation, washed twice in PBS, then resuspended in complete medium and plated in T-75 flasks with complete MSC medium and allowed to adhere for 72 h. All nonadherent cells were then removed at the first medium change at 72 h and adherent cells were refed with medium. Medium was then changed every 72 h thereafter, and cells were passaged by trypsinization once BM-MSC reached ∼80% confluence.
Tri-lineage differentiation
Tri-lineage differentiation for both Ad-MSC and BM-MSC was performed between passages 2 and 3, using the StemPro Adipogenesis Differentiation Kit, the Chondrogenesis Differentiation Kit, and the Osteogenesis Differentiation Kit (Life Technologies Corp.), according to manufacturer's instructions. At the completion of the differentiation protocol for each cell lineage, cells were incubated with appropriate stains and photographed, using an Olympus CKX41 light microscope and attached SC30 digital camera. Images were captured using getIT software v5.2 (Olympus Soft Imaging Solutions).
MSC and T cell coculture assays
Canine peripheral blood mononuclear cells (PBMC) were prepared from EDTA-anticoagulated whole blood collected from the jugular vein of dogs. To prepare PBMC, anticoagulated blood was separated over a Ficoll gradient using lymphocyte separation medium (MP Biomedicals, Inc., Santa Ana, CA), according to manufacturer's instructions. To assess T cell proliferation, the purified PBMC were labeled with carboxyfluorescein succinimidyl ester (CFSE; ThermoFisher, Waltham, MA), according to manufacturer's directions. The CFSE stained cells were then plated in triplicate wells of a 96-well, flat bottom plate in 100 μL MSC medium containing 55 μM β-mercaptoethanol at a density of 5 × 105 cells per well. Activation of T cell proliferation was initiated by addition of Concavalin A (ConA; Sigma-Aldrich) at a concentration of 10 μg/mL. The effects of Ad-MSC and BM-MSC on T cell proliferation and cytokine production were assessed by adding unlabeled Ad-MSC or BM-MSC to PBMC cultures at ratios of 1:10 (MSC:PBMC), in a final volume of 200 μL per well of triplicate wells of 96-well plates. Cocultures of MSC and T cells in contact were maintained for 96 h, at which point the supernatant was collected (and stored frozen) for cytokine analysis, and the nonadherent cells were collected for flow cytometric analysis (see “T cell proliferation analysis” and “Flow cytometry” sections).
T cell proliferation analysis
T cell proliferation with the addition of inhibitors was analyzed using EDU (5-ethynyl-2′-deoxyuridine) staining. EDU (Life Technologies Corp.) 3 μM was added to coculture at 48 h after plating. At 96 h nonadherent cells were collected and surfaced stained for detection of canine CD5+ cells, using an anti canine CD5-Alexa Fluor®488 (clone YKIX322.3; ABD Serotech, Raleigh, NC). After surface staining, cells were fixed with 4% PFA (Affymetrix, Inc., Cleveland, OH) and permeabilized with saponin-based permeabilization and wash reagent (Life Technologies Corp.). Fifty microliters of reaction cocktail was mixed in PBS containing copper(II) sulfate pentahydrate (CuSO4·5H2O) 1 mM and
Flow cytometry
For analysis of the surface phenotype of Ad-MSC and BM-MSC, cells were harvested by trypsinization and resuspended at a concentration of 1 × 106 cell/mL in FACS buffer (PBS, 2% FBS, and 0.1% sodium azide) on ice. Cells (1 × 105/well) were immunostained in 96-well polystyrene round bottom plates. Nonspecific staining was blocked with normal dog serum, MSC were then incubated with primary antibodies for 30 min at room temperature. The primary antibodies used in these studies were as follows: CD34-FITC (clone 1H6; ABD Serotech), CD44-FITC (clone 1M7; eBioscience, Inc., San Diego, CA), CD45-FITC (clone YKIX716.13; ABD Serotech), CD73-biotin (clone TY/11.8; Biolegend, San Diego, CA), CD90-APC (clone YKIX337.217; eBioscience, Inc.), and unconjugated CD105 (clone 8A1; Abcam, Cambridge, MA). After primary antibody incubation, cells were washed with FACS buffer, then analyzed using a Beckman-Coulter Gallios flow cytometer. Samples stained with anti CD105 were incubated with additional donkey anti mouse IgG–FITC (Jackson ImmunoResearch Laboratories, Inc., West Grove, PA). Data analysis was done using FlowJo 9.0.8 software. In histogram plots of MSC phenotype (Fig. 1), the x-axis represented florescence intensity and the y-axis represented cell count. dark gray histograms depict isotype staining, while light gray histograms represent staining intensity using the specific antibody.
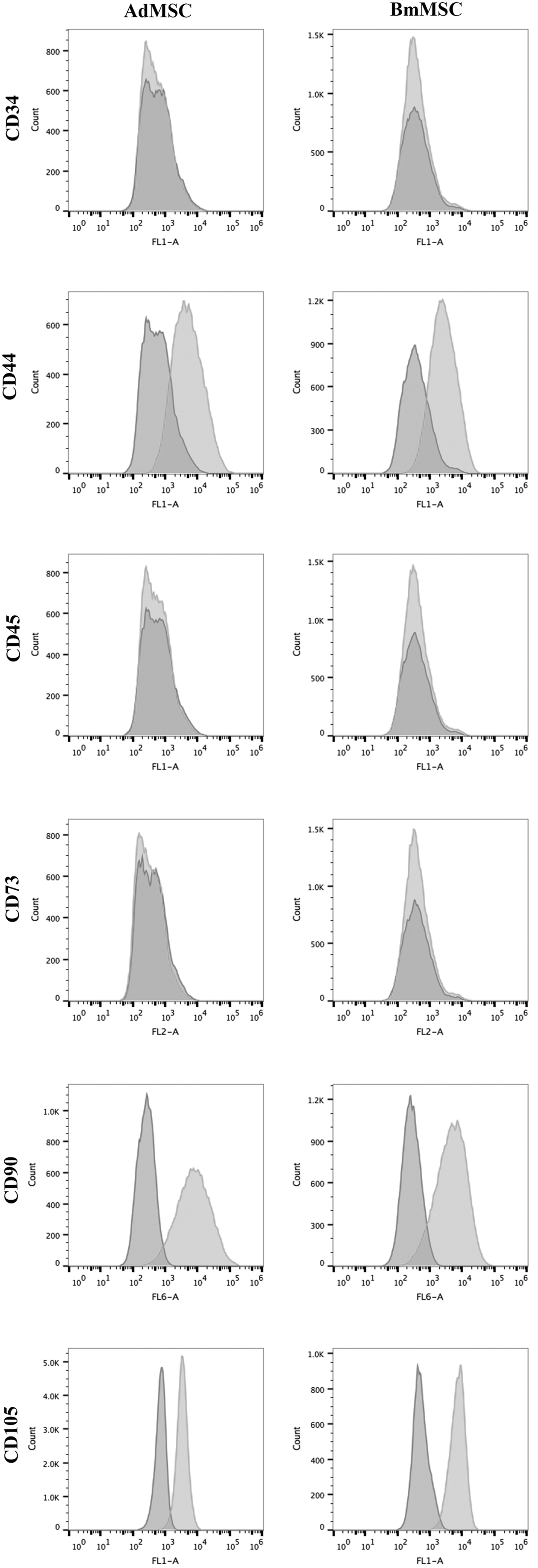
Immunophenotypic characterization of canine Ad-MSC and BM-MSC by flow cytometry. Canine Ad-MSC (left column) and BM-MSC (right column) at passage 2–3 were immunostained with antibodies to stem cell surface markers CD34, CD44, CD45, CD73, CD90 (or the intracellular marker CD105), and analyzed by flow cytometry using protocols described in the Materials and Methods section. Histograms depict fluorescence intensity on the x-axis and cell count on the y-axis. Dark gray histograms depict isotype antibody staining, while light gray histograms depict staining with relevant, directly conjugated primary mAbs. Each study was repeated at least twice with similar results, including studies with MSC from several unrelated animals. Ad-MSC, adipose-derived mesenchymal stem cell; BM-MSC, bone marrow-derived mesenchymal stem cell.
For analysis of T cell proliferation, nonadherent CFSE-labeled cells were collected from each well at 96 h, and then immunostained for detection of canine CD5+ cells, using an anti canine CD5-APC (clone YKIX322.3; ABD Serotech). After 30 min incubation with CD5 antibody, the cells were washed with FACS buffer, resuspended in 200 μL, and then analyzed by flow cytometry. Histograms were generated using FlowJo 9.0.8 software. The CD5+ population of cells was first gated, and then the proliferation percent measured as the frequency of dividing cells that had reduced CFSE florescence compared to unstimulated CFSE-labeled CD5+ T cells. T cell proliferation index was calculated as the proliferation percentage normalized to the value for stimulated PBMC+MSC, which was assigned a value of 1.
IFN-γ enzyme-linked immunosorbent assay
Supernatants were collected from 96 h cocultures of T cells and MSC and stored at −20°C before analysis. Concentrations of IFN-γ in supernatants was determined using a canine IFN-γ specific enzyme-linked immunosorbent assay (ELISA; R&D Systems, Minneapolis, MN), according to the manufacturer's directions. A cytokine stimulation index was calculated as the IFN-γ concentration (pg/mL) from the test sample normalized to the mean IFN-γ concentration present in supernatants from activated PBMC cocultured with MSC.
Neutralization of IFN-γ
IFN-γ present in T cell and MSC cocultures was neutralized using an anti-canine IFN-γ antibody (R&D Systems) at a concentration 10 ng/mL. At 48 h, an additional 5 ng/mL of the anti-IFN-γ antibody was added to cocultures to assure complete neutralization. T cell proliferation was analyzed as previously described above.
Immunocytochemistry
Expression of intracellular stem cell antigens by Ad-MSC and BM-MSC was determined by plating cells on glass coverslips in 24-well cell culture plates and allowing the cells to adhere for 24 h. Cells were fixed with 4% PFA for 10 min at room temperature, washed in PBS, and permeabilized with 0.1% Triton-X. Nonspecific binding was minimized by incubation with 10% secondary antibody species serum plus 0.1% Triton for 1 h before application of primary antibody. Each well was incubated with primary antibody overnight at 4°C. Primary antibodies used are as follows: Oct3/4 (clone H134; Santa Cruz Biotechnology, Inc.) and vimentin (clone V9; Merck Millipore, Billerica, MA). Corresponding rabbit and mouse IgG irrelevant isotype antibodies were used at concentrations matching the primary antibodies (eBioscience, Inc.). Cells were washed with PBS with 0.05% Tween and then incubated with secondary antibodies (donkey anti mouse IgG or donkey anti rabbit IgG; Jackson ImmunoResearch Laboratories, Inc.), then washed and mounted with DAPI counter stain. Visualization of florescence staining was performed using an Olympus IX83 confocal microscope. Images were imported as Tiff files to Photoshop CC 2015, and adjusted with high definition resolution (HDR) toning. For each antibody, adjusted HDR toning was saved as preset values and applied to corresponding isotype control stains as well.
T cell apoptosis and cell death measurement in cocultures
Cocultures of MSC and PBMC were prepared as noted above, except that the activated T cells were not labeled with CFSE. After culture for 24, 48, 72, and 96 h, nonadherent cells (primarily CD5+ T cells) were collected and prepared for analysis of apoptosis and cell death. Cells were immunostained for CD5 (eBiosciences), and for Annexin V expression (V405 Annexin V; Life Technologies Corp.). Assessment of cell death was done by adding 7-AAD viability stain (eBiosciences, Inc.) to cells immediately before flow cytometric analysis. Histograms were generated with Flowjo 9.0.8 software, and CD5+ cells were evaluated for expression of Annexin V and 7-AAD.
Comparison of gene expression profiles by microarray analysis
Triplicate, independent cultures of Ad-MSC and BM-MSC were established as previously described above and cellular RNA collected from semi-confluent cultures using Qiagen RNeasy mini kit. The RNA was then labeled and hybridized to Canine Gene 1.0 ST Arrays (Affymetrix, Santa Clara, CA) using standard Affymetrix protocols. Image files were converted to log2 expression values with Robust Multiarray Average background correction and quantile normalization, as implemented by a statistical/visualization package (Partek Genomics Suite v6.6, St Louis, MO). The normalized expression values used in the statistical and bioinformatics analysis for these studies, and the original raw visual data used to calculate these values, have been deposited in the publicly accessible database Gene Expression Omnibus (
The Ingenuity Pathway Analysis (IPA) platform (Ingenuity, Redwood City, CA) was used to perform systems analysis on the top 5% of genes that were significantly differentially expressed between Ad-MSC and BM-MSC. The IPA software uses Fisher's exact test to identify over-represented and connected biological units in a defined set of genes, which can include pathways, cellular functions, or known targets of regulatory genes. In some cases, a confidence or z score was generated based on the activation state of the pathway (or upstream regulators of the pathway) or based on the expression pattern of the associated genes. The IPA software was also used to group transcripts into gene-limited networks (containing 35 genes at most) based on evidence of direct or indirect relationships between molecules according to the IPA Knowledge Base.
Statistical analysis
Statistical comparisons between data sets with two treatment groups were done using an unpaired t-test. Comparisons between three or more groups were done using ANOVA, followed by Tukey multiple means comparison post-test. Analyses were done using Prism 6 software (GraphPad, La Jolla, CA). For all analyses, statistical significance was determined for P < 0.05.
Results
Phenotypic comparison of Ad-MSC and BM-MSC
The surface phenotypes of canine Ad-MSC and BM-MSC were compared using flow cytometry to quantitate expression of stem cell surface markers, as reported in earlier studies of canine MSC [12,50,55]. We found that both types of MSC were very similar in terms of their surface phenotypes, inasmuch as each cell type expressed high levels of CD44, CD90, and CD105, but were negative for expression of the hematopoietic stem cell antigens CD45 and CD34 (Fig. 1). Notably, neither MSC cell type expressed surface marker CD73, though this molecule has been reported to be expressed by canine MSC in other studies [50]. We did, however, observe CD73 expression using the same antibody by MSC derived from canine iPSC (Chow et al., manuscript in preparation), indicating that in some instances surface marker CD73 can be expressed by canine MSC. Ad-MSC and BM-MSC derived from two additional, unrelated dogs in the study displayed a very similar surface phenotype (data not shown).
MSC were also evaluated by immunocytological staining for expression of intracellular proteins associated with MSC in other species (Fig. 2). Both Ad-MSC and BM-MSC expressed high levels of the mesenchymal cytoskeletal intermediate filament protein vimentin (Fig. 2A, B). Both cell types were also positive for intra-nuclear expression of the nuclear transcription factor OCT3/4 (Fig. 2C, D). However, the two cell types did exhibit morphological differences, with Ad-MSC being more elongated, versus the flattened and oblong BM-MSC (not shown). These findings are consistent with the previously reported phenotypes of both human and rodent MSC [56,57].
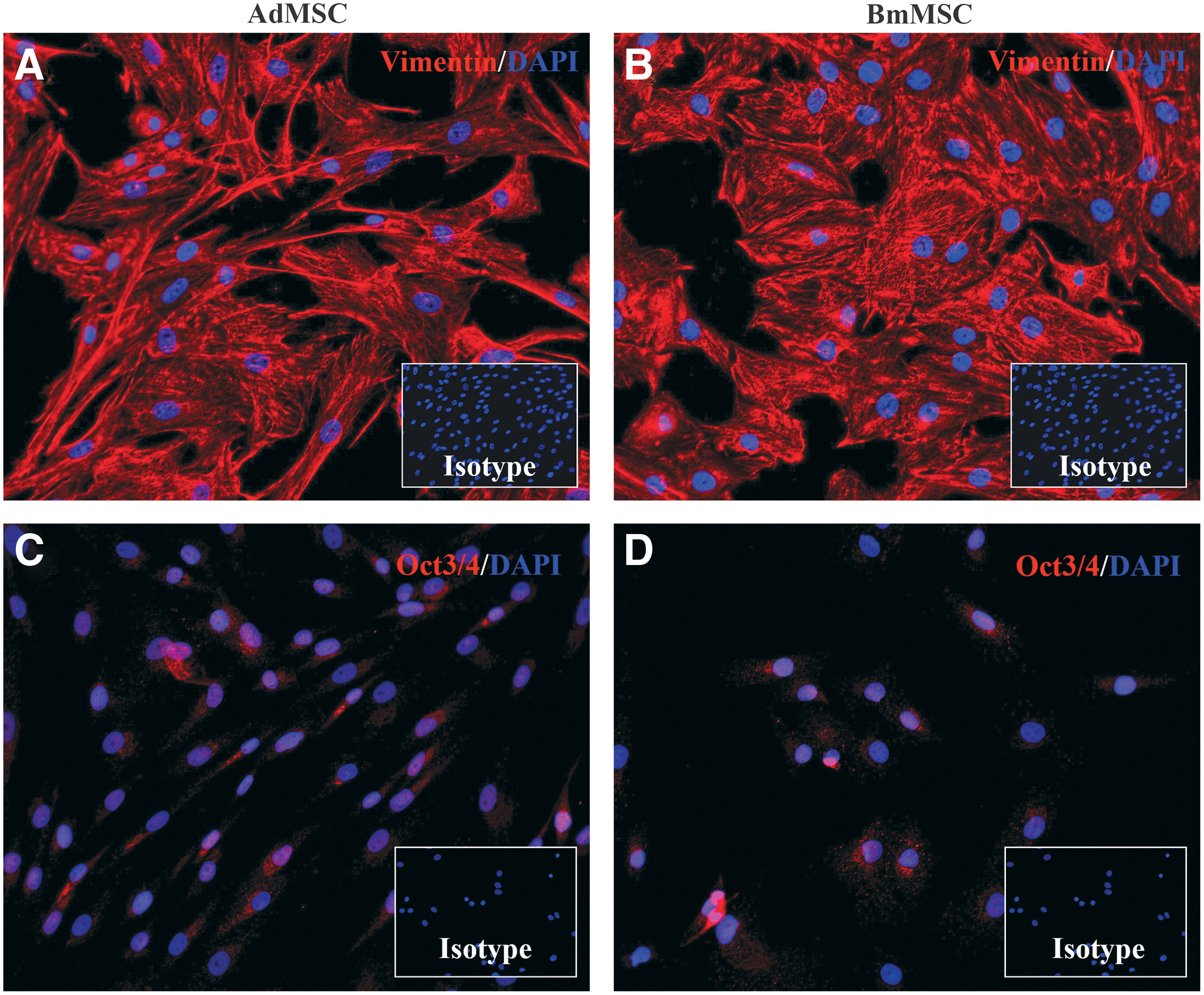
Immunocytological characterization of Ad-MSC and BM-MSC by immunofluorescence staining. MSC were cultured on glass slides and immunostained for detection of expression of intracellular stem cell antigens, as described in the Materials and Methods section. Slides were counterstained with DAPI for nuclear detection. Inset boxes depict staining with isotype control antibodies. In
Tri-lineage differentiation
The ability of Ad-MSC and BM-MSC to undergo tri-lineage differentiation, an important property of MSC, was assessed next. Using standard differentiation induction conditions, Ad-MSC and BM-MSC both readily differentiated into cells with properties consistent with chondrocytes, adipocytes, and osteoblasts (data not shown). Thus, tri-lineage differentiation properties were equally shared by the two MSC types.
Suppression of T cell proliferation and cytokine release by Ad-MSC and BM-MSC
The immune modulatory properties of MSC have been widely described, both in vitro and in vivo, based in large part on assays that assess T cell suppression [8,58,59]. Therefore, we used PBMC obtained from healthy donor dogs (unrelated to the source dog for MSC cultures) and Ad-MSC and BM-MSC to assess and compare immune modulatory properties of each cell type. Triplicate wells of activated T cells were incubated with MSC at an MSC to PBMC ratio of 1:10, for 96 h. At the initiation of the assays, T cells were activated with ConA (10 μg/mL). To assess proliferation, PBMC were labeled with CFSE, as described in the Materials and Methods section (MSC were unlabeled in these assays). At 96 h, supernatants were collected for IFN-γ quantitation by ELISA, and T cell proliferation was analyzed by flow cytometry.
T cells were immunostained for CD5 expression and the dilution of CFSE staining intensity of CD5+ T cells was assessed as a measure of T cell proliferation (Fig. 3). Both Ad-MSC and BM-MSC significantly suppressed T cell proliferation when incubated with MSC (Fig. 3A–D). Ad-MSC suppressed proliferation by an average of 88%, compared to 86% suppression for BM-MSC (this difference was not statistically significant).
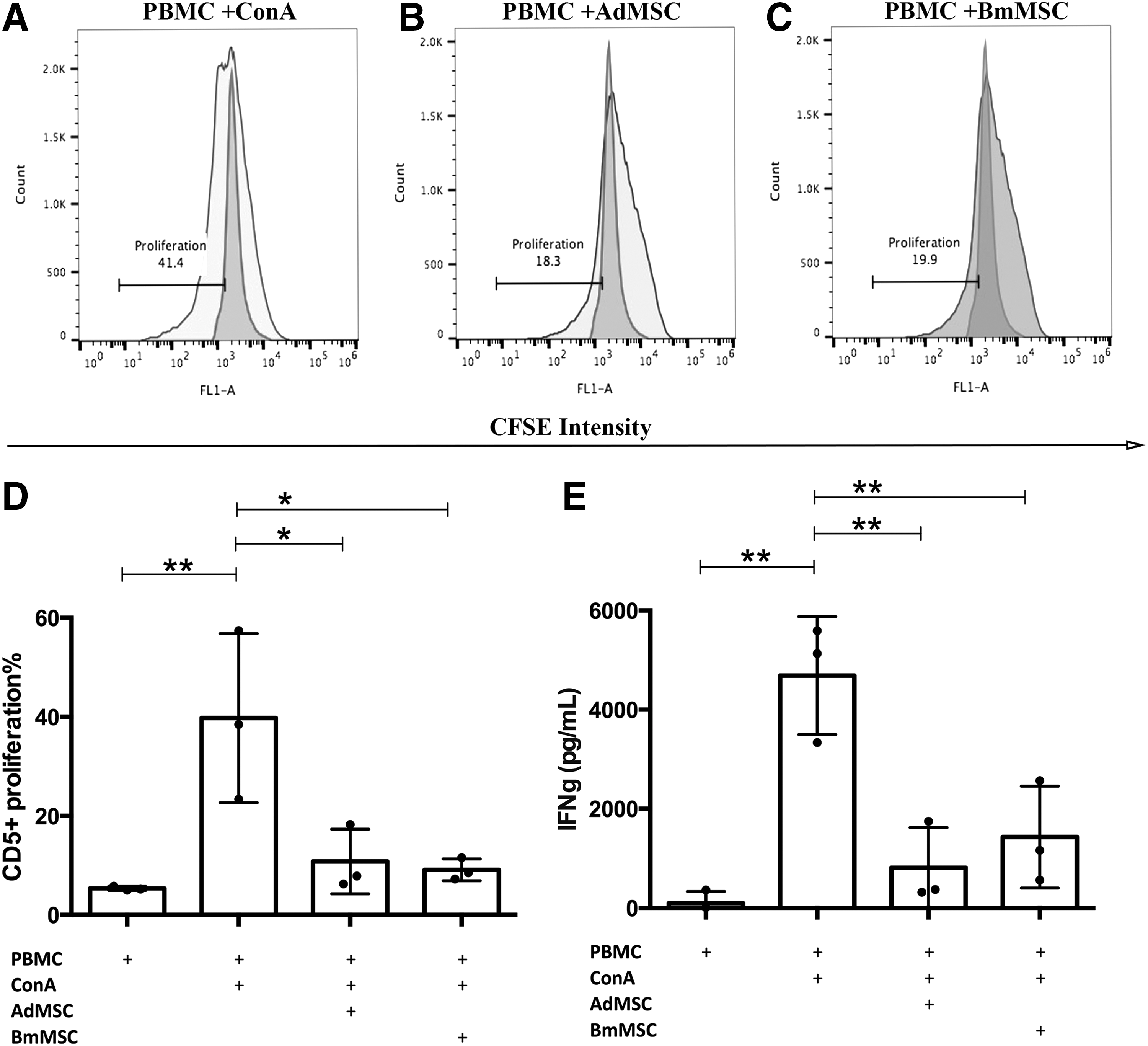
Effects of Ad-MSC and BM-MSC on T cell proliferation and IFN-γ production. PBMC from healthy donor dogs (n = 3) were activated with 10 μg/mL ConA and cocultured with MSC at a ratio of 1:10 (MSC:T cell) in triplicate wells of a 96-well plate for 96 h, followed by flow cytometric assessment of T cell proliferation, as described in the Materials and Methods section. In
In addition, coculture with Ad-MSC and BM-MSC also produced significant suppression of IFN-γ release by activated T cells (Fig. 3E), with 93% suppression of IFN-γ release induced by Ad-MSC and 86% suppression induced by BM-MSC (differences were not statistically significant). These results indicated that canine Ad-MSC and BM-MSC are both capable of inducing significant suppression of T cell function, at roughly equivalent potency.
Mechanisms of T cell suppression by MSC
Studies were conducted next to identify and compare mechanisms of T cell suppression induced by canine MSC (Fig. 4). These studies used both T cell proliferation and IFN-γ production as the primary read-outs (though IFN-γ release proved to be the more sensitive readout for these assays). Inhibitors of most of the major known pathways of MSC suppression of T cells in other species were evaluated, including the following inhibitors and pathways: NO pathway (aminoguanidine, L-NMMA); cyclooxygenase pathway (indomethacin, MFA); TGF-β pathway (SB431542, LY364947); IDO pathway (1-MT, CAY 10581); adenosine receptor pathway (ZM241385, CSC); and checkpoint molecule pathway (blocking antibodies to canine PD-L1). The doses of inhibitors used were determined in most cases by titration studies in our laboratory, or based on inhibitor concentrations used in previous publications.
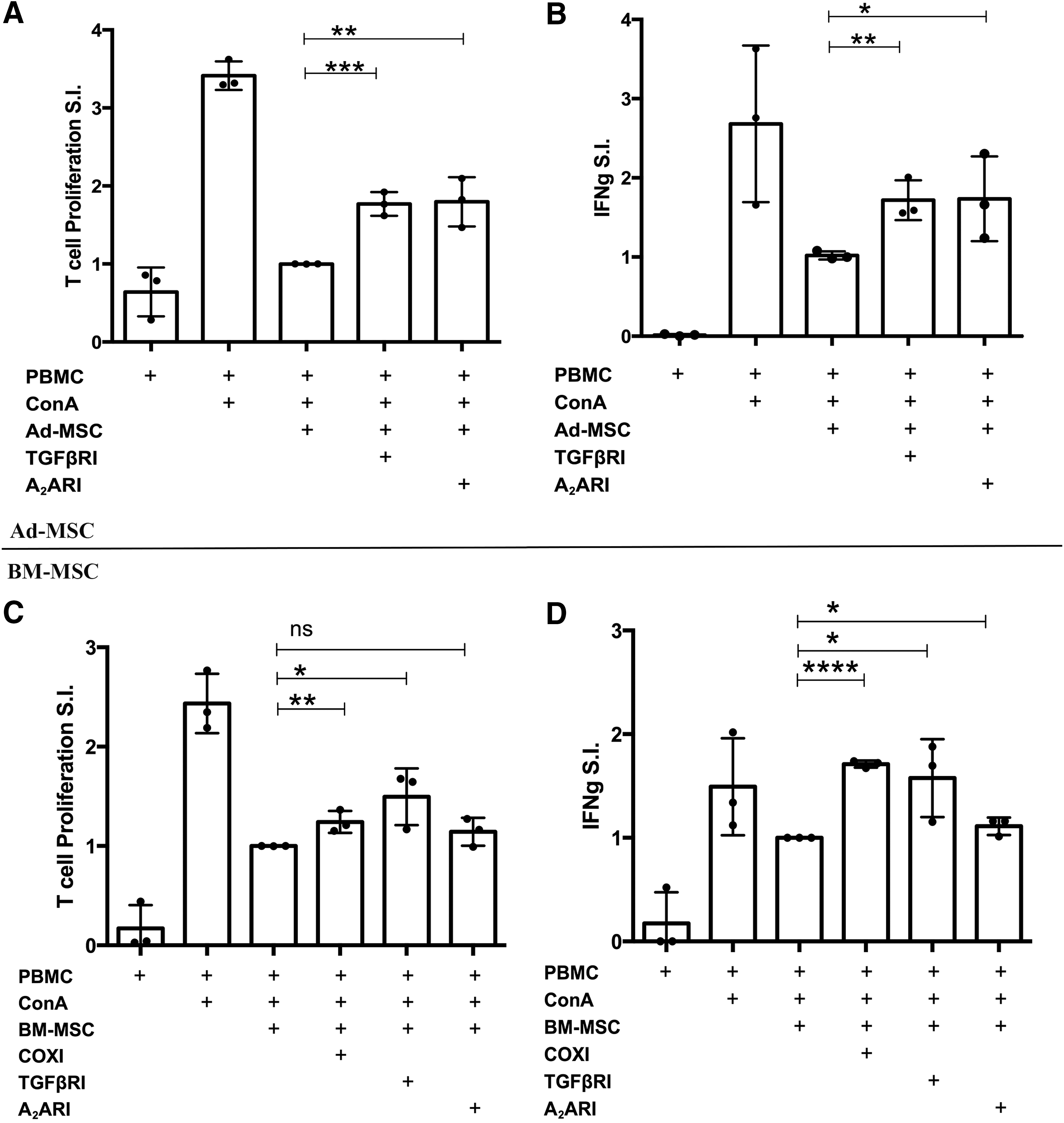
Identification of pathways regulating MSC suppression of T cell proliferation and cytokine production. Ad-MSC
We found that for suppression of T cell function by canine Ad-MSC, the predominant pathways utilized were the TGF-β pathway and the adenosine pathways, as revealed by significant reversal of inhibition of T cell proliferation and IFN-γ release following incubation with a TGF-β receptor inhibitor (SB431542) or with an adenosine receptor antagonist (ZM241385) (Fig. 4A, B). In contrast, blocking other signaling pathways using inhibitors or antibodies did not significantly reverse T cell suppression elicited by Ad-MSC (data not shown). TGF-β release in the range of 5–15 ng/mL was also detected in conditioned medium from T cells cocultured with Ad-MSC (data not shown). It was not possible, however, to measure adenosine concentrations directly.
For BM-MSC inhibition of T cells, the TGF-β and adenosine pathways were also found to be active (Fig. 4C, D). In addition, we also identified an important contribution from the cyclooxygenase pathway, inasmuch as significant reversal of suppression occurred following the addition of indomethacin or MFA to the BM-MSC and T cell cocultures (Fig. 4C, D). In addition, BM-MSC cocultured with T cells led to the secretion of high concentrations of PGE2 and TGF-β (data not shown). Thus, we concluded from these studies that canine Ad-MSC and BM-MSC use two overlapping immune modulatory pathways (TGF-β and adenosine signaling) for T cell suppression, while BM-MSC also utilize the cyclooxygenase pathway for T cell suppression.
MSC activation by IFN-γ required for full immune suppressive activity
Several studies have found that for MSC to suppress T cells, the MSC must first be activated by proinflammatory cytokines [18,19,23,60]. The major cytokine known to “license” MSC to become immune modulatory has been identified as IFN-γ [36,60]. Therefore, the role of IFN-γ in regulating the T cell suppressive effects of canine MSC was evaluated by neutralizing IFN-γ production during MSC: T cell cocultures (Fig. 5). These experiments revealed that the ability of MSC to suppress T cell proliferation was significantly attenuated when IFN-γ was neutralized at the initiation of the cocultures. Therefore, analogous to other species, IFN-γ produced by activated canine T cells also plays a critical role in the early activation of canine MSC, including both Ad-MSC and BM-MSC, resulting in an immunosuppressive phenotype.
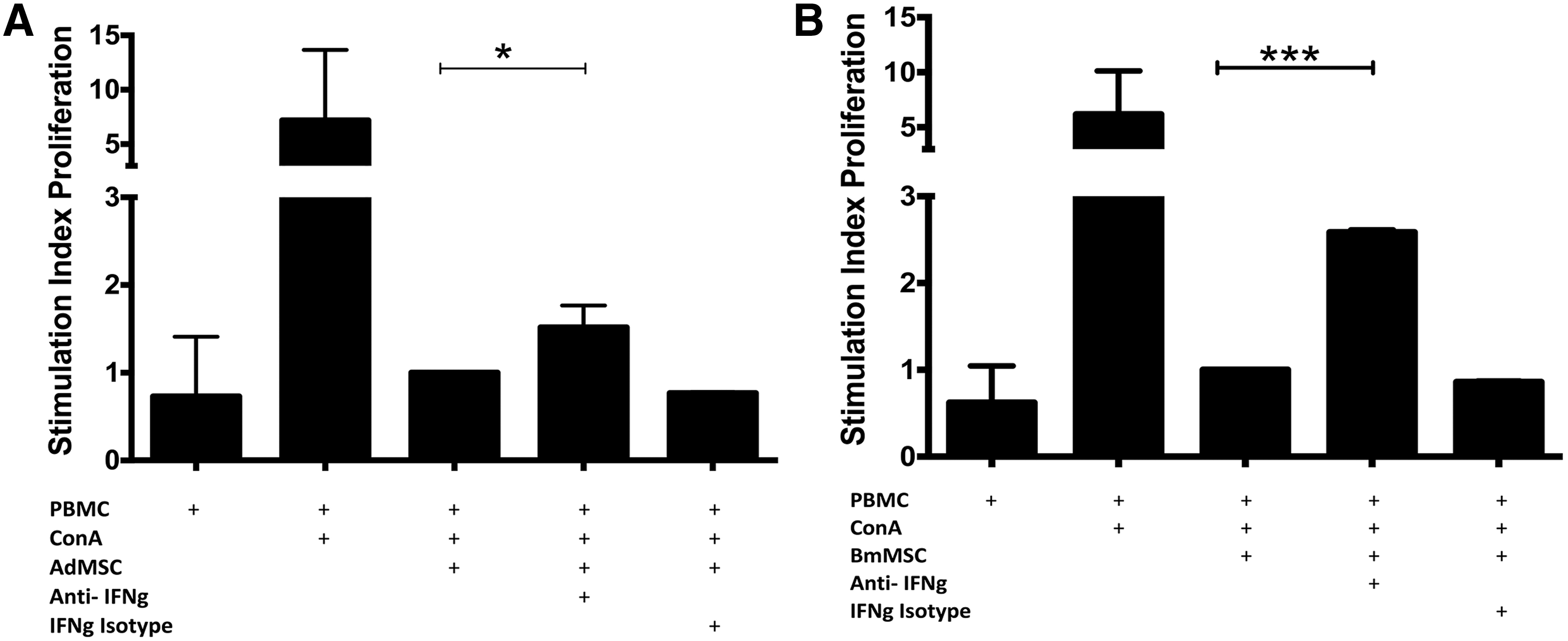
Effects of IFN-γ neutralization on MSC suppression of T cell proliferation. Cocultures of Ad-MSC or BM-MSC and activated T cells were incubated with anti-canine IFN-γ neutralizing antibody (10 μg/mL) added at the initiation of the coculture, with additional antibody added at 48 h at 5 μg/mL. The effects of IFN-γ neutralization on suppression of T cell proliferation by Ad-MSC
MSC induce T cell death in coculture
Previous reports have suggested that MSC may suppress the function of activated T cells in part by inducing T cell apoptosis [61 –63]. Therefore, the effects of addition of MSC on apoptosis of activated canine T cells were assessed in coculture assays, using flow cytometric expression of Annexin V combined with cell membrane permeability changes to detect early and late apoptotic and cell death events in T cells (Fig. 6). These studies revealed that the presence of MSC in cocultures with T cells did not significantly influence (either positively or negatively) the number of early or late apoptotic CD5+ T cells over a 96-h period. However, the numbers of dead T cells (7-AAD+CD5+) increased progressively in cocultures with both Ad-MSC and BM-MSC, compared to cultures of activated T cells alone, reaching the level of statistical significance at 96 h (Fig. 6C). For example, coculture with Ad-MSC increased the percentage of dead T cells to 20% (compared with 9% for activated T cell cultures alone), while coculture with BM-MSC increased the percentage of dead T cells to 29%. Thus, the presence of canine MSC can decrease the overall viability of activated T cells.
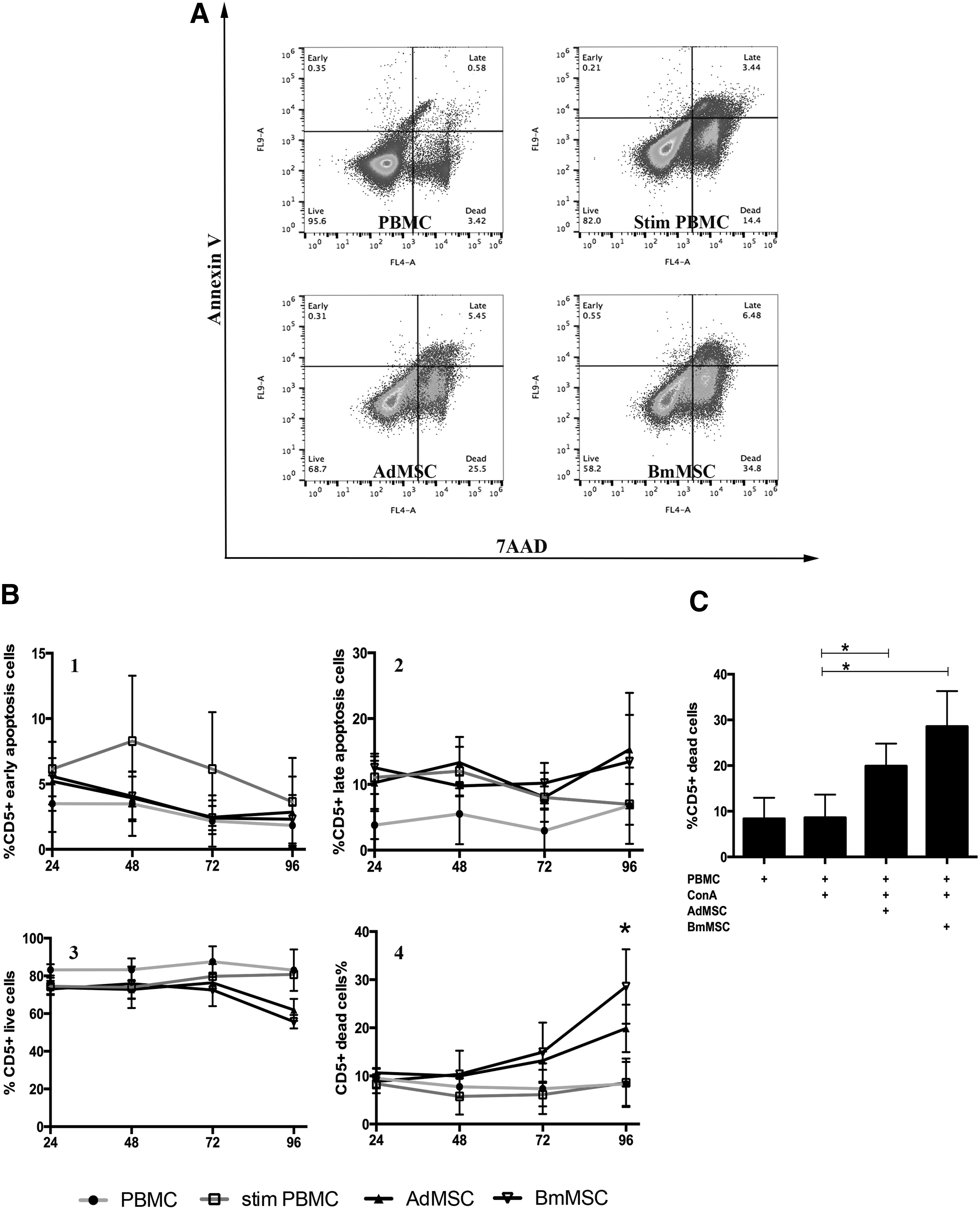
Coculture with MSC induces T cell death. Activated T cells alone, or T cells cocultured with MSC at a 1:10 ratio (MSC:T cells), were cultured in triplicate wells of 96-well plates as noted in the Materials and Methods section. At 24 h intervals, the nonadherent cells were collected and immunostained with anti-CD5 antibody, Annexin V for apoptosis detection, and 7-AAD for detection of dead cells. In
Gene expression patterns in Ad-MSC and BM-MSC
To compare the gene expression patterns of Ad-MSC and BM-MSC, microarray studies were done, using Affymetrix canine OST 1.0 chips. (The full array data have been deposited in the Gene Expression Omnibus database online.) Differential gene expression analysis showed that 698 genes were upregulated in Ad-MSC compared to BM-MSC, with a fold change >2 and P < 0.02. In addition, 695 genes were downregulated in Ad-MSC compared to BM-MSC, with a fold change of < −2 (raw data deposited in Gene Expression Omnibus under accession number (GSE90449). Principle component analysis (Fig. 7A) demonstrated that the biological replicates of the same cell type clustered together, within a standard deviation of 2, with over 74% variance accounted for by cell type. Over 60% of differentially expressed genes with a significance of P < 0.05 fell within a −2 to 2-fold difference (Fig. 7B). Using a complete list of immune function-related genes from the Immunology Database and Analysis Portal (ImmPort) system (
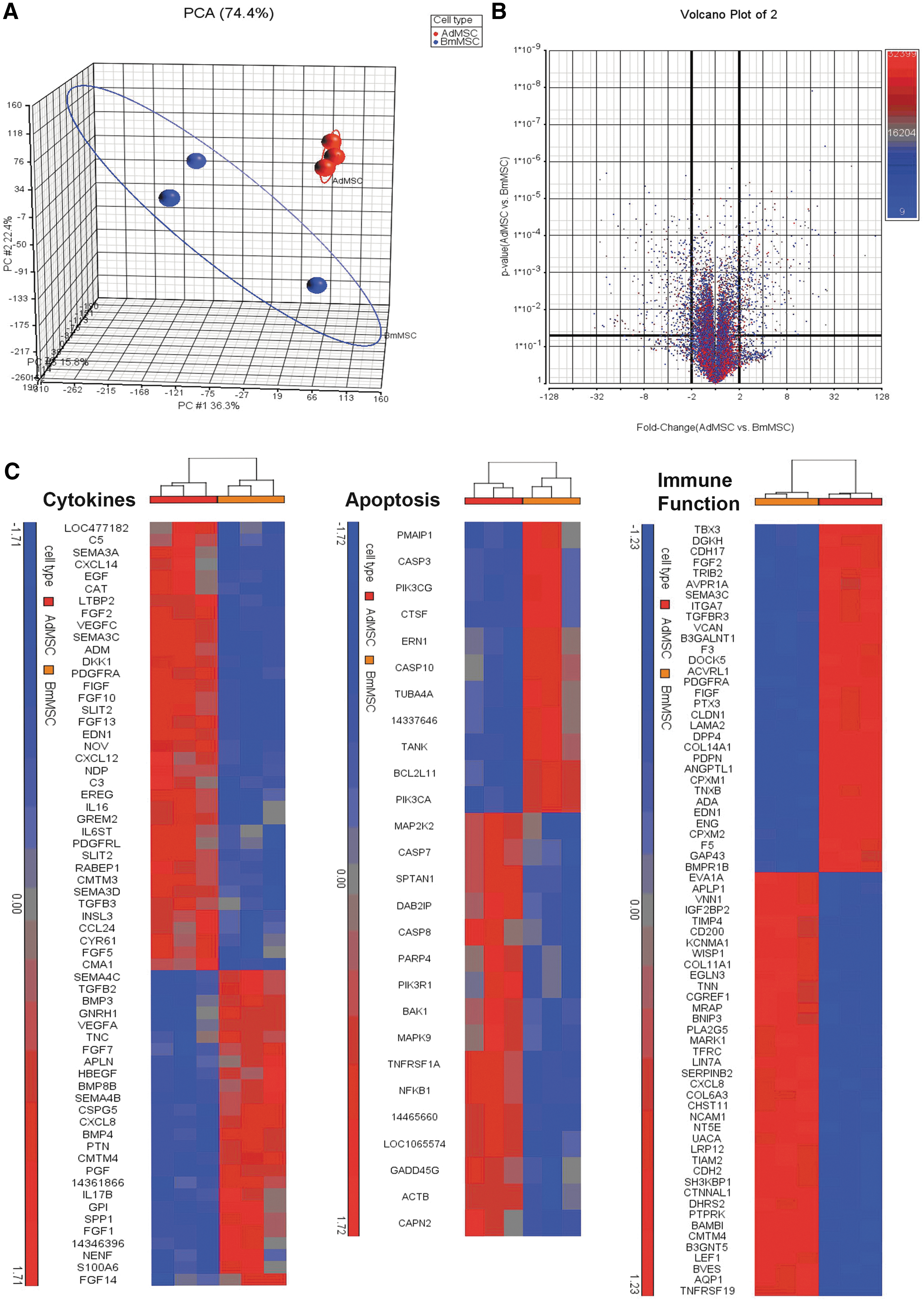
Differential gene expression by Ad-MSC versus BM-MSC. The Partek Genomics Suite was used to import microarray data from the canine 1.0 ST Affymetrix chip.
Finally, Gene Ontology (GO) Enrichment Analysis and Pathway Enrichment Analysis was also performed using the IPA platform to determine the biological significance of the genes differentially expressed between Ad-MSC and BM-MSC (Fig. 8), in which the top 25 upregulated and downregulated bio-functions and pathways are depicted. Ad-MSC have higher expression of genes in the VEGF signaling, G receptor alpha signaling, STAT3 pathways. BM-MSC have higher expression of genes in the corticotropin pathway, IL-1 pathway, B cell receptor pathway, and MAPK signaling pathways. Functional pathways upregulated in Ad-MSC included cardiovascular system development, endothelial cell development, carbohydrate metabolism, and other cell growth-related functions, while BM-MSC were upregulated in functional genes related to bone and connective tissue development, digestive system and other functions related to cytokinesis.

Over and under-expressed genes, pathways and functions. The IPA software was used to determine the biological significance and pathways associated with differentially expressed genes between Ad-MSC and BM-MSC.
Discussion
Studies in spontaneous, large animal disease models can serve as an important role for assessment of stem cell therapies ultimately intended for use in humans. Dogs develop many of the same inflammatory and autoimmune diseases as humans and thus clinical studies of stem cell therapy in these canine models can be particularly informative with respect to safety and potential efficacy, in time frames much accelerated compared to human studies [64].
However, it is also known that the mechanism(s) of action of MSC varies among species. For example, IDO-dependent mechanisms of immune modulation by MSC dominate in humans, nonhuman primates, and pigs, whereas in rabbits and rodents NO-dependent mechanisms of immune suppression are much more important [20,34,65,66]. Therefore, it is useful for evaluating MSC therapy in spontaneous disease models such as dogs to determine which immune modulatory pathways are operative. Based on the studies reported here, we conclude that MSC from dogs utilize different pathways than either humans or rodents, relying primarily on the TGF-β pathway, adenosine pathway, and cyclooxygenase pathways for T cell suppression.
We did not find evidence for utilization of either the NO or IDO pathways by canine MSC. These results differ from those of Kang et al. who reported that the IDO pathway was operative in canine Ad-MSC-mediated suppression of T cells [50]. However, Kang et al. also reported that canine coculture with MSC resulted in an increased production of the inflammatory cytokine IFN-γ, rather than suppression as we found in our studies. In their culture system irradiated MSCs were also used, along with multiple growth factors in the culture media, which may have had an effect on cellular responses and signaling. Nonetheless, in our studies we utilized two different inhibitors of the IDO pathway (1-MT and CAY 10581), at previously published doses [50,67,68], and failed to find evidence for IDO-dependent T cell suppression by canine MSC. Therefore, while we cannot fully explain the discrepancy between our studies and those of Kang et al., we believe that results are consistent with the conclusion that the IDO pathway does not play an important role in T cell suppression by canine MSC.
Previous studies have revealed important differences between Ad-MSC and BM-MSC with respect to a variety of different properties, including osteogenesis, protection from sepsis, and healing of cardiac infarction [69 –72]. Recently, the immune modulatory properties of human Ad-MSC and BM-MSC were compared [73]. In these studies, there were differences in cytokine secretion profiles between the two types of MSC, but when tested for T cell suppression activity, the differences observed were relatively minor. Likewise, our studies also did not detect important differences in potency between canine Ad-MSC and BM-MSC in terms of suppression of T cell activation. Moreover, the pathways used for immune modulation were also similar, with one important exception. In the case of canine BM-MSC, the cyclooxygenase pathway was identified as an important pathway for T cell suppression, in addition to the TGF-β and adenosine receptor-mediated pathways. Identification of the cyclooxygenase pathway for T cell suppression by canine BM-MSC is in agreement with a previous study that also identified PGE2 production as a major mediator of T cell suppression by canine MSC [51]. However, this previous study did not evaluate the activity of other T cell suppression pathways.
It is now accepted that MSC generally do not exert their immune suppressive effects until they are activated to upregulate T cell suppressive pathways by proinflammatory cytokines, especially IFN-γ [35,36]. Indeed, several studies have revealed that when MSC are administered in the absence of IFN-γ or other inflammatory signals, they fail to exert significant immune modulatory activity [33,35]. Thus, we conducted studies to determine whether MSC activation by IFN-γ was also required for activity by canine MSC. Thus, we observed that when MSC were cocultured with activated T cells, and IFN-γ release was neutralized at the initiation of the co-cultures, T cell suppression was also significantly abrogated (Fig. 5). Therefore, it appears that canine MSC, like human and mouse MSC, must also be “licensed” by inflammatory cytokines to become functionally active in immune modulation.
Our studies also investigated other processes by which MSC might elicit immune downmodulation, in addition to suppression of T cell proliferation and reduction in cytokine production. For example, a net loss of activated T cells via increased cell death could over time lead to immune suppression. Thus, we evaluated T cell apoptosis and cell death responses to MSC using in vitro cocultures. Activation of T cells with ConA in itself (without addition of MSC) induced a significant increase in early and late T cell apoptosis, and coculture with Ad-MSC or BM-MSC did not significantly accelerate this process (Fig. 6). However, T cell death was significantly increased when activated T cells were cocultured with MSC, particularly at later time points in culture (Fig. 6). These findings suggest that MSC may indirectly induce necrotic cell death in activated T cells, an effect that would result in an overall net loss of T cells in inflamed sites or where T cells might be exposed to accumulated MSC, such as the lungs, spleen, or lymph nodes.
The phenotype of canine Ad-MSC and BM-MSC was found to be relatively similar, based on cell surface marker expression, growth characteristics in culture, and expression of certain intracellular stem cell markers, though some morphological differences were noted (Figs. 1 and 2). Gene expression analysis, however, revealed several important differences in genes related to immunological functions between Ad-MSC and BM-MSC. For example, GO enrichment analysis of differential gene expression (based on P < 0.05 differences) revealed a significant enrichment of two genes in the adenosine metabolic process and also in two genes involved in the prostaglandin metabolic process, and enrichment of >5 genes in the TGF-β signaling and binding pathways in Ad-MSC versus BM-MSC (Supplementary Fig. S2). In Ad-MSC expression of the PLA2G5 gene for phospholipase A2 and the PTGS2 gene (prostaglandin-endoperoxide synthase 2) involved in PGE synthesis were both down regulated, which is consistent with the lack of response of Ad-MSC to PGE2 COX1 and COX2 pathway inhibitors.
Pathway analysis also showed differential expression of genes regulated downstream of the TGF-β receptor (Supplementary Fig. S2). These findings highlight the complexity of the cell regulatory networks operative in both Ad-MSC and BM-MSC and help explain how blocking TGF-β receptor signaling might have very tissue-specific effects. The adenosine pathway analysis also demonstrated upregulation of the adenosine deaminase (ADA) gene in Ad-MSC, a gene that is involved in purine synthesis (Supplementary Fig. S2).
In summary, we found that canine Ad-MSC and BM-MSC were roughly equivalent in terms of their immune modulation potency and their surface phenotypic properties, and they resembled in many respects analogous populations of human and rodent MSC. Overall, relatively minor differences were uncovered in gene expression patterns between Ad-MSC and BM-MSC; with the most notable differences being in the VEGF signaling pathway, which is upregulated in Ad-MSC, and the connective tissue and bone differentiation pathways, which are upregulated in the BM-MSC.
Canine MSC were found to rely on relatively distinct pathways of T cell suppression relative to MSC from humans and rodents, preferentially utilizing TGF-β, adenosine, and cyclooxygenase pathways to a much greater degree than IDO or NO-dependent pathways. Induction of T cell necrosis by canine MSC also appeared to be an additional mechanism of immune modulation. These findings help to provide a better understanding of canine MSC biology and will serve to facilitate studies of MSC-based therapies for modulation of inflammatory diseases in spontaneous models of canine disease.
Footnotes
Acknowledgments
These studies were supported by grants from the Shipley Foundation Program in Applied Regenerative Medicine and by an NIH T32 award 5T32OD010437-14 to D.R. The authors thank Dr. Michael G. Edwards of University of Colorado Denver for assistance in microarray analysis.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.