
Abstract
Tumors recruit bone mesenchymal stem cells (BMSCs) to localize to tumor sites, which induces their conversion into cancer-associated fibroblasts (CAFs) that facilitate tumor progression. However, this process is poorly understood on the molecular level. In this study, we found that 4T1 breast cancer cells promoted the migration of BMSCs, and bFGF neutralizing antibody inhibited the migration of BMSCs induced by a tumor-conditioned medium. In addition, exogenous bFGF enhanced the migration of BMSCs in a dose-dependent manner in vitro. Furthermore, BMSCs promoted the proliferation of 4T1 tumor cells under BMSC-conditioned medium and in tumor xenograft model. Dramatically, BMSCs expressed CAF markers and produced collagen in the tumor microenvironment, and this transition was blocked by bFGF antibody. In addition, exogenous bFGF induced CAF differentiation of BMSCs. And bFGF increased phosphorylation of Erk1/2 and Smad3 in BMSCs and Erk inhibitor PD98059 was shown to block bFGF-induced Erk and Smad3 phosphorylation, suggesting that Erk/Smad3 signaling pathway involved in BMSC transdifferentiation induced by bFGF. Collectively, our results indicate that bFGF signaling plays indispensable roles in BMSC recruitment and transdifferentiation into CAFs and the consequent protumor effects, and targeting tumor stroma through bFGF inhibition maybe a promising strategy to suppress tumor progression.
Introduction
I
Bone mesenchymal stem cells (BMSCs), which are characterized by their ability to self-renew and differentiate into tissues of mesodermal and nonmesodermal cell lineages, play crucial roles in supporting hematopoiesis [10,11]. BMSCs can be found throughout the body, but they are often involved in tissue remodeling after injury or chronic inflammation where they differentiate into a variety of connective tissue cell types [12,13]. Studies have increasingly shown that BMSCs play an important role in tumor development and progression. BMSCs recruited to tumors selectively proliferate and contribute to the formation of the tumor-associated stroma [14]. Furthermore, recruitment of BMSCs in breast carcinomas facilitates metastatic spread [15].
Previous studies have suggested that BMSCs have the ability to home to tumor sites [16,17] and differentiate into cancer-associated fibroblasts (CAFs) under the tumor-conditioned medium (TCM) [18]. These data indicate that secreted factors in tumor microenvironment play essential roles in BMSC recruitment and transdifferentiation. To date, BMSCs are known to respond to a number of chemotactic factors, including CXCL16/CXCR6 [19], IL-6 [20], IL-8 [21], and SDF-1/CXCR4 [22]. Using a combination of chromatography and electrospray tandem mass spectrometry, Lin et al. found novel soluble signaling molecules that induce BMSC chemotaxis to be present in the TCM of a breast cancer cell line [23]. Differentiation of BMSCs into CAFs under a tumor environment indicates that there are secreted factors from tumor cells to regulate this process. One study showed that inhibiting transforming growth factor-β (TGF-β) signaling in human BMSCs can block their differentiation to CAFs induced by TCM [24].
bFGF, which belongs to the heparin binding growth factor family, mediates various cellular events, including proliferation, motility, and differentiation [25 –28]. bFGF increases embryonic myogenic cell migration in a dose-dependent manner [29]. bFGF also plays an important role in the proliferation and stem cell plasticity of adipose-derived stem cells [30]. Furthermore, bFGF can activate normal fibroblasts and give them properties that are analogous to those of CAFs [31]. In addition, increased mRNA expression of α-smooth muscle actin (α-SMA), vimentin, and collagen in human BMSCs has been noted after 14–28 days of exposure to low-dose bFGF [28]. However, whether bFGF signaling participates in BMSC chemotaxis for tumor and CAF induction is still unclear.
In this study, we investigated the interplay between tumor cells and BMSCs. We found that bFGF secreted by tumor cells recruits BMSCs to the primary tumor site and induces the differentiation of BMSCs into CAFs to promote tumor growth. Furthermore, we identified the mechanism underlying bFGF-induced BMSC transdifferentiation, and our results demonstrated that Erk/Smad3 signaling pathway was essential for bFGF-induced BMSC transdifferentiation into CAFs.
Materials and Methods
Cell lines
Murine 4T1 breast cancer cells were obtained from the Tianjin Medical University Cancer Research Institute. The cells were maintained in RPMI 1640 (Gibco, Life Technologies, Paisley, United Kingdom) supplemented with 10% fetal bovine serum (FBS, Gibco). Mouse BMSCs were obtained from Cyagen Biotechnology Co., Ltd. (Guangzhou, China). BMSCs were maintained in low-glucose Dulbecco's modified Eagle's medium (Gibco) supplemented with 15% FBS. BMSCs between passage 8 and 15 were used for experiments. All cells were incubated at 37°C in 5% CO2 humidified cell culture incubator.
All experimental protocols were approved by Tianjin Medical University Cancer Institute and Hospital.
Flow cytometry
Phenotypic analysis of BMSCs at passage 10 was performed with flow cytometry. Briefly, BMSCs (1.0 × 106 cells) were trypsinized, washed twice in PBS, and stained with monoclonal antibodies against stem cell antigen 1 (Sca-1), CD29, CD44, CD105, CD11b (an immune cell marker), CD34 (expressed on endothelial cells), CD45 (a marker of all hematopoietic cells), and CD117 (a hematopoietic stem/progenitor cell marker) for 30 min on ice. Labeled cells were analyzed using a FACSCalibur flow cytometer (BD FACSCanto II; BD Biosciences). All flow cytometry antibodies were purchased from BioLegend (San Diego).
Cell differentiation
For adipocyte and osteogenic differentiation, BMSCs were exposed to an adipocyte induction medium (M3017; Tianjin Weikai Bioeng Ltd., China) or an osteogenic differentiation medium (M3015; Tianjin Weikai Bioeng Ltd.) and medium was replaced every 3 days for 2 weeks. Then cells were fixed with 4% paraformaldehyde and stained with oil red O stain [32] or 1-Step™ NBT/BCIP (S1015; Tianjin Weikai Bioeng Ltd.) solution for 30 min and then rinsed with water [33].
Preparation of conditioned medium
4T1 cells were grown in RPMI 1640 supplemented with 10% FBS. When the cells were 70%–80% confluent, the old medium was removed and a fresh serum-free medium was added. The TCM from 4T1 cells was harvested after 16 h of incubation and centrifuged at 1,500 rpm for 10 min to remove cell debris. A BMSC-conditioned medium (CM) was collected in the same way.
Transwell migration assay
The tropism of mouse BMSCs for tumor cells and growth factors was determined using a transwell migration assay. Briefly, BMSCs (1 × 104) were placed in a serum-free medium in the upper well of a transwell plate (Millipore, Billerica, MA). 4T1 cells (1 × 104) were plated on the bottom well for 12 h and the medium was changed to a serum-free medium before BMSCs were added to the top well. TCM was placed in the lower well of the plate. In selected experiments, 10 ng/mL recombinant mouse bFGF (Peprotech, Rocky Hill, NJ), 50 ng/mL bFGF, or 20 ng/mL bFGF antibody (Millipore) was added to the lower compartment. BMSCs were incubated for 12 h at 37°C. The cells attached to the top side of the membrane were removed using cotton swabs. The migrated cells on the bottom side were then fixed with 4% paraformaldehyde and stained with 0.1% crystal violet. Five random fields at × 400 magnification were photographed and counted.
Cell treatment with CM
For the observation of the effect of BMSC-CM on tumor cell proliferation, 4T1 cells were plated at a density of 5 × 103 cells per well in 24-well plates in a tumor cell culture medium (supplemented with 10% FBS or 1% FBS) or a mixture of tumor cell culture medium and BMSC-CM (4:1 or 1:1). Every group consisted of three wells. After culture for 72 h, the tumor cells in each well were collected and counted.
MTS assay
A cell suspension of 4T1 cells (100 μL, 3 × 103 cells/well) was placed in a 96-well plate and treated with a tumor cell culture medium (supplemented with 10% FBS) or a mixture of tumor cell culture medium and BMSC-CM (4:1 or 1:1) for 72 h. 3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetra-zolium (MTS; Promega, Madison, WI) was added (20 μL per well) at a concentration of 1.90 mg/mL to each well, and the plate was incubated at 37°C for 2–4 h. Then, the optical density (OD) of each well was read at 490 nm on a microplate reader.
Immunofluorescence analysis
BMSCs (2 × 104 cells per well) were plated on sterilized coverslips in 12-well plates and allowed to adhere overnight. The following day, 50% TCM, 10 ng/mL bFGF, 10 ng/mL recombinant mouse TGF-β (rm TGF-β, Cell Signaling Technology, Danvers, MA), or 20 ng/mL specific bFGF antibody was added to the cells. After 48 h of cell culture, the cells were fixed with 4% paraformaldehyde at room temperature for 30 min, followed by permeabilization with 0.1% Triton X-100, washing with PBS, and blocking with 1% BSA in PBS. Cells were then immunostained for α-SMA (1:500; Sigma, Saint Louis, MO,
Picric acid–sirius red staining
BMSCs (1 × 105 cells per well) were plated on six-well plates and allowed to adhere overnight. The following day, 50% TCM, 10 ng/mL bFGF, 10 ng/mL TGF-β, or 20 ng/mL specific bFGF antibody was added to the cells. After 48 h of cell culture, the cells were fixed with 4% paraformaldehyde at room temperature for 30 min, permeabilized with 0.1% Triton X-100, and washed with PBS. Then, the cells were stained with picric acid–sirius red (0.1% sirius red in saturated aqueous picric acid, Solarbio, Beijing, China) to detect collagen expression. Stained sections were examined by microscopy. Three randomly selected images per section were digitally captured (magnification × 400).
Western blotting
Proteins were extracted from BMSCs cultured in a normal medium, 50% TCM, 10 ng/mL bFGF, 10 ng/mL TGF-β, 20 ng/mL bFGF antibody, or 10 μM Erk inhibitor PD98059 (Selleckchem) using mammalian protein extraction reagent supplemented with a protease inhibitor and phosphatase inhibitor cocktail (Solarbio). The protein was fractionated by 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis, transferred to a PVDF membrane, and incubated with primary antibodies against α-SMA (1:2,000; Sigma), vimentin (1:2,000; Sigma), Erk (1:2,000; Cell Signaling Technology), p-Erk (1:2,000; Cell Signaling Technology), Smad3 (1:2,000; Cell Signaling Technology), p-Smad3 (Ser423/425, 1:2,000; Cell Signaling Technology), or β-actin (1:2,000; Sigma). Protein expression was detected using an ECL western blotting detection system.
Real-time reverse transcription–polymerase chain reaction
To measure the levels of α-SMA, vimentin, collagen I, collagen III, and GAPDH in BMSCs, the cells were treated in the same manner as for Western. Total RNA was extracted with TRIzol reagent (Invitrogen). cDNA was synthesized from 2 μg of total RNA using MMLV reverse transcriptase. Real-time polymerase chain reaction (PCR) was carried out with 2 μL of cDNA and SYBR Green Master Mix (Qiagen). The forward and reverse primers used were as follows: α-SMA, 5′-CCCAGACATCAGGGAGTAATGG-3′ and 5′-TCTATCGGATACTTCAGCGTCA-3′; vimentin, 5′-CGTCCACACGCACCTACAG-3′ and 5′-GGGGGATGAGGAATAGAGGCT-3′; collagen I, 5′-TAAGGGTCCCCAATGGTGAGA-3′ and 5′-GGGTCCCTCGACTCCTACAT-3′; collagen III, 5′-CTGTAACATGGAAACTGGGGAAA-3′ and 5′-CCATAGCTGAACTGAAAACCACC-3′; and GAPDH, 5′-AATGGATTTGGACGCATTGGT-3′ and 5′-TTTGCACTGGTACGTGTTGAT-3′. The thermal conditions were as follows: 3 min at 95°C, followed by 40 cycles of 95°C for 15 s, 55°C for 30 s, and 72°C for 1 min. For semiquantitative analysis, the target gene expression was normalized to that of GAPDH. Data are expressed as percentages compared with the control.
Immunohistochemistry
All tumor masses fixed in 10% neutral formalin were embedded in paraffin and cut into parallel consecutive 4-μm thick sections for subsequent immunohistochemical study. Briefly, the endogenous peroxidase activity was blocked with 3% H2O2 for 20 min. The sections were permeabilized with sodium citrate–hydrochloric acid buffer solution (pH 6.0) for 20 min in a microwave. Then, 1% BSA was applied for 30 min to prevent nonspecific adherence of serum proteins. Next, sections were incubated with primary antibody anti-mouse α-SMA (1:500; Sigma) or anti-PCNA (1:500; Abcam, Cambridge, MA) overnight at 4 C. After washing with PBS, sections were incubated with secondary antibody diluted 1:1,000 at 37 C for 45 min and washed with PBS. DAB (diaminobenzidine, BOSTER, Wuhan, China) was applied for 5 min and then washed off with PBS. Finally, sections were counterstained with hematoxylin. For a negative control, PBS was used instead of primary antibody. The sections were analyzed by light microscopy. The quantification of α-SMA and PCNA density was performed using Image-Pro Plus image analysis software.
Animal experiments
Six-week-old female BALB/c mice were purchased from the Beijing Vital River Experimental Animal Technical Company and used in accordance with the Animal Ethics and Welfare Committee of Tianjin Medical University Cancer Institute and Hospital. 4T1 cells, BMSCs, and 4T1-activated BMSCs (BMSCs that were exposed to 50% TCM for 7 days and culture medium changed every 2 days) were prepared as single-cell suspensions (2 × 106 4T1 cells in 0.1 mL PBS, a mix of 2 × 106 4T1 cells and 1 × 106 BMSCs in 0.1 mL PBS, or a mix of 2 × 106 4T1 cells and 1 × 106 4T1-activated BMSCs in 0.1 mL PBS, respectively). Cells were injected subcutaneously at one site on the back of BALB/c mice. Each group contained 10 animals. Mice were examined every 2 days, and tumor growth was evaluated by measuring the length and width of the tumor mass. Palpable tumors at the injection sites that were more than 3 mm in diameter were monitored as a tumor. At the end of the experiment, tumor masses were removed and fixed in 10% neutral formalin for histologic preparations.
Statistical analysis
All quantitative data are shown as mean ± standard deviation. All experiments were performed in triplicate. Statistical significance was analyzed by one-way ANOVA using SPSS 16.0 software. Comparisons with P < 0.05 were considered to be statistically significant.
Results
The characteristics of BMSCs
Morphologically, mouse BMSCs had a spindled, fibroblast-like appearance after expansion (Fig. 1A). Mouse BMSCs were tested for Sca-1, CD105, CD29 and CD44 positivity, CD34, CD45, CD11b, and CD117 negativity using flow cytometric analysis (Fig. 1D), and multilineage differentiation into adipocytes (Fig. 1B) and osteocytes (Fig. 1C). The surface marker expression pattern corresponding to mouse BMSCs was described previously [34].
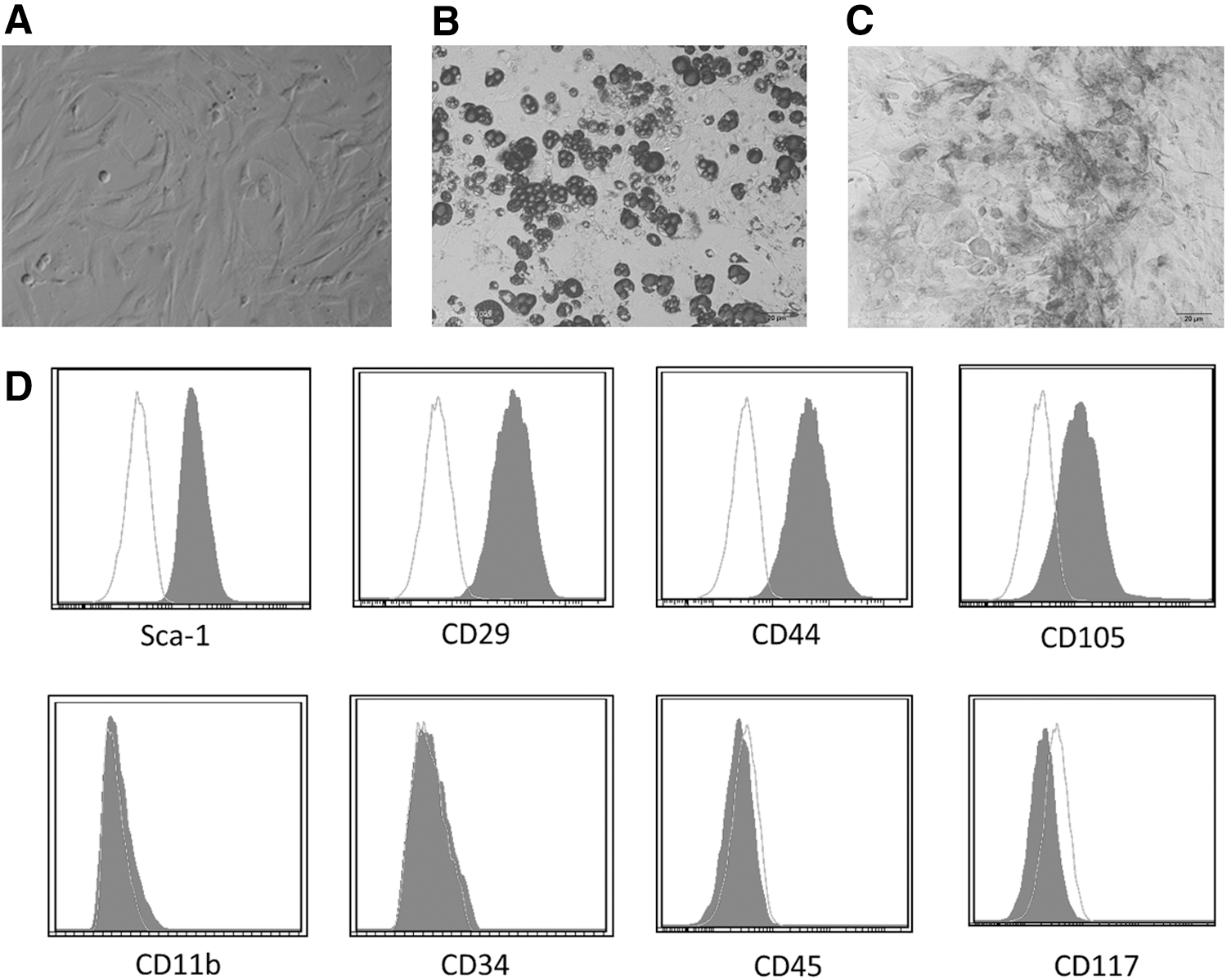
Characteristics of BMSCs (P10). Morphological, BMSCs were long, spindle shaped, and fibroblastic in appearance
Effect of tumor cell and TCM on BMSC migration
To examine the chemotaxis of BMSCs toward tumor environment, BMSCs were exposed to 4T1 tumor cells or TCM, and then their ability to migrate toward tumor environment was assayed in transwell chamber migration assay.
As shown in Fig. 2A, the migrating cell numbers of BMSCs in 4T1 and TCM groups were significantly higher compared with the control group (Fig. 2A, P < 0.001 and P < 0.001, respectively).

bFGF-induced BMSC migration.
Tumor cell-derived bFGF induces BMSC migration
As there was no direct contact between 4T1 tumor cells and BMSCs in the in vitro migration assay, a 4T1 cell-secreted factor(s) was most likely responsible for the stimulation of BMSC migration. To investigate whether bFGF secreted by 4T1 cells induces BMSC migration, we then added neutralizing bFGF antibody to the TCM to block the bFGF signaling activity. At a 20 ng/mL concentration of bFGF antibody, BMSC migration was attenuated in response to the TCM (Fig. 2B, P < 0.025). Taken together, these data suggest that bFGF in the tumor microenvironment stimulates BMSC migration.
bFGF enhances the migration of BMSCs in a dose-dependent manner in vitro
We then investigated the role of exogenous bFGF in BMSC migration in vitro. Both 10 ng/mL (P < 0.025) and 50 ng/mL (P < 0.001) bFGF promoted BMSC migration (Fig. 2C). Moreover, there was a 1.8-fold (P < 0.001) increase in the number of migrated cells toward 50 ng/mL bFGF compared to 10 ng/mL bFGF.
BMSC-CM promotes 4T1 cell proliferation in vitro
For the observation of the effect of BMSCs on tumor cell proliferation, 4T1 cells were maintained in a mixture of tumor cell growth medium (supplemented with 10% FBS or 1% FBS) and BMSC-CM (volume in 4:1 or 1:1). Compared to the control medium, both 20% BMSC-CM and 50% BMSC-CM had a proproliferative effect on 4T1 cells (Fig. 3A, P < 0.02 and P < 0.001, respectively). To exclude the influence of FBS on cell proliferation, we decreased the serum concentration to 1% FBS. Under 1% FBS, both 20% BMSC-CM and 50% BMSC-CM promoted the proliferation of 4T1 cells (Fig. 3B, P < 0.05 and P < 0.002, respectively).
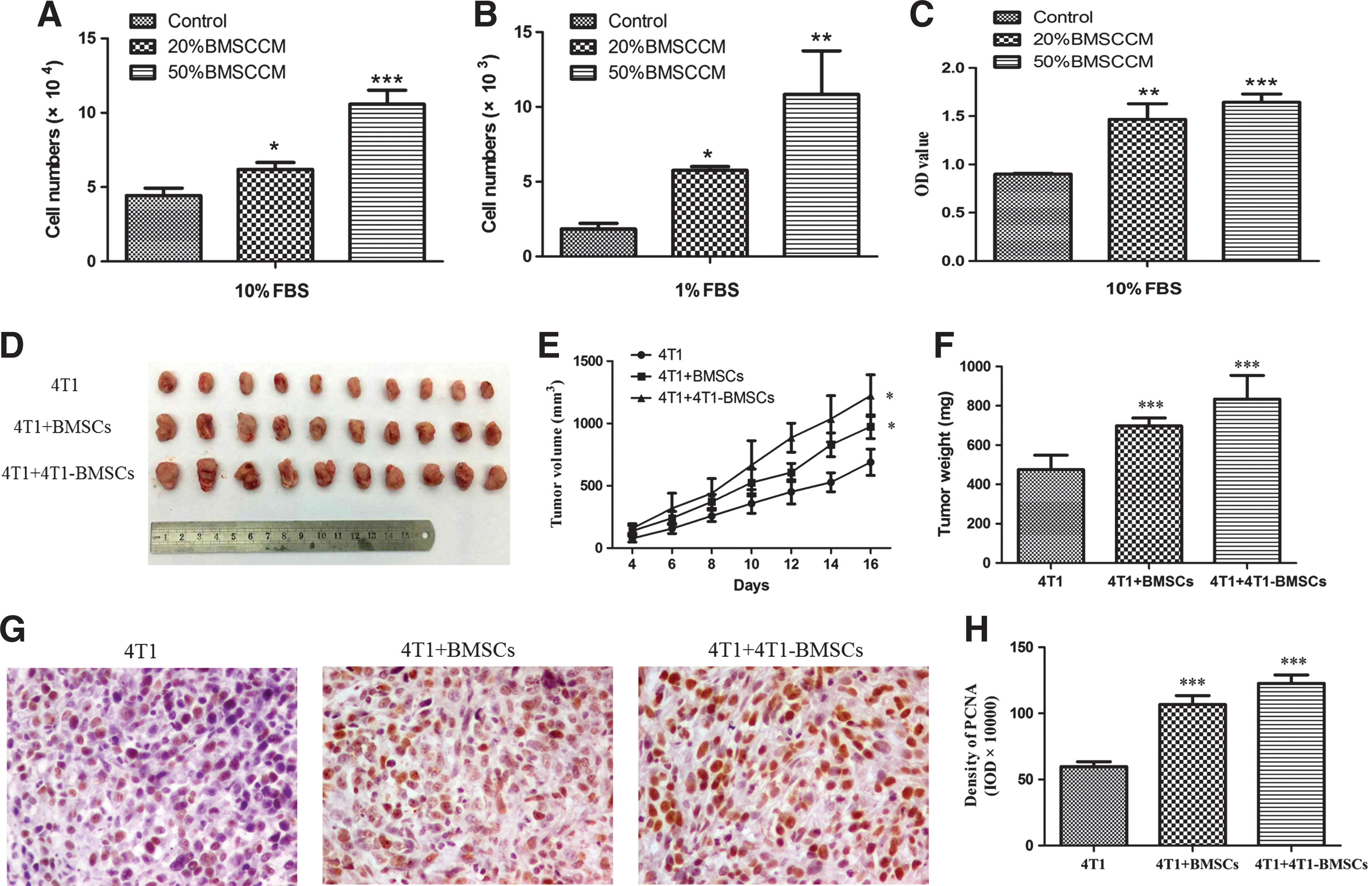
Effect of BMSCs on 4T1 breast cancer growth.
In accordance with the cell count assay, the MTS cell proliferation assay also showed that 20% and 50% BMSC-CM promoted 4T1 cell proliferation. 20% and 50% BMSC-CM group had a much higher OD value than the control medium (Fig. 3C, P < 0.002 and P < 0.001, respectively).
BMSCs promote tumor growth in vivo
To determine if BMSCs support tumor growth in vivo, a set of xenograft experiments were performed using mouse 4T1 breast cancer cells. Both the presence of BMSCs and TCM-pretreated BMSCs clearly supported tumor growth when compared with 4T1 cells alone (Fig. 3D). 4T1 cells mixed with BMSCs or TCM-pretreated BMSCs generated tumors of greater volume and weight when compared to 4T1 cells-alone group (Fig. 3E, F). The tumor weight in the BMSCs and TCM-pretreated BMSC group was increased by 46.7% (P < 0.001) and 75.5% (P < 0.001), respectively, compared to the control group.
Effect of BMSCs on the expression of PCNA in vivo
To quantify PCNA expression, the tumor sections were photographed with a digital camera at × 400 magnification (Fig. 3G), and the integral optical density of PCNA in each section was analyzed using Image-Pro-Plus image analysis software. We found that the expression of PCNA in tumor tissues significantly increased in the BMSCs (P < 0.001) and TCM-treated BMSC groups (P < 0.001) compared to the 4T1 group (Fig. 3H).
Effect of BMSCs on the expression of α-SMA in vivo
Previous studies have indicated that BM-derived cells such as mesenchymal stem cells (MSCs) are one of the origins of CAFs [22,35]. Therefore, tumor sections were processed for the immunohistochemical staining of α-SMA at the end of the observation period (Fig. 4A). Strong positive staining for α-SMA was seen in tumors derived from the BMSCs (P < 0.001) and TCM-pretreated BMSC groups (P < 0.001). This suggested that BMSCs coinjected with 4T1 cells may become functionally incorporated into the tumor stroma and further facilitate tumor growth.
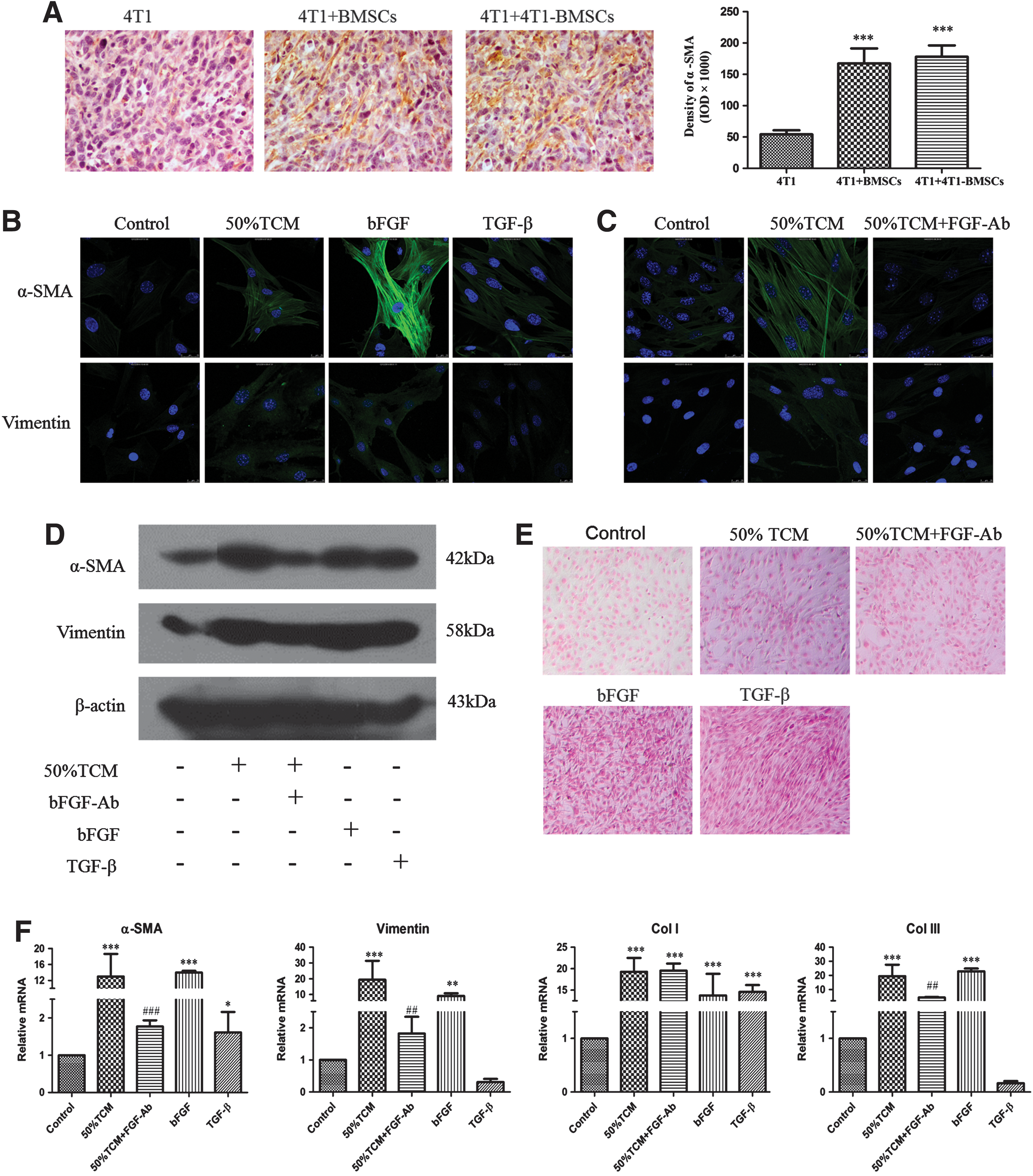
Expression of CAF markers in tumor issue sections and in BMSCs under 50% TCM, bFGF antibody, bFGF, or TGF-β treatment.
Activation of BMSCs by TCM—expression of markers specific to CAFs
CAFs can be characterized by an increased expression of proteins such as α-SMA and vimentin, and BMSC-derived cells expressing CAF markers contribute to the stroma of mixed xenograft tumor. We next examined the differentiation of BMSCs through exposure to the TCM in vitro. Immunofluorescence assay showed that native BMSCs expressed α-SMA and vimentin at a low level, while the expression levels of α-SMA and vimentin increased after exposure to the TCM, indicating that the TCM induced BMSCs to acquire a CAF-like phenotype (Fig. 4B).
To further confirm the effect of TCM on BMSC differentiation, we assessed the TCM-stimulated expression of α-SMA and vimentin through western blotting analysis (Fig. 4D). Accordingly, BMSCs exposed to the TCM resulted in the increased expression of α-SMA and vimentin. Furthermore, there were 14-fold and 19-fold upregulations in mRNA expression levels of α-SMA (P < 0.001) and vimentin (P < 0.001) after 48 h of TCM treatment, respectively (Fig. 4F).
CAFs also express collagen [35] and we further determined collagen expression to reflect the CAF activity. Prominent collagen deposition was found after picric acid–sirius red staining and the TCM group secreted slightly more collagen than the control group (Fig. 4E). The mRNA expression level of collagen I (P < 0.001) and collagen III (P < 0.001) also significantly increased after TCM exposure (Fig. 4F).
bFGF signaling induced BMSC differentiation
TCM exposure led BMSC differentiation to CAF-like phenotype, indicating that a soluble factor(s) was likely responsible for the differentiation stimulation. When specific bFGF antibody was added to the TCM, the expression of α-SMA, vimentin, and collagen was reduced in both protein and mRNA levels, compared to the TCM group without bFGF antibody (Fig. 4C–F). These data indicate that tumor cell-derived bFGF may involve in BMSC differentiation into CAFs.
To date, differentiation of BMSCs to CAFs has been induced in vitro mainly by TGF-β [24,36]. However, another study indicates that bFGF is capable of increasing the mRNA expression of α-SMA, vimentin, and collagen in human BMSCs after low-dose bFGF (3 ng/mL) exposure for 14 or 28 days [28]. In this study, the expression levels of α-SMA, vimentin, and collagen in bFGF-treated BMSCs were determined by immunofluorescence, western blotting, picric acid–sirius red staining, or reverse transcription-polymerase chain reaction (RT-PCR) analysis (Fig. 4B, D–F). These results showed that extraneous bFGF upregulated α-SMA, vimentin, and collagen expression in both mRNA and protein levels. Furthermore, immunofluorescence and RT-PCR analysis showed that bFGF had a stronger effect on α-SMA, vimentin, and collagen expression than TGF-β at the same concentration.
Erk/Smad3 signaling pathway involved in BMSC differentiation induced by bFGF
The progression from undifferentiated BMSCs to differentiated cells involves the activation or suppression of certain signaling pathways. These pathways may be specific to one lineage or may be important for the differentiation process in general. As shown in Fig. 5A, Erk1/2 was phosphor activated by bFGF, indicating that Erk1/2 signaling accompanied the upregulation of α-SMA and vimentin in transdifferentiated BMSCs. Additionally, compared to control group, phosphorylation of mothers against decapentaplegic homolog 3 (p-Smad3) was also elevated in bFGF induced cells (Figure 5A).

Erk/Smad3 signaling pathway is essential for BMSC differentiation to CAFs induced by bFGF.
To further elucidate the role of Erk/Smad3 signaling in bFGF-induced BMSC differentiation, we blocked the activation of Erk1/2 by treating BMSCs with a specific Erk inhibitor PD98059. In the presence of PD98059, bFGF-induced Erk and Smad3 phosphorylation was significantly decreased and both α-SMA and vimentin protein levels were dramatically downregulated (Fig. 5A), which indicated that Erk/Smad3 signaling pathway was involved in bFGF-induced BMSC differentiation (Fig. 5B).
Discussion
The surrounding tumor microenvironment appears to be an important determinant in the final outcome of the disease [37,38]. BM-derived cells, particularly BMSCs, seem to contribute to the tumor stroma and regulate tumor growth [8].
Previous studies have suggested that BMSCs have the ability to home to tumor sites [16,17]. Due to their tropism for tumors, BMSCs may serve as a platform for delivery of biological agents for anticancer therapy [39,40], an approach that has shown promise in preclinical models [41]. Secreted factors in tumor microenvironment maybe involved in tumor chemotaxis toward BMSCs. Recently, Schmidt et al. reported that bFGF increased the migratory activity of BMSCs through activation of the Akt/protein kinase B pathway [42]. Thus, whether tumor cells recruit BMSCs by secreting bFGF is a promising topic of study. In this study, we provided evidence that tumor-secreted bFGF promoted BMSC migration toward tumor cells by adding a specific bFGF neutralizing antibody in TCM. Moreover, exogenous bFGF induced BMSC migration in a dose-dependent manner. Notably, bFGF did not promote BMSC proliferation at the same dose and time (12 h) as the migration assay (Date not shown). Indeed, BMSCs are known to express the bFGF receptor on their surface [43]. Our results suggest that tumor cells recruit BMSCs by secreting bFGF.
Several studies have shown that BMSCs inhibit tumor growth in murine model of glioma by inhibition of angiogenesis [44] and hepatocellular carcinoma by inducing tumor cell apoptosis [45]. However, there are controversies in BMSC effects on tumor growth. BMSCs have been showed to support tumor growth and metastasis once integrated into the tumor microenvironment [15,19]. Our findings demonstrate that BMSCs promote the proliferation of 4T1 cells in vitro and tumor growth in a murine model of breast cancer. According to the observation of tumor mass, PCNA expression in tumor cells increased in the coinjection group. Therefore, BMSCs could promote 4T1 breast cancer growth in vitro and in vivo. Many studies indicate that BMSCs have several tumor-growth promoting functions in the tumor microenvironment, including expressing growth factors, enhancing tumor vessel formation, and creating tumor stem cell niches [8]. Interestingly, one study indicates that BMSCs play a dual role in tumor growth in vitro and in vivo. Human BMSCs inhibited lung and esophageal tumor cell proliferation and invasion in vitro; but, in vivo, BMSCs favored tumor formation and growth [46]. These controversies maybe attributed to the differences in tumor types or experimental conditions and further studies are needed to elucidate this phenomenon. MSCs, including BMSCs, have the capacity to differentiate into cells of connective tissue lineages, including bone, fat, cartilage, and muscle. These cells have generated a great deal of interest because of their potential use in regenerative medicine and tissue engineering and there are some dramatic examples, derived from both preclinical and clinical studies, which illustrate their therapeutic value [47]. However, according to our results and other studies, the safety remains a key problem to solve for tumor patients receiving BMSC or gene therapy.
Studies have indicated that BM-derived cells, especially BMSCs, not only regulate tumor growth [48] but are also sources of CAFs [35,49]. The CAF population differs morphologically and functionally from normal fibroblasts and expresses α-SMA and vimentin [50]. In our study, BMSCs presented with a bipolar and/or multipolar morphology and then acquired a uniform spindle-shaped morphology, forming parallel arrays and whorls at confluence after TCM treatment. Moreover, upregulated expression of α-SMA, vimentin, and collagen in BMSCs following exposure to the TCM was also confirmed. In addition, the expression of α-SMA in the tumor stroma increased in the coinjection groups in vivo, which suggested that BMSCs coinjected with 4T1 cells may become functionally incorporated into the tumor stroma and promote tumor growth. In addition, BMSCs activated using the TCM were injected together with 4T1 cells, resulting in an accelerated tumor growth. These results support the conclusion that BMSCs are precursors of CAFs under tumor environment and promote tumor growth.
BMSCs exhibited a capability to differentiate into CAFs under TCM treatment, indicating that a soluble factor(s) produced by tumor cells may mediate this process. To date, differentiation of BMSCs into CAFs has been induced in vitro mainly by TGF-β [24]. However, one study indicates that bFGF makes normal fibroblasts having analogous properties to CAFs [31]. In addition, an increased mRNA expression of α-SMA, vimentin, and collagen in human BMSCs has been noted after 14–28 days of exposure to low-dose bFGF [28]. However, the molecular mechanisms underlying bFGF regulation on BMSCs is unclear and there has been little investigation of whether tumor cells secrete bFGF to induce the differentiation of BMSCs to CAFs. In this study, blocking bFGF in the TCM using a specific antibody inhibited the expression of α-SMA and vimentin in BMSCs. Moreover, exogenous bFGF induced BMSCs to acquire a CAF-like appearance and increased α-SMA, vimentin, and collagen expression in both mRNA and protein levels. These data indicate that bFGF under tumor environment is capable of inducing BMSCs differentiation to CAFs.
bFGF is a basic growth factor that signals to the nucleus by binding to FGF receptor and activating multiple signal transduction pathways, including those involving Ras, mitogen-activated protein kinases (MAPKs), extracellular signal-regulated kinases (ERKs), Src, p38 MAPKs, phospholipase-Cγ, Crk, jun N-terminal kinase, and protein kinase C [51]. The ERKs are the main downstream of FGF signaling pathways. In this study, bFGF induced the phosphorylation of Erk1/2 in BMSCs and Erk inhibitor reduced α-SMA and vimentin expression and blocked bFGF-induced Erk phosphorylation, indicating that Erk1/2 signaling plays an indispensable role in BMSC differentiation into CAFs. Smad3 signaling participates in epithelial to mesenchymal transdifferentiation and pathological fibrotic diseases [52]. The Smad family is divided into different groups and Smad3 belongs to R-Smads, which are phosphorylated by receptor kinases. For example, Smad2 and Smad3 are phosphorylated by the TGF-β receptor kinase in the C-terminal tail [53,54]. Indeed, the Smad activity is also regulated by phosphorylation through nonreceptor kinases, such as ERK phosphorylation [55]. Interestingly, in this study, bFGF increased Smad3 phosphorylation and Erk inhibitor reduced bFGF-induced Smad3 phosphorylation, suggesting that the Erk/Smad3 signaling pathway was involved in BMSC transdifferentiation induced by bFGF.
Conclusion
In summary, these data identify bFGF secreted by breast cancer cells recruits BMSCs to primary tumor sites, and BMSCs support tumor growth partly by differentiating into CAFs, which are induced by bFGF (Fig. 6). Furthermore, our results indicate that bFGF promote CAF differentiation of BMSCs through Erk/Smad3 signaling pathway. bFGF is known for its proangiogenic effect; however, our results show that bFGF acts as a novel regulatory molecule that promotes migration and CAF differentiation of BMSCs. The cross talk between BMSCs and tumor cells is complicated and we suggest that the exploitation of BMSCs in new therapeutic strategies should be undertaken with caution in malignant conditions.

bFGF induces the recruitment and CAF differentiation of BMSCs to promote tumor growth. Model shows putative potential mechanisms underlying primary breast cancer progression by recruitment of BMSCs. Tumor-derived bFGF interacts with its receptor FGFR on BMSCs to recruit BMSCs into tumor sites and induces BMSCs into CAFs to promote tumor growth. BMSCs, bone mesenchymal stem cells.
Footnotes
Acknowledgment
This work was supported by the National Natural Science Foundation of China (81173376 and 81473441).
Author Disclosure Statement
No competing financial interests exist.