
Abstract
Human pluripotent stem cells (hPSC) have great clinical potential through the use of their differentiated progeny, a population in which there is some concern over risks of tumorigenicity or other unwanted cellular behavior due to residual hPSC. Preclinical studies using human stem cells are most often performed within a xenotransplant context. In this study, we sought to measure how undifferentiated hPSC behave following xenotransplant. We directly transplanted undifferentiated human induced pluripotent stem cells (hIPSC) and human embryonic stem cells (hESC) into the adult mouse brain ventricle and analyzed their fates. No tumors or precancerous lesions were present at more than one year after transplantation. This result differed with the tumorigenic capacity we observed after allotransplantation of mouse ESC into the mouse brain. A substantial population of cellular derivatives of undifferentiated hESC and hIPSC engrafted, survived, and migrated within the mouse brain parenchyma. Within brain structures, transplanted cell distribution followed a very specific pattern, suggesting the existence of distinct microenvironments that offer different degrees of permissibility for engraftment. Most of the transplanted hESC and hIPSC that developed into brain cells were NeuN+ neuronal cells, and no astrocytes were detected. Substantial cell and nuclear fusion occurred between host and transplanted cells, a phenomenon influenced by microenvironment. Overall, hIPSC appear to be largely functionally equivalent to hESC in vivo. Altogether, these data bring new insights into the behavior of stem cells without prior differentiation following xenotransplantation into the adult brain.
Introduction
S
The methods used to make hIPSC include a variety of genetic and nongenetic approaches, including conventional viral transduction, but also transient introduction of DNA [9], recombinant proteins [10], miRNA [11], and small molecules [12,13]. Shared among most of these approaches is the introduction of core pluripotency-related factors, including OCT4/POU5F1, SOX2, and NANOG as well as in some conditions MYC, KLF4, or LIN28A. These factors reprogram somatic cells such as fibroblasts into hIPSC by globally transforming the epigenetic landscape.
hIPSC possess most of the same properties as hESC, including gene expression, epigenomic patterns, and teratoma-forming activity. Their proteomes [14] as well as metabolome [15,16] are nearly identical. hESC and hIPSC also produce very similar differentiated derivatives in vitro [17]. While hIPSC and hESC are clearly distinct cell types, the fact that hIPSC are so similar to hESC, despite being derived from a variety of somatic cell types is encouraging for the prospect of using hIPSC for clinical applications. However, studies of human hIPSC behavior post-transplant employing transplantation into actual organs are relatively few. Most often hIPSC and hESC are compared using teratoma formation assays [18,19], but as useful as the teratoma assays are in one sense (practically speaking as a pluripotency assay), they have some notable weaknesses as well from a preclinical perspective. For example, transplanting large number of undifferentiated cells subcutaneously into an immunodeficient mouse has very limited relevance to how such cells or their derivatives might be used in a clinical setting and there is a lack of standardization in terms of how the assays are done [18], together representing a challenge in the field from a translational perspective [7,20]. Both mouse embryonic stem cells (ESC) and induced pluripotent stem cells (IPSC) have some notable similarities to cancer cells [21] that suggest that induced pluripotency and tumorigenicity share some elements, likely manifesting in the human cells as well. A key open question is the extent to which hIPSC and hESC derivatives are safe in a transplantation setting involving injection directly into the brain.
Some published transplantation studies of hIPSC or their derivatives provide important insights into the tumorigenic capacity of pluripotent cells. For example, in the recent landmark study of clinical use in a human patient of hIPSC-derived retinal pigmented epithelial cells (RPEs), no evidence of tumors or even precancerous lesions was present [22]. Related studies with hIPSC-derived RPEs for retinal disease and oligodendrocyte progenitors for spinal cord injury in mice reported no evidence of tumors [23,24]. In another study, IPSC lines were termed as “safe” or “unsafe” based on the residual teratoma-forming activity of their neurosphere derivatives [25]. One unsafe hIPSC line failed to produce teratoma in the recipient spinal cord, but importantly it did produce clusters of Nanog+ cells indicating that hIPSC potentially have tumorigenic potential beyond teratoma as do hESC [26]. Another study transplanted adenovirus-generated IPSC derived from rat tail-tip fibroblasts into the striatum of immunocompetent rats. They observed that these IPSC produced region-specific neuronal phenotypes, in the absence of tumor formation, at 90 days post-transplantation [27]. hIPSC-derived dopaminergic neurons improved function in a rat model of Parkinson's disease and no teratoma formation was evident at 12 weeks [28]. However, using a distinct differentiation protocol, another study of hIPSC found that hIPSC derivatives generated proliferative, Nestin+ precursor cells in the rat brain [29] following transplant, indicating that even predifferentiation is not a complete guarantee of safety. In addition to the apparently intrinsic ability of hIPSC and hESC to form teratoma in certain contexts, hIPSC also can possess genetic and epigenetic changes that could have some relevance for their behavior, including their tumorigenicity [30 –34], but whether these alterations have functional consequences in a clinical setting remains unknown. The central issue for both hESC and hIPSC intended for differentiation and clinical use is likely to be validation by thorough screening of the aforementioned properties such as through genome sequencing [35].
One broad, likely accurate assumption in the regenerative medicine field is that stem cell-based therapies will require predifferentiation of pluripotent cells and in some cases sorting-based negative selection against potential residual stem cells before transplantation [36 –38]. A concern of injecting undifferentiated pluripotent cells is the unknown potential of undifferentiated pluripotent cells to give rise to teratoma and other tumors once transplanted into patients. However, while both hESC and hIPSC give rise to teratoma when injected subcutaneously into immunodeficient mice, the behavior of these naive cells when injected into recipients with healthy immune systems remains less clear. Previous studies on transplantation of undifferentiated IPSC into the brain in animal models have yielded mixed results in terms of teratoma formation. While some studies did show teratoma formation, others did not [39]. Experiments where IPSC were transplanted into striatum and cortex produced tumors when the recipient brains had suffered an ischemic event [40], whereas transplanted IPSC did not form tumors in normal brains [28,41], indicating a potential role of the microenvironment in the behavior of the transplanted cells.
Ideally, whether in the context of pre- or postdifferentiation, the potential clinical utility of hIPSC should be evaluated in parallel to hESC by transplanting both cell types into specific tissues of interest within immunocompetent recipient mice. Here, we have conducted such studies, finding that both hIPSC and hESC engraft robustly and share similar cell fates post-transplantation when incorporated into the immunocompetent, adult mouse brain. Within brain structures, transplanted cell distribution followed a very specific pattern, suggesting the existence of distinct microenvironments that offer different degrees of permissibility for engraftment. Transplanted hIPSC and hESC migrated and differentiated into a wide variety of cell types, including most prominently NeuN+ neurons, but also oligodendrocytes and microglia, whereas no astrocytes were found. They also appear to have integrated into the host blood vasculature and pia mater. Importantly, transplanted hIPSC and hESC behaved much the same in the recipients. In both cases, at more than 1 year (15 months) post-transplant, the undifferentiated human stem cells exhibited no teratoma-forming activity, no evidence of even preneoplastic lesions, and there were no cells located at off-target locations outside the central nervous system (CNS). In contrast, allotransplantation of mouse ESC into the mouse brain did induce, to a low degree, the formation of teratoma. It is possible that in the xenotransplantation experiments, a cross-species response eliminated the tumor-forming capacity of the overall populations of injected cells, as we did find that transplanted mouse ESC led to tumor formation. Both hIPSC and hESC exhibited high and similar rates of fusion with host cells. These data overall shed significant new light on the behavior of human stem cells following transplantation into the adult brain.
Materials and Methods
hESC culture and preparation for transplantation
H9 (WA09), H1, and HSF6 hESC were obtained from the National Stem Cell Bank. GFP-labeled mouse ESC (LB10; GSC-5003) were obtained from MTI-Global Stem. hESC were propagated in six-well plates on an irradiated mouse embryonic fibroblast feeder layer in a medium containing Dulbecco's modified Eagle's medium:F12 (DMEM:F12; Hyclone), 15% KnockOut Serum Replacement (Invitrogen), 10 ng/mL recombinant human bFGF (R&D Systems), supplemented with nonessential amino acids,
hIPSC production
We generated three independent hIPSC cell lines derived from the same parental human dermal fibroblasts (HDFs; Cell Applications, Inc.). A fourth hIPSC cell line (Thomson Laboratory) was used as a control. HDFs were cultivated in DMEM with 10% fetal bovine serum, 4 mM
hIPSC characterization
Alkaline phosphatase staining was performed using the Vector Blue Alkaline Phosphate Substrate Kit (Vector Laboratories). Live cell immunostaining was performed using TRA-1-60 primary antibody (1:100, MAB4360; Millipore) with goat anti-mouse IgM secondary antibody (1:100, A21426; Invitrogen). For teratoma production, 1.5 × 106 cells were suspended in 100 μL phosphate-buffered saline (PBS) containing 30% hESC-qualified Matrigel (BD Biosciences) and injected subcutaneously into the hind limb of NOD/SCID/IL-2Rγ−/− mice (n = 4 mice per cell line). Teratomas were fixed with formalin, embedded in paraffin, cut into 10 μm sections, and stained with Hematoxylin and Eosin (H&E) for analysis.
Reverse transcription–polymerase chain reaction
RNA was extracted from cultured cells using the RNeasy Mini Kit (Qiagen). cDNA was produced using Superscript III First-Strand Synthesis System (Invitrogen). Reverse transcription–polymerase chain reaction (RT-PCR) was performed using the GoTaq Flexi DNA polymerase (Promega) using primers for human endogenous POU5F1, endogenous SOX2, LIN28A, REX1, and NANOG [42]. Primers for GAPDH were designed using NCBI Primer Blast (Fwd: 5′-TGACGCTGGGGCTGGCATTG, Rev: 5′-GGCTGGTGGTCCAGGGGTCT).
Cell transplantation
We transplanted hESC and hIPSC into the lateral ventricles of 41 Swiss Webster mice (20 hESC and 21 hIPSC). Mice (25–35 g, 6–8 weeks old) were anesthetized with 4% isoflurane to induce and 2% to maintain the surgical plane. Before surgery, reflexes were checked by pinching the hind paws and when insensitive to pain, heads were immobilized within a stereotaxic frame in flat skull position. The eyes were protected with Puralube Vet Ointment. A two-centimeter midsagittal skin incision was made on the scalp to expose the skull. The coordinates for the lateral ventricle location were determined based on the Paxinos and Franklin adult mouse brain atlas [43]. A hole was drilled through the skull at the appropriate coordinates, and a glass capillary micropipette stereotaxically advanced so that the internal tip of the pipette was located within the right lateral ventricle. The micropipette had a 50 μm diameter tip and was filled with the stem cells in aCSF solution. The number of transplanted cells was quantified using a hemocytometer and 200,000–250,000 cells in a total volume of 1 μL were transplanted in each animal. The stem cell solution was slowly injected into the brain. The micropipette was kept at the site for an additional 4 min before being slowly withdrawn. The wound was cleaned with 7.5% povidone–iodine. The skin incision was closed with stainless steel wound clips. After surgery, an analgesic (buprenorphine hydrochloride, 0.1 mL of 0.3 mg/mL) and an anti-inflammatory (meloxicam, 0.1 mL of 1.5 mg/mL) were subcutaneously injected. For BrdU studies, BrdU (50 mg/kg I.P.) was injected each day during four weeks after cell transplantation. The animal studies were approved by the UC Davis IACUC.
Immunostaining
Adult mice were perfused intracardially with PBS followed by 4% paraformaldehyde in PBS (PFA). Brains were removed, postfixed for 24 h in 4% PFA and 40 μm coronal slices prepared on a vibratome (Leica). Free-floating sections were blocked in 10% donkey serum (Gibco), 0.1% Triton X-100, and 0.2% gelatin (Sigma). Sections were incubated for 24 h at room temperature in one of the primary antibodies: mouse human nuclei antibody, clone 235-1, 1:500 (cat# MAB1281; Millipore), rabbit anti-laminin (1:500; Abcam), rabbit anti-NeuN (1:1,000; Chemicon), goat anti-DCX (1:500; Santa Cruz), rabbit anti-S100 (1:500; Abcam), rabbit anti-GFAP (1:1,000; Sigma), rabbit anti-Olig2 (1:500; Abcam), rabbit anti-Sox10 (1:500; Abcam), rabbit anti-nestin (1:200; Chemicon), rabbit anti-GAD67 (1:1,000; Abcam), rabbit anti-parvalbumin (anti-PV, 1:1,000; Swant), goat anti-calretinin (anti-CR, 1:1,000; Swant), rabbit anti-calbindin (anti-CB, 1:2,000; Swant), rabbit anti-Ki67 (1:1,000; Abcam), rabbit anti-OCT4 (1:1,000; Chemicon), rabbit anti-SSEA4 (1:1,000; Chemicon), rabbit anti-alpha smooth muscle actin (1:200; Abcam), chicken anti-BrdU (1:1,000; Abcam), and rabbit anti-DsRed antibody (1:100; Clontech Living Colors DsRed). Sections were rinsed and incubated for 1 h in one of the secondary antibodies: Cy2-, Cy3-, or Cy5-conjugated polyclonal anti-mouse/goat/rabbit antibodies (1:100; Jackson Laboratories). Antibodies were diluted in incubation buffer containing 2% normal donkey serum, 0.02% Triton X-100, and 0.04% gelatin. We omitted the first antibody as a control for each immunostaining experiment. TUNEL assay (Roche) was performed as per instructions and included TUNEL-positive (slices treated with DNase I) and -negative controls. All imaging were performed on an Olympus Fluoview Confocal Laser Microscope and analysis performed in Fluoview v.3.3 (Olympus).
Cell quantification
The survival of transplanted cells after transplantation, and the percentage of transplanted cells that expressed cell-specific markers were quantified through confocal microscopy. Brains were perfused and cut as described above. The number of cells that survived transplantation was estimated by counting the number of cells in every third 50 μm-thick section throughout the rostrocaudal axis of the brain, and multiplying this number by three. The percentage of transplanted cells that expressed cell type-specific markers was calculated using at least three animals for each marker. Immunopositive cells were quantified in every six 50 μm-thick section throughout the rostrocaudal axis of the brain.
DsRed animals
Two homozygous males of the strain
Human and mouse Cot-1 probe synthesis
Human Cot-1 DNA (15279-011; Invitrogen) and mouse Cot-1 DNA (18440-016; Invitrogen) were obtained to hybridize to human repetitive AluI and KpnI family DNA elements or to mouse repetitive B1, B2, and L1 DNA elements. Human and mouse probes were labeled using fluorescein-12-2′-dUTP (FITC, 11-373-242-910; Roche) or tetramethyl-rhodamine-5-dUTP (TRITC, 11-534-378-910; Roche) using nick translation (ENZ-42910; Enzo). Four micrograms each of mouse Cot-1 DNA and ligated human Cot-1 DNA were nick translated for 12 h at 15°C. Probes were then ethanol precipitated overnight and resuspended in hybridization buffer (50% formamide, 5× saline-sodium citrate buffer [SSC], 1× Denhardt's, 0.3 mg/mL tRNA, 0.2 mg/mL salmon sperm DNA, 100 μg/mL heparin, 0.1% Tween-20, 0.1% CHAPS, and 5 mM EDTA) at 50 ng/μL.
Fluorescent in situ hybridization on mouse brain sections
Mouse brain sections (5 or 50 μm) were processed as according to Solovei et al. [44] with a few modifications. Sections were unmasked in sodium citrate solution initially for 3.5 min in a microwave, then allowed to cool for 2 min. This was followed by seven repeats of 1 min reheating and 2 min cooling. Sections were then allowed to rest for 15 min in sodium citrate solution, followed by a 5 min wash in 2× SSC buffer. Sections were prehybridized with 750 ng of human and mouse Cot-1 probe (for 50 μm sections) or 500 ng of human and mouse Cot-1 probe (for 5 μm sections) for 1 h at room temperature. Sections were then denatured with probe for 5 min at 80°C, followed by hybridization for 2 days at 37°C. Following hybridization, sections were washed 3× for 5 min each in 2× SSC buffer at 37°C, and then 3× for 5 min each in a high-stringency wash (0.1× SSC) at 60°C. Slides were then costained with DAPI and sealed. For analysis, two hESC-injected mouse brains (n = 33 5 μm section images; n = 14,048 cells) and one hIPSC-injected mouse brain (n = 19 5 μm section images; n = 7,959 cells) were used.
Results
Generation and validation of hIPSC lines
hIPSC lines were generated from HDFs using retroviral transduction with POU5F1/OCT4, SOX2, KLF4, and MYC. The hIPSC were morphologically indistinguishable from hESC, forming compact colonies with well-defined borders and high nucleus-to-cytoplasm ratio (Fig. 1A). They expressed markers of pluripotency, including alkaline phosphatase (Fig. 1B), TRA-1-60 (Fig. 1C), LIN28A, REX1, NANOG, and endogenous OCT4 and SOX2 (Fig. 1D). The hIPSC shared the normal female karyotype of the parental HDFs (Fig. 1E). When injected subcutaneously into immunodeficient mice, the hIPSC formed teratomas containing endodermal, mesodermal, and ectodermal lineages (Fig. 1F). In addition to these three hIPSC lines that we generated, for some studies we used an additional fourth hIPSC line produced by the Thomson Laboratory.
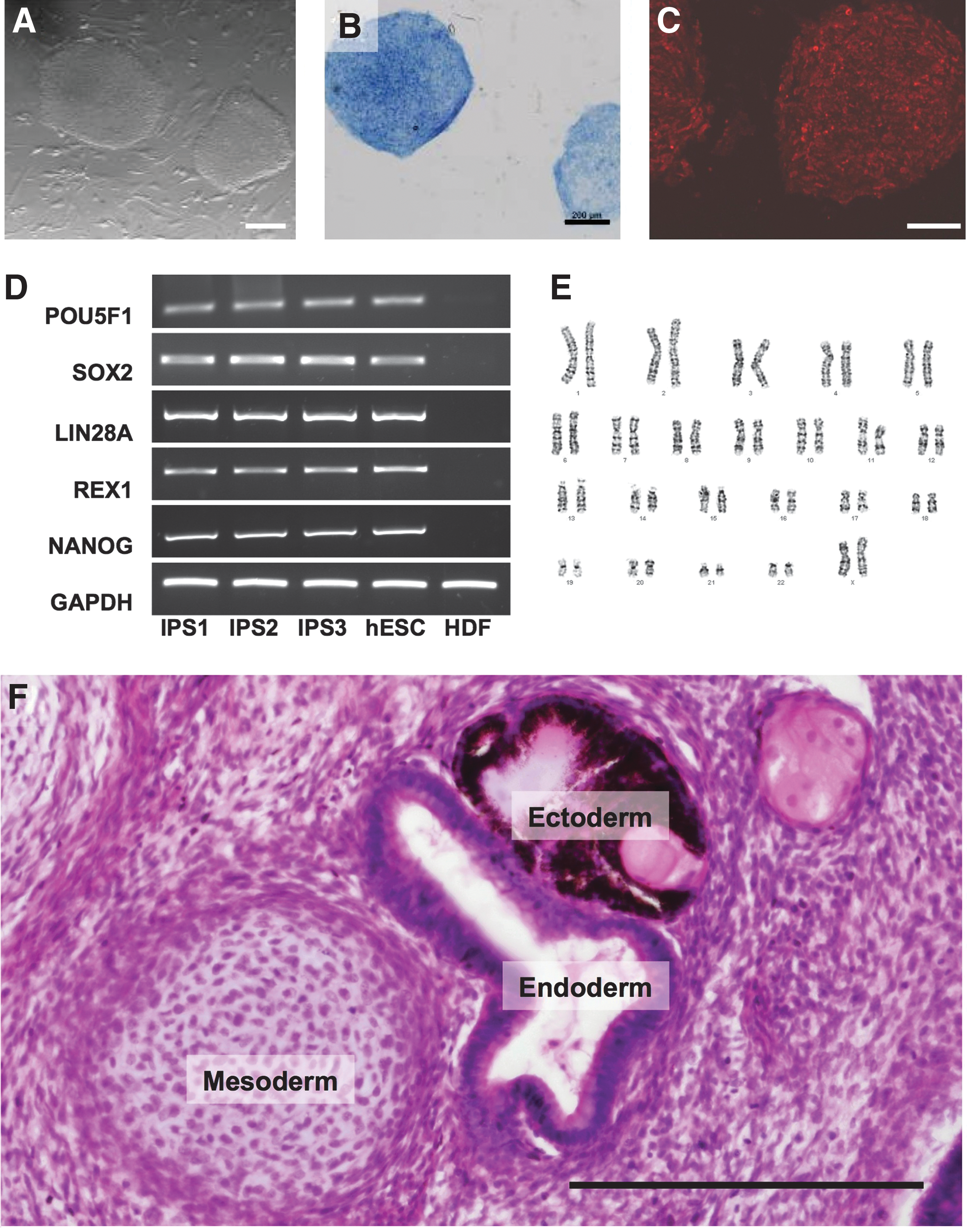
hIPSC exhibit hESC-like characteristics. hIPSC were assessed for hESC properties, including morphology
hESC and hIPSC exhibit high rates of integration in mouse brain
To examine the relative behavior of undifferentiated hESC and hIPSC following transplant into the adult brain, we used stereotactic injection to transplant undifferentiated hESC or hIPSC into the lateral ventricle of the adult mouse brain. Approximately 200,000–250,000 human cells were injected in the right lateral ventricle for each of the hESC and hIPSC lines. Transplanted hESC and hIPSC, as well as their derivatives were identified using immunostaining with a human-specific nuclear antibody [45 –47] and confocal microcopy. Human-specific nuclear antibody labeled the nucleus in some cases and always the cytoplasm of the human cells as previously shown [45 –47]. No staining was evident in untransplanted mouse brain. Overall, we found that both hESC and hIPSC had similar capacities to penetrate into the brain parenchyma after transplantation. Further analysis of the H9 hESC and for one of the hIPSC lines generated from HDFs was conducted. We found that although hESC and hIPSC had similar migration capacities, the rate of survival after transplantation was higher for hESC than for hIPSC. Four weeks after transplantation, the measured net engraftment of hESC was 151% ± 9.4% versus 39.47% ± 9.2% of hIPSC (P < 0.001).
Evidence of a higher in vivo rate of hESC versus hIPSC proliferation
Since we found a greater number of hESC than were injected, we predicted that some level of transplanted cell proliferation occurred. We performed hESC and hIPSC transplantations and injected the animals with BrdU every day during four weeks (Fig. 2). We quantified the number of BrdU+ cells and found that 13.85% ± 0.3% hESC were BrdU+, whereas only 5.15% ± 1.5% hIPSC were BrdU+ (P < 0.001). The transplanted hESC and hIPSC and their derivatives did not exhibit overt signs of being unhealthy. More specifically, we did not observe cell shrinkage, blebbing, or apoptotic body formations that are characteristic of unhealthy or dying cells [48]. We performed TUNEL staining one, two, four, and 12 weeks after transplantation and we did not detect any TUNEL+ transplanted cells (not shown). Nevertheless, in some cases one week after transplantation we observed a small amount of immunopositive debris that was located near the ventricles at the level of the striatum and the hypothalamus that we interpreted as remnants of transplanted human stem cells that had died much earlier, shortly after transplantation (not shown). Therefore, we concluded that the rates of proliferation after transplantation differed between hESC and hIPSC, and that the rate of cell death was not significantly affecting net cell rates of survival.

In vivo proliferation characteristics of transplanted hESC and hIPSC. BrdU (red) was injected each day during 4 weeks after hESC
Migration and regional engraftment of transplanted hESC and hIPSC follow a region-specific pattern
After transplantation into the lateral ventricle, some of the human stem cells distributed throughout the ventricular system, including lateral ventricles as well as the third and fourth ventricles, and the central canal. Human cells were located as far as 3 mm from any ventricle both within one-week post-transplantation and at later time points. These data suggest that the migration of transplanted cells into the parenchyma is nearly complete by one week post-transplantation. One week after transplantation, hESC and hIPSC within the brain parenchyma exhibited an immature morphology with a rounded soma. Two weeks after transplantation, cells acquired a more mature morphology, including the presence of multiple processes protruding from the soma and directed toward specific surrounding brain areas. By week four, evidence of differentiation was clearly apparent, and at week 12 post-transplantation, cells had achieved the maximum morphological complexity we observed. By 6, 12, and 15 months after transplantation, hIPSC derivatives survived and their morphology and localization were similar to that observed 4 weeks after transplantation (Supplementary Fig. S1; Supplementary Data are available online at
Qualitative analysis indicates that hESC and hIPSC colonized similar areas of the brain after transplantation in the ventricular system. They appeared to preferentially colonize some specific areas of the cerebral cortex, including cingulated cortex, piriform cortex, and occasionally sensory–motor cortex. Stem cells also colonized the olfactory bulbs, hippocampus, septum, thalamus, hypothalamus, corpus callosum, subfornical organ, substantia nigra, and other areas (Figs. 3 –5). A small number of human cells were found to have engrafted in the spinal cord in some animals as well. Occasionally we also detected a very small number of human cells in the striatum (four or five cells in one single section), whereas the septum, located nearby the striatum, showed a relatively large number of human cells (up to 500 cells) in the same brain section. Within brain structures, transplanted cell distribution followed a very specific pattern. For example, in the habenular nuclei of the thalamus, cells were more abundant in the lateral nucleus and had preference for the area separating both nuclei (lateral and medial) suggesting the existence of two microenvironments within the habenula with different degrees of permissibility for engraftment (Fig. 3). The most permissive area of the brain for engraftment and survival of both hESC and hIPSC was the subfornical organ, where hundreds of human cells were observed in each single tissue section from rostral to caudal in each of the experiments. In some of the recipient animals we found derivatives of transplanted cells forming part of the subventricular zone, where the adult host stem cells reside. Human cells were not found in the cerebellum. Since all the cells were injected into the same location (lateral ventricle), the differences in engraftment in distinct locations may reflect variable subdomains that are more amenable to engraftment of pluripotent stem cells.

hESC and hIPSC derivative distribution at 4 weeks after transplantation. Most of the hESC
No teratomas or precancerous lesions originated from hPSC more than 1 year post-transplantation
Following transplantation, both types of human stem cells appeared to have differentiated into many types of cells, including neurons, glial cells, ependymal cells surrounding the ventricles, blood vessel cells, and cells in the epithelium surrounding the surface of the brain. We injected BrdU each day after transplantation, and on day seven we found that only a very small number of human cells (0.2%), mostly with glial morphology, were BrdU+. Most of these cells were located in regions of white matter, such as in the corpus callosum and the hippocampus fimbria. None of the transplanted cells with neuronal morphology expressed BrdU. No nests of BrdU+ human cells were evident. At one week and 12 weeks post-transplantation, we performed immunostaining with the proliferative marker Ki67 and did not observe Ki67+ human cells. No Oct4+ or NANOG+ cells were evident as well. We performed H&E staining in tissue of transplanted brains at 4 and 12 weeks, plus 6 and 15 months after transplantation of hESC or hIPSC and did not observe tumor formation in the brain or outside the brain (Supplementary Fig. S2). These data suggest that transplanted, undifferentiated hIPSC and hESC are not inherently tumorigenic and pluripotent cell tumorigenesis may be context dependent with the adult brain being nonpermissive.
To attempt to quantify engraftment of human cells we conducted quantitative polymerase chain reaction (qPCR) for human- and mouse-specific genomic DNA for human ERV-3 and mouse GAPDH, respectively, on genomic DNA isolated from recipient brains. We verified that this assay is sensitive enough to detect five human cells among 50,000 mouse cells (290 ng gDNA) in an in vitro context with pure DNA. In mice that received one of the three hIPSC lines, we were able to detect human cell engraftment by the qPCR assay in diencephalon and hippocampus (Supplementary Fig. S3A). While the detected levels of human DNA were relatively low, we did not observe detectable background PCR amplification in the absence of added DNA from transplanted brain samples, suggesting our qPCR detection of human DNA represents bona fide hIPSC engraftment. Most likely we predict that the apparent low level of human DNA in the mouse brain was due to issues related to the prior fixation of the brain as our control in vitro experiments used purified human cellular DNA from culture never subject to fixation.
To test if transplanted hIPSC traveled to off-target regions outside the brain, we also performed the same qPCR. We perfused two mice injected with hESC and two injected with hIPSC at 12 weeks after transplantation, dissected kidneys, lungs, heart, and liver, performed qPCR, and did not detect human cells at these off-target locations (not shown). We analyzed the same organs plus the spleen from mice one year after injection (Supplementary Fig. S3B). We included human DNA spiked in at two different concentrations (equivalent to 5 or 50 human cells among 50,000 mouse cells) as a positive control to verify that we can detect low levels of human DNA (Supplementary Fig. S3B). We also performed a pathological analysis of kidneys, lungs, heart, liver, and spleen in animals injected with hIPSC after one year and did not detect any tissue abnormality (not shown). Therefore, hPSC transplanted in the ventricle do not appear to exit the CNS in detectable numbers.
Mouse ESC can generate teratoma/teratocarcinoma-like tumors after transplantation in the mouse brain
We hypothesized that a potential explanation for the lack of teratoma formation after transplantation of hIPSC and hESC in the mouse brain could be due to the heterologous, cross-species nature of this transplantation. To test this hypothesis we transplanted mouse ESC labeled with enhanced green fluorescent protein (EGFP) into the lateral ventricles of mice using the same protocol we had previously used for hESC.
We transplanted mouse ESC into 11 mice and in nine of them cells survived and no teratoma were generated. Transplanted cells penetrated the brain parenchyma, however, did not migrate as much as in the case of the human cells and we observed a reduced number of cells, compared with the transplanted human cells in the cerebral cortex and hippocampus (Fig. 4A). In the other two transplanted brains, a tumor developed. In one, by H&E staining, the tumor presented as a primitive neuroectodermal-kind tumor with neural rosette-like structures; however, it was not teratoma-like in overall appearance. The final brain presented with a teratoma-like tumor and adjacent to it, there was a collection of small cells clustered together with dark nuclei that presented as a malignant undifferentiated tumor. We classified this tumor as most likely a teratocarcinoma (Fig. 4B, C), but further studies will be needed for definitive determination. Overall, we determined that in some cases, teratomas were generated after transplantation of mouse ESC, supporting our hypothesis that hPSC may fail to form tumors in the mouse brain due, at least in part, to cross-species transplantation.
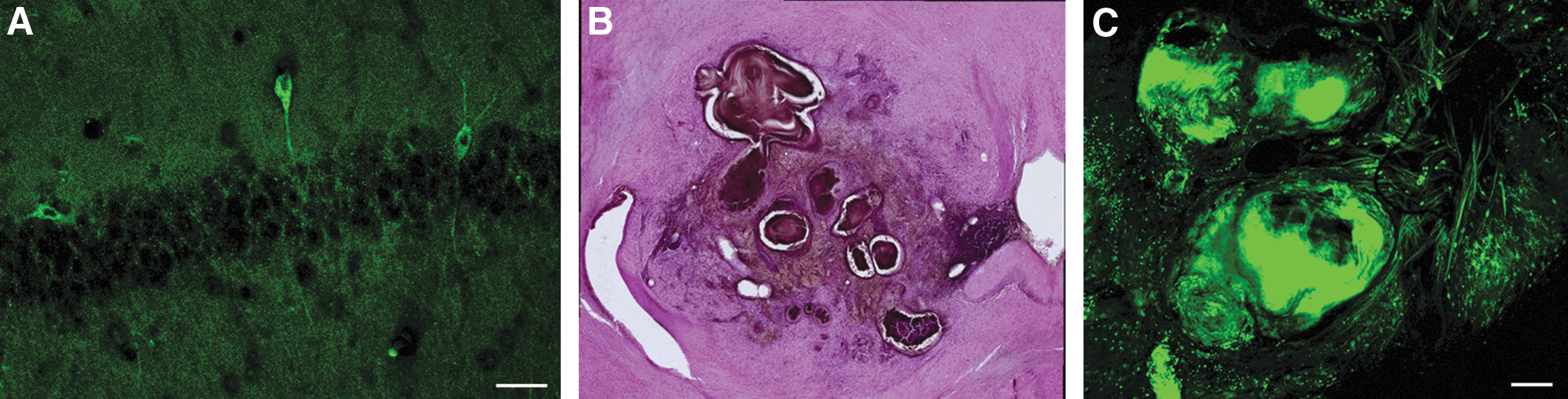
Outcomes from mouse ESC transplanted into the lateral ventricle of mice.
Generally similar fates of transplanted hIPSC and hESC
We costained tissue with the human cell antibody and with specific cell markers and took confocal images to determine the cell fate of the transplanted hESC (Fig. 5) and hIPSC (Fig. 6). Transplanted cells were integrated into the wall of blood vessels (Fig. 3, arrows) and into the pia (Fig. 3, arrowheads) in most areas of the brain that were colonized. Cells in both structures expressed laminin (Figs. 5A and 6A), a protein present in the basal lamina, one of the layers of the basement membrane that lines the surface of the brain, and the interior surface of blood vessels. The transplanted cells or their derivatives were abundant in blood vessels in most of the areas of the cerebral cortex and in the thalamus. The size of the vessels with human cellular constituents varied from very small capillaries (5–10 μm) to wider vessels (30–50 μm). Transplanted cells also acquired the position, morphology, and marker expression of ependymal cells. Human cuboid cells were evident surrounding the ventricles and stained positive for nestin, a marker of stem and progenitor cells (Fig. 6B).
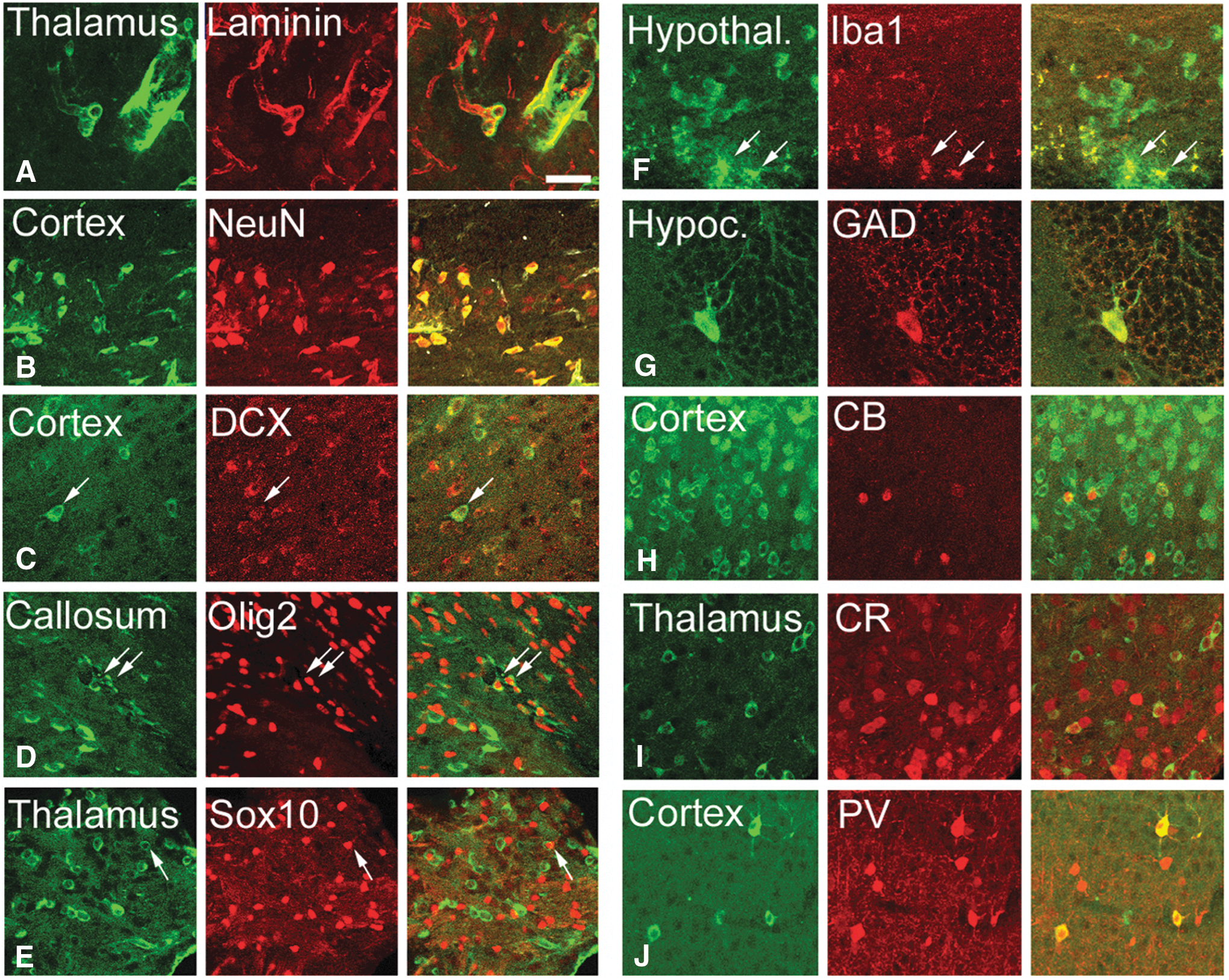
hESC derivative cells expressed specific cell markers (red) four weeks after transplantation.
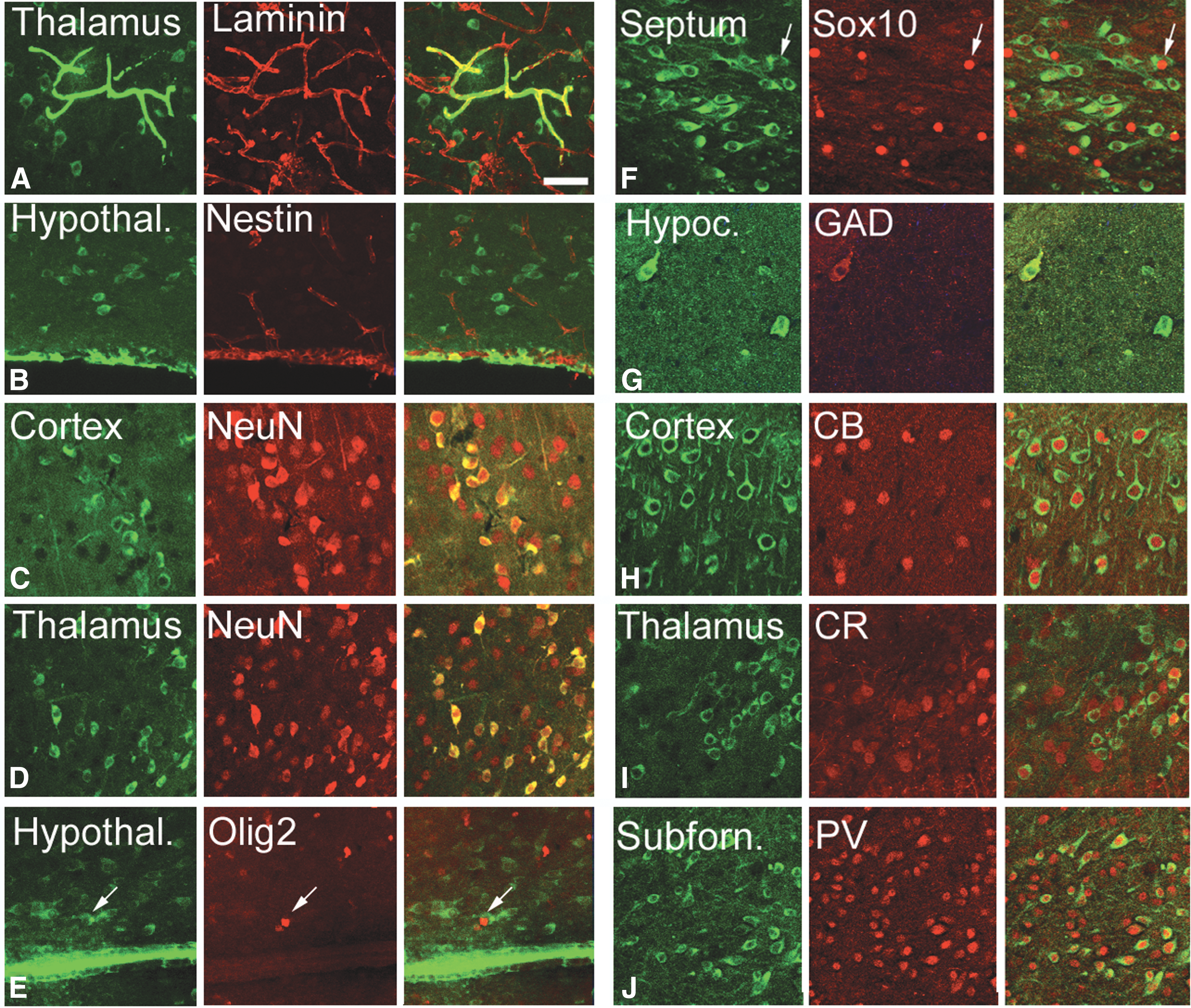
hIPSC derivative cells expressed specific cell markers (red) four weeks after transplantation.
Our results indicate that the rates of differentiation into specific nervous system cells of both transplanted hESC and hIPSC were mostly similar. We stained cells for the mature neuronal marker NeuN, which is expressed by all neurons in the nervous system, except cerebellar Purkinje cells, olfactory bulb mitral cells, and retinal photoreceptor cells [49]. We also labeled cells with the immature neuron/neural precursor cell marker DCX [50], the oligodendroglial markers Olig2 [51] and Sox10 [52], and the astroglial markers S100 [53] and GFAP [54] (Figs. 5 and 6; Supplementary Table S1). We observed that, four weeks after transplantation, 82.9% of transplanted hESC derivatives were NeuN+ neurons (953 cells), 5.4% were Olig2+ oligodendrocytes (352 cells), and 4.2% Sox10+ oligodendrocytes (182 cells). Cell numbers in parentheses represent total numbers of cells couned per marker. Fifty-nine percent were DCX+ cells (763 cells; not shown). Some cells were double positive for multiple markers. A similar proportion of hIPSC adopted the same phenotype after transplantation: 78.3% were NeuN+ neurons (299 cells), 56.4% DCX+ cells (1,083 cells), 5.5% Olig2+ oligodendrocytes (1,916 cells), and 2.9% Sox10 (793 cells). None of the human cells expressed the astrocyte markers S100 (hESC: 0/217 cells; hIPSC: 0/1,083 cells) or GFAP (hESC: 0/321 cells; hIPSC: 0/1,306 cells). A small number of the hESC or hIPSC surrounding the ventricles was positive for Iba1, a marker for microglia (Fig. 5F). Some of the transplanted cells that expressed NeuN also expressed the GABA synthesizing enzyme GAD67, suggesting that the transplanted cells differentiated both into GABAergic and non-GABAergic neurons (Figs. 5 and 6). Non-GABAergic neurons included projection neurons such as cortical pyramidal cells. Our data show that the human GABAergic neurons were mostly interneurons based on the expression of the calcium-sequestering proteins CB, CR, and PV. Cells that were located in the thalamic reticular nucleus expressed PV, cells in the paraventricular nucleus expressed CR, and cells in the nucleus reuniens expressed CB [55]. None of the transplanted cells expressed cell markers that normally are absent in brain such as alpha smooth muscle actin (not shown).
Transplanted hESC and hIPSC both exhibit substantial rates of cell and nuclear fusion with murine host cells and nuclei
To test the possibility that the expression of specific brain cellular markers by the transplanted cells could be, in part, a result of cell fusion between the transplanted cells and the host murine cells, we transplanted hIPSC into mice that express DsRed under the actin promoter. If cell fusion occurs, we expected to find that individual, transplanted human cells (stained green) fluoresce both green and red at the same time. We quantified the number of red cells and the number of DAPI+ nuclei and calculated the percentage of host cells that expressed DsRed. Despite an actin promoter that would be predicted to be fairly ubiquitous in driving expression, not all brain cells were DsRed+. The percentage of DsRed+ host cells was fairly similar in different regions of the brain with an average of 39.5% (cerebral cortex: 41.4% ± 3.9%, granular layer of the hippocampus: 35.5% ± 4.7%, molecular layer of the hippocampus: 51.1% ± 6.9%, subfornical organ: 31.8% ± 2.3%, septum: 37.5% ± 1.0%, thalamus: 40.1% ± 6.0%) (Fig. 7B).
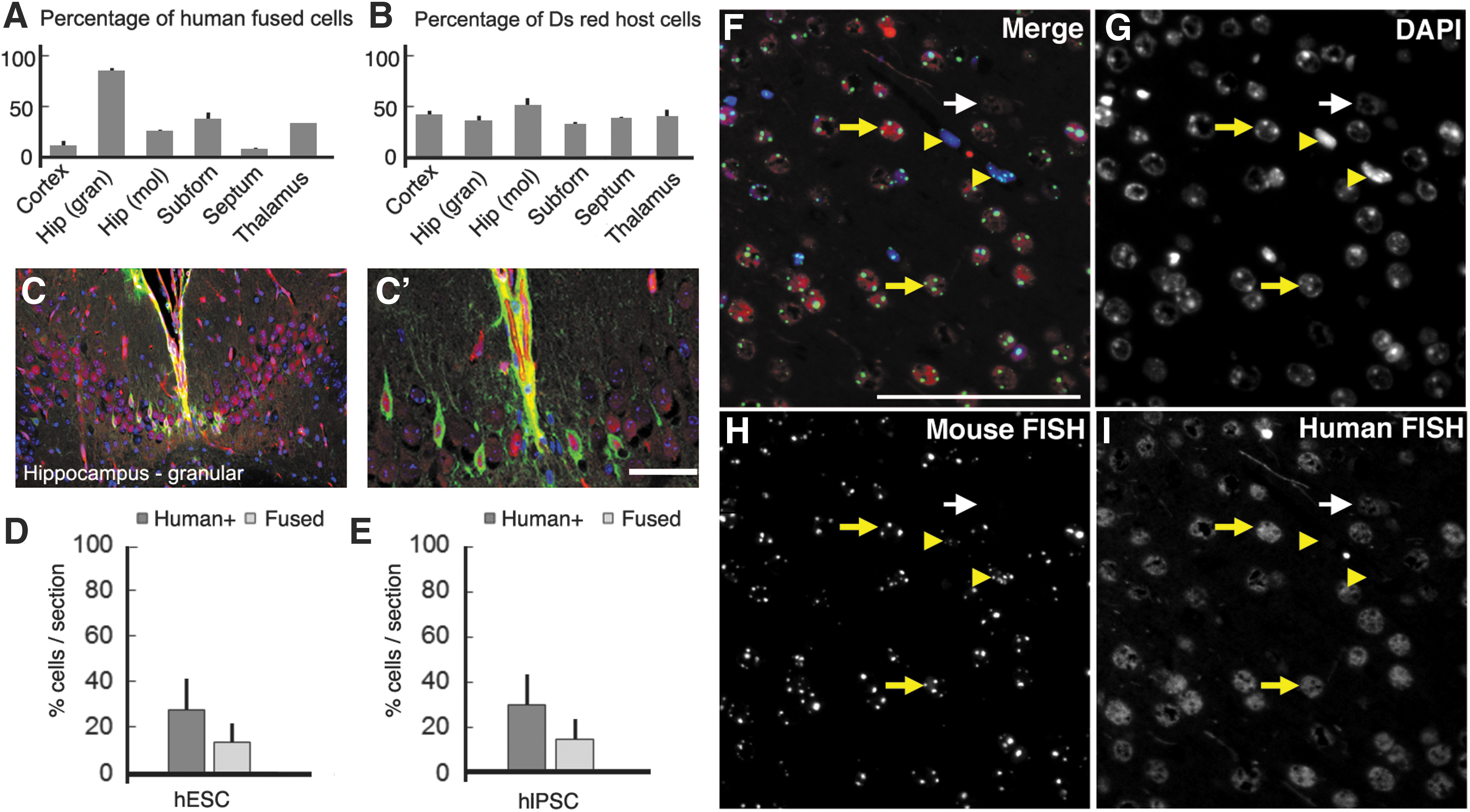
hESC and hIPSC fusion with the host cells after transplantation.
We found evidence of substantial human:mouse cell fusion in the transplanted brains. Furthermore, the percentage of transplanted cells that colocalized with Ds-Red was very different from region to region. For example, while in the cortex and the septum only 10% and 7% of the transplanted hIPSC derivatives colocalized with DsRed, respectively, in the granular layer of the hippocampus 85% of the hIPSC derivatives were DsRed+ (cortex: 10.8% ± 4.4%, granular layer of the hippocampus: 85.5% ± 2.0%, molecular layer of the hippocampus: 25.5% ± 0.4%, subfornical organ: 37.1% ± 6.0%, septum: 7.0% ± 0.8%, thalamus: 32.04% ± 0.2%, and hypothalamus: 28.0% ± 0.6%) (Fig. 7A). We looked at the number of nuclei inside each double colocalized cell and interestingly we only found single nucleated cells (Fig. 7C, C′). Based on recently published data we hypothesized the nuclei fused soon after cytoplasmic fusion [56]. Transplanted hESC and their derivatives fused with host cells at a similar rate as hIPSC (Fig. 7D). We concluded that at least some of the transplanted hPSC fused with the host cells, but not all of them, and that the fusion rate was specific to the region of the brain where the cells were present. Therefore, the cell-specific marker expression of the transplanted cells may be in some cases a result of hPSC host cell fusion rather than de novo differentiation of human stem cells, whereas in other cases it may be a result of cell autonomous differentiation.
To confirm nuclear fusion between host and transplanted cells, we performed fluorescent in situ hybridization to detect species-specific mouse Cot-1 DNA (repetitive B1, B2, and L1 elements) and human Cot-1 DNA (AluI and KpnI elements) sequences within single nuclei [57,58] (Fig. 7F–I and Supplementary Fig. S4). Probe specificity was confirmed by cohybridizing human Cot-1 and mouse Cot-1 to human HeLa cells and mouse 3T3-D6 cells as positive controls (data not shown). We measured the percent of fused mouse and human nuclei in the cerebral cortex of hESC-injected and hIPSC-injected murine brains. hESC nuclei displayed a very similar rate (P = 0.580) of nuclear fusion (56.1% ± 19.7% of hESC) relative to hIPSC (59.5% ± 24.4% of hIPSC). In the regions analyzed, we determined that 13.6% ± 9.34% of host cells had fused nuclei with hESC (Fig. 7D) and 15.1% ± 9.81% with hIPSC (Fig. 7E), although there was not a significant difference in the amount of fused nuclei between hESC and hIPSC (P = 0.582). Therefore, overall we conclude that there was significant hPSC fusion with mouse brain cells, and that following fusion, the resulting individual cells are mouse–human hybrid cells containing both sets of chromosomes.
Transplanted cells expressed synaptophysin
We examined whether the transplanted cells established synaptic connections. Synaptophysin is a synaptic vesicle glycoprotein that is present in all the neurons in the brain and spinal cord that participate in synaptic transmission [59]. We found that four weeks after transplantation all neuronal derivatives of the transplanted hIPSC and hESC expressed synaptophysin puncta along their processes, indicating their participation in synaptic transmission (Fig. 8).
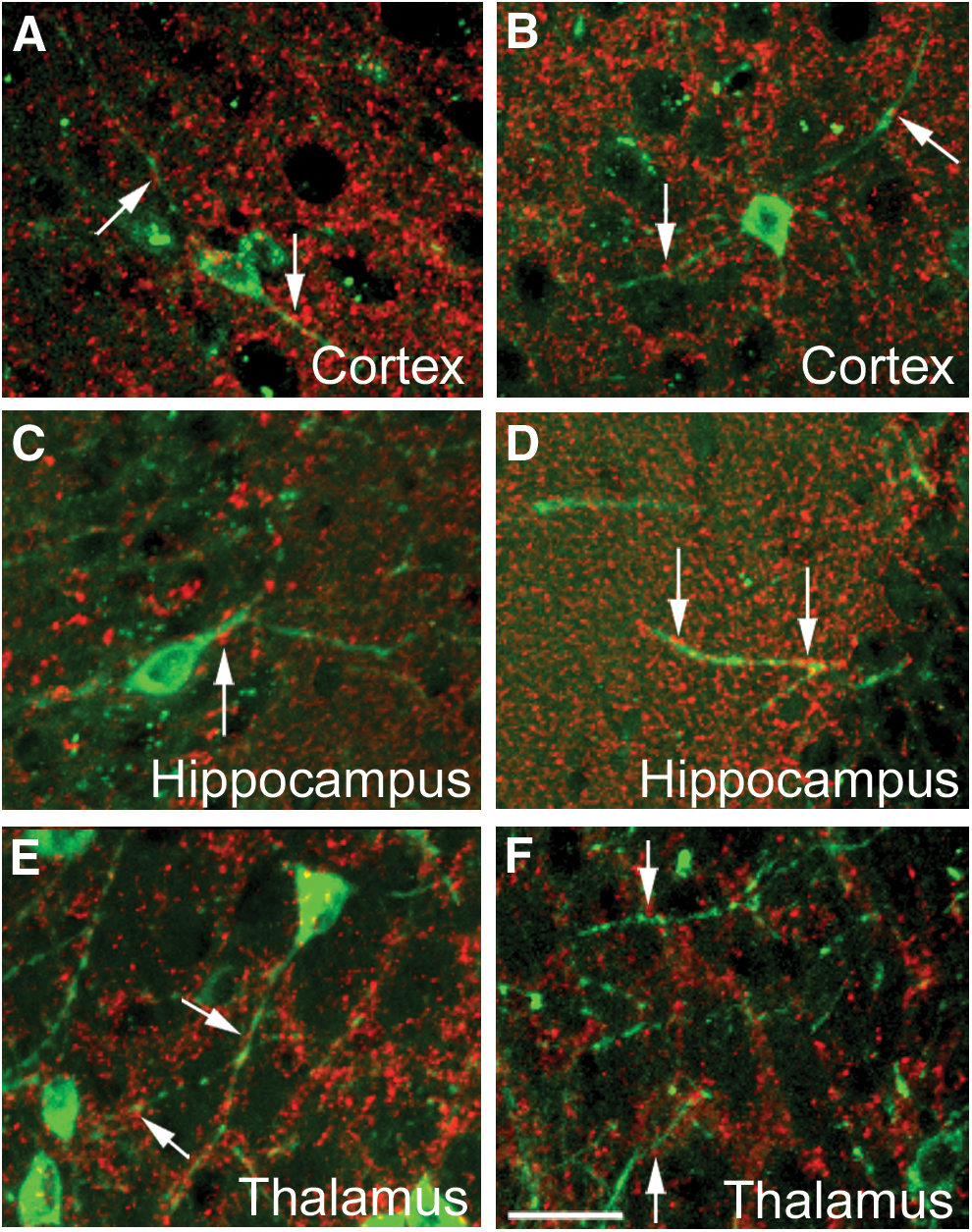
Synaptophysin expression in transplanted hIPSC derivatives. Transplanted hESC
Similar properties of four distinct transplanted hIPSC lines
Our results regarding the safety and engraftability of hIPSC were obtained using hIPSC from three different lines derived from the same parent fibroblasts. In addition, we performed experiments using a fourth hIPSC line derived by a different laboratory from another parent fibroblast (iPS(IMR90) clone #4; Thomson laboratory) and obtained similar safety and engraftment results [4] (Supplementary Fig. S5). Multiple hESC lines (H9, H1, and HSF-6) behaved in similar manners following transplantation as well.
Discussion
We found mostly similar properties of undifferentiated hIPSC and hESC transplanted into the mature, immunocompetent mouse brain. Undifferentiated hIPSC and hESC engrafted, and migrated in vivo in an analogous manner. However, the rate of proliferation of hESC was relatively higher than that of the hIPSC. Even for hESC we observed a low rate of in vivo proliferation and for both transplanted hESC and hIPSC no off-target cells were observed outside the central nervous system. Surprisingly, no teratoma was observed in our studies as well with either undifferentiated hESC or hIPSC transplantations to the adult mouse brain.
Why did teratoma fail to form in our allotransplantation studies? The model that we favor is that hESC and hIPSC have inherent low or absent tumorigenic potential in the context of the mouse adult brain, which may have low tumorigenic permissiveness for hESC and hIPSC. In this model, the adult brain may be an environment in which hESC and hIPSC are generally not able to proliferate significantly more than a few days after transplant, perhaps due to potent differentiation-inducing signals. In addition, the presence of substantial fusion between transplanted stem cells and recipient cells, which has been noted in the past [56,60], could influence the apparent fate of transplanted cells, including potentially blocking tumorigenicity. Cell fusion could potentially complicate the clinical utility of transplantation of stem cells or their derivatives in human patients; however, since we did not observe teratoma or other tumors with hIPSC transplantation, we believe that such fusion is not inherently tumorigenic. As observed in a recent study, we found that most fused cells expressing both human and mouse markers contained only a single nucleus (synkaryon) suggesting a relatively high rate of nuclear fusion as well [56].
Another possible explanation for the absence of teratoma in the immunocompetent recipient mice in our studies is immune system-mediated killing of specific hESC and hIPSC that contain teratoma-forming activity. While in some past teratoma studies, the absence of teratoma was theorized to be due to immune cells killing the transplanted cells [61]; in our studies in the brain we have documented a substantial survival and engraftment rate of transplanted human pluripotent cells eliminating the possibility of large-scale rejection of transplanted human cells. However, it remains formally possible that a small subset of hESC and hIPSC that possessed all teratoma-forming activity died during or after transplantation. The notion of the cross-species nature of our transplantation protocol being in part responsible for a lack of tumorigenicity is supported by our observation that mouse ESC at least sometimes led to teratoma or teratocarcinoma formation in the recipient mouse brain. These results indicate that the murine brain environment can be permissive for tumor formation. This may be because of a differential immune response built against stem cell belonging to the same versus different species. The different genetic background (C57Bl6/N) of the mouse ESCs from that of the recipient mice (Swiss Webster) may have also played some role in only observing teratoma in a few mice in that context. Based on these data allotransplantation experiments could be used as a preclinical predictor for safety.
The relatively high rate of engraftment we observed with transplanted hESC and hIPSC in immunocompetent murine recipients was also somewhat unexpected. Engraftment with immune tolerance in an allogeneic setting has been observed previously for certain stem cells [62,63], but in our studies, the mechanisms allowing relatively robust survival and engraftment are not entirely clear. One possibility is that our method of injection of human stem cells into the adult mouse brain may have led to minimal if any disruption of the blood–brain barrier and immune stimulation. It is also possible that the specific stem cells that we used or a particular aspect of their preparation rendered them less prone to stimulate an immune response. The use of distinct detection methods may have allowed us to more readily detect human cells as well.
Our studies provide evidence that different regions of the adult brain have inherently distinct properties in terms of their influence on transplanted hESC and hIPSC. While overall the transplanted cells had a high rate of survival, some regions of the brain were more permissive than others for cell engraftment and survival. For example, human cells were present in the septum of all transplanted mice, but not in the striatum and only sometimes in the hypothalamus. The subfornical organ was the brain structure that was most highly populated by the transplanted human cells. The capacity of the transplanted cells to widely penetrate this structure could be related to its location protruding into the third ventricle or alternatively to a more permissive microenvironment. For example, in the habenula, the hESC and hIPSC and/or their derivatives had a tendency to migrate into the boundary between lateral and medial habenular nuclei. Both types of human stem cells differentiated into neurons, glial cells, ependymal cells surrounding the ventricles, and basement membrane cells in blood vessels and in the pia. Most hIPSC became neurons that were excitatory or inhibitory and a very small portion into oligodendrocytes. Generally, the transplanted cells that differentiated into neurons acquired the fate of the surrounding host cells, as was clearly illustrated in the thalamus, where cells that were located in the reticular nucleus expressed PV, cells in the paraventricular nucleus expressed CR, and cells in the reuniens nucleus expressed CB. These data suggest that factors dictating cell specification are still present in the adult brain. The rates of cell fusion also greatly varied depending on the brain region in which the human cells were located suggesting that the local cellular environment is a key factor dictating rates of cell fusion as well.
Interestingly, the transplanted hESC and hIPSC did not produce astrocytes. This result contrasts with the capacity of embryonic neuronal stem cells to generate astrocytes after transplantation into the brain [64]. Given that the hESC and hIPSC both have proven pluripotency in vitro, the simplest explanation for the absence of astrocytes in our studies is that the adult brain lacks the appropriate signals to direct these specific stem cells to that lineage. Alternatively, relative potency may manifest in vivo distinctly from the context of in vitro and teratoma assays. In this way of thinking, the adult brain is formally permissive for astrocyte differentiation from hESC and hIPSC, but in the context of an in vivo environment, hESC and hIPSC nonetheless exhibited strong preferences for producing nonastrocytic differentiated cells. In other words, even pluripotent stem cells, or possibly the specific cell lines used in these studies, may have strong tendencies to produce certain cell types, whereas almost never making others. Alternatively, hIPSC may be unable to fuse with murine astrocytes.
Assessing the behavior of undifferentiated human stem cells is vital for understanding the biology of the cells, their clinical potential following differentiation, and also how residual undifferentiated cells in differentiated populations may behave in patients following transplants. Our study uniquely has documented the specific behavior of undifferentiated stem cells in allo- versus xenotransplantation, indicating that allotransplantation experiments could be used as a preclinical predictor for safety. Our work documenting high rates of hIPSC fusion in vivo also reinforces the importance of this issue for the field. More broadly, our findings encourage future studies of transplantation of undifferentiated hIPSC and hESC into other settings, such as the liver and heart in animal models, which will determine whether the apparent strong long-term safety profile and efficacy of undifferentiated pluripotent stem cells in the xenograft context is unique to the brain as a target organ or manifests in other transplanted tissues. Transplantation of undifferentiated hIPSC into injury models may also yield valuable insights. Overall, such studies will lead to a greater understanding of hIPSC biology.
We concluded that allotransplantation of undifferentiated hIPSC and hESC into the adult mouse brain ventricle yielded substantial populations of engrafted, differentiated human cells. Most human cells were NeuN+ neuronal cells, but other differentiated cell types were present as well. Fusion occurred between a significant number of host and transplanted cells, a phenomenon strongly influenced by microenvironment and that could impact apparent hIPSC marker expression and cellular behavior. At more than 1 year post-transplant, no tumors or precancerous lesions were present. Overall, following transplant into the mature mouse brain, hIPSC appear to be largely functionally equivalent to hESC, but future studies with additional hPSC lines and in particular hESC will help to further clarify their relative properties.
Footnotes
Acknowledgments
The authors thank Jasmin Camacho for technical assistance. They thank Jamie Thomson for supplying hIPSC lines generated in his laboratory. This work was supported by NIH Grants 1R01GM100782 and 1R01GM116919 (to P.S.K.), Shriners grant (to V.M.C.), CIRM Grant RN2-00922-1 (to P.S.K.), CIRM training fellowship TG2-01163 (to A.M., B.L.B., and B.T.K.Y.), and in part by the HHMI:IMBS Fellowship Program Grant 56006769 (to B.T.K.Y.).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.