
Abstract
Human induced pluripotent stem cells (hiPSCs) hold great hopes for application in regenerative medicine due to their inherent capacity to self-renew and differentiate into cells from the three embryonic germ layers. For clinical applications, a large quantity of hiPSCs produced in standardized and scalable culture processes is required. Several groups, including ours, have developed methodologies for scaled-up hiPSC production in stirred bioreactors in chemically defined medium. In this study, we optimized the critical steps and factors that affect hiPSC expansion and yield in stirred-suspension cultures, including inoculation conditions, seeding density, aggregate size, agitation rate, and cell passaging method. After multiple passages in stirred-suspension bioreactors, hiPSCs remained pluripotent, karyotypically normal, and capable of differentiating into all three germ layers.
Introduction
H
Microcarriers provide a high surface-to-volume ratio for hiPSC attachment and expansion [5].
Stirring motion in the microcarrier-based system further minimizes the heterogeneity of the culture environment and distribution of oxygen. There are currently different types of microcarriers with variable cell attachment properties as well as bead-to-bead variability in cell confluency [11]. To further minimize variability in the culture system, other groups have explored microcarrier-free systems with different vessel types [12 –24].
Table 1 provides a summary of suspension-based bioprocessing for hESC and hiPSC expansion, including the current study. The culture vessels mainly fall into five categories: spinner flask with a magnetic stir bar (NDS), spinner flask with a glass stirrer pendulum (Integra Biosciences), spinner flask with a suspended impeller (Corning, Inc.), Erlenmeyer flask on an orbital shaker (Corning, Inc.), and bioreactor systems (DASGIP, BioLevitator™). Specific operational protocols have been developed for these culture vessels with different hiPSC expansion rates. Key parameters that affect cell yield are cell inoculation condition, cell inoculation density, stirring speed, and culture period. In this study, we systematically assessed and optimized these parameters for our microcarrier-free bioreactor system (Fig. 1) with chemically defined medium, mTeSR1. A strong emphasis was placed on the inoculation condition as this is a critical initial step for microcarrier-free systems, where hiPSCs are grown and expanded as aggregates.

Bioreactor setup for suspension culture of hiPSCs. The 100-mL NDS stirred-suspension bioreactor (spinner flask)
CM, conditioned medium; hESC, human embryonic stem cells; hiPSC, human induced pluripotent stem cell.
Currently, two methods, single cell inoculation and aggregate inoculation, are used for the initiation of microcarrier-free suspension culture. For single cell inoculation, dissociated single cells are transferred directly into stirred culture vessel and then aggregates form from the union of individual cells from the stirring/agitation. The aggregate inoculation method employs preformed aggregates from static culture, which are then transferred to the culture vessel for stirred-suspension culture.
Variable cell yields have been reported using the two inoculation methods with some even lower than that from the static culture. In this study, we compared the two methods and developed a highly efficient method for aggregate formation in static suspension before cell inoculation. By using this method, majority of the single cells participated in aggregate formation. We further examined the combinatorial effects of expansion period, aggregation size, agitation rate, and pH on the number of viable hiPSCs in stirred-suspension culture. Our optimized protocol consistently yielded a 12-fold expansion of viable hiPSCs per culture run. The hiPSCs maintained their pluripotent state, differentiation potential, and normal karyotype after multiple expansions and passages in suspension culture. Our efforts contribute to the translation of scalable, chemically defined suspension culture systems toward good manufacturing practice conditions for clinical applications.
Materials and Methods
In this study, the suspension culture system we employed includes 125-mL spinner flasks with a magnetic stir bar (NDS) (Fig. 1). Every experiment related to bioreactor culture was performed in triplicate.
Static culture and maintenance of hiPSCs
We cultured hiPSC lines, 4YA and 4YF, which were derived from infant fibroblasts (BJ cell lines) and reprogrammed using retrovirus four factors (OSKM) with EOS reporter. Both hiPSC lines have normal karyotypes (46, XY) and the cells were obtained from Dr. James Ellis' laboratory at the University of Toronto (Toronto, Canada). Cells were cultured and maintained in chemically defined, feeder-free Matrigel/mTeSR1 system under standard culture conditions (37°C, 5% CO2, 95% relative humidity). We coated 35 and 60-mm culture dishes with hESC-qualified BD Matrigel (BD Biosciences) before cell culture for at least 2 h at room temperature. Cells were grown on these Matrigel-coated dishes containing complete mTeSR1 medium (STEMCELL Technologies).
Cells were passaged every 4 days as small cell clumps through enzymatic and mechanical treatment.
Briefly, cultures were washed once with Dulbecco's modified Eagle's medium (DMEM)/F12 (Invitrogen) and then treated with Dispase (1 mg/mL; STEMCELL Technologies). After incubating the culture dishes for 7–9 min at 37°C, the hiPSC colonies started to curl-up around the edges and Dispase was removed followed by two washes with DMEM/F12. Colonies were scraped from the bottom of the culture dishes and broken into smaller cell clumps by trituration using a 1-mL pipette tip. Cell clumps were then replated onto Matrigel-coated dishes in mTeSR1 medium at a 1:6 split ratio.
hiPSC inoculation in the bioreactor
Inoculation conditions were previously reported to have a significant effect on cell survival and growth characteristics during the initial stages of suspension culture in bioreactors. In this study, we explored three types of hiPSC inoculum: (1) single cells, (2) cell clumps, and (3) sphere-like hiPSC aggregates.
hiPSC aggregate formation
We developed three different methods of generating hiPSC aggregates for suspension culture in bioreactors.
Method 1
Aggregates were formed on poly-
Method 2
Dissociated single cells were grown in static suspension culture condition where cells are cultured in noncoated 35 or 60-mm Petri dishes containing complete mTeSR1 medium supplemented with 10 μM Y-27632. After 3–4 h of static suspension cultures, hiPSC aggregates were observed, and these aggregates were pipetted up and down several times to generate smaller aggregates (25–50 μm) before inoculating them in 100-mL bioreactors.
Method 3
Dissociated single cells were cultured in a 15-mL centrifuge tube containing complete mTeSR1 medium supplemented with 10 μM Y-27632. After 3–4 h, hiPSC aggregates were observed, and these aggregates were inoculated in 100-mL bioreactors. Several steps were taken to achieve the optimized size of aggregates. First, the single cell density we employed for aggregate formation in static condition was around 106 cells/cm2 in mTeSR1 medium supplemented with 10 μM Y-27632. Second, we incubated the suspension cultures in a standard cell culture incubator for 3–4 h, depending on different cell lines. Third, after incubation, we gently pipetted the aggregates about 10 times and broke them into smaller ones. Then, randomly measured 30–60 aggregates under microscope using measuring software.
Aggregate-forming efficiency
To determine the effectiveness of the three methods in forming hiPSC aggregates, we estimated the percentage of viable hiPSCs that participated in aggregate formation by using this following equation:
Dead cells were stained positive for Trypan Blue and counted with a hemocytometer.
For Method 1, dead cells were observed floating in the culture medium and some were also attached to the culture surface. Both viable aggregates and dead cells attached to the culture surface were detached by introducing streams of culture media across the surface with a pipette.
Cell suspension was collected and then filtered through a 40 μm cell strainer. The flow-through contained dead cells that were calculated after centrifugation and resuspension. For Methods 2 and 3, the dead cells were observed floating in the culture medium. Cell suspension was collected and then filtered through a 40 μm cell strainer. The flow-through contained dead cells that were calculated after centrifugation and resuspension.
Removal of dead cells before inoculation
Cell suspension containing hiPSC aggregates and dead cells were collected into a 15-mL centrifuge tube and centrifuged at 300 rpm for 3 min. The supernatant that contained the dead cells was aspirated, leaving the hiPSC aggregates at the bottom of the tube for downstream inoculation in bioreactors.
Suspension culture in stirred bioreactors
For single cell inoculation, single hiPSCs were seeded in stirred-suspension bioreactors at low (1 × 105 cells/mL), medium (2.5 × 105 cells/mL), and high (5 × 105 cells/mL) cell densities, which were calculated from the starting cell number minus the number of dead cells. For other inoculation conditions, either cell clumps or cell aggregates were seeded in the bioreactors at low (∼2 × 104 cells/mL), medium (∼4 × 104 cells/mL), and high (∼8 × 104 cells/mL) densities, which were estimated from the starting cell number minus the number of single cells remaining in the culture dish after cell aggregation. First, we estimated the percentage of viable hiPSCs that participated in aggregate formation by using this following equation:
Dead cells were stained positive for Trypan Blue and counted with a hemocytometer. Second, we estimated live cell density in the form of cell aggregates by using this following equation:
Suspension cultures were passaged every 5 days through enzymatic dissociation with Accutase and the hiPSC inoculation processes were repeated, and new cultures were initiated through the same single cells and aggregation process, respectively, as described above for each passage. Agitation speeds were set at 80, 100, and 120 rpm for low, medium, and high, respectively.
Determination of hiPSC number and viability from bioreactor suspension cultures
Bioreactors were removed from the incubator and briefly swirled to even out the distribution of aggregates before withdrawing 2.0 mL of cell suspension for downstream cell count and viability assessment. Each collected sample was placed in a 35-mm culture dish for observation and photomicrographs. Then, the cell aggregates in the dish were treated with 0.25% Trypsin-EDTA (Invitrogen) for 5–7 min at 37°C for dissociation into single cells. Subsequently, two 100 μL samples were used for cell counting with a hemocytometer and cell viability was determined using the Trypan Blue exclusion assay. The fold expansion at each passage was determined by the cell number at the time of harvest divided by the starting cell number.
pH value measurements
It has been well documented that pH in the culture environment affects cell metabolism and growth. In general, an alkalescent environment supports cell growth, whereas acidic environment induces apoptosis. For each culture period, we monitored the pH of the culture media and used it as an indicator of medium replacement. Beginning from the third day (low and medium densities) or the second day (high density) after cell inoculation, a 5-mL sample was taken from each bioreactor each day. When collecting medium sample, the bioreactor was transferred into the cell culture hood, let it stand still for several minutes, gently aspirated a 5-mL sample from the supernatant medium, and then transferred it into a 50-mL tube. A digital pH meter with a probe (VWR sympHony B10P pH Meter) was used for the pH measurement of the culture media.
In vitro and in vivo hiPSC differentiation
For in vitro differentiation, aggregates grown in the suspension culture were dissociated into single cells and replated on 35-mm agar-coated dishes in a differentiation medium consisting of DMEM supplemented with 20% fetal bovine serum, 10 μM Y-27632, 1 mM
For in vivo differentiation, hiPSC aggregates were harvested and injected into the rear leg muscles of 6- to 8-week-old SCID Beige mice (1–2 × 106 cells per injection). Teratomas consisting of all three germ layers were removed from injection sites after 10–12 weeks, fixed overnight in 4% paraformaldehyde (PFA), embedded in paraffin, sectioned, and examined histologically after staining with Hematoxylin and Eosin.
Immunocytochemistry
Cells were fixed in 4% PFA for 15 min at room temperature, washed three times with 1 × phosphate-buffered saline (PBS), permeabilized with 0.1% Triton X-100 (Sigma-Aldrich) for 15 min at room temperature, and then washed three more times with PBS before blocking with 10% normal serum or 3% bovine serum albumin (BSA) in PBS for 30 min at room temperature to minimize nonspecific binding of antibodies. Fixed hiPSCs were incubated with primary antibodies (Millipore) against pluripotency markers, Nanog, Oct4, SSEA-4, TRA-1-60, and TRA-1-81, at 1:100 dilution, 4°C overnight. Differentiated EBs were incubated with primary antibodies (Sigma-Aldrich) against β-tubulin III (ectoderm marker), smooth muscle actin (mesoderm marker), and α-fetoprotein (endoderm marker) at 1:400 dilution, 4°C overnight. The next day, cells were incubated with the appropriate secondary antibodies, Alexa Fluor 546 goat anti-mouse and Alexa Fluor 594 donkey anti-rabbit (1:200; Invitrogen), for at least 2 h at room temperature and then washed three times with 1 × PBS before imaging with a confocal microscope.
Flow cytometry
The expression of pluripotency markers, Oct4, Nanog, and SSEA-4, of hiPSCs grown in stirred bioreactors was analyzed by fluorescence-activated cell sorting (FACS). Cell aggregates were dissociated into single cells through trypsinization, fixed with 4% PFA, washed three times with PBS, and permeabilized with 0.75% Saponin (Sigma-Aldrich). After two washes with PBS, cells were then resuspended in PBS containing 3% BSA for 30 min at 37°C. Resuspended cells were incubated with the following antibodies (Millipore) for at least 1 h at 37°C: anti-Oct4 (Alexa Fluor 488 conjugate; 1:50), anti-Nanog (FITC conjugate; 1:50), and anti-SSEA-4 (PE conjugate, 1:50 dilution). Mouse IgG Alexa Fluor 488, mouse IgG1 FITC, and mouse IgG3PE were used as isotope controls for the primary antibodies. Flow cytometric analysis was performed using BD FACSAria III.
Karyotype analysis
Karyotype analysis of hiPSCs was carried out using the G-banding method. Briefly, hiPSCs were incubated with a culture medium containing 0.1 mg/mL of colcemid (Sigma-Aldrich) at 37°C for an hour, trypsinized into single cells, resuspended, and incubated in 68 mM KCl for 20 min at room temperature, and then fixed with fixative (3:1 methanol:glacial acetic acid), and dropped to make the spread of chromosomes on the slides. The slides were dried at 37°C on slide warmer overnight, baked at 80°C for 90 min, treated with 0.05% trypsin (Life Technologies) for 45 s to 1 min, and then stained with the Giemsa and Leishman's solution (Sigma-Aldrich). At least 10 metaphase spreads were analyzed for each of hiPSC lines grown in stirred bioreactors.
Statistical analysis
Statistical analysis on data was carried out using unpaired Student's t-test for the comparison of two groups with a minimal significance of P < 0.05.
Results
Effect of inoculum format on hiPSC expansion in stirred-suspension bioreactor culture
Cell aggregate formation and size distribution are critical for suspension bioreactor expansion. In this report, we compared the two methods of bioreactor inoculation: single cells and aggregates to develop a highly efficient and reproducible method for expanding iPSCs in suspension culture (see flow chart; Fig. 2). Different inoculum forms, including single cells, cell clumps, and cell aggregates, have been reported for the expansion of mammalian stem cells in suspension bioreactors [14,25].
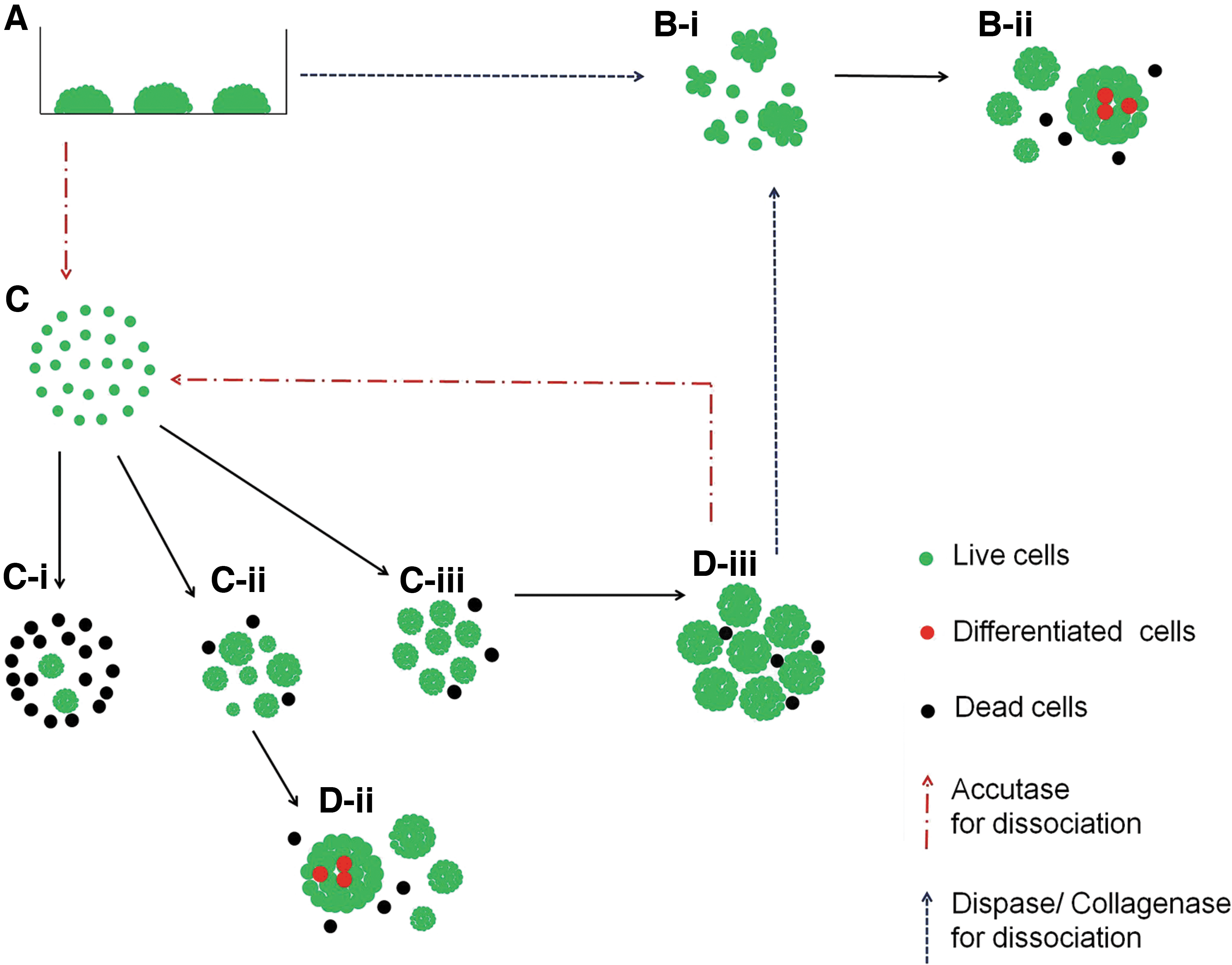
Summary diagram outlining different cell dissociation and aggregation methods used for hiPSC bioreactor expansion. hiPSCs in static culture
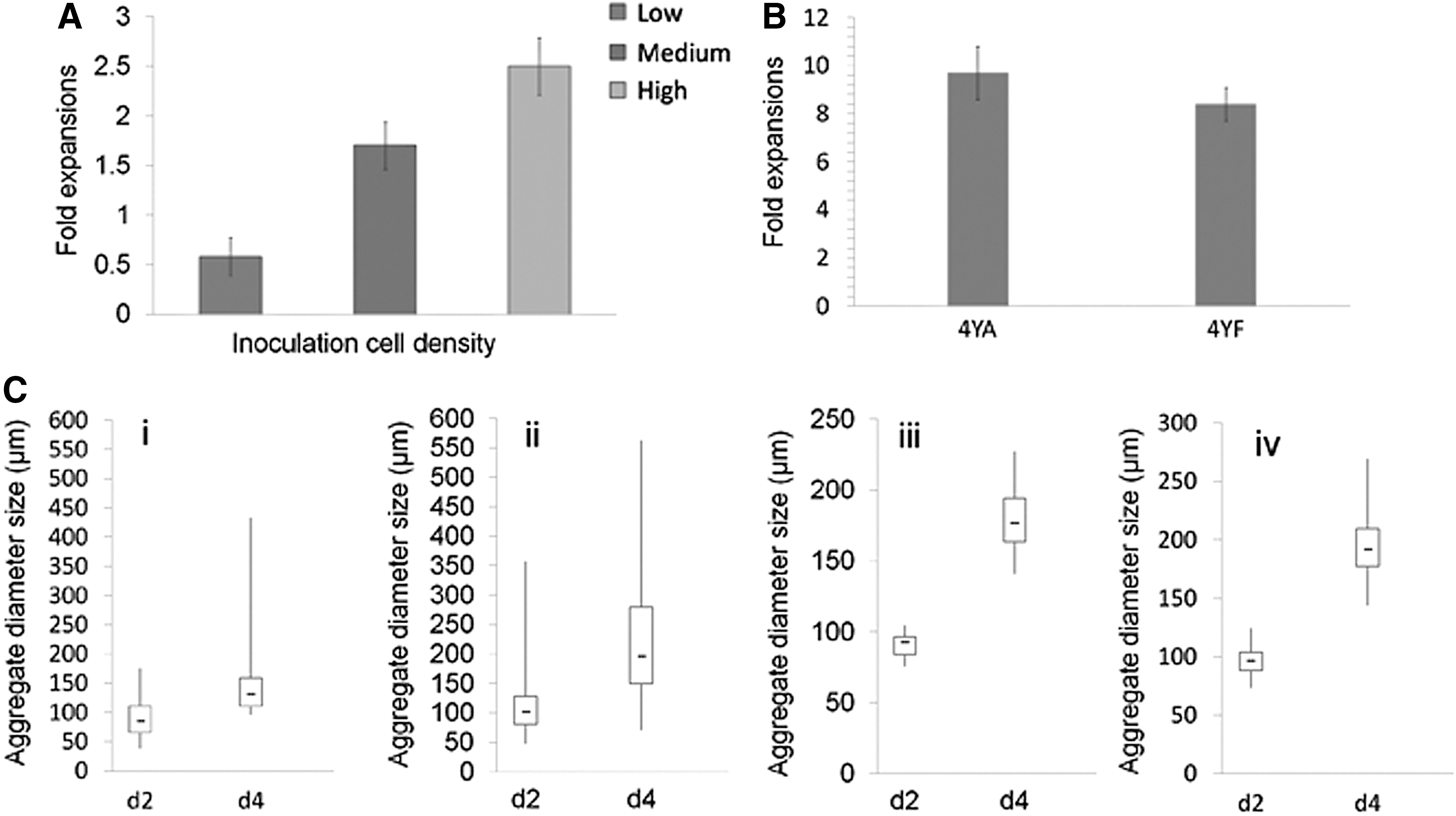
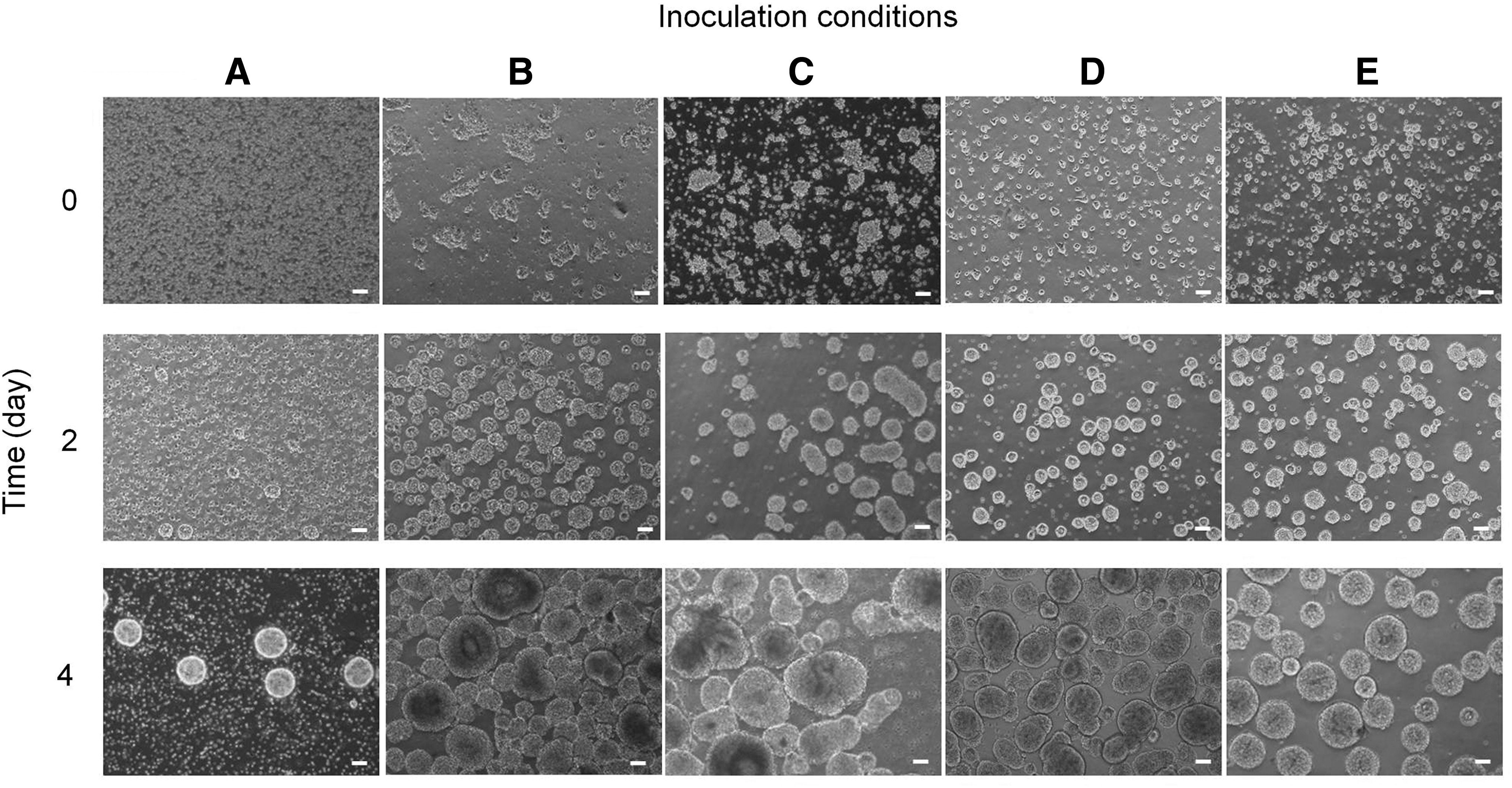
We first examined the outcome of single cell-inoculated bioreactor culture of hiPSC at low (1 × 105 cells/mL), medium (2.5 × 105 cells/mL), and high (5 × 105 cells/mL) densities. All cells were exposed to the same agitation rate of 100 rpm and cultured in the same type of bioreactor with 100 mL of mTesR1 supplemented with 10 μM Y-27632. At day 5 of stirred-suspension culture, fold expansion estimated at 0.58, 1.7, and 2.5 was detected for low, medium, and high seeding densities, respectively (Figs. 3A and 4A). These values were not higher than the fold expansion observed in the static culture (Fig. 3B). We next examined the outcome of cell clumps that were generated from the enzymatic dissociation of hiPSC colonies followed by inoculation in bioreactors. Enzymatic dissociation with either Dispase or Collagenase IV generated clumps at various sizes and, these in turn, resulted in various cell aggregate sizes in the stirred-suspension culture. Dark necrotic centers were observed in some of the larger aggregates (≥300 μm) due to limited access of nutrient, growth factors, and O2 at the center. For more uniform aggregate size distribution in the stirred-suspension culture, we explored methods to generate small aggregates (∼10 cells) as inocula for stirred-suspension bioreactor culture.
Cell expansion following different inoculation formats.
Inoculation conditions for the expansion of hiPSCs in suspension bioreactors.
We generated small hiPSC aggregates from dissociated single cells using three methods. One method was the formation of cell aggregates in conical centrifuge tubes for 3–4 h under standard culture conditions. Approximately 75% of the seeded single cells participated in aggregate formation, but the cell aggregates resulted in various sizes. Another approach involved transferring single cells onto PDL-coated dishes and incubating overnight in standard culture conditions. Approximately 70% of the seeded single cells participated in aggregate formation.
The resulting aggregates from this method were more uniform in size compared with the first method. Our third method, which involved culturing dissociated single cells in uncoated Petri dishes, gave similar yield in aggregate number and size as the PDL approach. Both the second and third methods were appropriate to generate small aggregates to inoculate the stirred bioreactor. Compared with the third method, one drawback with the second method was the long period (18–24 h) needed to form aggregates, whereas cell aggregates preformed through static suspension culture only needed ∼4 h with homogeneous size. So, in this study, we adopted the third method for generating small cell aggregates to inoculate the spinner bioreactor (Figs. 2C and 4D, E).
Effect of splitting method on cell survival and reaggregation
We explored different methods of hiPSC dissociation for optimal hiPSC culture in stirred-suspension bioreactor. Aggregates could be split as either single cells or cell clumps. We dissociated aggregates into cell clumps using both enzymatic and mechanical methods. Both methods generated nonhomogeneous cell clumps that later gave rise to cell aggregates of variable sizes (Figs. 3C-i, ii and 4B, C). In contrast, dissociation of aggregates into single cells followed by reaggregation resulted in more uniform cell aggregates in the bioreactor (Figs. 3C-iii, iv and 4D, E). We also compared the cell recovery rate of single cells generated using two enzymatic dissociation reagents, Accutase and TrypLE. We detected that ∼1.4-fold higher number of Accutase-dissociated cells participated in reaggregation compared with dissociated cells using TrypLE (Supplementary Fig. S1; Supplementary Data are available online at
Effect of inoculum level and agitation rate for stirred-suspension bioreactor culture
We determined the effect of inoculum level by seeding cells at different densities: low (∼2 × 104 cells/mL), medium (∼4 × 104 cells/mL), and high (∼8 × 104 cells/mL). We further examined the effect of agitation rate (ie, 80, 100, and 120 rpm) on cell aggregation in the bioreactor containing 100 mL of mTeSR1 medium supplemented with 10 μM Y-27632. At low agitation rate (80 rpm), cells inoculated at all three densities failed to form homogeneous aggregates (Fig. 5A-i). Similarly, cells inoculated at high agitation rate (120 rpm) also failed to form homogeneous aggregates with aggregates disintegrating probably due to the high stirring speed/shear force in the bioreactor (Fig. 5A-iii). Cells inoculated at medium agitation rate (100 rpm) showed the best growth kinetics and aggregation size compared with the other two agitation rates (Fig. 5A-ii). Figure 5B shows the growth curves with viable cell densities for each condition grouped by agitation rates, and Fig. 5C shows the respective fold expansions. The results indicated that the medium agitation rate (100 rpm) is optimal for hiPSC expansion in the bioreactor. However, if we extended expansion period up to day 6, a small portion of oversized aggregates would undergo apoptosis or differentiation in the central areas (Fig. 6A, day 6) for both cell lines. Based on this reason, we used 5 days as the expansion period in this protocol.
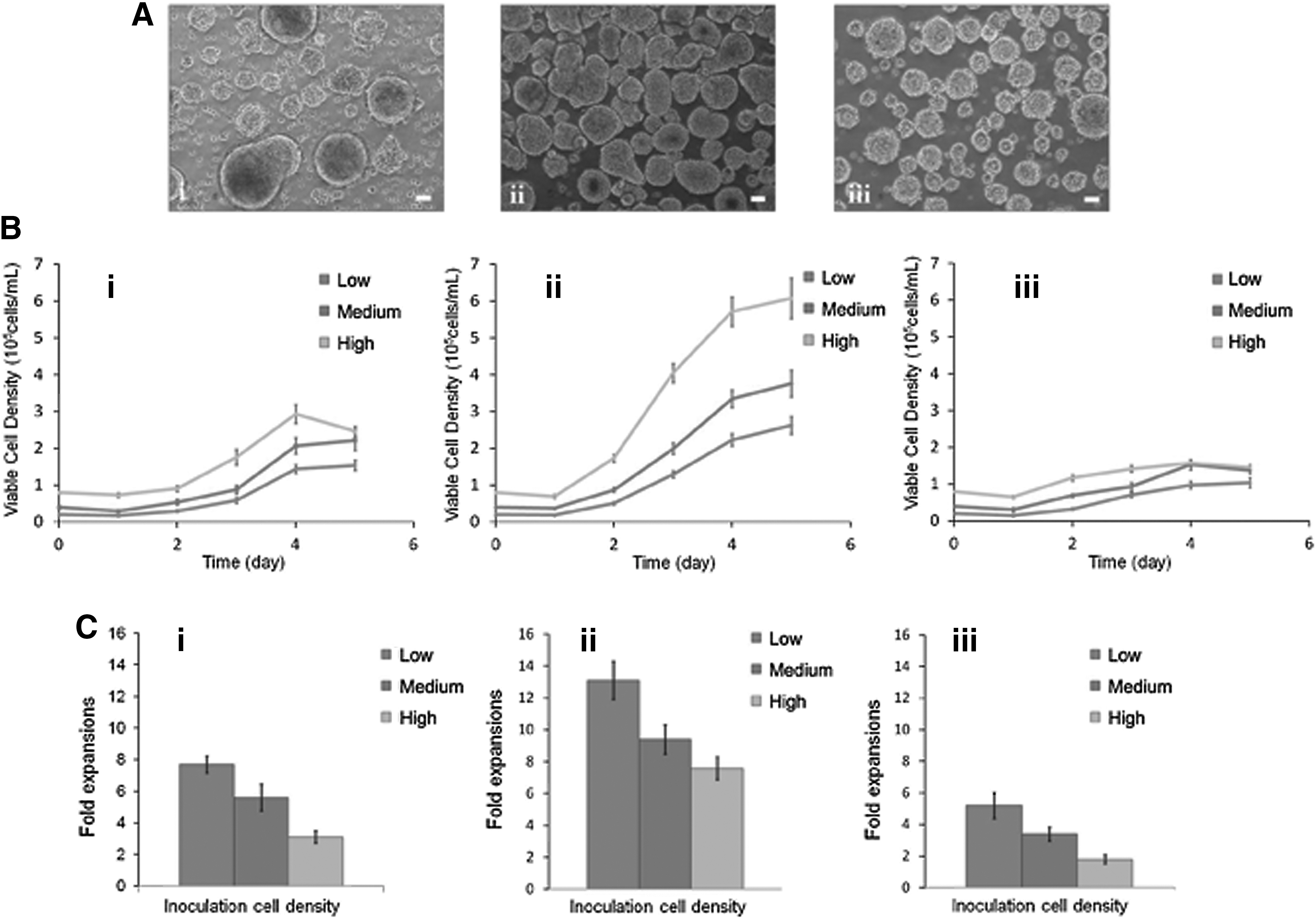
Effects of agitation rates and inoculation densities on hiPSC bioreactor expansion following the inoculation of cell aggregates.
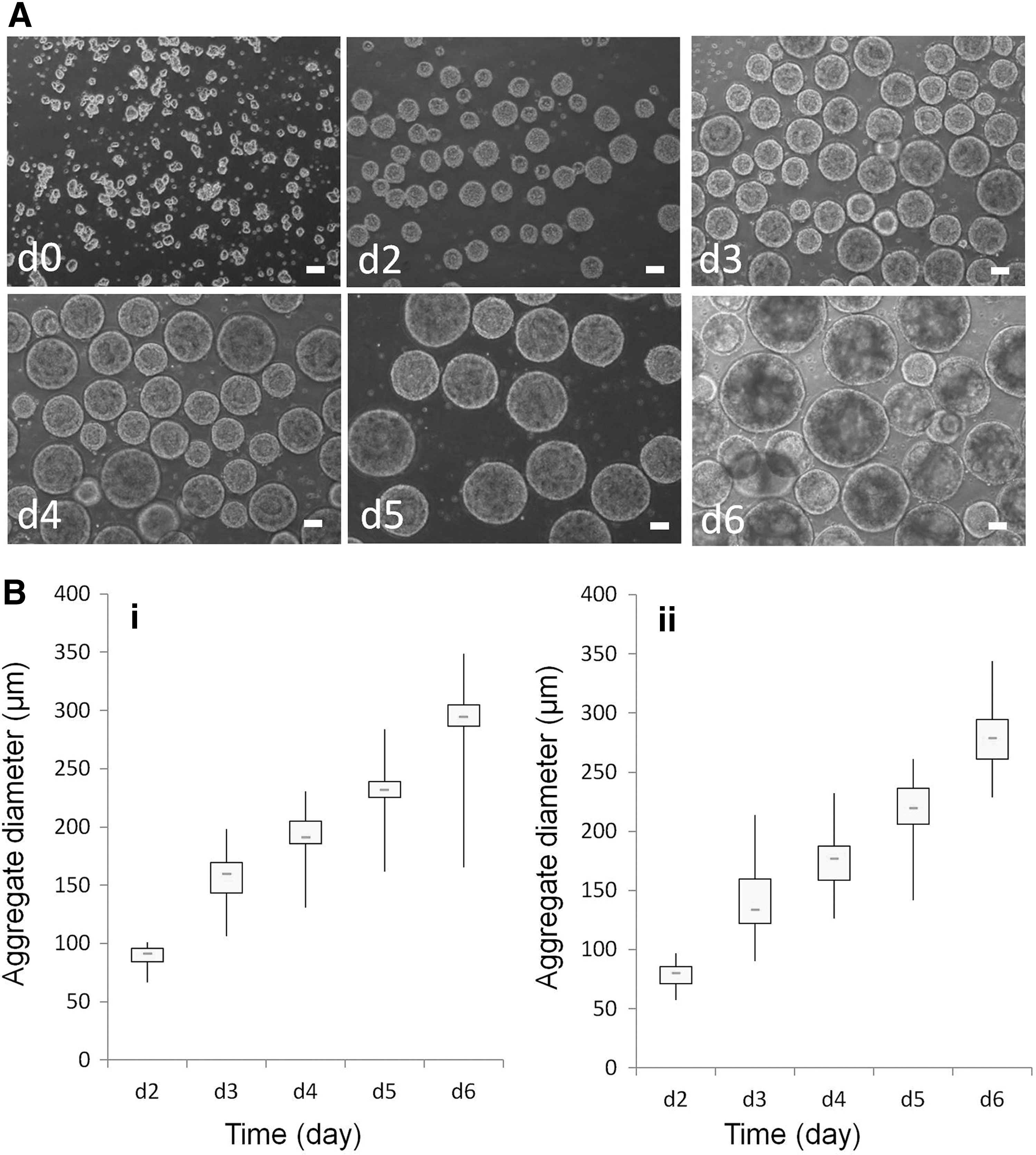
Average aggregate size and the expansion period in the bioreactors inoculated with cell aggregates in medium density at 100 rpm.
At 100 rpm, we observed increased viable cell density with increased inoculum level for hiPSC lines, 4YA and 4YF (Fig. 5B-ii and Supplementary Fig. S2A). The fold expansion, however, decreased with the increased inoculum levels (Fig. 5C-ii and Supplementary Fig. S2B).
Effect of aggregate size and expansion period
The average size of cell aggregates is one of the crucial factors that affect the pluripotency of hiPSCs grown in bioreactors. If the size is too large, then cells in the center of the aggregate tend to undergo spontaneous differentiation and/or cell death. To determine the optimal aggregate size, we measured the diameter of the cell aggregates in stirred-suspension bioreactor culture on a daily basis. Figure 6A shows the single cell-derived aggregates as inocula at day 0 (Fig. 6A, d0) and aggregates grown in bioreactor from day 2 to 6 (Fig. 6A, d2–d6). Our results suggested that the optimal size for cell harvesting or splitting is ∼250 μm, which was obtained at day 5 of cell culture. Aggregates larger than 250 μm tend to form dark centers that are indicative of spontaneous differentiation and/or cell death (Fig. 6A, d6). These findings were consistently observed in both hiPSC lines 4YA (Fig. 6B-i) and 4YF (Fig. 6B-ii).
Effect of pH
The pH of the medium changes based on cell density in the bioreactor and it should be closely monitored as low pH values have negative effects on stem cell proliferation [25]. We observed that the hiPSCs grew well as aggregates (Fig. 7A-i) in a weak alkaline medium (7.0<pH <7.5). The pH value dropped as the cells expanded in the bioreactor. When the pH value was 6.9, the rate of cell expansion and cell survival significantly decreased (Fig. 7A-ii). The number of days in culture for the medium to reach this slightly acidic pH value was about 4, 3, and 2 days for experimental groups with low, medium, and high inoculum levels, respectively. At these time points, we changed the medium for different experimental groups. If the medium remained unchanged, then the pH value dropped even further, which resulted in significant decrease in aggregate size (Fig. 7B-i) and viable cell percentage (Fig. 7B-ii).
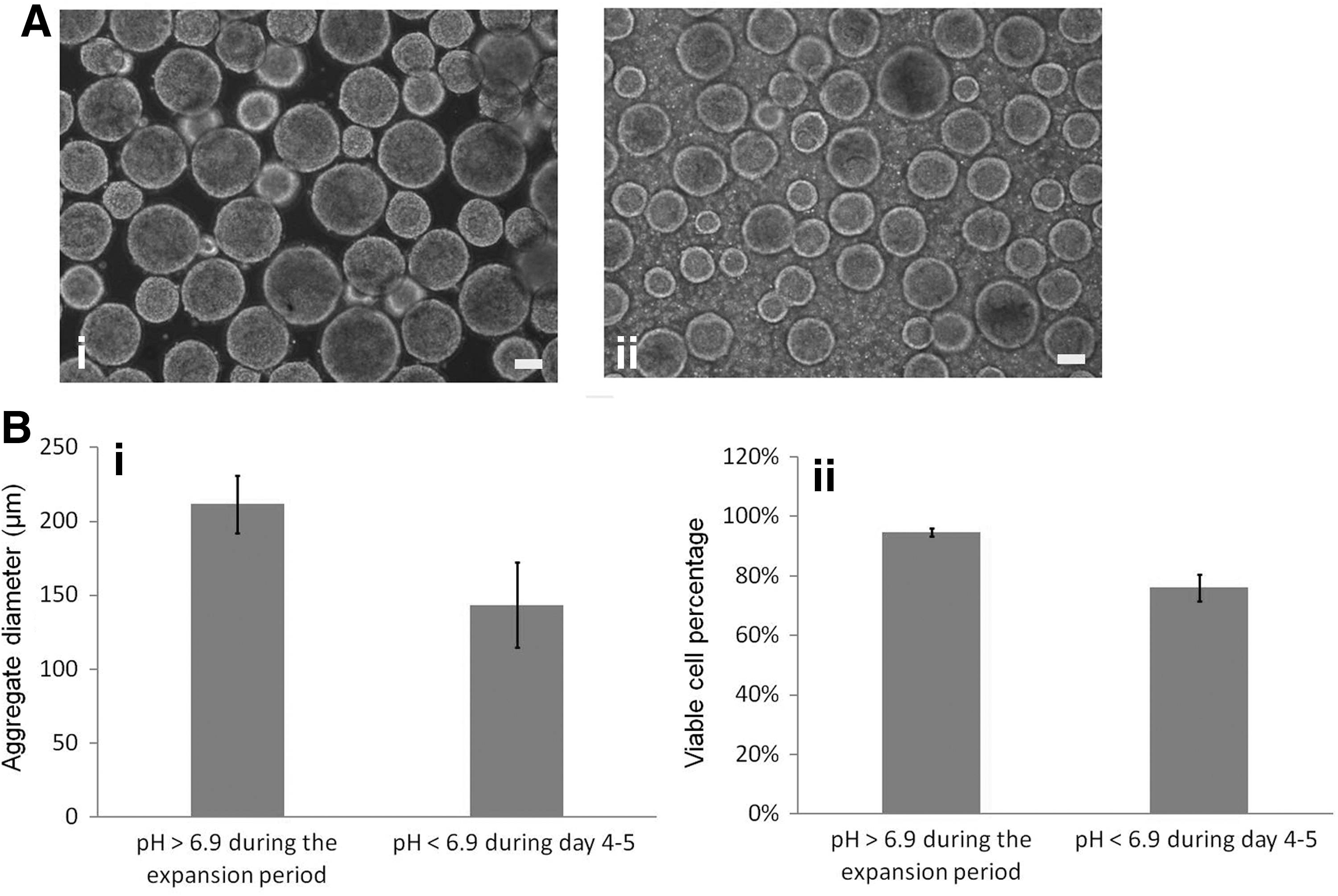
Effect of pH on hiPSC suspension culture.
Effect of Y-27632 and growth factors
ROCK inhibitor (Y-27632) is commonly added to stirred-suspension bioreactor culture to promote cell survival [12,14,26]. In this study we determined the effect of Y-27632 on the number of viable cells at three different concentrations, 5, 10, and 15 μM. We observed lower number of viable cells and fold expansion in the presence of 5 μM Y-27632, whereas, at 10 and 15 μM Y-27632, similar cell yields were measured (Fig. 8A). However, we found that cell aggregates obtained from the 15 μM condition were tightly packed and were more difficult to dissociate into single cells compared with aggregates from the 10 μM condition.
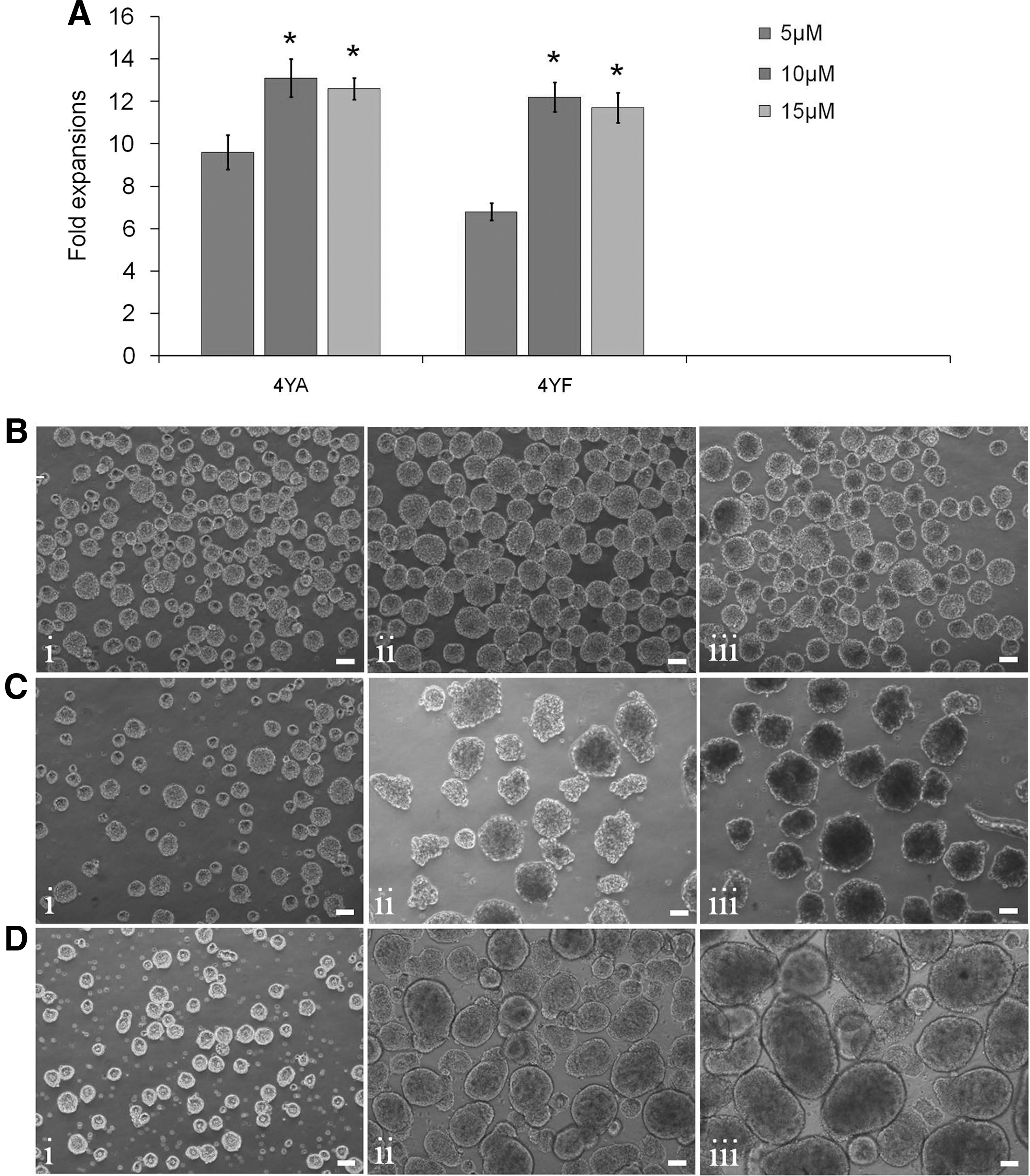
Effect of Y-27632 and growth factors on hiPSC suspension culture.
We also explored whether growth factors bFGF and TGF-β are necessary for hiPSC expansion in stirred-suspension bioreactor culture. In the absence of both growth factors in the mTeSR1 medium, hiPSCs stopped growing starting at day 3 in culture (Fig. 8B-i, iii). In the absence of bFGF, decreased cell growth was observed (Fig. 8C-i, iii) compared with control that had both bFGF and TGF-β (Fig. 8D-i, iii). These results suggest that optimal hiPSC expansion requires both bFGF and TGF-β.
Optimal culture conditions for hiPSC expansion in stirred-suspension bioreactor
Based on aforementioned comparative experiments, we defined the optimal conditions for hiPSC suspension culture in 125-mL stirred bioreactors containing 100 mL culture medium supplemented with 10 μM Y-2762. These include: (1) Accutase for the generation of single cells, (2) small, single cell-derived aggregates as inoculum, (3) optimum inoculum level of 2–4 × 104 cells/mL, (4) optimum agitation rate of 100 rpm, (5) replacement of medium when pH dropped to 6.9, (6) optimum cell aggregation size of 250 μm, which could be achieved at day 5 of culture. We continued to culture two different hiPSC lines, 4YA and 4YF, under these culture conditions for 10 and 7 passages, respectively. The hiPSCs at the end of these passages were collected and assessed for pluripotency.
Expanded hiPSCs retain their pluripotency and differentiation capacity
Karyotype analyses indicated that hiPSCs obtained from our defined culture conditions had normal karyotypes (Fig. 9A and Supplementary Fig. S3A). The cells also expressed pluripotency markers, including alkaline phosphatase (AP), Oct4, Nanog, TRA-1-60, TRA-1-81, and SSEA-4 (Fig. 9B-i, vi and Supplementary Fig. S3B-i–vi). Flow cytometry analysis indicated that the hiPSCs expressed high levels of Oct4, Nanog, Sox2, and SSEA-4 in both cell lines (Fig. 9C-i, iv and Supplementary Fig. S3C-i–iv).
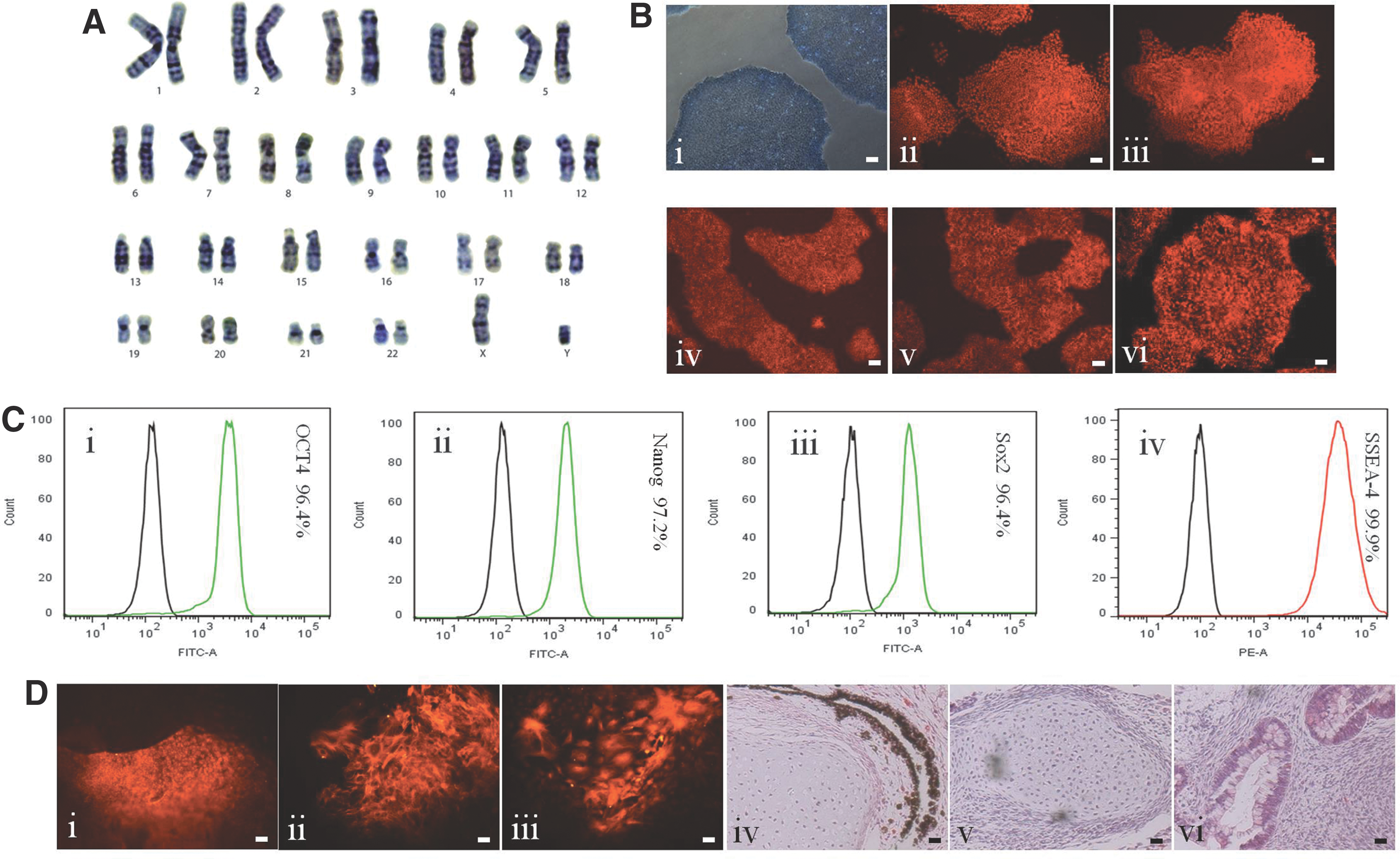
Characterization of hiPSCs grown in the stirred bioreactors.
The differentiation capability of the hiPSCs from stirred-suspension bioreactor culture was also assessed. EBs generated from hiPSCS were able to spontaneously differentiate into the three germ layers as differentiated cells were stained positive for AFP (endoderm), smooth muscle actin (mesoderm), and β-III tubulin (ectoderm) (Fig. 9D-i, iii and Supplementary Fig. S3D-i–iii). We also used the teratoma assay as a complementary in vivo test. Teratoma produced from 4YA (Fig. 9D-iv–vi) contained pigment cells (ectoderm), cartilage (mesoderm), and columnar glands (endoderm). Teratoma produced from 4YF (Supplementary Fig. S3D-iv–vi) contained rosettes of neural epithelium (ectoderm), cartilage (mesoderm), and columnar glands (endoderm).
Discussion
Human pluripotent stem cells (hPSCs, including hESCs and hiPSCs) share the key properties of unlimited self-renewal and pluripotency to differentiate into any cell types in the body. hiPSCs have been considered as an effective replacement for hESCs and a promising candidate cell source for regenerative medicine and cell-based therapy. Additionally, hiPSCs also have their own advantages. First, they avoid the ethical issues surrounding the use of human embryo-derived hESCs. Second, they can be autologously derived from patient tissue thus eliminating immune rejection upon transplantation. However, critics have warned that regenerative medicine may be too expensive to implement, particularly from an iPSC personalized medicine perspective [27,28]. Thus, governments have begun to invest in the development of hiPSC biobanks in anticipation of clinical trials [29]. To make hiPSC technology more cost effective, the same cell lines (from biobanks) will need to be used for multiple applications, making good cell manufacturing extremely important [30].
Currently, in the initial, investigator-led clinical trials being developed for hiPSCs, cells are cultured in dishes under static conditions [31], which are laborious, and results in low cell yields, with limited quality control. In contrast, bioreactors, which are central to good cell manufacturing, offer advantages over culture dishes [30]: key physiological parameters can be controlled and standardized, allowing for batches with large cell numbers.
In our experiments, we attempted to develop methods that improve the expansion of hiPSCs in stirred-suspension bioreactors. mTeSR1 is commercially available and the most commonly used medium for hESC and hiPSC culture and expansion in laboratories worldwide. An optimal protocol developed for suspension culture in this medium has universal significance. Compared with previous reports, many more parameters were studied and lots of comparative experiments were performed to define the optimal culture conditions for this system. Our results indicate that several factors influence the quality and quantity of hiPSCs produced in bioreactors. These included: (1) colony splitting, (2) cell aggregate formation, (3) inoculation methods (single cells, cell clumps, or aggregates from single cells), (4) cell inoculation density, (5) agitation rate, (6) pH, (7) average size of cell aggregates for inoculation and harvest, and (8) expansion period.
We recognize that the inoculation condition has a significant effect on cell survival and growth characteristics during the initial stages of suspension culture in bioreactors. So, we explored three types of hiPSC inoculum: (1) single cells, (2) cell clumps, and (3) sphere-like hiPSC aggregates, and excluded type (1) and type (2) through comparative experiments. Type (3) contains three different methods for hiPSC aggregate formation. The results verified that method 1 (aggregates formed on PDL-coated dishes) and method 2 (aggregates formed in static suspension culture condition) are ideal methods to generate uniform aggregates in size.
Differing from mouse, single cells dissociated from hiPSC colonies or aggregates have a poor ability to form aggregates in stirred bioreactors even at high cell density (∼5 × 105 cells/mL) in the presence of Y-27632. Using Dispase, Collagenase IV and/or mechanical dissociation, cell colonies (grown in static) or aggregates (grown in suspension) were dissociated into cell clumps or clusters for inoculation of bioreactors. However, as these cell clumps or clusters were produced in a large range of sizes, the resulting cell aggregates grown in bioreactor also have a large difference in size. In the central portions of large aggregates, cell necrosis and/or differentiation could be observed under the dissecting microscope.
We observed that the inoculation of cell clumps or clusters into bioreactors was not a good method for the suspension culture, so we tried to preform small aggregates with uniform size from single cells for inoculation of bioreactors. We used three methods to make cell aggregates. When cell aggregates were preformed in a conical tube, we observed a huge range of sizes, even when using a small number of cells. In contrast, cell aggregates preformed on PDL substrate were robust, similar in size, and suitable for suspension culture. However, one drawback was the long period (18 ∼ 24 h) needed to form aggregates under this condition. When cell aggregates were preformed through static suspension culture, this method only needed ∼4 h to form small aggregates with homogeneous size, which are appropriate for suspension culture in stirred bioreactors. Aggregates could also be formed in low attachment dishes (plates) and agar-coated dishes for around 4 h. At the same cell density, there is no obvious difference in rates of aggregate formation between them.
If cells (in the form of aggregates) were inoculated into the bioreactor at high density, aggregates reached a plateau in cell number in a short time (3–4 days). However, at this time point, such cell aggregates failed to reach the appropriate size for passaging. This can lead to some bad consequences, which are harmful to cells, such as a very rapid change in pH and fast nutrient depletion. The frequent medium replacement and cell splitting required would be a costly waste of media and growth factors.
pH monitoring is a key step in the process of suspension culture, as cell aggregates grew well in a weak alkaline medium, but induced apoptosis and gradually died in a weak acid environment (below 6.9). This suggests that hiPSCs are intolerant to the weak acid environment.
Based on our experimental design, the pH value would drop to 6.9 or below, in about 3 days after inoculation of hiPSC aggregates to the bioreactor at the density of 2–4 × 104 cells/mL. At this point, the cells need to be refed with fresh medium for 2 more days' expansion. About 5 days after inoculation, the cell number and average size of aggregates are ready for subculture. Certainly, these parameters should be altered according to the rate of cell proliferation. If cell proliferation is obviously faster or slower than the two cell lines used in this study, the parameters, such as cell inoculation density, medium replacement, and expansion period should be changed to the appropriate state.
According to our results, culture medium can maintain the undifferentiated state of hESCs (data not shown) and hiPSCs in stirred bioreactors before the pH drops to 6.9. We conclude that bFGF in the culture medium is relatively stable and can maintain hPSCs in an undifferentiated state for 3–4 days without changing medium when cells are seeded at low or medium density. This is different from static culture of hPSCs, wherein culture medium needs to be changed every day or at most every other day. We speculate that the shear forces produced by stirred bioreactor probably have the synergistic effect with bFGF on the pluripotency of hPSCs.
In collaboration with STEMCELL Technologies, we tested if growth factors, bFGF and TGF-β, are necessary for hiPSC expansion in a stirred-suspension bioreactor culture. We have previously observed that fluid shear promotes the pluripotency of mouse PSCs, allowing for the removal of LIF from bioreactor culture [32,33]. However, for human iPSCs, we observed that cells would stop proliferating in the absence of bFGF and/or TGF-β.
The average size of cell aggregates is a crucial factor that affects the pluripotency of hiPSCs in this suspension culture system. During expansion, the average size of aggregates increased more than five times from a starting size (25–50 μm) to the ending size, (∼250 μm), whereas the cell number increased 12- to 13-fold. We can also control the initial aggregate size in 25–50 μm by seeding cells with 3–5 × 104 cells/cm2 combined with trituration. During each expansion period, the size of the aggregates should be monitored and harvested in time to prevent them from getting too large. In the central portions of the oversized aggregates, cells would undergo apoptosis or differentiation, due to the lack of O2, nutrient, and cytokines in these areas.
The average diameter of preformed aggregates used for inoculation of bioreactors should be <50 μL. According to the aggregate morphology and measurement results, we concluded that the appropriate average size of aggregates is around 250 μm. When the aggregate size is close to 300 μm in diameter, they would be getting worse due to abovementioned reasons.
We tested three different agitation rates, 80, 100, and 120 rpm, at varying inoculation densities (2, 4, 8 × 104 cells/mL). At 120 rpm cells failed to thrive, as fluid shear produced in these conditions destroyed aggregates. In contrast, at 80 rpm aggregates grew to sizes approaching 400 μm and possessed large necrotic/differentiated centers (see Results section). We obtained the best uniformity of the aggregates, cell number, and viability with the medium agitation rate (100 rpm).
ROCK inhibitor, Y-27632, plays a crucial role in this study. Similar to hESCs, hiPSCs also undergo massive cell death in culture after single cell dissociation. Y-27632 helped to resolve this problem. Previous studies have demonstrated that Y-27632 is a safe reagent, which can greatly improve the cell survival rate after single cell dissociation [26]. As of now, the evidences suggested that Y-27632 has no negative effect on both hESCs and hiPSCs, but instead increases the cell–cell and cell–substrate interactions. As single cells are apt to undergo apoptosis and die in a short time, they must be maintained in the presence of Y-27632. So, the cell dissociation solution, media for freezing, thawing, static suspension, and stirred-suspension culture are all supplemented with 10 μM Y-27632. In the process of suspension culture in stirred bioreactors, even if the small aggregates were preformed in static suspension, Y-27632 also needs to be added to the culture medium after they were inoculated to the bioreactors. Without its addition, aggregates disperse into irregular cell pieces in suspension conditions.
We designed and performed lots of comparative experiments to define the influencing factors on hiPSC scale-up in stirred bioreactor. During each expansion period, the cell number increased 12- to 13-fold in 5 days. Theoretically, a large quantity of cells can be obtained for clinical purposes by using a large number of stirred bioreactors. However, it is tedious, labor intensive, and there is a risk of contamination. To address this problem, we adopted a two-step upscaling approach to expand hPSCs in different size vessels. We made small aggregates in a 60-mm ultra-low attachment dish under static conditions, perform suspension culture in a 100-mL stirred bioreactor, collect cell aggregates after an expansion period, and made smaller aggregates through trituration in a 50-mL Erlenmeyer flask or a 100-mL stirred bioreactor, which has enough small aggregates to serve as the initial inoculum for further scale-up culture in 1,000 mL stirred spinners.
To conclude, several factors stand out in this study. First, the initial static cultured cells used for inoculation of bioreactors should be in a highly undifferentiated state to prevent the propagation of differentiated cells in the bioreactor. We have previously reported a surgical method for efficiently removing differentiated cells from hPSC colonies [34]. Even if differentiation is <5%, differentiated regions of colonies should be removed completely before transferring, as it is impossible to remove differentiated cell bioreactor culture. Second, the average size of initial aggregates should be between 25 and 50 μM. Large initial aggregates shorten the expansion period in suspension and frequent splitting is harmful to cells. Third, the pH value and the nutritional status of culture medium is a key factor for suspension culture of hiPSCs, as cells can undergo apoptosis or differentiation in a weak acidic environment. Although we do not need to change the medium every day, as we do in static culture, we should feed cells in time with fresh medium according to the pH value of the culture medium. Finally, cells in the center of overgrown aggregates undergo necrosis/differentiation due to problems with mass transfer of nutrients and/or O2.
At an optimal agitation rate, we observed the increase in viable cell density and the decrease in expansion fold with increased inoculum level for both hiPSC lines. For example, when seeding 4YA hiPSCs at different densities, low (∼2 × 104 cells/mL), medium (∼4 × 104 cells/mL), and high (∼8 × 104 cells/mL), the viable cell density was 2.62 × 105, 3.76 × 105, and 6.08 × 105 cells/mL, respectively, the fold expansion was 13.1, 9.4, and 7.6, respectively. The maximum cell number obtained in a 100-mL stirred bioreactor was about 6 × 107, which can be used to preform small aggregates for starting inoculum to large-scale bioreactors (eg, containing 1,000 mL medium). Two or more 1,000-mL bioreactors would generate 109 hiPSCs, which would be enough for clinical trials.
Footnotes
Author Disclosure Statement
The authors declare no conflict of interest. No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.