
Abstract
Elucidation of the spaceflight (SF) effects on the adult stem and progenitor cells is an important goal in space biology and medicine. A unique opportunity for this was provided by project “BION-M1”. The purpose of this study was to evaluate the effects of 30-day SF on biosatellite, 7-day recovery (SFR), and subsequent ground control (GC) experiment on the mononuclear cells (MNCs) from C57/BI/6N murine tibia bone marrow. Also, hematopoietic and stromal precursor functions were characterized ex vivo. There was no significant difference in the total MNC number between experimental groups. After SF, immunophenotyping revealed an increase of large-sized CD45+MNCs corresponded to committed hematopoietic progenitors. The total hematopoietic colony-forming unit (CFU) number decreased after SF and did not restore after 7 day of recovery due to predominant reduction of bi- and multipotent CFUs and primitive burst-forming units in favor of unipotent CFUs. Functional activity of stromal precursors in vitro was only slightly altered. SF cells displayed the enhanced expression of alkaline phosphatase. The data of the GC experiment demonstrated the preservation of the functional activity of progenitor cells from mice bone marrow. The activation of erythropoiesis in expense of burst-forming units of erythrocytes elevation was detected. After 7 days of recovery, the number of colony-forming units of fibroblast (CFUs-f) was similar to the vivarium control, while the proliferative activity of bone marrow stromal precursors decreased. The present study demonstrated that certain hematopoietic progenitors are susceptible to SF factors, while the stromal precursors displayed a certain degree of resistance. These data indicate mild and reversible alterations of bone marrow progenitors after SF.
Introduction
S
A number of serious functional and structural changes in different tissues, including hematopoietic, have been revealed in rodent experiments carried out in the frame of BION program (1973–1997) on “Cosmos” biosatellites and Spacelab 1–3 [1 –3]. Hematopoietic tissue of the bone marrow represents a depot of stem and progenitor cells of hematopoietic and stromal lineages, which are the most susceptible to SF factors among the other tissues. The adaptation to the adverse effects of SF and the subsequent return to normal gravity are largely dependent on bone marrow state. After biosatellite experiments, a decrease in hematopoietic precursor number in bone marrow, attenuation of lymphopoiesis, enhancement of granulocyto- and monocytopoiesis, and inhibition of erythropoiesis in the bone marrow and spleen with simultaneous decrease in reticulocyte number in the peripheral blood was detected [3 –5].
Studies of the stromal precursors' compartment, now called multipotent mesenchymal stromal cells (MMSCs), in biosatellite experiments are still few. A reduction in the number of osteoblasts, while maintaining the number of osteoclasts, and inhibition of periosteal remodeling in long bones were revealed in a BION project [6]. Furthermore, the reduction in the number of bone marrow-derived stromal progenitor cells (colony-forming unit of fibroblast [CFU-f]) in rats after the flight had been demonstrated earlier [4,6,7]. More recent studies onboard the ISS (Mission STS-131, NASA, 2010) have shown a delay of the osteoblast cell cycle and increased osteoclast-mediated osteolysis under microgravity as well as downregulation of mesenchymal cell marker gene expression [8,9]. However, many aspects of bone marrow low-committed stromal precursor adaptation to SF factors are still undercovered.
The goal of the C57BL/6N mice experiments in the BION-M1 project was to elucidate cellular and molecular mechanisms underlying the adaptation of key physiological systems to long-term exposure in microgravity [10]. In the present article, we examine the total mononuclear population of the murine tibia bone marrow and characterized the morphology and functions of stromal and hematopoietic precursors shortly after prolonged 30-day SF on a biosatellite, 7-day recovery, and a subsequent ground control (GC) experiment.
Materials and Methods
Ethical statement
The permission for experiments was obtained from the Biomedical Ethics Commission of IBMP (No-319, April 4, 2013). Experiments were organized in compliance with the principal bioethical requirements and European Convention for the Protection of Vertebrate Animals used for Experimental and Other Scientific Purposes [11].
Experimental groups
The launch of “BION-M1” was implemented in April 2013. Male C57BL/6N mice weighing 22–25 g were purchased from the Animal Breeding Facility–Branch of Shemyakin and Ovchinnikov Institute of Bioorganic Chemistry. The following experimental groups were formed (Fig. 1A): SF–30 days of flight on biosatellite “BION-M1”; control for the SF animals (vivarium control to SF [SFV]); recovery after the flight (SFR); control for recovery after the flight (vivarium control-recovery [VCR]); subsequent GC experiment; vivarium control to GC (GCV) [10]; the recovery after the GC (GCR) experiment; and control for recovery from the GC experiment (GCRV) [12]. In this study, we have described the data from two experiments in frame of “BION-M1” Project: the biosatellite and the ground-based experiment (Fig. 1B). The purpose of ground-based experiment was to evaluate the effects of flight habitat parameters other than SF factors (microgravity, hypergravity, radiation). Due to this goal, the ground-based experiment was performed after SF in the biosatellite animal boxes. Accordingly, two different animal pools were used, which did not allow us to consolidate all control groups. Meanwhile, there were no significant differences (U test) between the data of the control groups inside of flight and ground-based experiments, accordingly. This was a rationale to join the control groups inside of certain experiment (Fig. 1B).

Study design.
Study design
The animals were euthanized by cervical dislocation and tibias were dissected for further studies. Bone marrow was isolated from the tibia by aspiration and the suspension of bone marrow mononuclear cells (MNCs) were isolated as described below. Cell suspension was pooled and aliquoted for further experiments. A part of samples was used immediately for total nucleated cell counting and immunophenotyping. Another part of aliquots was frozen and further used for stromal and hematopoietic cell characterization in primary cultures.
Isolation of bone marrow MNCs
MNCs were isolated from the tibia bone marrow of C57BL/6N mice as described elsewhere [13]. MNCs were counted after lysis of erythrocytes with OptiLyse C buffer (BD) according to manufacturer's instructions. In brief, a lysis buffer was added to the cell suspension in a ratio of 10:1 and incubated for 15 min at room temperature. After lysis of erythrocytes, MNCs were placed in a hemocytometer, counted, pooled, and aliquoted for further experiments. MNC viability was evaluated with trypan blue exclusion test [12].
Primary culture of stromal progenitors
MNCs were seeded in 35 mm Petri dishes in full culture medium a-MEM (Gibco), 2 mM glutamine, 20% FCS (Hyclone), 1% antibiotic/antimicotic (PanEco, Russia). The seeding density and time-in-culture was chosen according to the experimental design. The culture medium was changed every 3 days. For estimation of cell proliferation, MNCs were seeded as 12 × 104/cm2 and expanded for 14 days. Simultaneously with initiation of the culture, five view fields were randomly selected and marked, and further images were captured on day 3, 7, and 10 using a Nikon Eclipse TiU microscope (Nikon, Japan) equipped with a digital camera. For CFU-f evaluation, MNCs were seeded as 4 × 104/cm2, 12 × 104/cm2, and 36 × 104/cm2, expanded for 14 days, and stained with 0.5% crystal violet in methanol for 5 min. For evaluation of differentiation, MNCs were plated as 22 × 105/cm2. After 7 days in culture, alkaline phosphatase (ALP) activity was detected in cells with a Leucocyte ALP kit (Sigma) to demonstrate spontaneous osteocommitment. Spontaneous adipodifferentiation was detected after 7 days in the culture with Oil Red O staining [12].
Hematopoietic CFU assay
MNCs (50 × 103 cells/mL) were cultured in a methylcellulose-based medium MethoCult GF M3434 (StemCells) according to manufacturer's protocol. After 14 days, the number of CFU-tot and CFUs of certain hematopoietic lineages was calculated.
Immunophenotyping of bone marrow MNCs
An aliquot of cells from each pool was stained with monoclonal phycoerythrin (PE)-conjugated antibodies against CD45, CD34, CD90.2 (Thy1) antigens (Biolegend). As a control, nonmatched PE-IgG of the same subclass were used (Biolegend). Cells were washed with a phosphate-buffer saline (PBS) by centrifugation (5 min, 1500 rpm), resuspended in PBS, and counted and aliquoted in 100 μL with 1–3 × 105 cells per probe. Antigen-specific or isotype antibody (1 μl) was added to each tube, vortexed, and incubated for 15 min at +4°C in the dark. Then, 400 μL of PBS was added into each sample. Analysis of cell fluorescence was carried out with flow cytometer Epics XL, using System II software, to capture and analyze histograms (Beckman Coulter). The proportion of CD45+, CD34+, and CD90+ MNCs was determined [14].
Immunophenotyping of cells in primary cultures was performed as follows: MNCs were grown on coverslips according to the protocol, described above. Cells were fixed with 4% formaldehyde in PBS for 15 min at RT; then, cells were washed with PBS three times and incubated with the PE-conjugated anti CD45, CD34, and CD90.2 (Thy1) antibody (Biolegend) for 1 h at 4°C. Nonmatched PE-IgG were used as a negative control. Then, the samples were washed and mounted in Fluoroshield with DAPI (Sigma-Aldrich). Images were acquired using an LSM 780 (Carl Zeiss, Oberkochen, Germany) confocal microscope.
Morphometry and statistical analysis
Cell cultures were examined with a Nikon Eclipse TiU microscope (Nikon, Japan). Confocal Images were analyzed with NIS Elements 32 (Nikon) or Sigma Scan Pro5 (SPSS Corp.) software. Statistics were performed with the “Statistica 7.0” software package using a nonparametric Mann–Whitney test. Differences were considered significant at p < 0.05.
Results
Bone marrow MNCs
Cellularity
In the flight experiment, the number of MNCs isolated from the single bone varied significantly (from 14 to 32 × 106) within certain groups of mice. There was no significant difference in the total cell number between experimental groups. Meanwhile, a decrease in cell number as a tendency was observed in SF and SFR in comparison with SFV [14] (Fig. 2A).
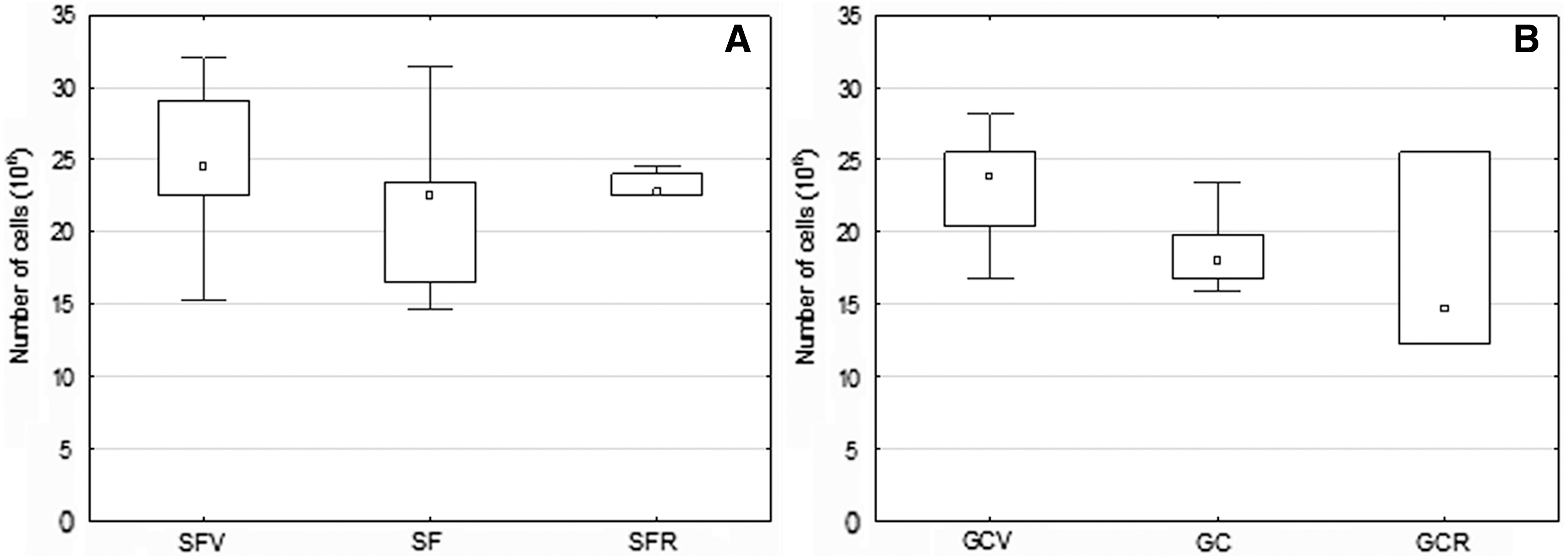
An average number of nucleated cells in the tibia of mice bone marrow.
Subsequent GC experiment revealed a significant decrease in total cell number in the GC versus GCV group (Fig. 2B). The same decline was observed in the GCR group, but was not significant due to high data dispersion.
Viability of MNCs was high and reached up to 95%–100% (Trypan blue exclusion test). Flow cytometry analysis revealed the heterogeneity of MNCs in forward/side scattering (FSC/SSC) density plots, which is typical for bone marrow cells. Total MNCs were divided into two subpopulations according to cell size (Gate 1 and Gate 2, respectively) (Fig. 3A). Small-sized MNCs were in Gate 1, and on average, two times larger MNCs were in Gate 2 [14]. From the data on MNC number in Gate 1 and 2, we calculated the ratio of MNCs of different size. In both control groups (SFV and GCV), MNCs were distributed almost equally between the gates. In the SF and SFR groups, the number of MNCs in Gate 2 was significantly higher than in the Gate 1 (the increase amounted to 20% and 30%, respectively) (Fig. 3B). From the physiological point of view, small-sized cells represent lymphocytes, and larger cells are leukocytes and hematopoietic stem progenitor cells (HSPCs) of different commitment. An increase in the number of large-sized MNCs after SF may indicate the alteration of lymphocyte/leukocytes ratio. As we did not find any significant difference in the total cell number of MNCs between the experimental groups, we have assumed that the increase of proportion of large-sized cells may be considered as an elevation of total number of larger MNCs represented by leucocytes and HSPCs.
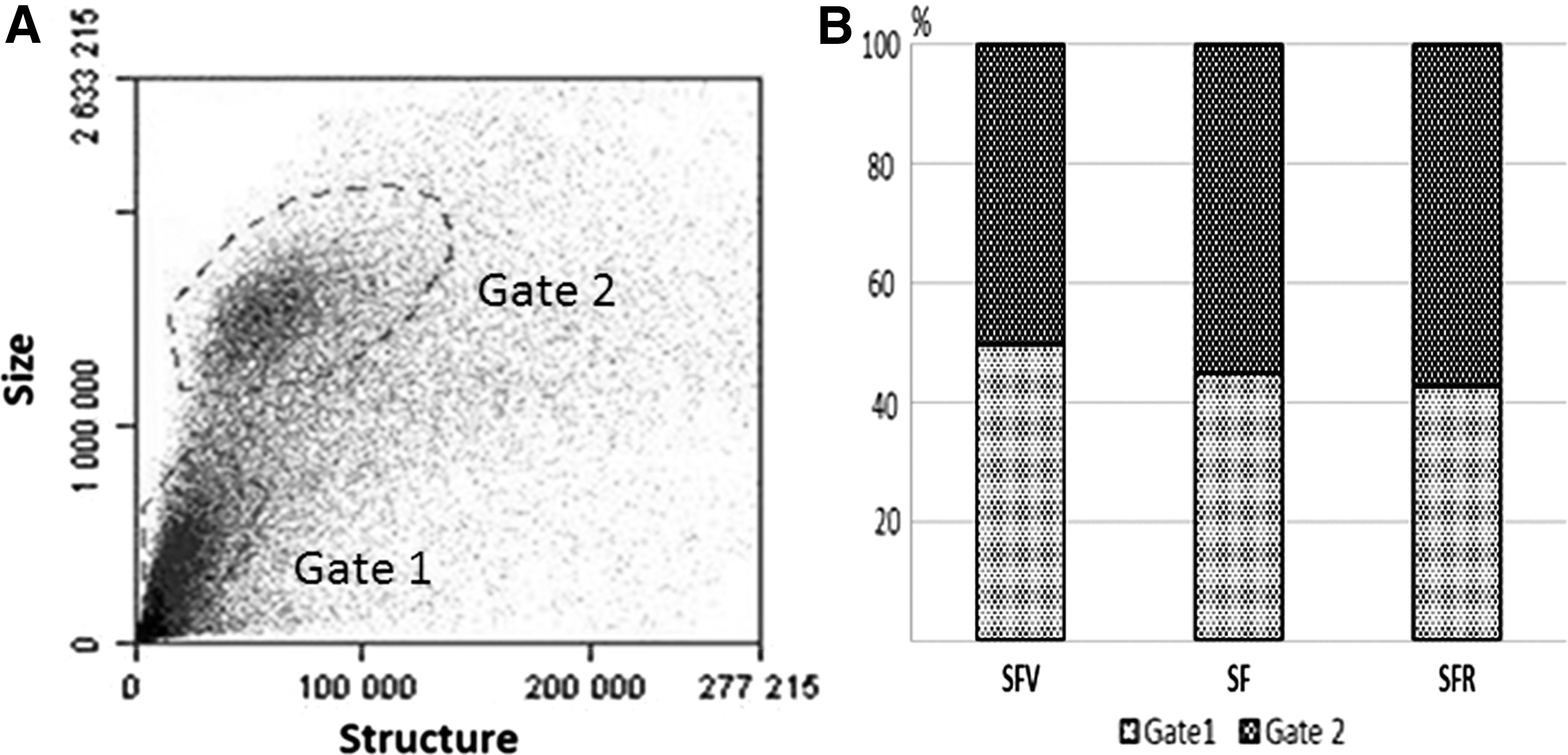
Size/structure distribution of MNCs from tibia bone marrow of C57BL/6N mice.
MNCs in primary culture
The bone marrow MNCs included several cell types. In cell cultures, they represent a quite heterogeneous population because of the tight contacts between stromal and hematopoietic cells; the ratio of those is dynamically changed along culture duration [15]. To identify stromal and blood-borne cells more precisely, CD45, CD34 and CD90.2 positive cells were detected immunocytochemically after 14 days of primary culture. CD45-positive cells were represented as small-sized cells with low nuclear-cytoplasmic ratio (Fig. 4). A part of CD45-positive cells bore the CD34 antigen as well. The CD90.2 antigen was expressed on the medium- and large-sized fibroblast-like cells with processes, which had a large nucleus with 2–3 nucleoli and a high nuclear–cytoplasmic ratio (Fig. 4). CD45-positive cells were clustered on the surface of large stromal cells named “nursing” cells.
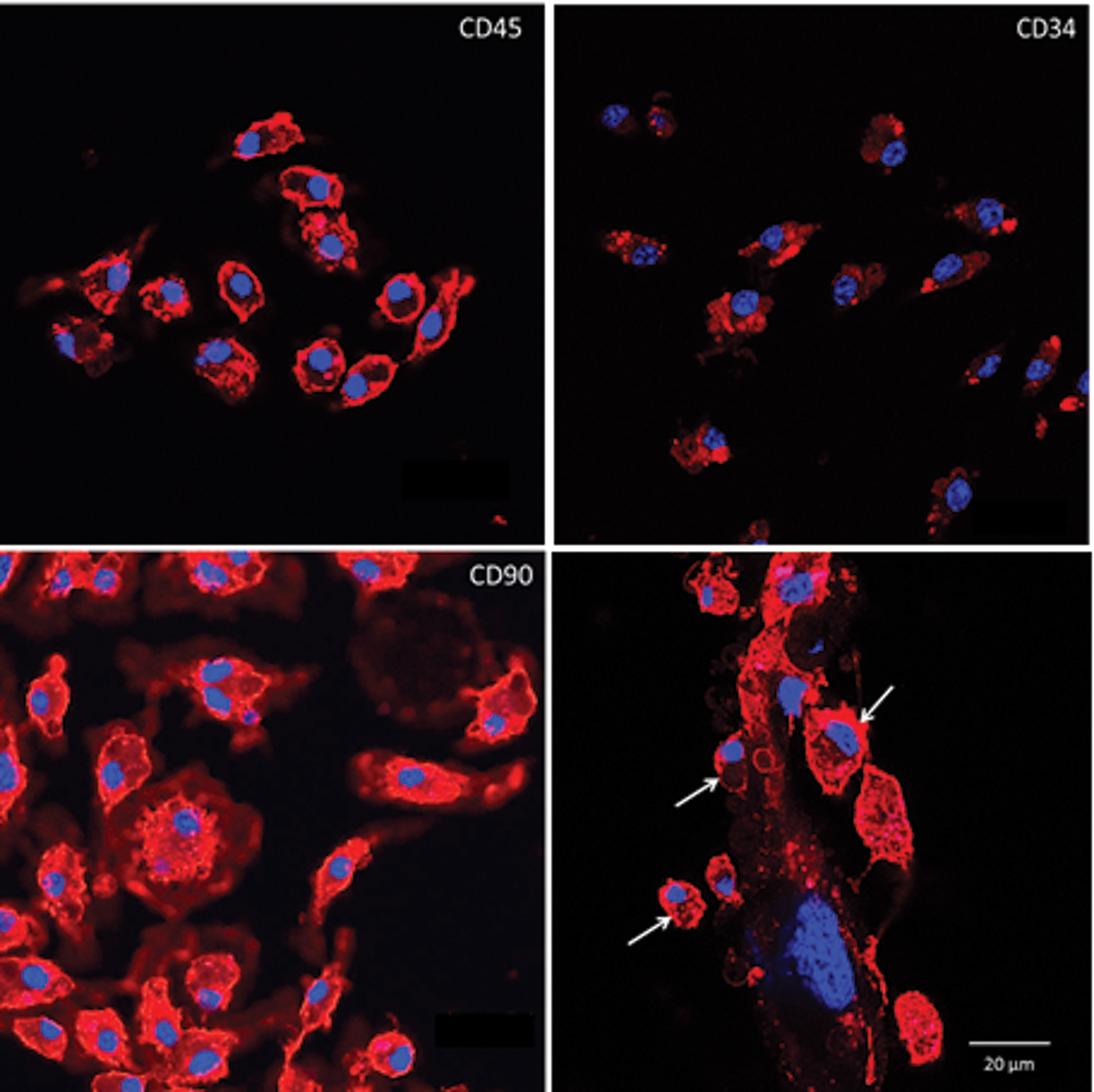
Immunophenotype of bone marrow MNCs in primary culture. Upper left: cells expressing pan-leukocyte marker CD45. Upper right: cells bearing CD34—adhesion molecule showing expression on early hematopoietic cells. Lower left: cells expressing CD90 marker of early stromal progenitors. Lower right: CD45-positive cells (white arrows) on the surface of large stromal cell. Confocal microscopy. Scale bar, 20 μm. In collaboration with Prof. S.V. Buravkov, FBM MSU, Moscow. Color images available online at
After 10 days in the culture, the SFV population was heterogeneous, including fibroblast-like stromal and hematopoietic-like precursors (Fig. 5A). The stromal cell population comprised cells of different sizes (small, medium, large) as well as a number of nursing cells (Fig. 5B). In the SF population, the same cell types were represented (Fig. 5A), although the share of nursing cell was substantially decreased (Fig. 5B). Small-sized hematopoietic-like cells were practically absent (Fig. 5A). After 7 days of recovery, the morphology of SFR cultures was differed from SFV and SF groups (Fig. 5A). The decrease of the number of small and large cells in expense of medium-sized cells were observed, the number of nursing cells was partly restored versus SF, but did not reach the SVF group (Fig. 5B). Ground-based experimental conditions did not influence the composition of cultured bone marrow cell populations (Fig. 5A).
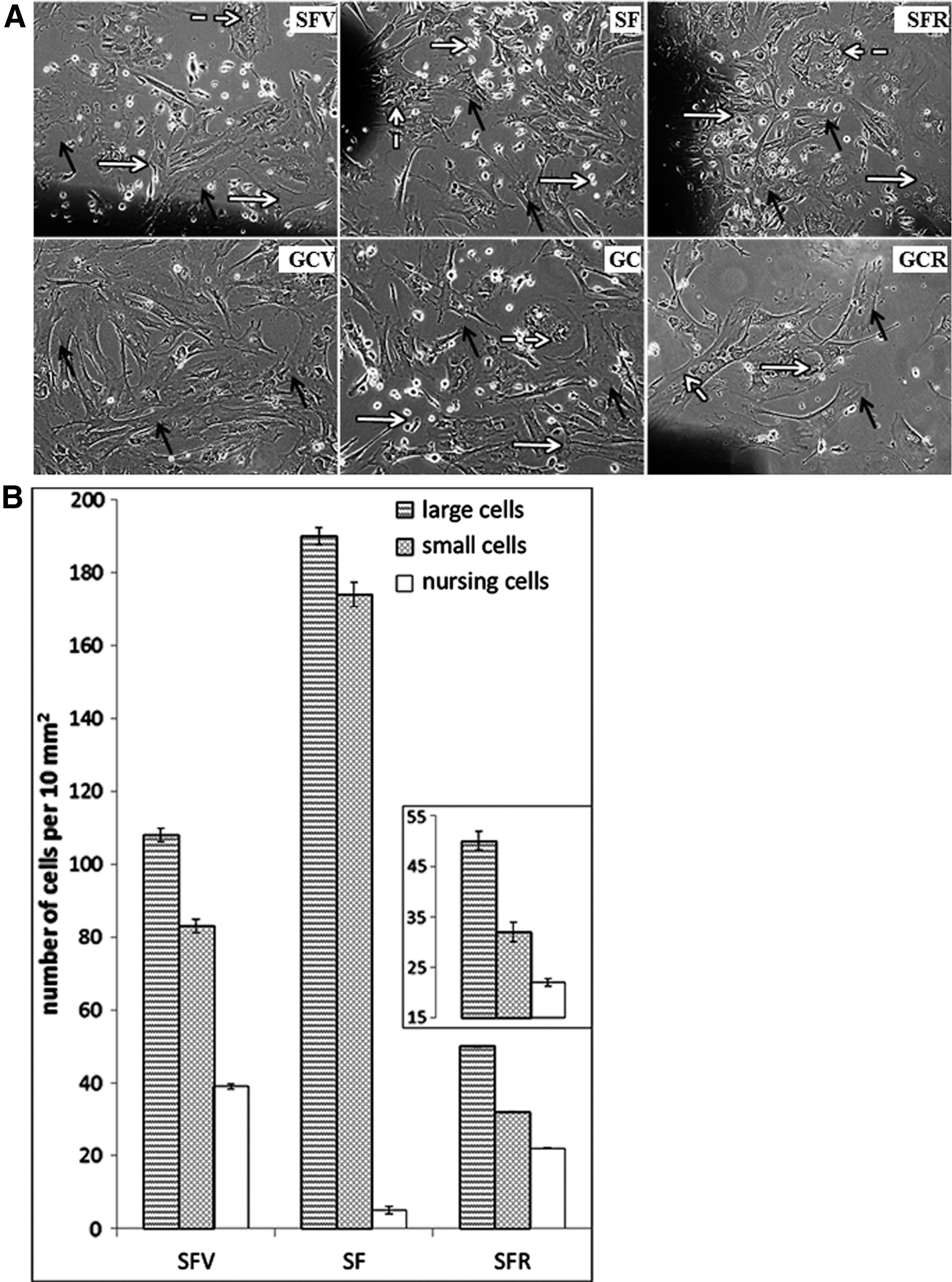
Bone marrow MNC population heterogeneity in primary culture.
The presented data demonstrated that the cellular composition of bone marrow was significantly affected in SF. The number of hematopoietic cells was reduced drastically; that provoked the decrease in the quantity of cellular associations.
To characterize cell growth along propagation, population doublings (PDs) in each experimental group were calculated. We did not find any significant differences in PDs either in the SF or GC experiments (it was in range from 2.3 to 3.2).
Thus, SF factors did not affect growth rate of bone marrow-derived MNC in vitro. Herewith, the number of adhered cells was higher in the SF group versus SFV, while the number of adhered cells in the SFR and GCR was lower in comparison with appropriate controls.
MMSCs in primary culture
Colony-forming units of fibroblasts
Ability to raise a colony from a single cell in vitro is the most distinguished feature of bone marrow low-differentiated stromal precursors, aka MMSCs [16]. To detect MMSCs among the MNC population, we evaluated the CFUs of stromal cells (CFU-f). MNCs were seeded in a primary culture with density usually applied for detecting of CFU-f (4 × 104 cells/mm2) [15] and with two higher densities (12 and 36 × 104). After 14 days in the culture, CFUs-f were detected at all cell densities. The number of colonies did not differ in plates with 4 × 104 density in all experimental groups and was about 300–500 CFU-f per one tibia bone marrow cell population. Only in the GCR group, the number of CFU-f was significantly higher in comparison to GCV (Fig. 6A). With increase in seeding density in 3 (12 × 104) and 9 (36 × 104) folds, the number of CFUs-f multiplied linearly; this confirmed the clonal nature of these colonies (data not shown).
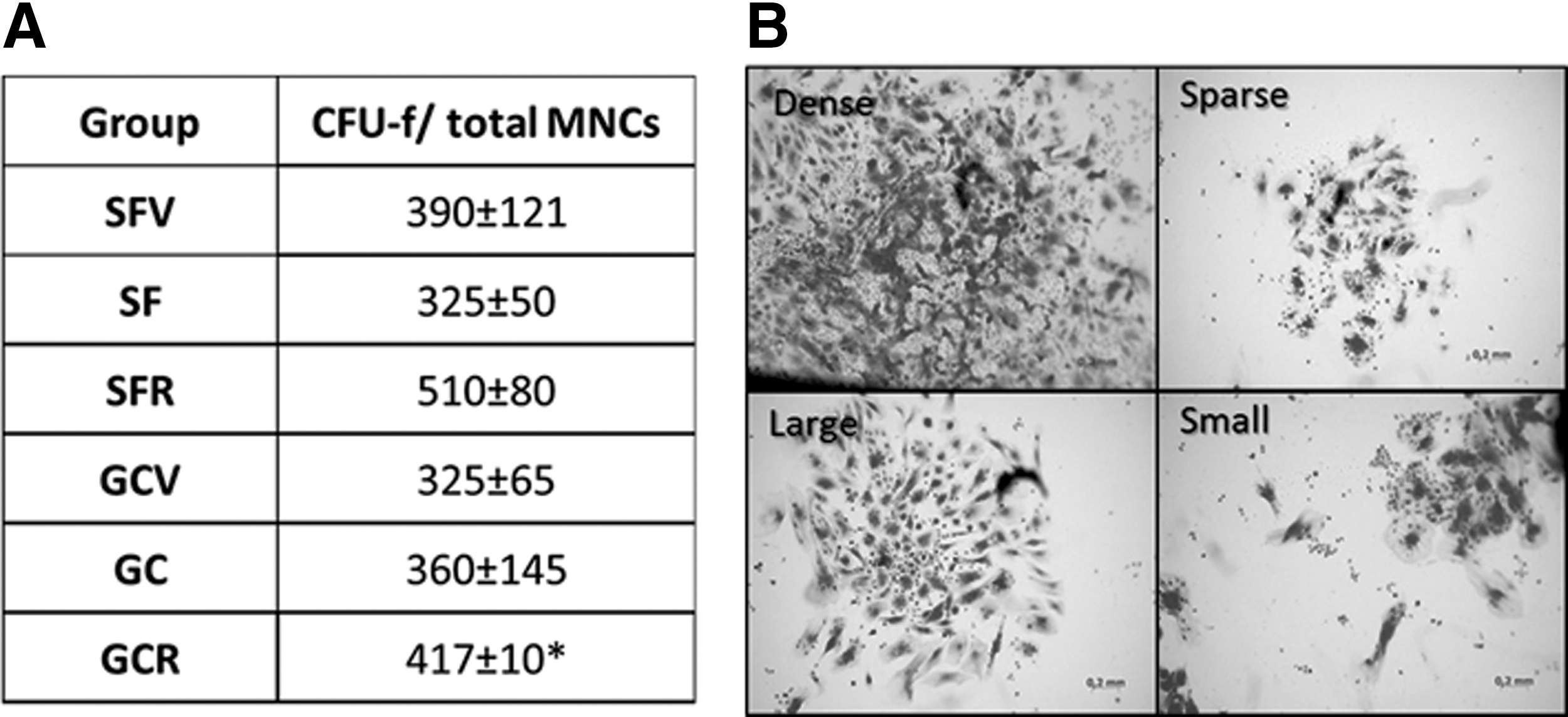
CFU-f evaluation among bone marrow MNCs.
CFUs-f varied in size and morphology (Fig. 6B). There were four types of colonies: small, less than 3 mm in diameter; large, more than 3 mm; sparse, weakly stained with crystal violet, each single cell was well distinguishable; and dense, strongly stained, single cell was not seen. The size of colonies is a characteristic of the proliferation capacity of clone-forming cells; actively proliferating clones appear to be dense, while slow-growing clones are sparse [17]. The dense colonies dominated in SF and SFV, while most of colonies were sparse in SFR group. It is noteworthy that in ground-based experiment, the CFUs-f displayed similar sparse morphology in all groups, indicating similar proliferating activity.
Spontaneous differentiation
It is well demonstrated that in vitro MMSCs exhibit at least three-lineage potential differentiating into osteo-, adipo-, and chondrodirection. The SF simulation provoked intensification of adipo- and attenuation of osteocommitment, as was demonstrated in our laboratory earlier [18].
In this study, we examined spontaneous MSC osteogenic commitment after 7 days in culture. The activity of ALP—an early marker of osteodifferentiation—was detected histochemically in cultured MMSCs (Fig. 7). The cells with different intensity of staining were detected in the same culture (Fig. 7A, C). The most homogenous staining was revealed in the SF group. Nevertheless, the mean staining intensity did not differ between experimental groups. Herewith, the total area occupied with stained cells in SF group was 1.5-fold higher than that in SFV (Fig. 7B), while in the SFR group, the area of stained cell was similar to that in SFV. The same effect was observed in the GC experiment (Fig. 7D).
The efficacy of spontaneous osteodifferentiation in cultured MSCs from mice bone marrow after SF and GC conditions.
Spontaneous adipodifferentiation of MMSCs was detected with Oil Red O after 7 days in culture. The intracellular lipids are described as characteristic features of mice bone marrow cells in culture [19]. In all experimental and control groups, the positively stained lipid vacuoles were found in fibroblast-like cells of different sizes.
Hematopoietic CFUs
Recently, we published the data of the preliminary analysis of relative CFU number per 50 × 103 mice bone marrow MNCs after the “BION-M1” flight and GC experiments, demonstrating a significant decrease in the number of erythroid progenitors, including the common myeloid precursor—CFU-GEMM—after SF [20]. Here, we calculated the absolute number of CFUs per bone marrow and the ratio of uni- and bi-/multipotent CFUs after 14 days of MNC expansion in a semisolid MethoCult medium. The number of CFU-tot among MNCs from SF was lower than in SFV (Fig. 8A). The share of unipotent CFUs was similar in the SF and SFV groups and comprised about 70% of total CFUs. After 7 days of recovery, the number of CFU-tot in the SFR group was close to SF—still lower than in SFV, indicating incompleted restoration of hematopoiesis (Fig. 8A). The significant twofold decrease in bi-/multipotent CFUs was noted in SFR versus SFV and SF groups, indicating the attenuation of multipotent recovery (Fig. 8B). The most prominent alteration was detected among primitive erythroid precursors-burst-forming units of erythrocytes (BFU-E). In comparison to SFV, sixfold reduction of BFU-E in SF was detected, which was not fully recovered after 7 days (Fig. 8A). These data demonstrated that SF factors affected CFUs number of hematopoietic stem cells, mainly due to the suppression of more primitive bi- and multipotent progenitors (Fig. 8B).
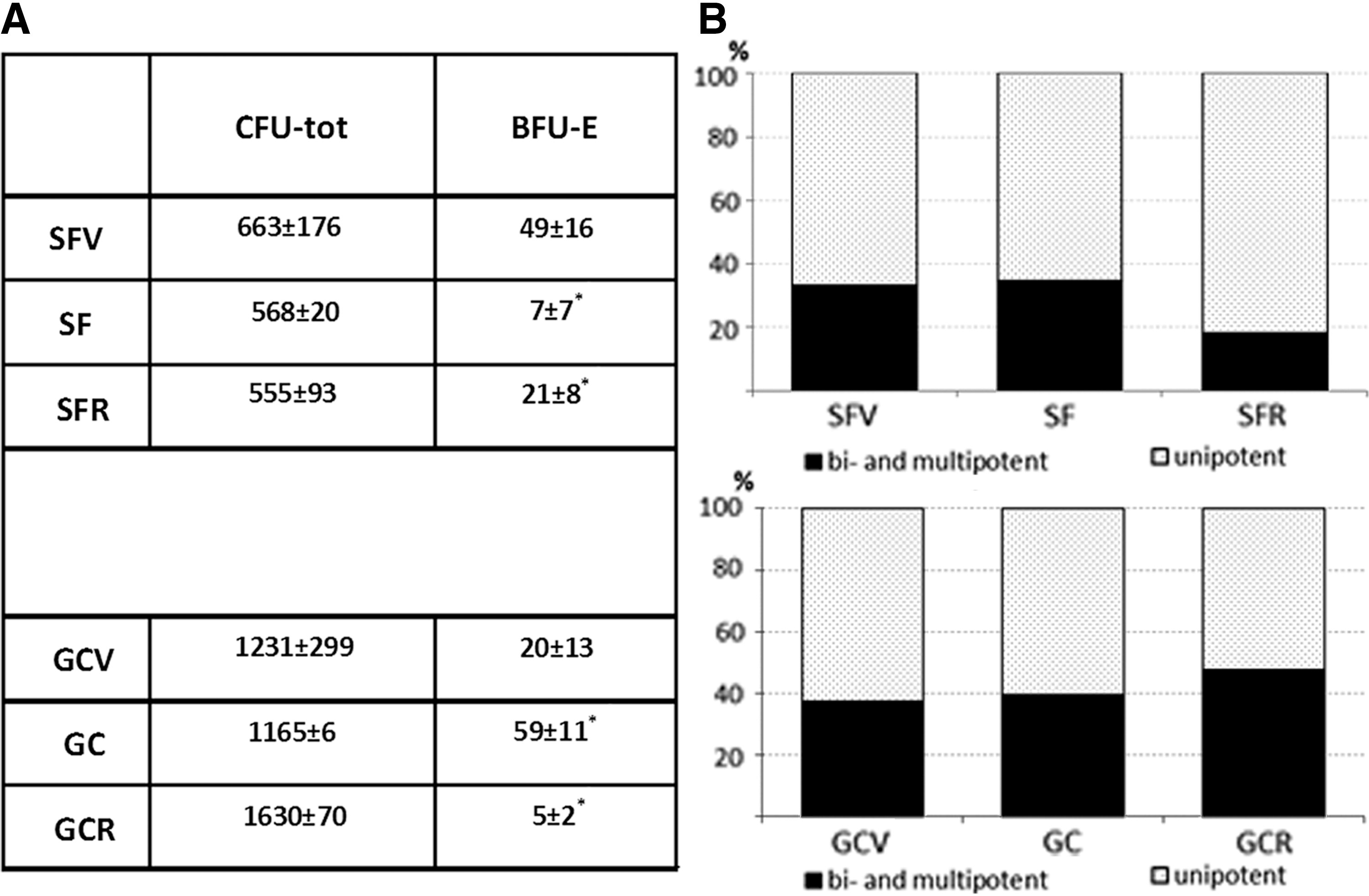
The hematopoietic CFUs among total MNC population from mice bone marrow after SF and GC conditions.
In the GC experiment, CFU-tot was slightly lowered in GC, but was significantly elevated in GCR in comparison with GCV (Fig. 8A). The number of BFU-E was significantly increased in GC both versus the GCV and GCR groups. The share of uni- and multipotent CUFs was not affected in GC experiment (Fig. 8B).
Discussion
In the present article, we have evaluated cellularity, viability, and phenotype of mononuclear (MNC) population from C57BL/6N mice tibia and provided in vitro characterization of MNC hematopoietic and stromal stem/progenitors after a 30-day “BION-M1” space flight, 7-day recovery, and ground-based control experiment.
Analysis of functional activity in vitro did not reveal significant negative consequences of SF on clonogenic, proliferative, and differentiating activity of stromal precursors of mice bone marrow. Meanwhile, the total hematopoietic CFU number decreased after SF and did not restore after 7 days of recovery. This reduction affected mainly bi- and multipotent CFUs and primitive BFUs.
The data of the GC experiment demonstrated the preservation of the functional activity of progenitor cells from the mice bone marrow. The activation of erythropoiesis as specific reaction of hematopoietic compartment was detected. After 7 days of recovery, the number of CFUs corresponded to the vivarium control, while the efficiency of attachment and proliferative activity of bone marrow stromal precursors decreased.
It is generally assumed that the influence of SF factors on stem and progenitor cells in the bone marrow could be a background to bone and hematopoietic tissue alterations. The analysis of Wistar rat bone marrow after 7–22 day's flights on “Cosmos” and “BION” biosatellites revealed the decrease of the hematopoietic progenitor number, while the total cellularity and mitotic activity of bone marrow cells were maintained at the control level. There were no detectable alterations in the bone marrow composition; most of cells were mature neutrophils with few premature progenies such as myeloblasts and promyelocytes. Erythroid and lymphoid precursors in the bone marrow were presented as rare groups of a few cells, indicating the inhibition of these lineages, which was accompanied by the decrease in the number of reticulocyte in peripheral blood. The enhancement of granulocytopoiesis occurred. This was evidenced from the increase of the granulocytes/early precursors ratio in the myelograms and the number of the leukocytes and neutrophils in peripheral blood. The physiological regeneration of hematopoiesis was slowed after SF [1 –7].
Evaluation of CFUs in a spleen assay revealed a significant (20 times) reduction in the number of CFUs among bone marrow MNCs. Erythroid differentiation of these progenitors was significantly inhibited, while granulocytic, on the contrary, was strengthened. These results are in agreement with the observations in bone marrow. Based on the above data, it was assumed that 7–22 day's flights negatively affected rat bone marrow hematopoietic compartment. The detected changes were mainly addressed to mature and premature cells. These alterations were reversible and fully compensated after 25–27 days of recovery [2,21].
The results on Sprague–Dawley rats in the frame of SLS-2 experiments were consistent with the earlier data cited above. After 13–14 days of microgravity, the signs of the inhibition of erythropoiesis and lymphopoiesis were present in the rat bone marrow; an increase of granulocytopoiesis was noticed as well. The number of colony-forming units of granulocytes, colony-forming units of granulocytes and macrophages, colony-forming units of macrophages, and BFU-E was less compared with the control. The effect of microgravity on the activity of rat hematopoietic cells was studied in vitro as well. Rat CD34+ cells were inoculated into the stromal sublayer and exposed on the Space Shuttle STS-63 and STS-69 for 8–10 days. A significant decrease in CD34
The purpose of biosatellite “BION-M1” flight was to broaden the earlier data on the effects of SF on mammals. The novelty of this project was in the extension of flight duration up to 30 days, the elevation of the altitude up to 575 km and C57BL/6N mice as a principal species for biomedical research. Despite this, our data are in good agreement with the results of the previous series of studies on biosatellites. We assumed that this evidences common patterns of the SF-induced changes. Proliferative activity and the total number hematopoietic colonies remained at the control level, while the inhibition of erythroid lineage occurred. Upon 7 days of recovery, an increase in the share of committed precursors—mainly granulocytic—was noticed. The decrease in the number of erythroid precursors may be associated with a reduction in total blood volume and consequent reduction in erythrocyte number. In addition, the decline in the number of osteoblasts during SF could lead to the reduction of paracrine regulation of hematopoiesis as a number of important regulators of hematopoiesis (CXCL12, osteopontin, angiopoietin-1, TPO, SCF, EPO, HIF1/2α) are produced by osteoblasts [23 –27]. This assumption would be interesting to test in future experiments on a biosatellite.
The results obtained in GC and SF have demonstrated opposite effects on hematopoietic progenitors, especially those of erythroid lineage. Apparently, the changes in the bone marrow of flight and synchronous animals have a different nature. These data are correlated well with the results of Shvets et al. [28]. In the rat's unloading model, they demonstrated a two- to threefold increase of erythroid CFUs after 14 day's limitation of motor activity. On the contrary, the changes in flight and synchronous animals could demonstrate a similar character, as for example, the reduction of the weight of thymus and spleen, but the extent of reduction was more pronounced after the flight [29].
MMSCs are involved in the remodeling of bone, and hematopoiesis support plays an important role in the maintenance of bone marrow and bone homeostasis. In the frame of the BION experiments, the decrease of mineralized tissue, increase of adipose tissue in the bone marrow, reduction of the osteoblast number with sustained number of osteoclasts, and inhibition of periosteal remodeling—mainly in long bones—was revealed [6]. In addition, a decrease in bone marrow stromal progenitor cells (CFU-f) in the flight animals was demonstrated [4]. “Cosmos” biosatellite experiments showed a decrease in mineralization, especially of the long tubular bone, and a decline of osteoblast number and activity. Meanwhile, the number of osteoclasts was constant, or even increased, and early osteoclast precursors appeared [30,31]. These events could provoke a switching of collagen synthesis from bone-specific type I to unspecific type III. It is known that type III is normally produced by smooth muscle cells, fibroblasts, and reticular cells, but not osteoblasts [32]. After 14 days on the biosatellite board, the number of stromal progenitor cells (CFU-f) in the rat bone marrow was notably decreased, compared with control animals [4]. The exposure to weightlessness along 9–14 days during SLS-1 and SLS-2 flights provoked the early signs of osteopenia of the tibia and inhibition of bone growth in length. The presence of mononuclear precursor cells of osteoclasts was noticed [33]. The downregulation of osteonectin, osteocalcin, and type I collagen genes in the cells of the bone tissue of the femur was detected [34].
More recent studies have elucidated the mechanism of the involvement of different bone cells in bone homeostasis under weightlessness. The slowdown of the osteoblast cell cycle and increased osteocyte-mediated osteolysis has been shown under microgravity [8]. Furthermore, the suppression of MMSC commitment was detected, as was evidenced by the downregulation of differentiation-associated genes without affecting the expression of “stemness” genes. In these circumstances, the increase in the number of undifferentiated precursors occurred [11]. However, many issues of undifferentiated stromal precursor potential in the bone marrow remain uncovered.
Our data on the changelessness of mice CFU-f number immediately after SF and recovery period in comparison with vivarium control evidenced the preservation of stromal lineage stability. These observations differ from previous data from earlier experiments (“Cosmos” biosatellite) where the decrease of CFU-f from rat bone marrow was described [4]. This mismatch is probably due to the use of another experimental model (mice), increase of the flight duration, and effects of the overload during the landing. The in vitro analysis of the mononuclear population of bone marrow revealed decreased total cell number in comparison with vivarium control. This observation could be explained by enhanced proliferation of stromal cells during flight due to exposure to low doses of ionizing radiation [35]. In addition, the cells from the SF group seem to have greater attachment efficiency, since the number of PDs in SF and vivarium control groups had no difference. The predominance of loosely packed colonies and reduction of attached cells in the recovery group compared with SF and vivarium controls suggests that both proliferative and adhesive properties of stromal precursors were suppressed after 7 days of recovery.
In the GC experiment, similar events took place. The number of attached cells after 3 days in culture and the area of stromal colonies in the GC group exceeded the same parameters in the vivarium control group, while in the recovery group these parameters were lower than in both mentioned groups.
The larger area occupied by the ALP-stained cells and less heterogeneity in staining evidence the preservation of spontaneous osteocommitment and increased number of stromal cells in the flight group compared to the control and recovery groups. This is consistent with data on the efficiency of cell attachment and morphology of the colonies in the above-mentioned groups.
The present study showed that SF factors did not have a damaging impact on the total MNC population from the C57/Bl/6N mice tibia bone marrow after a 30-day flight. The mild impairment of hematopoietic lineage was observed. In primary cultures, the functions of stromal precursors were well preserved. So far, our data evidence that those hematopoietic cells are susceptible to SF factors, while MMSCs display a high degree of resistance. These data are consistent with previously assumed moderate manifestation and reversibility of the alterations after SF.
Footnotes
Acknowledgments
We are thankful to Dr. Aleksandra Gornostaeva, PhD, for providing technical support. This work was supported by the Program for Basic Research of SSC RF–IBMP RAS.
Author Disclosure Statement
No competing financial interests exist.