
Abstract
Mesenchymal stem cells (MSCs) are the primary cell source in cell therapy and regenerative medicine due to its extraordinary self-renewing capacity and multilineage differentiation potential. Clinical trials involving MSCs are being conducted in a range of human diseases and the number of registered cases is continuously increasing. However, a wide gap exists between the number of MSCs obtainable from the donor site and the number required for implantation to damage tissues, and also between MSC scalability and MSC phenotype stability. The clinical translation of MSCs necessitates a scalable expansion bioprocess for the biomanufacturing of therapeutically qualified cells. This review presents current achievements for expansion of MSCs. Issues involving culture condition modification, bioreactor systems, as well as microcarrier and scaffold platforms for optimal MSC systems are discussed. Most importantly, the gap between current MSC expansion and clinical application, as well as outbreak directions for the future are discussed. The present systemic review will bring new insights into future large-scale MSC expansion and clinical application.
Introduction
R
A considerable amount of clinical studies has shown very promising results in terms of both therapeutic efficacy and safety. Currently, there are more than 500 registered clinical trials involving MSC-based therapy worldwide, with 159 studies completed for potential treatments of type 1 diabetes, knee osteoarthritis, multiple sclerosis, acute graft-versus-host disease (aGVHD), Crohn's disease, and myocardial infarction [5 –10]. In the past decade, stem cell industries have released multiple MSC-based products into the market, such as Osteocel®, OvationOS®, and BIO4®, which are viable bone matrix products designed to stimulate bone repair and regenerative, developed by NuVasive, Inc., Osiris Therapeutics, Inc., and Stryker, respectively. Osiris's other current product line in the market also includes Grafix® for acute and chronic wounds, and Cartiform® for cartilage repair. Recently in 2016, Mesoblast launched TEMCELL® HS Inj through JCR Pharmaceuticals Co. Ltd., the first allogeneic cell-based product to receive full approval in Japan for children and adults with aGVHD. In the near future, inevitably several more cell-based products will reach the market, namely MSC-based products.
Although MSCs show effectiveness for various therapeutic applications with multiple launched products, clinical translation of these therapies are hindered by challenges in scalable expansion technology for the biomanufacturing of MSCs. Even though presently there are still no standardized therapeutic cell doses to be administered into patients, previous and current MSC-based clinical trials have shown that generally, several million cells per kilogram of body weight are required per treatment (Table 1) [11,12]. However, only a very small quantity of MSC can be isolated from the previously mentioned sources, for instance, MSC only represents ∼0.01% of the total mononuclear cell fraction in the bone marrow [13,14]. This reason has led to the emergence of multiple research groups focusing their attention solely on improving and developing new culture methods toward clinical-scale expansion of MSC. Nevertheless, most studies focus on questions starting with how and what, for instance, research results have shown how to optimize MSC expansion methods, and what to add or alter to increase MSC proliferation. However, the questions that were seldom asked is why and which, why are we using this procedure or technology to expand MSCs, and by using this particular method, which therapeutic application is most suitable? Most groups aim at finding a culture condition or method not affecting differentiation and promote cell expansion while disregarding the effect of particular culture condition on differentiation preference toward a specific lineage. Therefore, it is important to address the missing crucial linkage between regenerative medicine bioprocessors and translation into actual clinical applications. In this review, we give an overview of the different MSC expansion methods, including the utilization of chemical and biological additives, bioengineered scaffolds, and microcarriers, together with the dynamic culture system using bioreactors.
Two-Dimensional Conventional Culture Condition for Expansion of MSCs
Growth factor additives for MSC expansion
MSCs for clinical use are typically expanded using static adherent cultures, predominantly combining with fetal bovine serum (FBS) [15]. However, to avoid risk related to the use of animal products, the use of human-derived products, such as human serum, platelet-rich plasma, and platelet lysate (hPL), has been suggested, with hPL proving to be the most effective cell culture additives [16 –18].
Within recent years, many laboratories adapted their culture conditions to hPL. Between 2007 and 2013, more than 10% of clinical trial cases used hPL for their studies, as they can simply be obtained from apheresis products or buffy coats of healthy volunteers [16 –20]. Bernardo et al. illustrated that the release of growth factors from the platelets, including PDGFs, b-FGF, VEGF, IGF-1, and TGFβ stimulated proliferation and expansion of MSCs [20]. However, Trubiani et al. proved that culture conditions with hPL have limitations as MSCs culture with hPL exhibited a reduction of osteogenic or adipogenic differentiation potential, as well as decreased immunosuppressive capability with modified surface marker expression [21]. Also similar to FBS, hPL is not precisely defined and also suffered from batch-to-batch variation, and therefore the capability to maintain growth and therapeutic potential of MSCs varies between different batches. Thus, it is essential to establish a well-defined culture system free of human and animal-derived products, using optimized xeno/serum-free media capable of stimulating efficient isolation and expansion of MSCs without altering their therapeutic functions, and at the same time in accordance with Good Manufacturing Practice (GMP). The first FDA-approved commercial xeno/serum-free product is StemPro® MSC SFM, launched by Invitrogen. Crapnell et al. showed an enhanced effect of StemPro MSC SFM to support highly efficient growth while maintaining the surface markers and characteristics of MSCs [22].
Growth factors can also be directly added to MSC cultures. The choice of growth factor additives is determined based on the present knowledge on the influence of the particular growth factor on cell behavior. Since individual growth factors influence differentiation at different rates and some have more impact on proliferation, synergistic pairs of growth factors should be applied to optimize the culture condition of MSCs. For instance, TGFβ is known to have a profound influence on cells from the chondrogenic lineage in vivo, and in vitro application of TGFβ on MSCs has shown increased cell proliferation and a bias toward the chondrogenic lineage [23]. Our laboratory constructed a sponge comprising of bone marrow-derived MSCs (BMSCs) and plasmid DNA encoding TGFβ1 to repair articular cartilage defects; results showed better chondrogenesis of BMSCs and complete repair of cartilage and subchondral bone when compared with constructs without gene insertion [24]. Moreover, Stewart et al. showed that BMP-2 through BMP-7 induced MSCs to form osteoblasts, whereas another member of the same family, BMP-3, stimulated MSC proliferation by threefold [25]. Similarly, Tokunaga et al. revealed that within the PDGFs family, PDGFRβ inhibited osteogenesis and induced MSC proliferation and migration, whereas PDGFRα was observed to facilitate osteogenesis [26]. Buckley and team also showed that MSCs expanded in the presence of FDFs proliferate more rapidly with higher cell yields while maintaining their differentiation potential; however, eventually, all of the MSCs differentiated into the chondrogenic line [27,28]. Other growth factors VEGF and IGF have also been tested and proved to play a role in promoting MSC proliferation [19,29].
Small-molecule additives for MSC expansion
As an alternative to growth factors, small-molecule drugs have been used in MSC-related studies to promote proliferation. Small molecule poses some advantages over growth factors, as they are not vulnerable to protease degradation or unfolding, and they can be chemically synthesized in the laboratory; therefore, reducing the production cost [30]. For example, MSC proliferation has been increased using a small-molecule Pam3Cys, a synthetic Toll-like receptor ligand, by 1.4-fold 48 h after treatment [31]. Similarly, Hoffman and Benoit observed a 1.8-fold increase in MSC proliferation in normal 2D culture conditions, as well as 1.3-fold increase when encapsulated within hydrogels with the small-molecule 6-bromoindirubin-3′-oxime (BIO), agonism of Wnt/β-catenin. BIO elevated several Wnt target-gene expressions such as Sox2, Oct4, NANOG, and cyclin D1 [32]. Moreover, Narcisi et al. showed that Wnt agonist small molecules, such as CHIR99021 (CHIR) and lithium chloride (LiCl), also stimulated MSC proliferation [33].
To provide better clinical results, the balance between MSC proliferation and ex vivo directed differentiation of MSCs to particular cell types for treating target diseases is required. Combinations of different small molecules may be able to synergistically regulate MSC proliferation and lineage-specific differentiation. Many studies have illustrated a number of small molecules that can be used to regulate the fate of MSCs (Table 2) [31,32,34 –39]. For example, CHIR and LiCl were also demonstrated to increase chondrogenic and osteogenic capacity, respectively [33]. Similarly, Park et al. demonstrated that the small-molecule Phenamil induced osteoblast differentiation and mineralization through the Trb3-dependent promotion of BMP action [38]. However, these studies also focused mainly on MSC differentiation; therefore, the study of small molecules should be directed toward its application for lineage-specific MSC expansion.
Large-Scale MSC Expansion with 3D Culture Systems
As mentioned earlier, the dosage of cells required to treat an adult patient in clinical applications significantly exceeds the number of cells accessible from donors. For a scalable expansion of MSC, dynamic culture systems have been utilized as an alternative to the classical planar culture systems, offering simpler operation protocols and harvesting procedures, a more flexible online monitoring and control, and a more cost-effective solution for large-scale production of MSCs for cell therapy [40].
For this purpose, various designs of bioreactors were developed for MSC amplification according to specific cell types and tissue structure (Fig. 1). Bioreactor microenvironments can be engineered to have a specific niche to accurately regulate stem cell fate decision; parameters that are widely explored include oxygen tension, biophysical cues (synthetic or natural scaffolds, physical forces; eg, shear stress), and biochemical cues (growth factors and bioactive molecules) (Fig. 1). For instance, an avascular and acellular tissue, such as cartilage, heavily relies on convection and molecular diffusion with loading-enhanced flow rates. On the other hand, the system for densely packed tissue, such as cardiac muscle, depends on efficient oxygen transport, whereas culturing bones and ligaments depend on mechanical inputs [41]. Due to these differences, developing a single standard bioreactor model for all tissue products still remains a challenge.
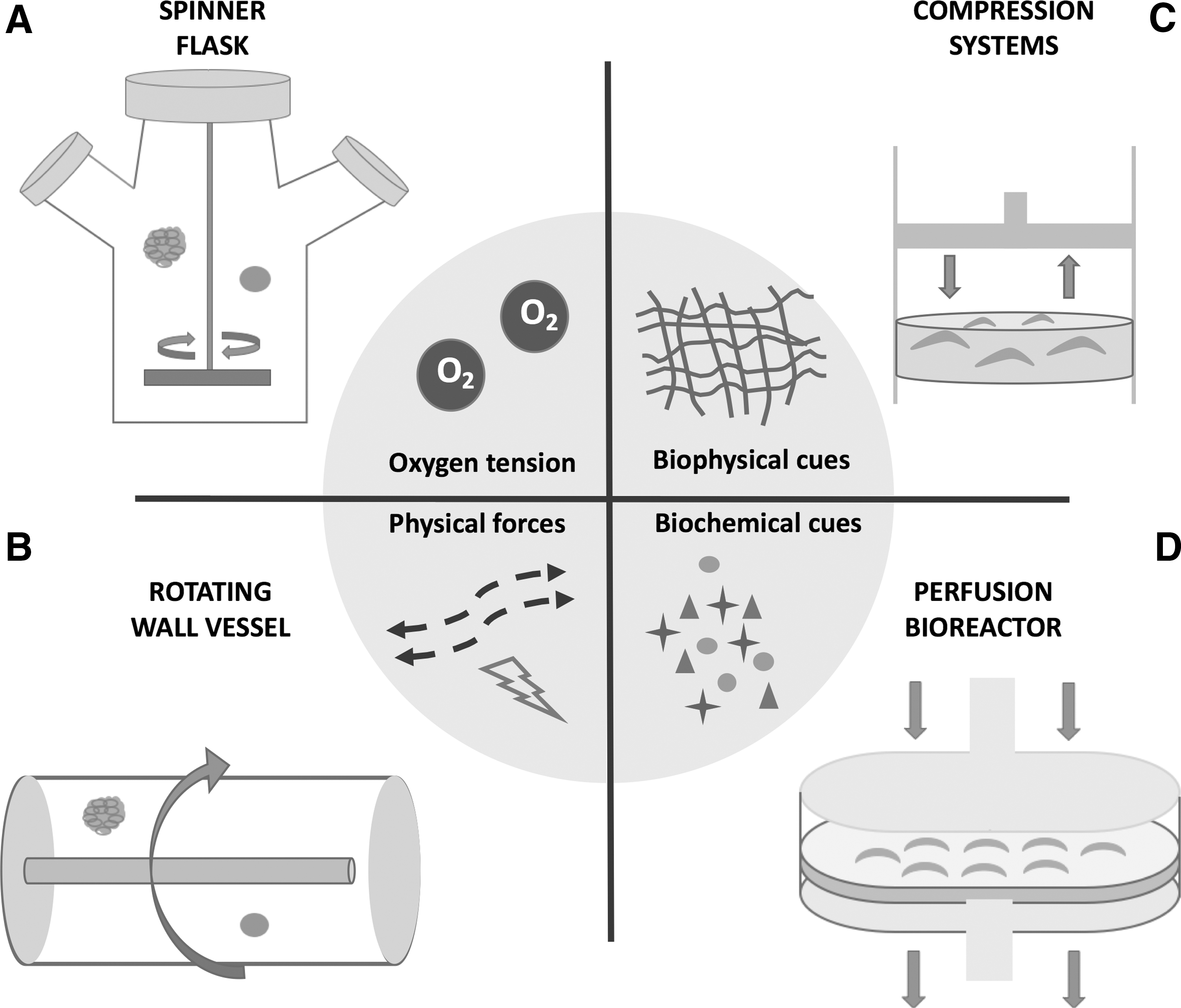
Different types of bioreactors to regulate MSC microenvironment, and crucial stem cell niche factors directing the fate of MSC proliferation and differentiation in bioreactors.
Hydrodynamic shear stress-dependent system
One of the most vital parameters for MSC expansion and differentiation in a bioreactor is the local internal shear stress generated by hydrodynamic forces. However, different flow rates have been applied in different studies based on trial and error, and there are not many studies that compare the variation of flow rates. The three major types of bioreactors that utilized this system are stirred suspension, rotating wall vessel (RWV), and perfusion bioreactors. The use of these bioreactors for MSC expansion and directed differentiation is listed in Table 3.
↑, increase; BM-hMSCs, bone marrow-derived hMSCs; AT-hMSCs, adipose tissue-derived hMSCs; ALP, alkaline phosphatase; Ca2+, calcium deposition; OC, osteocalcin; OP, osteopontin; PCL, polycaprolactone.
Stirred suspension bioreactor
Currently, stirred suspension bioreactor (SB), with the prototype of spinner flasks (SF), is mainly used for the large-scale production of MSCs with a stirring arm that mixes the media in the flask [42 –45]. In stirred SBs, cells can be cultured as single cells, as aggregates, or attached to a microcarrier or scaffold for anchorage-dependent cells such as MSCs. Kehoe et al. demonstrated that in conventional SBs, 106–107 cells/mL cell concentration can be obtained; other results demonstrated that long-term cultivation of MSC spheroid in SB also inhibits aggregate agglomeration, enhances cell viability, minimizes concentration gradients (metabolites, pH, dissolved oxygen), and improves osteogenesis and adipogenesis of MSCs [46 –48]. However, spatial shear stress distribution remains heterogeneous with high shear stress close to the stirrer and low shear stress nearer to the top, which may alter cell properties and therapeutic potency [49]. Grellier et al. exposed human MSCs to 1.2 Pa shear stress for either 30–90 min and reported an upregulation of genes related to osteogenesis [50]; for this reason, the shear stress within bioreactors could be manipulated to prime stem cells into differentiating into the desired cell type. However, there are not many reports comparing the effects of the variation of flow rates, so more studies are needed to evaluate the influence of bioreactor's microenvironment on stem cells.
Rotating-wall vessel bioreactor
To minimize heterogeneous shear stress and turbulence in cell culture in SB, NASA's Biotechnology Group developed a bioreactor design that allows for a more homogenous distribution of shear stress and encourages the round shape formation of MSCs, namely RWV bioreactor [51]. The design comprises of a cylindrical chamber with a gas exchange membrane, where the vessel body rotates around its axis. Chen et al. used the RWV system for bone marrow MSCs expansion, where a 29-fold increase of MSC number was observed while maintaining MSC markers and differentiation capability [52]. Lineage-specific MSC expansion has also been documented using RWV bioreactors [52,53]. Diederichs et al. cultured human adipose-derived MSCs on Sponceram® ceramic carrier discs in the RWV bioreactor where the result showed both an increase in cell proliferation and osteogenic gene expressions [53]. Similarly, Granet et al. showed that osteogenic and chondrogenic differentiation of MSCs were superior when using the RWV comparing to static culture, again suggesting the possibility of lineage-specific stem cell expansion [54]. Nevertheless, the computational fluid analysis shows heterogeneous shear stress and fluid velocity within scaffolds. In addition, scaffolds utilized in RWV are restricted in size and are vulnerable to collision with the bioreactor wall [55].
Perfusion bioreactor
Bioreactors can also maneuver with perfusion, where the system allows for a continuous exchange of media, introducing fresh medium while removing exhausted medium [56 –59]. In perfusion bioreactors, MSC-seeded scaffold constructs are usually placed in the chamber in a forced manner to ensure that the medium is flowing through the scaffold, allowing a more homogeneous cell distribution [60]. These configurations allow for a more flexible regulation of the internal environment and the physical structure of scaffolds. Perfusion bioreactors consist of parallel plates, hollow fibers, and fixed bed and fluidized bed bioreactors (FBB).
Parallel plate bioreactors comprise of an upper gas section separated from the liquid-filled bottom section by a gas-permeable membrane [61]. MSCs attach on the plastic surface of the bottom compartment as a monolayer, while the system provides continuous and automated feeding of media. Dennis et al. illustrated that the parallel plate bioreactor was successfully used to significantly proliferate CFU-F and progenitors with osteogenic potential from BMSCs [62].
Similar to parallel plate bioreactors, hollow fiber bioreactors operate in continuous perfusion mode and provide a low-shear stress environment. This type of bioreactor is characterized by a high surface area for cell culture, consisting of a bundle of parallel hollow fibers enclosed within a cylinder-shaped cartridge with a sterile closed loop for flow of medium and gas exchange around the fibers [61]. Numerous studies have reported the use of hollow fiber bioreactors in MSC expansion, the most commonly used being the Quantum® bioreactor [63,64]. Lechanteur et al. have reported an efficient isolation and expansion of MSCs in the Quantum bioreactor without altering therapeutic and differential potential of the cells; the bioreactor was able to reliably produce cell at a faster expansion rate when compared with normal flasks (double time 40 compared with 56 h) [65]. Nonetheless, both hollow fiber bioreactors and fixed bed bioreactors are presented with difficulties in scalability, culture monitoring, and cell harvesting [66].
As an alternative, fixed and FBB, or also known as a packed-bed bioreactor, also offer uniform hydrodynamic surroundings. In FBB, cells are seeded on an immobilized scaffold positioned in cylindrical vessel columns, also known as the bioreactor bed; the column consists of particles such as alginate beads, glass fibers, and hydrogel, either packed (fixed bed) or floating (fluidized bed) [58]. In one study, Tsai et al. expanded human MSCs in a 2.5-L CelliGen® 310 bioreactor, which consisted of immobilized Fibra-Cel® disks. Results revealed a 9.2-fold increase in cell number after 9 days of expansion without altering MSC characteristics [58]. In addition, FBB allows for controllable parameters such as pH and O2 and has been successfully utilized in embryonic stem cells, human amniotic fluid-derived MSC, and umbilical cord blood MSC expansion [67].
Compression stimulus-dependent system
Mechanical strains in vivo are also the main influencer of cell behaviors and morphologies, leading to inventions of bioreactors utilizing repeated mechanical stimulation. Examples of studies using compression bioreactors and combined systems for MSC expansion and lineage-specific differentiation are illustrated in Table 4.
↑, increase; PU, polyurethane; PU-BDI, polyurethane-based, 1–2 butanediisocyanate.
Compression bioreactors and combined systems
Compression bioreactors are constructed of a compression chamber with a piston that directly applies mechanical loads on the scaffold [68]. To optimize mass nutrient transport, compression bioreactors are usually combined with perfusion systems to further promote cell proliferation and differentiation, mainly toward osteogenic and chondrogenic lineages [69 –72]. For example, Jagodzinski et al. investigated the effect of mechanical cues in BM-derived hMSCs [70]. MSCs were cultured on bovine spongiosa discs in either a perfusion or a combined compression/perfusion bioreactor for 3 weeks; results showed that both conditions enhanced MSC proliferation, but the combined system also promoted osteogenesis. Similarly, Liu et al. reported that a combination of perfusion and cyclic compression system promoted MSC proliferation and osteogenic differentiation [72]. For cartilage, Guo et al. cultured alginate hydrogel beads with encapsulated human MSCs under, applying both shear and compressive forces [69]. This dynamic culture maintained higher cell density when compared with the static group, with an enhanced chondrogenic marker expression. Thus, conclusively, the combined system showed promising results for MSC expansion and differentiation toward bone tissue engineering; however, more studies remain to be explored to evaluate its potential for other tissues.
Tissue engineering for MSC proliferation
Scaffolds are commonly utilized in perfusion bioreactors to enhance nutrient supply and distribution, allowing culture medium to continuously flow through the pores of the scaffold. Scaffold-constructed materials range from synthetic biodegradable polymers, including poly(ethylene terephthalate) (PET), polylactic acid, polycaprolactone (PCL), or poly(
These findings illustrated that different scaffolds fabricated from different materials and methods all positively enhanced MSC proliferation; however, little is known whether these differences in properties have any influence or a slight favor toward a specific MSC lineage. Biophysical signals, including topography, stiffness, and elasticity, stimulated by scaffolds as synthetic extracellular matrices (ECM) play an essential part in priming stem cells in bioreactor microenvironment toward the desired lineage- and tissue-specific function. For instance, Engler and Discher showed that substrates with high stiffness promoted MSCs into osteoblasts, and medium and low stiffness directed MSCs into myogenesis and neurogenesis, respectively [76]. Our laboratory investigated the divergent differentiation pathways of MSCs and neotissue formation influenced by aligned and randomly oriented fibrous scaffolds [77]. We observed that the aligned group formed more mature tendon-like tissues, whereas the randomly oriented group demonstrated chondrogenesis and bone tissue formation. Optimal scaffold design should be based on the mechanical and structural characteristics of the desired native tissues, ideally bioactive, biodegradable, and biocompatible. Even so, the combination of these materials into scaffolds for MSC proliferation and differentiation is still currently under exploration.
Microcarrier-based technology
In some aforementioned mixed vessel systems, such as SBs and RWVs, suitable physical supports for anchorage-dependent cells, such as MSCs should be accommodated. Innovative three-dimensional (3D) scalable methods have been developed and improved for MSC manufacturing, namely microcarrier-based culture platform. Microcarriers are small, often spherical, particles that offer a large surface area to unit volume for cells to adhere and expand; one liter bearing 5 mg/mL microcarriers provides a total area of 1.35 m2 for cell growth [43,78]. Distinct microcarriers have their own unique physical properties, such as stiffness and nanotopography, and can be generated with a variety of materials, including gelatin, dextran, polystyrene, and alginate, and manufactured to be either nonporous (eg, SoloHill® Plastic microcarriers), macroporous (eg, CultiSpher®-S), or microporous (eg, Cytodex®-1) [78].
MSCs from different sources uniquely express distinct adhesion molecules; therefore, the best choice of microcarriers, surface coating, and culture medium crucially depends on the type of MSC to be expanded. Several types of microcarriers are now available in the market for researchers to choose the most suitable one that meets the demand of their specific cell types and targets. In several studies where microcarriers were screened, Cytodex-1 and -3, and CultiSpher S microcarriers were commonly selected for MSC expansion. Schop and his team have evaluated nine different microcarriers for the culture of bone marrow MSCs, and have concluded that the highest adhesion efficiency was obtained with Cytodex-1 microcarriers [79]. Frauenschuh et al. utilized Cytodex-1 microcarriers in SB for the attachment and expansion of porcine MSCs, where osteogenic and chondrogenic potential was maintained over a cultivation time of 28 days [45]. Timmins et al. also compared different microcarriers to promote human placental-derived MSCs proliferation in a wave bioreactor, and demonstrated that seeding efficiency was over 70% for Cytodex-1 and -3, and 90% efficiency and a 15-fold expansion were observed when cultured on CultiSpher S [80]. Using CultiSpher S, Eibes and his team also discovered a 10-fold increase in MSC numbers in spinner flask cultures using a low-serum-containing medium [44]. More recently, Goh et al. examined various microcarriers for fetal bone marrow MSC expansion in serum-supplemented medium; results concluded that Cytodex-3 led to the highest cell expansion with initial adhesion efficiency of 90% [43].
Missing Linkage Between Scalability and Clinical Applications of MSCs
Controlled differentiation of MSCs
In vivo, MSCs localize in well-defined niches, a 3D structure driving MSCs toward self-renewal and differentiation. MSC surrounding provides a balance between MSC quiescence, and proliferation and differentiation; thus, the stem cell niche must be viewed as a complex dynamic composition. Expansion and differentiation of MSCs using microcarrier-based technology have been established to be possible for large-scale expansion; however, it is important to consider the operating culture conditions that not only affect cell expansion but also cell phenotype, which influence MSC lineage commitments. Most research groups focus mainly on one of the aspects, expansion or differentiation, disregarding the missing link between the two important factors. The ideal stem cell culture condition is to combine expansion and differentiation within the bioreactor as differentiated cells are required for final clinical applications (Fig. 2). The major hurdle is the identification of the accurate time point between differentiation and expansion for different cell lineages, to prevent differentiation from competing with expansion and thus leading to insufficient cell dosage.

A diagram of lineage-specific/directed differentiation MSC expansion toward MSC-based therapies; MSCs need to be isolated, expanded, and differentiated in an enclosed, large-scale, automatic system in a combination with suitable microspheres/scaffolds, bioactive molecules, and growth factors.
Because MSC phenotype is different after expansion at different culture conditions, tissue-specific MSC expansion systems based on the target tissues also need to be developed. A few teams have had some success in this pursuit with human embryonic stem cells (hESCs). Bardy et al. demonstrated a microcarrier system for hESCs expansion and its subsequent differentiation into neural progenitor cells (NPC) [81]. A 20-fold hESC expansion was achieved using stirred SF, followed by a simple serum-free differentiation protocol into NPCs. This integrated procedure yielded 333 NPCs per seeded hESC as compared with 53 in the classical two-dimensional static cultures, suggesting a possible method toward scalability of lineage-specific stem cells. The use of microcarrier-based bioreactors for the manufacturing of endoderm progeny cells from hESCs has also been explored, as Lock and Tzanakakis illustrated the expression of definitive endoderm markers, including FOXA2 and SOX17, after hESC expansion on microcarriers in stirred SBs [82]. Osteogenic differentiation is another area where microcarriers and scaffolds play a crucial role in expansion. Shekaran et al. cultured human early MSCs (heMSCs) on PCL microcarriers coated with ECM in a wave bioreactor and successfully enhanced cell expansion and osteogenic induction [83]. Similarly, Goh and his research team demonstrated improvement in cell expansion capacity when they cultured human fetal MSCs on Cytodex-3 microcarrier, as well as higher osteogenic potency when differentiated in vitro onto 3D scaffolds or implanted in vivo [43]. Since MSC differentiation or priming is necessary for final therapeutic applications, understanding the influence of cellular microenvironments on MSC differentiation toward a specific lineage is crucial to optimize bioreactor strategy.
Therapeutic mechanisms of expanded MSCs in vivo
Exogenously administered MSCs in animals displayed the ability to migrate to the damaged site, and exert beneficial effects through bioactive factor secretions [84]. Bioactive factors released by MSCs were illustrated to have angiogenic and antiapoptotic properties, preventing further tissue damage and establishing new blood supply [84]. Moreover, the immunosuppressive properties of MSCs relating to the immune system are well documented; in vitro studies demonstrated that MSCs suppressed T lymphocyte and natural killer (NK) cell activation and proliferation [85]. Nevertheless, the mechanism responsible for these outcomes is not yet completely understood as it is still unclear if the MSC phenotype exists in vivo.
Understanding the therapeutic mechanisms of MSCs in vivo is crucial for the development and translation of cell-based therapies into clinical practices. The corresponding correlation concerning the phenotypic characteristics and the biological function of MSCs is not yet clearly understood, as there are no evaluation parameters to predict expanded MSC functions and safety in vivo. Current universal evaluations mainly base on CD markers and three lineages' differentiation; therefore, it is necessary to also focus on the cellular and genetic signature of MSCs and develop a predictable evaluation system to quality MSCs after expansion.
Our insufficient knowledge of the basic biological workings of MSCs is reflected in the diverse culture conditions utilized by different research groups. Multiple research groups are attempting to mimic site-specific conditions; however, once isolated, MSCs are subjected to environments that are different from those found in their native location. For instance, one study evaluated the effects of a spinner flask culture system on BMSCs seeded on BMP-2-loaded silk scaffold toward osteogenesis [86]. Results showed that the dynamic conditions promoted bone-like tissue formation in vitro; however, no significant differences in bone formation capability were observed in vivo when compared with the static conditions. Additionally, MSCs may possibly exist in vivo as heterogeneous populations, further adding complexity to their study in vitro, as the purification protocols or the culture condition imposed may select for specific subsets of cells or cellular characteristics. Taken together, more studies need to be reported to draw a definite conclusion regarding the actual potential and mechanisms of bioreactor-expanded MSCs in supporting tissue formation and regeneration in vivo. Moreover, the therapeutic viability and proliferative capacity of the bioreactor-expanded MSCs need to be assessed before they are applied clinically.
In addition, for future advances, studies should be focused on better understanding the function of endogenous MSCs. Learning about the function of these cells will provide more information on their true potential, revealing molecules and pathways to therapeutically target. From this information, certain small molecules could be utilized to expand the MSC pool in vivo for regeneration, instead of going through the expansion process.
Sources of MSCs
Over the last several years, the choice of MSC sources varied significantly among research groups, and is based on “trial and error.” Increasing attention on MSCs has led to questioning the equivalence and characteristics of MSCs isolated from different sources. As mentioned, MSCs can be isolated from various tissues, including bone marrow, adipose tissue, placenta, or umbilical cord; however, although they all illustrate comparable basic biological features, considerable differences among them still exist [3]. Kwon and his team isolated and cultured MSCs from amnion (AMSCs), chorion (CMSCs), and umbilical cord (UC-MSCs) from a single donor, and illustrated that they have different proliferation and differentiation potential [87]. Results from microarrays and RT-qPCR also showed that they all have distinguished gene expression profiles; AMSCs displayed higher expression of genes associated with apoptosis and cellular aging, whereas CMSCs and UC-MSCs overexpressed some genes related to heart morphogenesis and neuronal development, respectively. A similar study was conducted to investigate the differences of MSCs derived from BMSCs, adipose tissue, umbilical cord tissue, and menstrual blood (MenSCs) [88]. Based on gene expression data, 738 MenSC genes were distinctively upregulated or downregulated when compared with BMSCs and UC-MSCs from the same donor; MenSC genes were found to be superior in terms of neural differentiation capacities, whereas BMSCs and UC-MSCs were superior in terms of osteogenic and chondrogenic differentiation potentials.
Another study compared BMSCs isolated with different procedures, namely isolation based on adhesive properties (BM-MSCs) versus density gradient centrifugation (iBM-MSCs), and revealed that even though they have similar behavior in vitro, the two types illustrated different therapeutic properties when injected in vivo in a critical limb ischemia model. BM-MSCs were superior to iBM-MSCs in maintaining tissue viability and secreting cytokines crucial for vessel remodeling [89]. Hence, a high-throughput screen of the functional diversity and the possible clinical applicability of each cell types need to be extensively evaluated, as it can help select an adequate cell source for utilization in specific diseases.
Final Remarks and Future Perspectives
MSCs with their abilities for extensive proliferation and multilineage differentiation can be the solution to a renewable source of cellular materials for cell-based therapy. As mentioned, the development of manufacturing process for large-scale production of MSCs will be required before MSC-based therapy becomes a reality, and that process must also be well defined and be under controllable parameters. Multiple technologies and procedures, including culture condition modifications, bioreactor technologies, and microcarrier and scaffold applications, have been successfully generated and demonstrated to expand MSCs. However, given the heterogeneity of cell types, signaling pathways, and disease indications, it is unlikely that a “one-size-fits-all” standardized manufacturing platform will be a solution for all cell-based therapies.
Increasing interest also geared toward another approach for undifferentiated MSC culture where cells are cultured as scaffold-free 3D aggregates/spheroids or multicellular aggregates, termed microtissues [90]. Since studies have shown that this approach may enhance MSC therapeutic and multilineage differentiation potential, methods have been developed allowing the culture of these 3D aggregates in bioreactor systems, including SF and RWV bioreactors [48,91]. Even though microtissues may have attractive features, more understanding of their formation mechanism and structure–function relation is needed to guide their clinical applications.
Conclusively, there are still obstacles ahead in implementing MSC therapy in mature clinical practices. Further research is needed on the therapeutic mechanisms of MSCs in vivo. Moreover, there are still insufficient understandings on which source of MSC should be applied and which administration route is proper for a particular disease. Suitable scalable manufacturing processes for MSCs, including expansion protocols, product quality, and safety controls, must also be developed without negatively affecting the therapeutic properties of the cells to meet the clinical need.
As discussed, currently much research has gone into experimenting and optimizing different variables in MSC culture environment for optimal MSC expansion. However, as lineage-primed cells or differentiated cells are often required for translation into clinical practices, careful consideration must be given not only to the expansion process but also to the effect a particular process has on cell properties. Thus, we can conclude that bioreactor systems for cultivation of stem cells are expected to contribute toward the future development of platforms appropriate for clinical-grade manufacturing of stem cell-derived products. Progress in this area toward clinical applications will most likely rely on the complete understanding of MSC underlying mechanisms, accurately engineered cell microenvironment, and computational models for high-throughput screening.
Footnotes
Acknowledgments
This work was supported by the National High Technology Research and Development Program of China (863 Program) (2015AA020303), National Key Research and Development Program of China (2016YFB0700804), NSFC grants (81630065, GZ1094, 81472115, and 81672162), Key Scientific and Technological Innovation Team of Zhejiang Province (2013TD11).
Author Disclosure Statement
No competing financial interests exist.