
Abstract
Transplantation of neural stem/progenitor cells (NSPCs) following spinal cord injury (SCI) is a promising strategy to enhance regeneration but is limited by poor survival of grafted cells. Determining methods to enhance survival of NSPCs is therefore essential. Positive modulation of AMPA receptors has been shown to enhance neurogenesis in various models of brain injury. Here we examined the effect of selective AMPA receptor modulation in adult rat spinal cord-derived NSPCs using a class of allosteric AMPA receptor modulators known as ampakines. NSPCs from the periventricular region of the adult rat spinal cord were treated with ampakines CX614 and CX546 for 72 h either alone or in the presence of low-dose glutamate (50 μM). Treatment with either agent in the presence of glutamate significantly increased cell survival and proliferation and reduced cell death. Moreover, ampakine/glutamate treatment reduced cell death in the setting of oxidative stress. Treatment with ampakines did not significantly alter cell phenotype. These findings offer important insight into a potential therapeutic strategy to positively regulate transplanted and endogenous adult spinal cord-derived NSPCs after SCI.
Introduction
S
Previously, we have demonstrated that the excitatory neurotransmitter glutamate, which is released after SCI, promotes survival and proliferation and attenuates oxidative stress-induced cell death in adult spinal cord-derived NSPCs in vitro. Moreover, these effects were mediated by AMPA/kainate glutamate receptors [5]. Determining methods to selectively target AMPA receptors while avoiding the toxic effects of glutamate on mature neuronal and glial cells may therefore offer a therapeutic strategy to promote NSPC survival after SCI.
A class of allosteric AMPA receptor modulators, known as ampakines, has been shown to positively regulate neurogenesis in the brain [6]. Ampakines stabilize the AMPA receptor in an open state following the binding of glutamate thus prolonging current flow through the receptor and enhancing synaptic response. Ampakines have been shown to facilitate long-term potentiation, upregulate BDNF in the brain, promote synaptic plasticity in rodent models of neurodegenerative diseases [7 –9], and enhance functional recovery following stroke [10]. Recently, the ampakine CX546 was found to increase proliferation and neuronal differentiation of subventricular zone stem/progenitor cells derived from neonatal mice brains [11]. However, the effect of allosteric AMPA receptor modulation using ampakines on adult spinal cord-derived stem cells remains unknown.
This study examined the effect of two ampakine compounds, CX614 and CX546, on adult rat spinal cord-derived NSPCs. We show for the first time that treatment with CX614 or CX546 increases survival and proliferation of spinal cord NSPCs and protects against oxidative stress-induced cell death.
Materials and Methods
Cell isolation and culture
Cryogenically preserved NSPCs previously isolated from the central canal region of the spinal cord of transgenic adult female Wistar rats expressing green fluorescent protein were used in this study. The methods of isolation were previously described by our laboratory [3]. Cells were grown as free floating neurospheres in growth media containing neurobasal media (Gibco-Invitrogen), B27 neural supplement (Gibco-Invitrogen), 2 mM L-glutamine (Gibco-Invitrogen), 100 μg/mL penicillin–streptomycin (Gibco-Invitrogen), 20 ng/mL epidermal growth factor (EGF; Sigma), 20 ng/mL basic fibroblast growth factor (Sigma), and 2 μg/mL heparin (Sigma). NSPCs were passaged every 7 days and experiments were conducted on cells from passages 3–6. NSPCs (10,000 cells/well) were dissociated using Accutase (Gibco-Invitrogen) and seeded on Matrigel-coated (BD Biosciences, Inc.) 24-well plates containing growth media.
Treatments
Cultures were treated with ampakines CX546 (5, 50, 100 μM; Tocris) or CX614 (5, 10, 50 μM; Tocris) either alone or in the presence of glutamate (50 μM; Sigma). To induce oxidative stress, hydrogen peroxide (H2O2; Fisher Scientific) was added to each well to reach a final concentration of 500 μM.
Cell viability
Cell viability was assessed using live/dead staining (Invitrogen) as we have previously described [5]. Images were captured using a Nikon Eclipse TE 300 microscope. Five random images were taken for each well at 10× magnification and a minimum of three wells were counted per condition. Experiments were repeated a minimum of three times. The number of calcein+ cells was counted in each image to assess cell survival. The percentage of ethidium homodimer+ cells relative to total cells was calculated to determine percent cell death.
Immunocytochemistry
Cell differentiation and proliferation were assessed using immunocytochemical staining, as we have previously described. Cells were fixed with 4% paraformaldehyde for 20 min at room temperature and washed with 0.1 M phosphate-buffered saline (PBS). Cells were blocked with 10% normal goat serum with 0.3% Triton X-100 and 1.5% bovine serum albumin (depending on the primary antibody) for 1 h at room temperature. Afterward, cells were incubated with the primary antibody overnight at 4°C. The following primary antibodies were used: Ki67 (1:1,500; Abcam) for proliferating cells, nestin (1:100; BD Sciences) for NSPCs, GFAP (1:200; Millipore) for astrocytes, RIP (1:20; Developmental Studies Hybridoma Bank) for oligodendrocytes, and βIII tubulin (1:500; BioLegend) for neuronal progenitor cells. Cells were then washed with 0.1 M PBS, incubated with the fluorescent Alexa 568 secondary antibody (1:500; Invitrogen) for 1 h, and washed with PBS. To counterstain for cell nuclei, cells were incubated in Hoechst and washed with PBS. Immunocytochemical staining was imaged using a Nikon Eclipse TE 300 microscope. Ten random images were taken for each well at 20× magnification and the percentage of positive cells was quantified per image.
Statistics
All data are presented as mean ± standard deviation of the mean and were analyzed and graphed using SigmaStat and Prism software. Statistical differences between multiple groups were assessed using one-way or two-way analyses of variance and Bonferroni's or Dunnett's posthoc corrections. Differences between two groups were assessed using a two-tailed Student's t-test. A P value of <0.05 was considered statistically significant.
Results
AMPA receptor modulation increases adult spinal cord NSPC survival and proliferation
To examine the effect of AMPA receptor modulation on NSPCs, cells were treated with varying concentrations of ampakines CX614 or CX546 for 72 h. Cells were treated with each agent alone and in the presence of glutamate (50 μM). Given that there are basal levels of glutamate in the spinal cord, a concurrent treatment regimen would be most clinically relevant. There was a significant increase in the number of live cells following ampakine treatment in the presence of glutamate (Fig. 1A, B, E). The most robust response was seen at a concentration of 5 μM CX614 or 50 μM CX546. Moreover, these treatment regimens were associated with a significant reduction in cell death (Fig. 1C) and an increase in cell proliferation as measured by Ki67 staining (Fig. 1D). Higher concentrations of ampakines in the presence of glutamate led to a decrease in live cells.
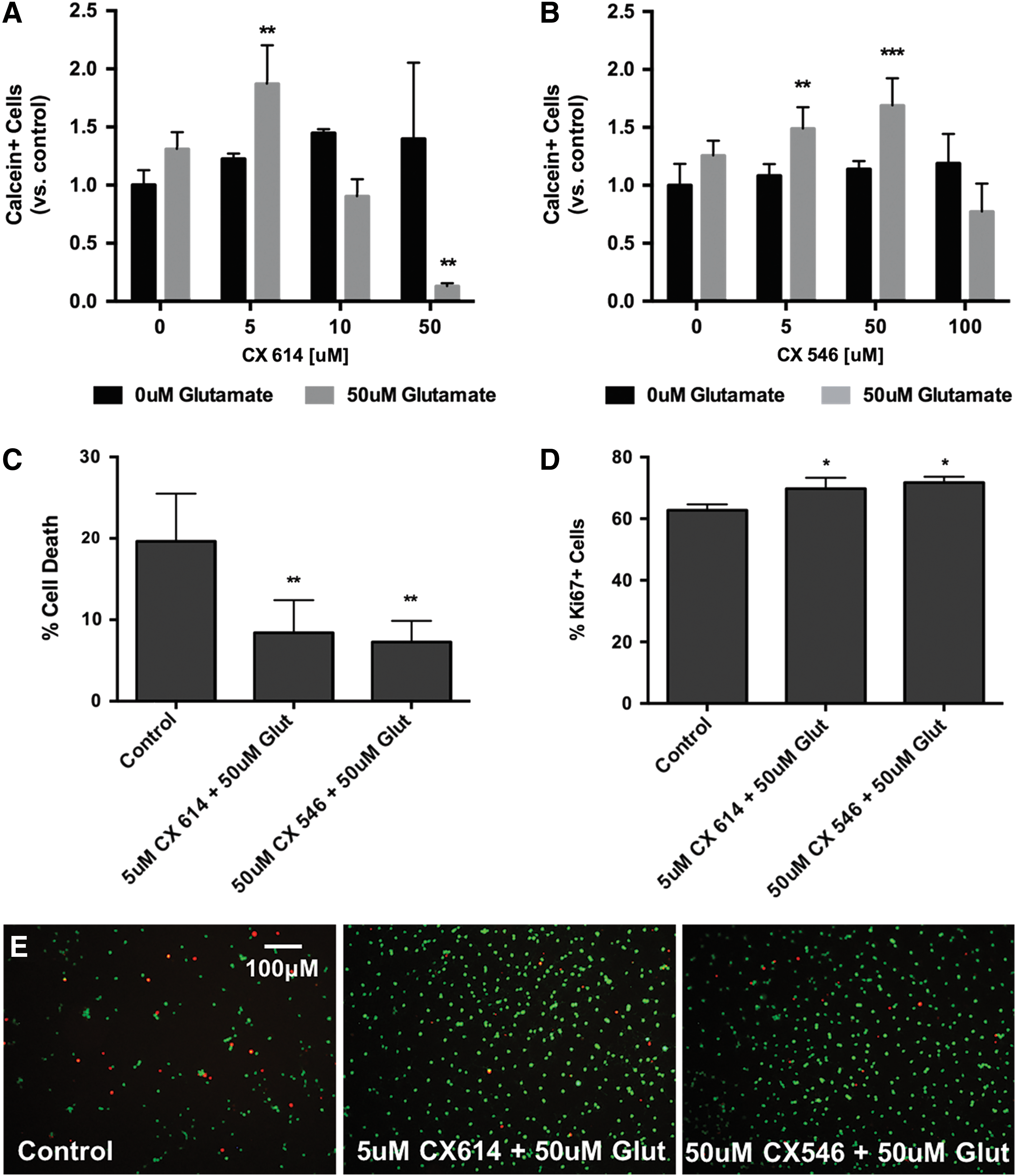
Effect of ampakines on adult rat spinal cord-derived NSPC survival and proliferation. Treatment with CX614 for 72 h led to a small but nonsignificant increase in live cells (calcein+) (black bars), however, this effect was further enhanced in the presence of low-dose glutamate (50 μM; gray bars)
Neuroprotection by modulation of AMPA receptors in the setting of oxidative stress
The effect of AMPA receptor modulation was further tested in the setting of oxidative stress. NSPCs were exposed to H2O2 (500 μM) for 6 h and cotreated with either CX546 or CX614 in the presence or absence of glutamate (50 μM). Treatment with either ampakine resulted in a significant increase in cell survival compared to controls exposed to H2O2 alone (Fig. 2). There was no difference in the percentage of Ki67+ cells between treated cells and controls under oxidative stress (control 58.6% ± 8.9%, 5 μM CX614 + 50 μM glutamate 64.2% ± 11.8%, 50 μM CX546 + 50 μM glutamate 62.3% ± 10.0%, P > 0.05; Fig. 2C), thus suggesting that the observed increase in live cells was due to enhanced survival and not proliferation.
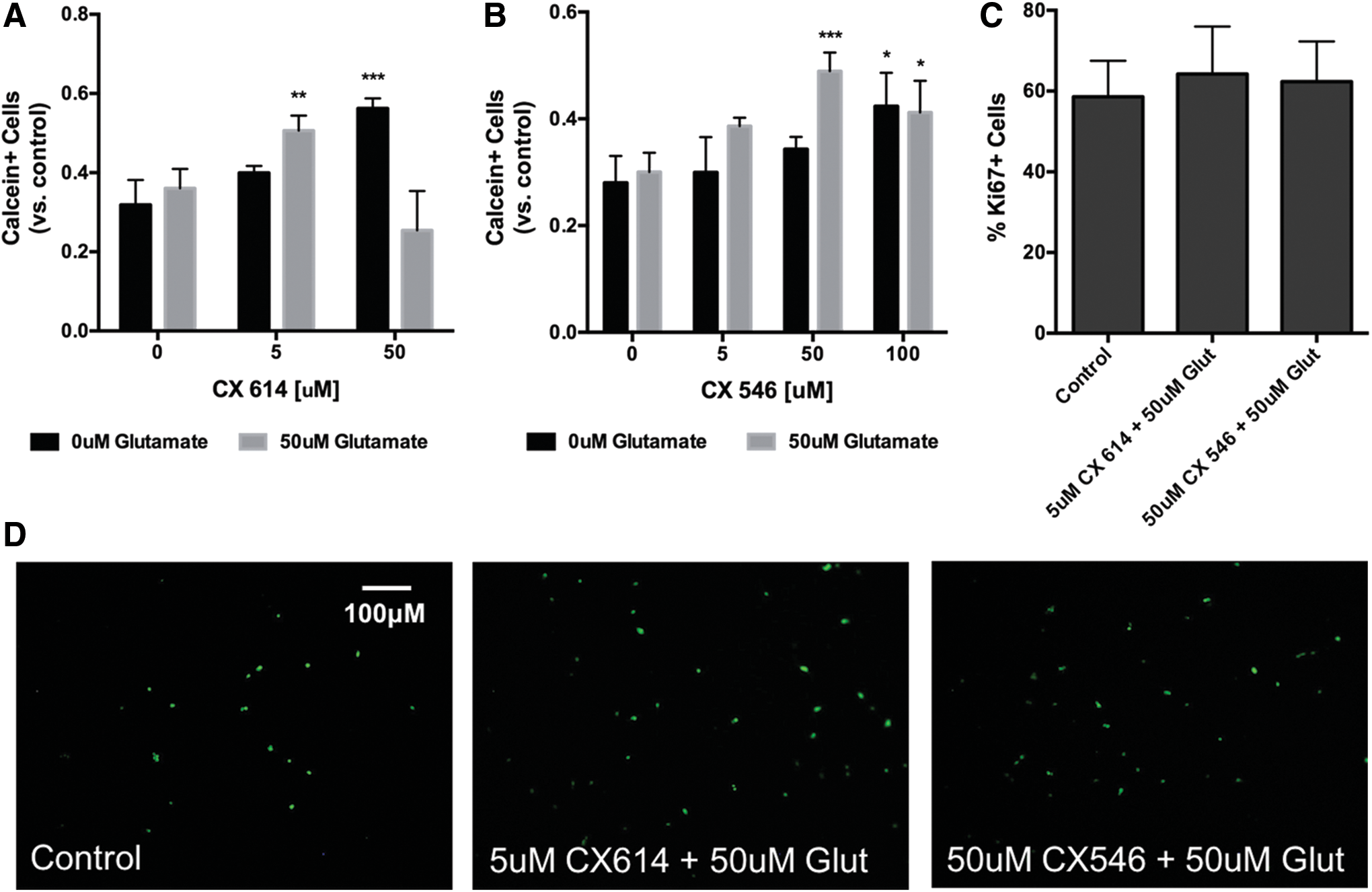
Effect of ampakines on adult rat spinal cord NSPCs under oxidative stress. NSPCs were exposed to 500 μM H2O2 for 6 h to induce oxidative stress. Treatment with CX614 either alone or in the presence of 50 μM glutamate led to increased cell survival in the presence of H2O2 compared to cells treated with H2O2 alone
Effect of AMPA receptor modulation on adult spinal cord NSPC phenotype
To determine whether ampakine treatment induced any changes in cellular phenotype, NSPCs were treated with CX614 (5 μM) or CX546 (50 μM) in the presence of glutamate (50 μM) for 6 days, after which immunostaining was performed (Fig. 3). The majority of cells were nestin positive, with low levels of neuronal progenitor (βIII tubulin), oligodendroglial (RIP), or astrocytic (GFAP) markers. While there was a similar percentage of βIII tubulin+ cells across treatments, cells treated with ampakine/glutamate had significantly longer neurites compared to controls (Fig. 3C). The percentage of RIP+ and GFAP+ cells was also similar across treatments, demonstrating that AMPA receptor modulation does not alter the phenotype of the NPSCs in vitro.

Phenotype of spinal cord NSPCs after 6 days of ampakine/glutamate treatment. Rat NSPCs were treated with control media (containing growth factors), control media +5 μM CX614 + 50 μM glutamate, or control media +50 μM CX546 + 50 μM glutamate for 6 days. The majority of cells in all treatment groups were nestin+
Discussion
Multipotent NSPCs that reside in the adult spinal cord have been the target of many therapeutic strategies for enhancing regeneration after SCI. Although the endogenous population of NSPCs initially proliferates after SCI, their numbers remain low, and regeneration is impeded by the acute toxic environment of the injured cord. Transplantation of exogenous NSPCs in the subacute period following injury has become a promising strategy to enhance regeneration but is limited by the poor survival of grafted cells. Strategies to better harness the potential of this population of adult stem cells remain an area of active research. In this study, we show that positive modulation of AMPA receptors using ampakines enhances survival and proliferation of adult spinal cord NSPCs in vitro and attenuates oxidative stress-induced cell death. These findings may have important therapeutic potential in enhancing survival of endogenous or transplanted NSPCs following SCI.
The therapeutic utility of ampakines has been investigated in the setting of a number of neurological conditions. Administration of ampakines has been shown to facilitate long-term potentiation, upregulate BDNF in the brain, and promote synaptic plasticity and learning in various rodent models [9]. Treatment with ampakine CX929 in a mouse model of Huntington's disease slowed the progression of striatal degeneration and motor dysfunction [7,8]. Moreover, administration of ampakines 5 days after injury in a rodent model of stroke was found to induce BDNF levels in the peri-infarct cortex leading to improved motor recovery [10]. Recently, treatment with ampakine CX546 (50 μM) for 48 h was shown to increase in vitro proliferation and neuronal differentiation of subventricular zone stem/progenitor cells derived from 1- to 3-day-old mice brains [11]. Here we present the first analysis on the potential use of ampakines to promote survival of NSPCs derived from the adult rat spinal cord.
Previously, microdialysis experiments in rodents have demonstrated that intrathecal levels of glutamate peak immediately after SCI reaching concentrations near 500 μM, which leads to extensive neuronal and glial cell death [12]. Recently, we showed that this same concentration of glutamate in vitro leads to increased survival and proliferation of spinal cord NSPCs and this is dependent on AMPA receptors [5]. It is thus possible that glutamate signaling via AMPA receptors may be a stimulus that facilitates the immediate proliferation of endogenous neural stem cells seen after SCI [13,14]. However, this acute period is characterized by a cascade of other toxic mediators that impedes sufficient regeneration. For this reason, transplantation of exogenous NSPCs is often delayed until the subacute period to avoid the early hostile environment of the injured spinal cord.
The initial rise in glutamate concentration after SCI has been shown to level off within a few hours of injury [12], however, there are no investigations examining the exact time course of intrathecal glutamate concentrations in the subacute or chronic periods of SCI. Previous studies conducted in a canine model of thoracolumbar SCI found that CSF glutamate concentrations reach 7.5 ± 7.2 μM in the acute period after injury and decrease to 4 ± 0.7 μM in the chronic period [15]. However, concentrations of glutamate have been shown to dramatically decrease even a few millimeters from the lesion epicenter, and thus, CSF estimates of glutamate likely underestimate intrathecal concentrations [12]. In this experiment, we chose a concentration of 50 μM glutamate to obtain a relatively low dose that may reflect intrathecal glutamate concentrations in the injured spinal cord in the subacute period. This concentration of glutamate did not have a significant effect on NSPC numbers alone, however, cotreatment with either CX614 or CX546 enhanced the effect of glutamate leading to increased cell survival. Although CX614 had a more potent effect on NSPCs compared with CX546, it was also associated with a narrower therapeutic range and thus careful dosing calibration would be required when translating to an in vivo setting.
Clarkson et al. found that delayed treatment with ampakines 5 days after stroke in mice led to enhanced recovery of limb control, and this effect was mediated by BDNF [10,16]. Conversely, AMPA receptor blockade impaired recovery. Importantly, administration of ampakines at the time of stroke led to an increase in infarct volume, therefore suggesting an inflection point in the role of AMPA receptor modulation after stroke. Similarly, AMPA receptor modulation may be a potential strategy in the subacute or chronic phases after SCI to increase neural stem cell numbers and enhance regeneration when low levels of glutamate are present. This may be particularly useful as exogenous NSPCs are often transplanted at 1 week after injury. The ampakine strategy may also be applicable to improving the fate of endogenous neural stem cells after SCI. The implications and therapeutic utility of this strategy in the setting of SCI thus warrant further study.
The exact downstream mechanism underlying the positive effect of ampakines on spinal cord NSPCs warrants further investigation. Ampakines have been shown to increase BDNF levels in the brain and enhance neuronal differentiation of neural stem cells [6,11,17]. Previously, we have demonstrated that NSPCs produce detectable levels of BDNF [18]. Moreover, we have shown that treatment of NSPCs with BDNF is protective against oxidative stress [19]. It is possible that ampakines induce BDNF production by NSPCs, which act in an autocrine manner to enhance survival. Moreover, BDNF is known to induce neuronal differentiation of NSPCs [20] and treatment with ampakine CX546 has previously been shown to induce neuronal differentiation of neonatal mouse brain stem cells [11]. Although we did not find a significant increase in the percentage of βIII tubulin+ cells under ampakine/glutamate treatment, there was a marked difference in cell morphology as βIII tubulin+ cells in the treatment groups had significantly longer neurites compared with control cells. The lack of a quantitative difference in βIII tubulin+ staining may be due to the growth factor containing media used in the present study. Culturing cells in media without growth factors in the presence of ampakines/glutamate may have led to a more robust increase in neurons as was previously demonstrated in brain-derived neural stem cells [11]. Further study into the mechanisms of the observed effects is therefore necessary.
Conclusions
We present the first examination of the effect of allosteric AMPA receptor modulators on adult spinal cord-derived NSPCs. We show that ampakines CX546 and CX614 promote the survival and proliferation of NSPCs and attenuate oxidative stress-induced cell death. These findings offer important insight into a potential therapeutic strategy to positively regulate adult spinal cord-derived NSPCs after SCI.
Footnotes
Acknowledgments
We thank Linda Lee and Rita van Bendegem for their technical assistance. We also thank the Krembil Foundation, Toronto General and Western Hospital Foundation, and Spinal Cord Injury Ontario for support.
Author Disclosure Statement
No competing financial interests exist.