
Abstract
Sjögren's syndrome (SS) is characterized by autoimmune activation and loss of function in the salivary glands. Recent studies reported that bone morphogenetic protein 6 (BMP6), which is a member of transforming growth factor beta (TGF-β) superfamily, was highly expressed in SS patients. To investigate the role of BMP6 in SS, we treated the salivary gland-derived mesenchymal stem cells (SGMSCs) with BMP6 and found that BMP6 could impair immunomodulatory properties of normal SGMSCs by downregulating the Prostaglandin E2 synthase through DNA-binding protein inhibitor-1. Neutralizing the BMP6 could significantly restore the SGMSC's immunoregulatory function in vitro and delay the SS disease activity in vivo. In conclusion, BMP6 could not only affect the secreting function of epithelial cells in the salivary gland but also influence the immunomodulatory properties of SGMSCs, which may trigger or enhance the autoimmune reflection in SS.
Introduction
S
Stem cell-based therapies have been applied to repair damaged tissues and organs. Mesenchymal stem cells (MSCs) are multipotent stem cells with the capacity to differentiate into osteoblasts, chondrocytes, adipocytes, and neural cells. Most importantly, MSCs have regulation capacity on the immune system particularly in the context of inflammatory-mediated disorders [4 –6]. A previous study has suggested that the pathogenesis of SS may be related to the impaired function of salivary gland-derived mesenchymal stem cells (SGMSCs) in SS patients [7]. However, the underlying mechanisms responsible for the deficiency of function in MSCs from SS patients and animals remain unclear.
A recent study reported that bone morphogenetic protein 6 (BMP6) was highly expressed in minor salivary glands from SS patients, and overexpression of BMP6 in normal mice can induce hypofunction of salivary glands [8], which shed light on the studies of the pathogenesis of SS. In the present study, we found a higher level of the BMP6 expression in SGMSCs from SS animal model (NOD/Ltj mice), and BMP6 could impair immunomodulatory properties of normal SGMSCs. Moreover, anti-BMP6 treatment could attenuate inflammation and delay the loss of function in the salivary gland of NOD/Ltj mice. Our study provides the first demonstration of a link between BMP6 and the immunomodulation disorder in SS.
Materials and Methods
Mice
Female NOD/Ltj mice served as SS animal model in this study, whereas female BALB/c mice served as control. NOD/Ltj mice were purchased from Beijing HFK Bioscience Co. BALB/c mice were obtained from the Institute of Laboratory Animal Science, Chinese Academy of Medical Sciences. All animal experiments were approved by the Animal Care and Use Committee of Capital Medical University.
Patient samples
Labial gland samples were collected from 10 patients with dry mouth complaints in the Salivary Gland Disease Center, Capital Medical University School of Stomatology (Beijing, China). Six patients (all female, 55.00 ± 6.708 years of age) were diagnosed as primary SS [9] four patients (all female, 41.75 ± 6.6 years of age) without saliva secretion loss or infiltration inside salivary gland served as control. All patients gave written informed consent before the study commenced. This study was approved by the Ethics Committees of Beijing Stomatological Hospital and Capital Medical University (Beijing, China).
Isolation and culture of MSCs in vitro
Mouse SGMSCs were isolated and cultured as described previously [10,11]. In brief, submandibular glands of 8-week-old BALB/c mice or NOD/Ltj mice were dissected and minced with a scalpel, digested in HBSS with 4 mg/mL collagenase II at 37°C with constant shaking for 30 min. Then the tissue solution was filtered through 40 μm cell strainers, and centrifuged at 500 g for 5 min at 4°C. Next, the cell pellets were resuspended with Dulbecco's modified Eagle's medium (Invitrogen) supplemented with 20% fetal bovine serum (FBS), 2 mM
Initially incubated for 2 h under 37°C at 5% CO2 condition, the cell culture supernatant was transferred to a new 100-mm dish and incubated for another 2 h, and then transferred to a new 100-mm dish. Two hours later, supernatant in all the dishes was removed and nonadherent cells were discarded. The attached cells were cultured for additional 7 days. Attached cells formed by the colonies were passed once for further experiments. CD34, CD45, CD44, CD90, CD105, and CD146 were stained for flow cytometry as the positive or negative surface markers of MSCs.
Lymphocyte proliferation assay
T cell and B cell proliferation was analyzed by Carboxyfluorescein succinimidyl ester (CFSE) distributions. Mouse lymph nodes were taken from BALB/c mice. Naive T cells were isolated from the lymphocytes by the Naive CD4+ T Cell Isolation Kit (Miltenyl Biotec) or the Naive CD8+ T Cell Isolation Kit (Miltenyl Biotec), B cells were isolated by the CD19 Isolation Kit (Miltenyl Biotec) following the instructions. For Naive CD4, CD8 T cell or B cell coculture, cells were labeled by 5 mM CFSE (Invitrogen) for 15 min and washed twice by cold PBS. For Th1, Th2, or Treg coculture, naive CD4 T cells were first cultured with mIL-12 (10 ng/mL), mIL-4 (10 ng/mL), or mTGF-β1 (2 ng/mL) separately for 1 day, and then labeled with CFSE.
For cocultured system, MSCs were pretreated with BMP6 or anti-BMP6 for 2 days and washed three times by PBS. T cells or B cells were seeded at the ratio of 1:10 (or 1:1 for naive CD4+ T cells) and cultured for 3 days. Soluble anti-CD3 (1 μg/mL) and anti-CD28 antibody (1 μg/mL) were used to activate T cells, and soluble anti-CD40 (10 μg/mL) was used to stimulate B cells. RPMI 1640 media (Invitrogen) supplemented with 10% FBS, 2 mM
In vitro osteogenic and adipogenic differentiation assays
For osteoinduction in vitro, SGMSCs were cultured under osteogenic culture conditions containing 2 mM
For adipoinduction in vitro, the Adipogenesis Differentiation Kit (Invitrogen) was used. Two weeks after the adipoinduction, cultures were stained with Oil Red O (Sigma). After photographing, Oil Red O stains were extracted with 100% isopropanol for 5 min and read at 492 nm. Total messenger RNA (mRNA) was isolated from the mouse SGMSCs 1 week after osteoinduction or adipoinduction. Expressions of bone gamma-carboxyglutamate protein (Bglap), Alpl, peroxisome proliferator-activated receptor-γ (Pparg) and fatty acid-binding protein 4 (Fabp4) were assayed by real-time reverse transcription polymerase chain reaction (RT-PCR). All experiments were repeated in three independent samples for each group.
Small interfering RNA transfection
Around 5 × 105 SGMSCs were seeded on a six-well culture plate. Id1 small interfering RNA (siRNA; sc-35632; Santa Cruz) and control siRNA (sc-36869; Santa Cruz) was used to treat the SGMSCs according to the manufacturer's instructions.
Mice salivary flow rate test
Mice salivary flow rates were monitored as described previously [12]; mice were anesthetized with a solution of Ketamine (100 mg/mL; Beijing Double-Crane Pharmaceutical Co.) and Xylazine (20 mg/mL; Sigma) in sterile water, given intraperitoneally (1 μL/g of body weight). Salivary secretion was stimulated using 0.1 mL/kg body weight of pilocarpine (50 mg/mL; Beijing Double-Crane Pharmaceutical Co.) subcutaneously. Saliva collection began within 2 min of pilocarpine administration. Animals were positioned with a 75-mm hematocrit tube placed in the oral cavity and whole saliva was collected into preweighed 0.75-mL Eppendorf tubes for 10 min. The amount of saliva collected was determined gravimetrically.
BMP6 and neutralization antibody treatment
For in vitro study, the MSCs were treated with 10 ng/mL BMP6 based on a dose-dependent experiment (Please see Supplementary Methods; Supplementary Fig. S1C; Supplementary Data are available online at
Histological analysis of salivary glands
Salivary glands were isolated and fixed with 4% PFA for 24 h at 4°C; paraffin sections were used for Hematoxylin and Eosin staining. After Hematoxylin and Eosin staining, the sections were photographed by microscope (BX51; Olympus) with CCD (DP72; Olympus). Area of inflammatory focus (containing >50 lymphocytes per 4 mm2 tissue) was calculated per field at × 200 magnification ( × 20 objective lenses) by Image-Pro Plus Version 6.0 software (Media Cybernetics). Six entire salivary gland sections for each animal were counted with an average of 10 fields/gland by an experienced expert of histopathology under blinded fashion.
Real-time RT-PCR
Total RNA was isolated by an RNA Isolation Kit (Tiangen) following the manufacturer's instructions, and complementary DNA (cDNA) was synthesized from 100 ng of total RNA using the Reverse Transcription Kit. Real-time polymerase chain reaction (PCR) was analyzed using the δδ-Ct method.
Primers were used as follows: BMP6 (Hs01099594_m1; Invitrogen), GAPDH (Hs02786624_g1; Invitrogen), Bmp6 (Mm01332882_m1; Invitrogen), Gapdh (Mm99999915_g1; Invitrogen), Bglap (Mm03413826_mH; Invitrogen), Alp (Mm01187117_m1; Invitrogen), Pparg (Mm00440940_m1; Invitrogen), Fabp4 (Mm00445878_m1; Invitrogen), Ido1 (indoleamine 2,3-dioxygenase, Mm00492590_m1; Invitrogen), Nos2 (nitric oxide synthase, Mm00440502_m1; Invitrogen), TGFb1 (Mm00441727_g1; Invitrogen), Id1 (Mm00775963_g1; Invitrogen), Id2 (Mm00711781_m1; Invitrogen), Id3 (Mm00492575_m1; Invitrogen), Id4 (Mm00499701_m1; Invitrogen), Cox2 (cyclooxygenase 2, Mm03294838_g1; Invitrogen), Aqp5 (aquaporin 5, Mm00437578_m1; Invitrogen), Slc12a2 (Mm01265951_m1; Invitrogen), Itpr3 (inositol 1,4,5-trisphosphate receptor type 3, Mm01306070_m1; Invitrogen), and Chrm3 (cholinergic receptor muscarinic 3, Mm00446300_s1; Invitrogen).
Enzyme-linked immunosorbent assay
Mouse peripheral blood was collected from the retro-orbital plexus and centrifuged to obtain serum. Tissue lysates were extracted from salivary glands with a Sonics Ultrasonic process on ice in NP-40 buffer (150 mM NaCl, 1.0% NP-40, 5 mM EDTA, 50 mM Tris pH 8.0) with 2 μg/mL Aprotinin, 5 μg/mL Leupeptin, 1 μg/mL Pepstain A, 1 mM PMSF, and 1 mM sodium orthovanadate. The resulting homogenates were centrifuged at 1,000 g and supernatants were kept for future use. Protein concentrations were tested by the Bicinchoninic Acid Protein Assay Kit (Sigma). Prostaglandin E2 (PGE2) was assessed by the Mouse ELISA Kit (R&D Systems) according to the manufacturer's instructions.
Statistical analyses
All data have a normal distribution and are presented as mean ± standard error of the mean (SEM; in data of infiltrating area statistics) or standard deviation (in other data) of three independent experiments, and we used an α level of 0.05 for all statistical tests. The mice salivary flow rates were statistically analyzed with repeated measurement; other data were analyzed with student's t-test, one-way analysis of variance (ANOVA), or two-way ANOVA. Analysis was performed with SPSS version 17.0.
Results
BMP6 was expressed at a higher level in salivary gland from SS patients and NOD/Ltj mice and regulated SGMSCs function
Our previous studies suggest that abnormal MSCs play a role in primary SS [12]. A recent study suggests that MSCs from salivary glands of SS patients exhibit impaired function compared with those from healthy individuals [7], and an increased expression of BMP6 in patients with primary SS and animal model is associated with functional impairment of the salivary gland [8]. In this study, we found that the BMP6 expression in salivary glands from SS patients and from the animal model (NOD/Ltj mice) is higher (Fig. 1A, B). Salivary gland-derived MSCs (Supplementary Fig. S1A, B) from NOD/Ltj mice also expressed a higher level of BMP6 compared with BALB/c (Fig. 1C).
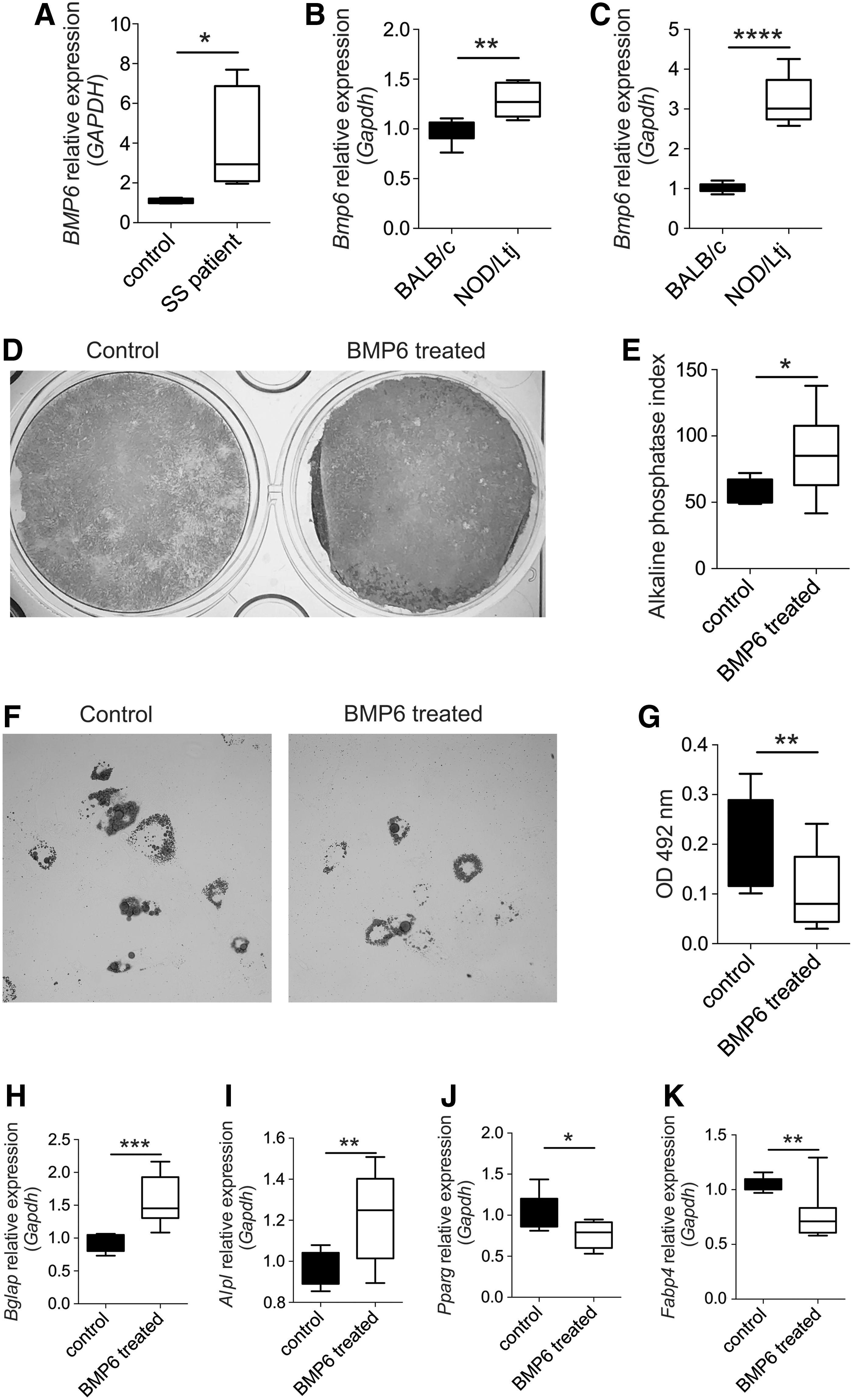
BMP6 was expressed at a higher level in salivary gland from SS patients and NOD/Ltj mice and regulated SGMSC function.
ALP staining and activity test revealed that ALP activity was much higher in BMP6-treated cells than in untreated cells (Fig. 1D, E). Consistently, real-time RT-PCR showed that the expression of the osteogenic markers Bglap and Alp were significantly higher in BMP6-treated cells (Fig. 1H, I). After adipogenic induction, cells treated with BMP6 showed less cellular lipid accumulation with control cells, as evidenced by Oil Red O staining, the optical density value of extraction (Fig. 1F, G). Real-time RT-PCR results revealed that following BMP6 treatment, the expression of adipogenesis-induced genes, including Pparg and Fabp4, was dramatically decreased (Fig. 1J, K).
BMP6 treatment impaired immunomodulatory properties of SGMSCs by downregulating Cox2 and PGE2 levels and promoting T cell proliferation
To investigate whether BMP6 treatment can affect dendritic cells, macrophages, or T cells directly, we treated those cells with BMP6 and did not find any significant change (Supplementary Fig. S2). Then, we treated BMP6 on SGMSCs to see if their immunomodulatory properties were impaired. Real-time PCR showed that BMP6 could only downregulate the PGE2 synthase Cox2 (Fig. 2A) but not Ido1 (Fig. 2B), Nos2 (Fig. 2C), or Tgfb1 (transforming growth factor beta 1) (Fig. 2D).
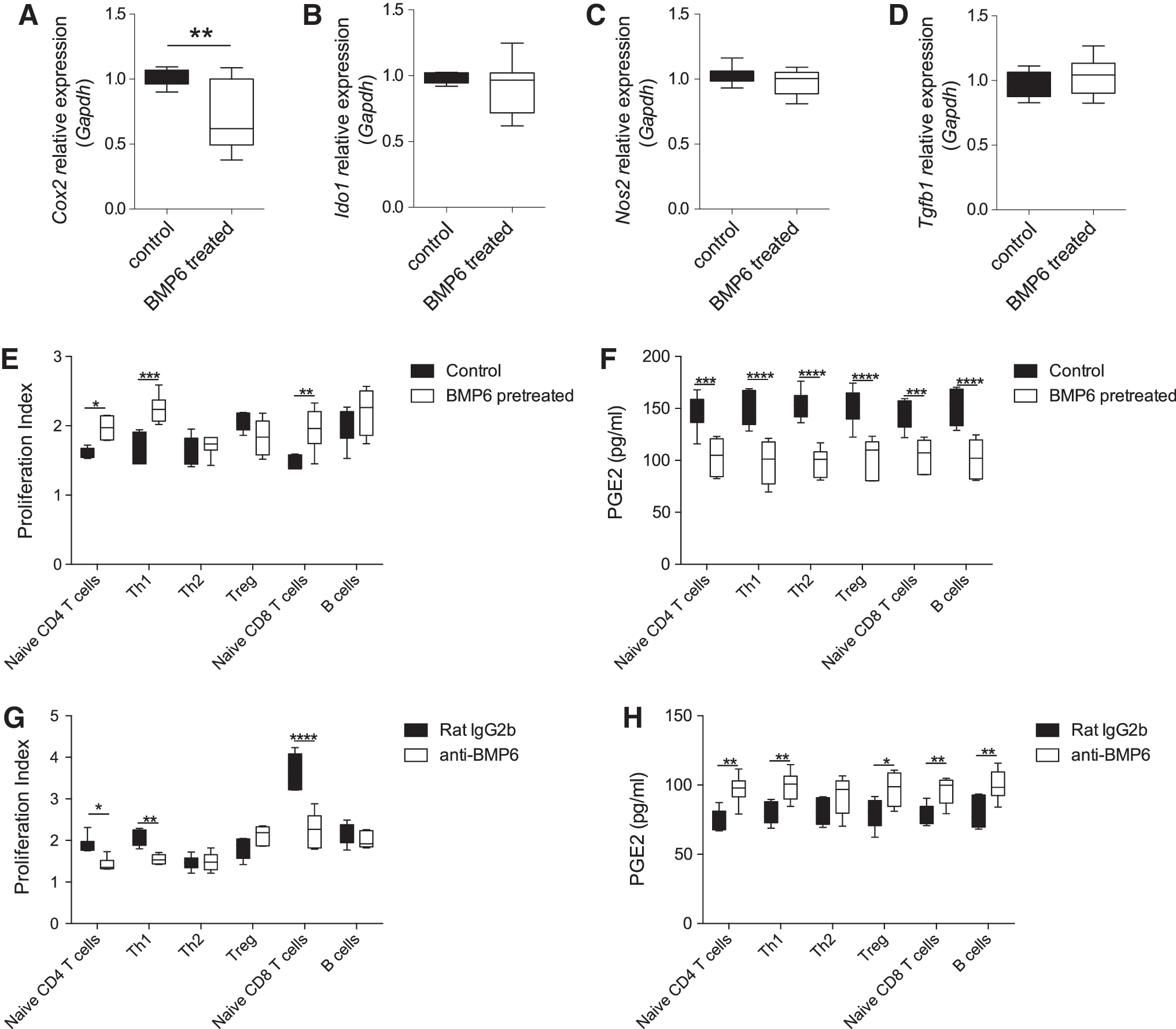
BMP6 treatment impaired immunomodulatory properties of SGMSCs by downregulating Cox2 and PGE2 levels and promoting T cell proliferation.
In the SGMSCs/T cell coculture system, the proliferation index of naive CD4+ T cells (Supplementary Fig. S3) cultured alone was 3.32 ± 0.21 (mean ± SEM), whereas SGMSC coculture group only reached 1.61 ± 0.03, and BMP6-pretreated SGMSC cocultured group is 1.97 ± 0.07, suggesting that SGMSCs have very active immunomodulatory functions, which can be impaired by BMP6. The proliferation of naive T CD4+ cells, Th1, and CD8+ T cells were inhibited significantly by coculturing with normal SGMSCs compared with BMP6-treated SGMSCs (Fig. 2E and Supplementary Fig. S3). Due to the proliferation, significant differences of cytokine expressions, such as IFN-gamma and TNF-alpha can also be found (Supplementary Fig. S4). ELISAs showed that SGMSCs treated with BMP6 decreased the concentration of PGE2 (Fig. 2F) in the supernatants of the coculture system.
In contrast, BMP6-neutralizing antibodies (anti-BMP6) could significantly enhance the immunomodulatory properties in SGMSCs from NOD/Ltj mice. Naive CD4+ T cells, Th1, and CD8+ T cells showed decreased proliferative capacity when cocultured with anti-BMP6-pretreated NOD SGMSCs compared with the isotype antibody group (Fig. 2G and Supplementary Fig. S3). Consistently, anti-BMP6-pretreated NOD SGMSCs significantly increased the levels of PGE2 (Fig. 2H) and decreased some cytokines (Supplementary Fig. S4) in the supernatants of the coculture system.
BMP6 impaired immunomodulatory properties of SGMSCs through inhibitor of DNA-binding proteins 1 (Id1)
Since inhibitors of DNA-binding proteins (IDs) are suggested to be BMP-response transcription factors [14], we next investigated whether IDs were involved in BMP6-mediated SGMSC function impairment. We first found that Id1 mRNA was expressed at a higher level in SGMSCs derived from NOD/Ltj mice compared with those from BALB/c mice (Fig. 3A), whereas Id2, Id3, and Id4 mRNA showed no significant difference between the two groups (Fig. 3B–D). When BALB/c SGMSCs were treated with BMP6, the expression level of Id1 mRNA was increased (Fig. 3E).
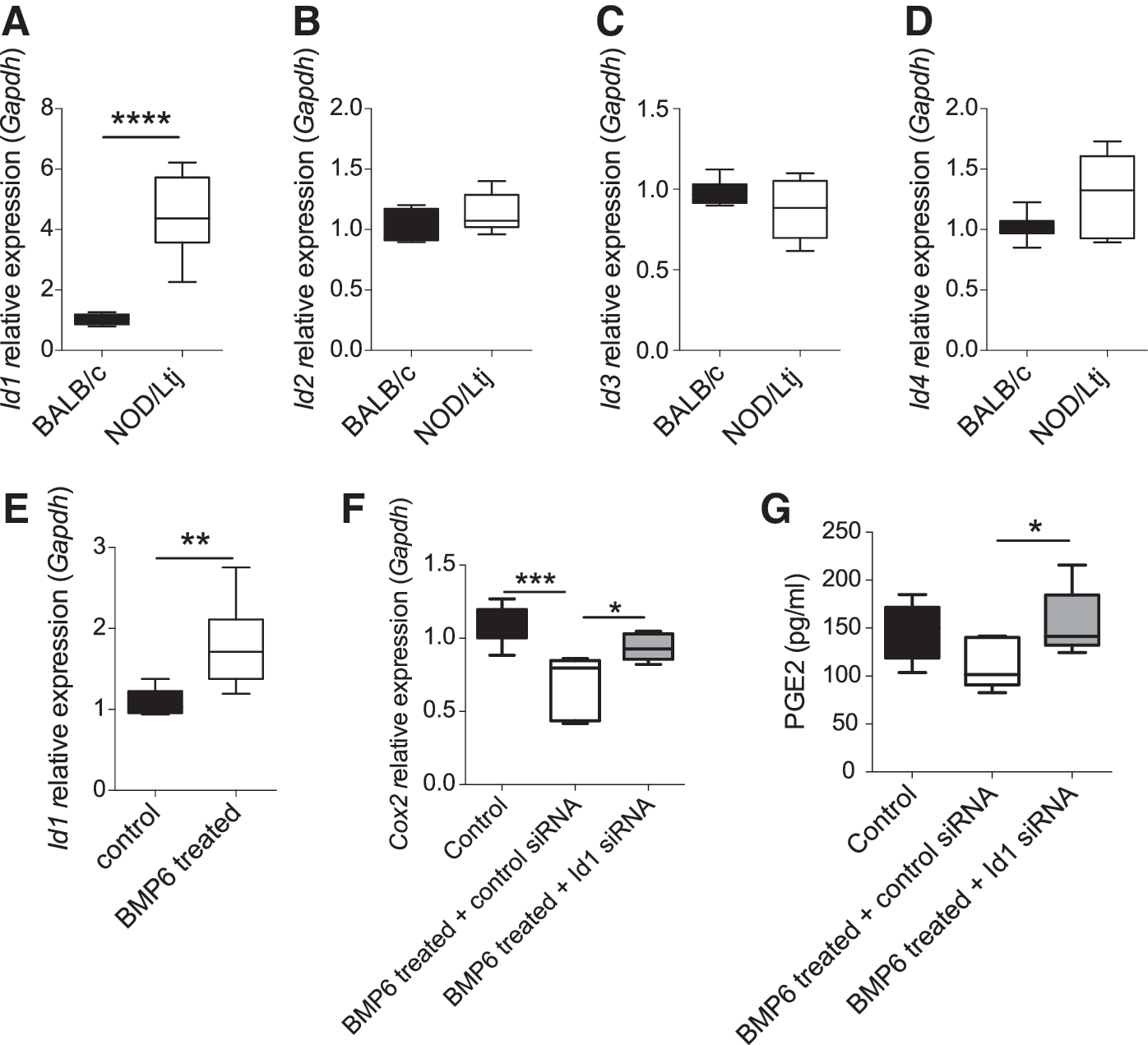
BMP6 impaired immunomodulatory properties of SGMSCs through Id1.
We next analyzed the expression levels of some molecules associated with the immunomodulatory properties of SGMSCs, including PGE2 and its synthase Cox2 in SGMSCs treated with or without BMP6 or Id1 siRNA. Real-time RT-PCR results revealed that knockdown of Id1 could increase BMP6-induced Cox2 downregulation (Fig. 3F) and ELISA of PGE2 also showed the same trend (Fig. 3G). These data indicate that Id1 may be involved in BMP6-mediated impaired immunomodulatory properties of SGMSCs by downregulating Cox2 and PGE2.
Anti-BMP6 treatment slow salivary gland hypofunction and attenuated inflammation in NOD/Ltj mice
To test the effect of BMP6 on salivary gland function, NOD/Ltj mice received intraperitoneal injections of BMP6-neutralizing antibody for 3 days. Histological analysis showed that the submandibular glands of the anti-BMP6-treated group contained less inflammatory infiltrations than glands of the isotype-treated group (Fig. 4A, C). Saliva was collected from 6 to 10 weeks old. Measurements of salivary flow rate showed that salivary flow rate gradually declined over time. However, the loss of saliva production in the group treated with anti-BMP6 was delayed compared with that of the isotype-treated group (Fig. 4B) in 8 and 9 weeks old.
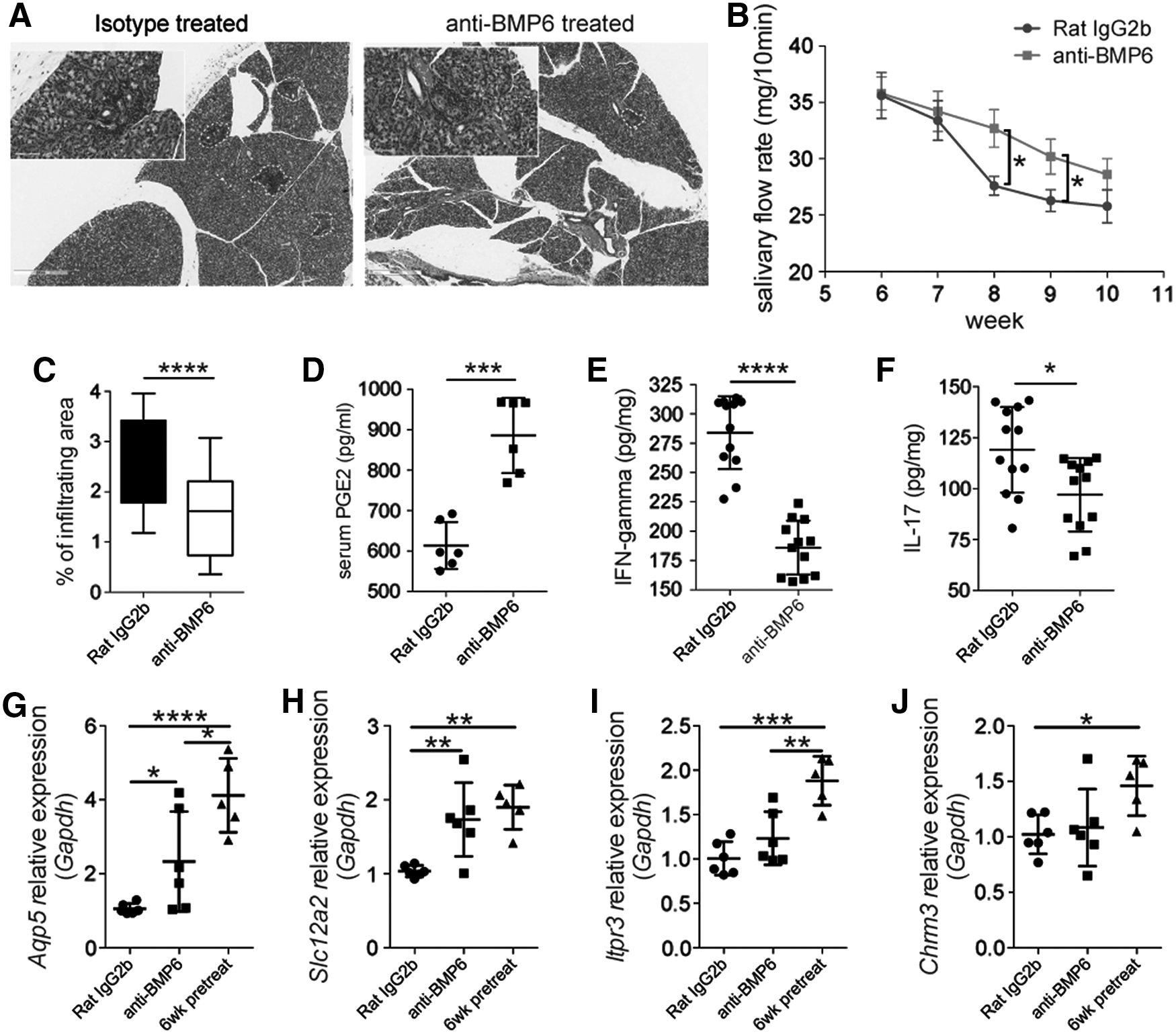
Anti-BMP6 treatment slowed salivary gland hypofunction and attenuated inflammation in NOD/Ltj mice.
We next examined whether anti-BMP6 treatment could alleviate local and systemic inflammatory response in vivo. Compared with isotype treatment, anti-BMP6 increased the levels of serum PGE2 (Fig. 4D), and decreased the levels of IFN-gamma (Fig. 4E) and IL-17 in salivary gland (Fig. 4F). Taken together, these data suggested that anti-BMP6 could slow the salivary gland hypofunction in NOD/Ltj mice by increasing systemic PGE2 and decreasing local IFN-gamma and IL-17. Real-time RT-PCR results revealed that anti-BMP6 could protect some salivary gland secreting function-related gene expressions, such as Aqp5 and Na-K-Cl cotransporter (Slc12a2), but had no effect on the expression of Itpr3 and Chrm3 (Fig. 4G–J).
Discussion
BMP6 was reported to be highly expressed in the epithelia of the salivary gland in patients with SS. Although people already knew that overexpression of BMP6 locally could induce a decrease of cellular water permeability and salivary gland hypofunction, as well as an increase of infiltrates in the glands [2,8], it is unknown how BMP6 effects immune cells locally. In the present study, we treated dendritic cells, macrophages, and T cells with BMP6 directly, and no significant change was observed (Supplementary Fig. S2). On the other hand, we found that BMP6 treatment can impair the immunomodulatory functions of MSCs derived from salivary glands, which may contribute to the inflammation of salivary glands in SS.
MSCs are multipotent stem cells that can differentiate into a variety of cell types, and were found to have immunoregulatory functions [15 –18] and can regulate the immune response in many diseases [17,19,20]. MSCs from patients with autoimmune diseases, including SS, were reported to have deficient immunoregulatory functions and biological properties [12,21,22]. Until now, factors that caused those functions to be impaired are not very clear.
It has been proposed that MSCs can regulate the adaptive and innate immune systems by several soluble factors to mediate immunosuppressive effect [20], including PGE2 [16], IDO [15], nitric oxide (NO) [4], and TGF-β1 [23]. In this study, we found that BMP6 can only downregulate the gene expression of PGE2 synthase Cox2 but not Ido1, Nos2, and TGFb1 in SGMSCs. PGE2 has a mostly immunosuppressive role in T cell activation and proliferation [24]. PGE2 can inhibit IL-2 production and also suppresses T cell receptor signaling [25,26], inhibits IL-2 and IFN-γ production [27], and upregulate IL-5 in T cells [28].
Moreover, several reports suggest that PGE2 enhances the induction and differentiation of regulatory T cells [29] and induce T cell anergy [30]. In the present study, BMP6 pretreating can decrease the PGE2 secretion in SGMSCs, leading to a lower level of inhibition to naive T CD4+, CD8+ cells, and Th1 proliferation in the SGMSC/T cell coculture system; anti-BMP6-neutralizing antibody can significantly increase PGE2 in SGMSCs from NOD mice. These results indicate that BMP6 downregulating PGE2 might be the primary reason for SGMSCs' immunomodulatory deficiency in SS.
We also directly used PGE2 to treat NOD mice at the dose of 6 mg/kg, twice a day for 16 days [31]; interestingly, the treatment group had observed slight diarrhea. We can see a decrease in inflammation area in the salivary glands (data not shown), but consistent results of salivary flow rates were not obtainable. We hypothesize that PGE2 produced by MSCs may be enriched locally to contribute to a “stem cell niche,” which is different from systemically distributed PGE2.
Id1 is a helix-loop-helix (HLH) protein that can form heterodimers with members of the essential HLH family of transcription factors [32]. It is reported that the TGF-β superfamily of transcription factors can regulate Id1. BMP is a member of the TGF-β family, which bind to transmembrane type I and type II receptors [33], trigger the phosphorylation of the receptor-regulated R-Smads, Smad1, Smad5, and Smad8 in the cytoplasm. The Id1 promoter is a BMP responsive element [14], Id1 expression can be increased in response to BMP signaling in different cells [34 –37]. In this study, we also found that SGMSCs from SS mice model expressed at a higher level of Bmp6 and Id1; for control mice, treatment of BMP6 can also upregulate Id1 gene expression in SGMSCs. Moreover, knocking down the Id1 by siRNA can decrease the BMP6-induced Cox2/PGE2 inhibition, indicating that Id1 can downregulate the Cox2/PGE2 level triggered by BMP signaling.
Taken together, BMP6 was expressed at a higher level in both salivary glands and MSCs derived from NOD/Ltj mice, which regulated the immunomodulatory properties of MSCs through Id1—Cox2/PGE2 chain. Neutralizing the BMP6 can significantly restore the MSCs function in vitro and delay the SS disease activity in vivo. The results herein support both the need for further study of the possible role of impaired SGMSCs in SS and the use of PGE2 for autoimmune diseases. Importantly, these studies also suggest the possibility that pharmacological inhibitors of BMP6 may be useful locally or systemically, in limiting the inflammatory infiltration and salivary hypofunction that occurs in patients with SS, a hypothesis that is directly testable.
Footnotes
Acknowledgments
This study was supported by grants from the National Science Foundation of China (81300896 to J.X., 81470759 to H.W., and 81600829 to Y.S.); Beijing Municipal Natural Science Foundation (7142069 to J.X. and 7123215 to H.W.); and Beijing NOVA program (2015B062 to J.X.)
Author Disclosure Statement
All authors have no potential conflicts of interest.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.