
Abstract
Most studies utilizing fish to study the effects of simulated microgravity (SMG) only observe the effects during the first week of development. They also do not take into account the potential impact on development of vibrations caused by the equipment. In this study we analyze the effects of both SMG and vibration on development of the skeleton. We analyze three different exposure durations and starting points that coincide with cranial neural crest cell migration. We use a combination of bone staining and morphometrics to analyze the effects. Our data show that both vibration and SMG affect vertebra number and body size; however, not all vertebrae are equally affected by each treatment. We also show that delayed ossification manifests during development, particularly after SMG exposure, and this translates into buckled and bent bones in adults. This study highlights the large impact of even very short exposure periods when they coincide with critical time points of development.
Introduction
M
Most of the studies utilizing fish to study the effects of SMG only examine the effects over the short term, up to 7 days postfertilization (dpf). For example, these studies have identified delays in development of the vestibular system [4,10], delays in the inflation of the swim bladder [8], reduction in pigmentation [11], and changes in gene expression in the lens, heart, eye, notochord, and neurons [12 –14].
Very few studies have examined the effect of microgravity on the development of the skeleton. In frogs, SMG results in altered morphology of the head cartilage [15], and in the medaka, microgravity has been shown to effect expression of Runx2, a key skeletogenic gene [6]. We previously examined the effects of short, medium, and long embryonic SMG exposure (ranging from 12 to 96 h exposure) on the developing skeleton in zebrafish [11] by exposing fish to SMG at the time of neural crest cell migration [beginning ∼14 h postfertilization (hpf)] [16] and examining the skeleton of adult fish. These data showed that exposing embryos to SMG for 24 h starting at 12 hpf has a long-term effect on some parts of the skeleton, and these effects can persist into adulthood. For example, the parasphenoid and the opercle bones were abnormal in adults. The skulls of these fish were also proportionally smaller than controls as was the entire body length; however, the number of vertebrae was not affected with this exposure timing and duration. Our previous data showed that skull morphology was not severely affected or altered since a functional skull was formed.
In this study, we expand our previous study to examine (1) whether vibration exposure alone can explain our previous observations, and (2) whether by examining different developmental time points we can gain a better understanding of when the skeletal defects arise. We study three different exposure groups (short, medium, and long) and examine fish from 4 dpf (hatching) until 120 dpf (adults).
Materials and Methods
SMG and vibration treatments
SMG environments can be achieved using a simple rotating device, a RWV, otherwise known as a bioreactor. The device used in this study was previously described [11] and has been used by several authors to study the SMG effects on zebrafish [4,8,10,11]. Briefly, the bioreactor (Synthecon, Houston, TX) consists of a hollow transparent Lexan cylinder (about 10 cm in diameter), which surrounds a solid Teflon core (about 5 cm in diameter). The bioreactor cylinder is closed at both ends by Teflon caps and is a water-filled container. The zebrafish embryos are placed in this container. At one end, the Teflon cap secures the bioreactor to a motorized base. The bioreactor was set to rotate on the horizontal axis, at 18.5 RPM, a speed previously established [4,8,10,11]. At this speed, the fluid-filled chamber will cause the zebrafish embryos within it to orbit the central core. The bioreactor however vibrates, and hence, the effect of vibration was also explored.
Groups of fish were exposed to different durations of either SMG or vibrations starting at different developmental stages. Fish were either raised under normal conditions [control no vibrations (CNV)], exposed to vibrations [control with vibrations (C+V)] or exposed to SMG. Embryos exposed to SMG were placed in the bioreactor from 10 or 12 hpf and exposed for 12, 24, or 96 h as follows: from 10 hpf for 12 h (short exposure) or for 96 h (long exposure), or from 12 hpf for 24 h (medium exposure), thus enabling us to compare different starting time points and different lengths of exposure. In the context of neural crest migration, these exposures occur during cranial and/or trunk neural crest migration (Fig. 1).
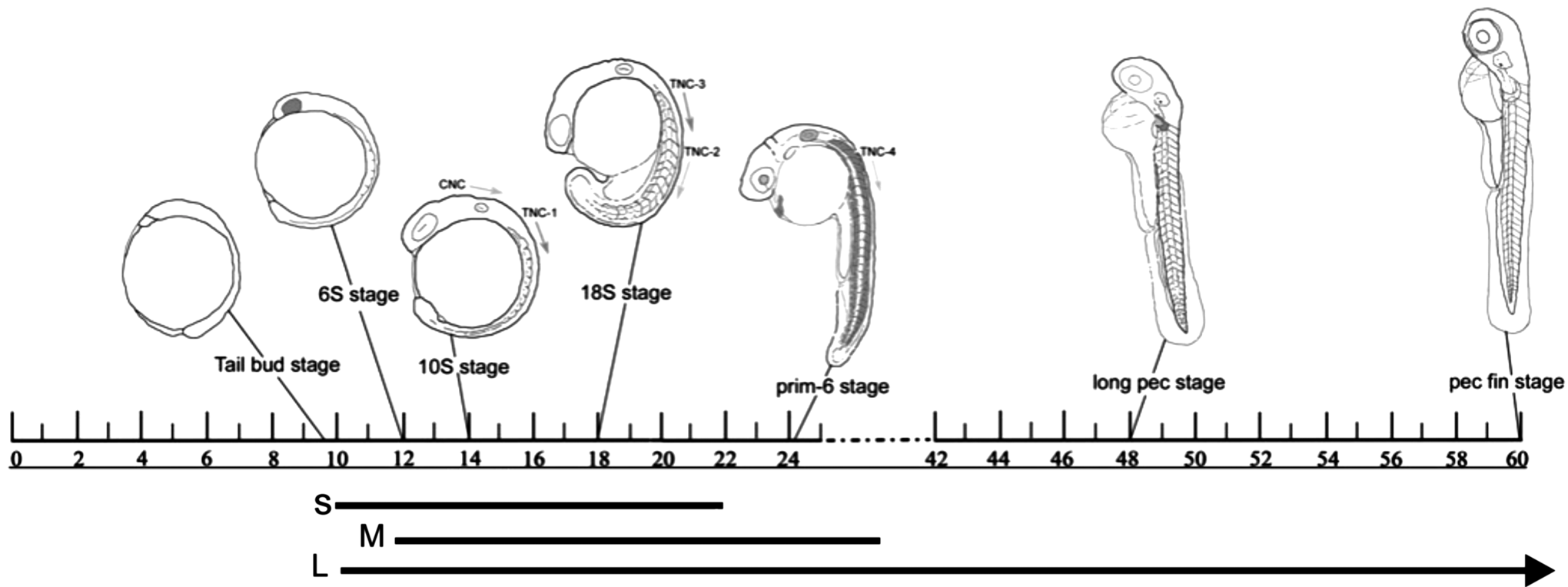
Schematic of neural crest cell migration in zebrafish (10–60 hpf). The 6S stage (6 somite stage) is characterized by the development of the optic primordia; CNC begin to migrate from the neural tube at 8S followed by four waves of TNC beginning at 10–12S stage. By 24 hpf (prim-6), embryos have a beating heart, lens, and optic placodes, and somitogenesis is complete. By 48 hpf (long-pec stage) organogenesis is complete with the exception of the gut; pectoral fin buds begin development. By 60 hpf (high-pec stage), circulation is strong throughout the body and muscle/cartilage differentiation is underway. Lines below the schematic indicate the three exposures used in this study (S, short; M, medium; L, long). Schematic after Ref. [16]. hpf, hours postfertilization; CNC, cranial neural crest cells; TNC, trunk neural crest cells.
Those embryos exposed to vibrations only (C+V) were placed in a water-filled plastic vessel with the same specifications as the bioreactor vessel and positioned on the edge of the bioreactor base, which vibrated. This arrangement ensured that these fish were exposed to vibrations emitted by the bioreactor at the same time points described above for the SMG fish. At the end of the exposure period, embryos were removed from the bioreactor vessel and raised under normal conditions until 4, 10, 35 dpf or 120 dpf. Survival rates were not equal between exposure groups, and some fish did not survive to each of these age groups.
All fish were kept at 28.5°C on a 12–12 h light cycle. Animal care protocols were submitted and approved annually by the SMU-MSVU Animal Care Committee, and guidelines set by the Canadian Council for Animal Care (CCAC) were followed.
Standard length measurements
Zebrafish raised under normal conditions were fixed at 4 dpf, 10 dpf, 35 dpf, and adulthood, 120 dpf (Table 1). The standard length (ie, the distance from the snout of the fish to the base of the tail [17]) of each fish was measured and then used to calculate the means for each age group. These means were compared with the mean standard lengths of fish raised under normal conditions (Table 1). To statistically compare the standard lengths at 120 dpf, independent two-tailed t-tests were performed using the raw values.
dpf, days postfertilization; SL, standard length; n, numbers of fish; CNV, control no vibrations; hpf, hours postfertilization; Indet., indeterminate, the sample size is too small for statistical analyses; C+V, control with vibrations; SMG, simulated microgravity.
Skeletal staining
Fish were euthanized in a lethal dose of MS-222 (ethyl 3-aminobenzoate methanesulfonic acid salt) and fixed overnight in 4% paraformaldehyde (PFA) in 0.01 M phosphate buffered saline (PBS) at 4°C or at room temperature in 10% Neutral-Buffered Formalin. Juvenile fish were stained at 4, 10, or 35 dpf using a standard acid-free double stain for bone and cartilage using Alizarin red S and Alcian blue, respectively (adapted from Ref. [18]). Adult fish were bone stained according to standard procedures using Alizarin red S [19]. Images of dorsal and lateral views were taken using a Nikon DXM1200C camera connected to a Nikon C-DSD115 stereomicroscope.
Vertebra counts
Precaudal and caudal vertebrae, as defined by Ref. [20], were counted for each fish excluding the Weberian apparatus. Nonparametric statistical tests were carried out to determine significance, the Kruskal–Wallis, a one-way analysis of variance of ranks, and the Mann–Whitney U test of significance. These tests were conducted on pairs of groups (eg, treatment vs. CNV and SMG vs. C+V) for the number of caudal vertebrae and the number of total vertebrae.
Morphometrics on the dorsal skull
To determine whether there were any quantitative changes to supplement the qualitative observations, morphometric analyses were conducted on the dorsal bones of the skull. This methodology is well established and has been used in several studies, including those involving zebrafish (eg, Refs. [21,22]). Landmarks were assigned to 15 anatomical reference points and include the premaxilla, the kinethmoid, the frontals and parietals, and the basioccipital (Fig. 2H, Table 2). Thin-plate splines were generated to determine where the changes in morphology (if any) occur. Splines compare the consensuses of two groups (eg, CNV vs. C+V). Landmarks are plotted on a grid, and warping of the grid occurs when there is a difference in landmark location between the reference and experimental consensuses. Statistical analyses (Goodall's F-test) were conducted on each set of comparisons to determine whether or not the warping observed is statistically significant (Table 3). This analysis could not be conducted for the 12 h C+V at 10 hpf group since there was only one fish in this group.
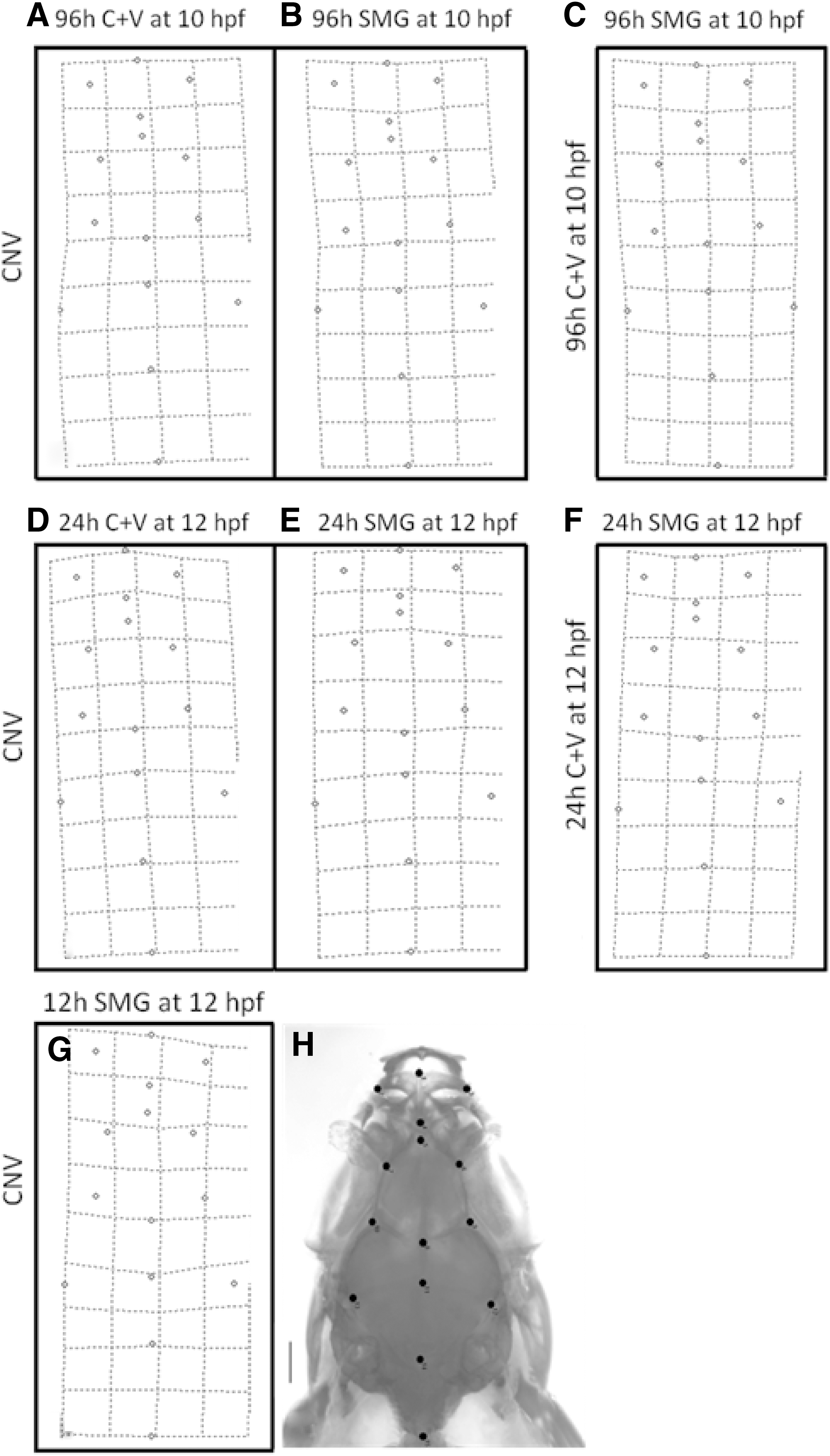
Thin plate splines showing differences in landmark location after treatment. The reference consensus for each spline is indicated on the left of the spline, and the experimental consensus is indicated above the spline.
Results
SMG and vibration exposure impacts body size
To determine whether SMG exposure and vibration exposure affected growth of the fish, we measured the standard length of fish after exposure at various developmental time points and compared this to fish raised under standard conditions in our facility.
We previously showed that adult fish were significantly shorter after short (12 h SMG at 10 hpf) and medium (24 h SMG at 12 hpf) exposure, but were not affected after long exposures (96 h SMG at 10 hpf) [11]. In this study we investigate whether fish are of a similar size at hatching, and whether exposure to vibrations alone can explain the previous results since the bioreactor vibrates when it turns. Our data show that fish are similar size at hatching (4 dpf) regardless of exposure duration or type (Table 1). At 10 dpf, significant differences in body length were observed after SMG exposure in the short exposure group only (Table 1). Several months later (120 dpf), fish in the short and medium SMG exposure groups (12 h SMG at 10 hpf and 24 h SMG at 12 hpf) are significantly different to the CNV group (Table 1). Interestingly, fish in the medium and long vibration groups (24 h C+V at 12 hpf, 96 h C+V at 10 hpf) are also significantly different from fish in the CNV group; the short exposure group sample size was too small to compare (Table 1). Thus based on these data, we conclude that both vibration exposure and SMG exposure affect adult body size and that this effect is dependent on both the starting point of exposure and the duration of exposure.
The effect of vibration appears less extreme than the effect of SMG exposure, when comparing the standard lengths of fish in SMG groups with the corresponding vibration exposure times and durations (ie, 24 h C+V at 12 hpf cf. 24 h SMG at 12 hpf, Table 1). This is supported by the survival rates (Table 1); more SMG fish survived to adulthood than C+V fish in both the medium and long exposure groups.
Effects on vertebra number
To further investigate the effects on growth, we examined the numbers of vertebrae in the vertebral column after each exposure group. The range of precaudal vertebrae was 10–11 in the control group (CNV); this is within the published range of 9–11 [20]. Fish from the SMG groups had 10–12 precaudal vertebrae, whereas vibration control groups (C+V) had 9–10 vertebrae (Table 4). However, only a small proportion of the fish differed from the CNV group with respect to numbers of precaudal vertebrae (only 2/21 SMG fish had 12 precaudal vertebrae, and only 2/19 vibration (C+V) fish had nine vertebrae). Thus it is possible that vibration exposure alone caused the variation observed in the precaudal vertebrae number (discussed later).
Regarding the caudal vertebrae in the CNV group, the majority of fish (10/15 fish) had 15 caudal vertebrae (range was 15–16 vertebrae, Table 4); 15 caudal vertebrae are the published number [20]. Interestingly, SMG exposed fish had a wide variation in caudal vertebra number (range was 13–17 caudal vertebrae), while the vibration groups (C+V) had 14–16 caudal vertebrae. Specifically, 58% (7/12) of medium exposure group (24 h SMG at 12 hpf fish) do not have 15 caudal vertebrae compared with 33% (2/6) of the corresponding vibration group (24 h C+V at 12 hpf), suggesting that SMG exposure has an additive effect on top of vibration exposure to contribute to the variable caudal vertebra number observed (Table 4).
For the treatments starting 2 h earlier, at 10 hpf, but of a longer duration, 96 h, 40% (2/5) of this long exposure group (96 h SMG at 10 hpf) and 41.6% (5/12) of the corresponding vibration exposure group (96 h C+V at 10 hpf) do not have 15 caudal vertebrae; this indicates that the effect of SMG and vibration exposure increases when treated earlier for a longer period of time. The sample sizes for the treatments for short duration (12 h starting at the 10 hpf) were low due to less fish surviving to adulthood. Of those that did survive, the number of caudal vertebrae is variable and inconclusive (Table 4). In summary, variation in caudal vertebra number was found after SMG and vibration treatment, and it appears that both act to effect caudal vertebra number and that longer duration of exposure results in a greater effect.
Our data also show that caudal vertebrae display more variation in number than the precaudal vertebrae. However, although the average vertebrae counts were not statistically significant to the CNV group, a statistical difference was found between the average number of vertebrae in the CNV group compared with the short exposure (12 h SMG at 10 hpf, P < 0.05) group. This treatment group however has a small sample size; therefore further research is needed in order to confirm this result.
Effects on skull morphology variable between adults and juveniles
The effects of SMG exposure and vibration exposure on the adult and juvenile skull morphology were accessed after short (12 h), medium (24 h), and long (96 h) exposures. In adults, the left lateral side and the dorsal view of the skull were assessed.
In adults, no morphological differences belonging to the three vibration (C+V) groups compared with the CNV group were observed (not shown); therefore, the SMG groups were compared with the CNV group. Our data show that the long exposure of 96 h SMG at 10 hpf has no effect on skull development, in contrast to the medium exposure group (24 h SMG at 12 hpf) and the short exposure group (12 h SMG at 10 hpf) (Fig. 3A–C). In these latter groups, notable changes in several bones were observed—the parasphenoid bone morphology is abnormal, delayed ossification was observed in the orbitosphenoid, and the supraoccipital and exoccipital bones varied in size [11]. Collectively these abnormalities contribute to a rounding of the frontal and parietal bones. The skulls of these fish were also notably truncated and/or narrower/wider than control fish.
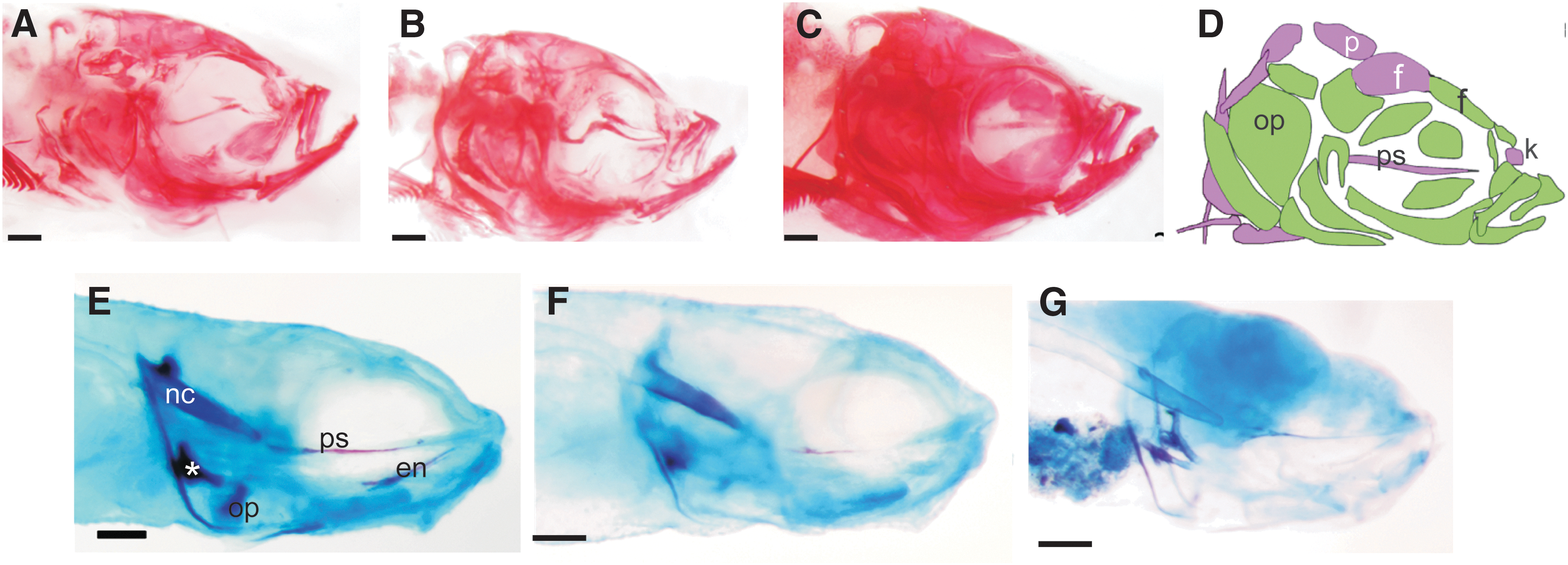
Morphological differences among SMG exposed fish.
To determine whether there were any quantitative changes to supplement the qualitative observations, morphometric analyses were conducted on the dorsal bones of the skull. Thin-plate splines were generated to determine where the changes in morphology (if any) occur (Fig. 2). They show a slight warping in the anterior and lateral regions of all the splines, suggesting variation in the premaxillae, kinethmoids, and along the lateral edges of the frontals and parietals in all groups. The warping, however, does not appear to be dramatic in any of the splines, particularly in the posterior end of the skull. Statistically significant differences were however found between five of the seven comparisons conducted (Table 3), and these differences were less significant between vibrations (C+V) and CNV than between SMG and CNV fish. The long exposure SMG group was an exception (and not significantly different to controls), unlike the short and medium exposure groups, which were significantly different to the both types of controls (C+V and CNV fish). These data suggest that SMG exposure has a greater effect on skull development than vibrations alone.
To further investigate when these defects may arise, the juvenile skull was also examined. Fish stained at 4 dpf do not exhibit abnormal skull morphologies (not shown); however, at 10 dpf differences were observed. We previously reported that fish from the medium exposure group (24 h SMG at 12 hpf group, n = 5) had delays in ossification of much of the skull [11]. Fish from the CNV group (n = 9) resembled fish from the C+V groups (not shown); however, they were more ossified than all the SMG exposed fish (Fig. 3). In this study we show that fish from the short exposure group (12 h SMG at 10 hpf, n = 8) show a similar progression of ossification of the parasphenoid, notochord, and ceratobranchial-5 as the CNV fish, while some fish in this group (28%) showed additional delays in the ossification of the opercula and entopterygoids (Fig. 3). Unfortunately, fish from the 96 h (long exposure) group did not maintain integrity during staining and could not be examined. Interestingly, at 35 dpf skull abnormalities and ossification delays were not observed in any of the fish (not shown). These data indicate that delays in ossification arise after 12 h SMG exposure to the larvae and that with persistent exposure, additional abnormalities such as odd bone shapes and buckling can occur. These ossification defects are not observed several weeks later (at 35 dpf, not shown) after rearing under normal conditions. In adults, only buckling is observed since ossification is completed sometime earlier. Since these ossification defects were not observed after vibration exposure, it suggests that the effects are due to SMG exposure.
Discussion
In this study, we investigated the effects of vibration exposure compared with SMG exposure during embryonic development on the adult skeleton of zebrafish. Data on the effects of simulated or true microgravity on the skeleton are not extensive, and the long-term effects are rarely investigated. Yet the use of zebrafish in these types of studies is valuable since the cells in zebrafish bone (osteoblasts, osteocytes, and osteoclasts) are the same as those in human bone [23].
Effects on body size (ie, standard length and vertebra counts)
Our data show that vibration exposure affects body length and vertebra number and that SMG exposure increases these effects. We previously showed that skull length after SMG exposure was proportional to body length and that the effects on body length are most likely due to stunted growth [11]. Although individual vertebra sizes were not measured, the data suggest that, at least in some cases, somites were smaller in some fish (ie, the smallest fish sometimes had more vertebrae than larger fish).
Somitogenesis is a highly controlled process during development that involves a molecular clock. Although the periodicity of this clock varies between organisms, within a species it is tightly controlled. Changes in somite number in zebrafish have been documented when fish are subjected to stress. Connolly and Hall [16] noted that the greatest difference in caudal vertebra counts occurred in fish heat-shocked between 13 and 17 hpf; this coincides with the timing of the 12 h SMG at 10 hpf, the short exposure group in the current study. Heat shock elicits a stress response that involves upregulation of heat-shock proteins [16].
Indeed, studies have shown that exposure to microgravity results in an upregulation of hsp70 [12]. Thus it is possible that exposure to vibrations and/or SMG results in upregulation of heat shock proteins and that these affect somitogenesis and ultimately the number of vertebrae that form. In our analyses, precaudal vertebrae number increased after SMG exposure but decreased after vibration exposure, suggesting that the molecular clock underlying somitogenesis was altered differently in each case. Stopping the clock involves Mesp, whereas changing the boundaries of the somites (ie, vertebra size) involves altering the determination front (fgf8 expression boundary or Notch signaling). Future research is needed to tease apart the effects of vibration versus SMG on heat-shock proteins and on somitogenesis.
Since the effects on the precaudal and caudal vertebrae were not the same, this suggests that either the timing of exposure relative to development of these two regions of the vertebral column was different or that the molecular clock underlying their development is not identical in all aspects. Connolly [24] noted that precaudal vertebrae displayed less variation than caudal vertebrae after heat-shock treatment at 12/13 somite stage (15 hpf) and at 60 hpf. Similarly, in this study we show that the caudal vertebrae are more variable in number than precaudal vertebrae, suggesting that the latter are more robust or that the timing of treatment coincided more closely with caudal vertebra development.
Effects on bone development
Bone is a dynamically remodeled tissue that requires mechanical stimulation for maintenance—a balance between the activities of osteoclasts, osteoblasts, and osteocytes is required. All three cell types are present in zebrafish [23]. Differences include the presence of both mono- and multinucleated osteoclasts in teleosts, including zebrafish [25], as well as the presence of different types of bone compared to in humans. For example, zebrafish have cellular bone and acellular bone [26,27]. These bones can be directly developing (ie, they form intramembranously without an association with cartilage, eg, calvariae) or indirectly developing (ie, they form endochondrally, perichondrally, or periskeletally, eg, mandible, scleral ossicles, and so on). Since fish don't have hematopoietic tissue inside the bone marrow, bone marrow spaces are filled with fat tissue, as well as nerves and blood vessels [27]. This fat-filled bone (known as tubular bone) serves as an adaptation to buoyancy [27]. Despite these differences, the cellular components and developmental origins of the bone-associated cells are conserved between zebrafish and humans, making zebrafish a valuable model organism in which to study conditions (such as SMG exposure) or diseases affecting human skeletal tissues.
Microgravity exposure can cause osteoclast-mediated bone resorption, as well as increased osteocyte-mediated bone resorption, known as osteocytic osteolysis [28]. There are some discrepancies between the effects caused by SMG versus true microgravity. Blaber et al. [9] suggested that SMG using RWVs may activate signaling that could increase differentiation of stem cells since these vessels do not reduce the overall mechanical stimulation from fluid flow shear and hydrostatic pressure; however, this likely only applies to cells adhering to these vessels and not to zebrafish embryos within the RWV chamber used in this study; embryos are suspended midway between the core and the wall of the chamber by the correct setting of 18.5 RPM [4,10].
Shimada et al. [5] demonstrated that various durations of exposure (18–60 h) to SMG starting at varying time points (8–56 hpf) affected gene expression in several zebrafish organ systems, but that these systems recover within 80 h of being removed from exposure. Although these researchers did not rear the fish to adulthood, the observed recovery at the gene level of the affected organ systems suggests that these organs would have developed relatively normally. Our data show that SMG exposure during embryonic development does not cause severe defects, and patterning of the skeleton is not affected supporting this hypothesis.
At 10–12 hpf, zebrafish embryos are undergoing somitogenesis, which ends at 22 hpf. Embryos were removed from the exposure treatment (SMG or vibration) after 12, 24, or 96 h starting at 10 or 12 hpf (ie, at 22 hpf, 36 hpf, and 106 hpf). This treatment period also spans the time when organogenesis occurs (including development of the heart, circulatory system, eye tissue differentiation, fin development, and jaw development). Since the fish appeared normal with respect to behavior, we assume that these structures were not affected; however the low survivability of some groups (eg, 12 h C+V at 10 hpf and 12 h SMG at 10 hpf) may indicate that some of these vital organs were indeed affected by SMG and vibration exposure. More research is needed to explore this further.
Research shows that subjecting bone to hypergravity by vibration results in accelerated osteoblastic and osteoclastic activity [29]. Low frequency vibrations can also affect early embryonic development of Xenopus laevis (frog) and Danio rerio (zebrafish) [30]. Specifically, embryos were exposed to vibrations from the one to two cell stage until the five somite stage (for the zebrafish). Results show that at 7 dpf, the zebrafish larvae had altered left-right patterning and tail morphogenesis. Skeletons were not examined in this study. Slow turning vessels used to assess effects of SMG and vibration on the development of cartilages during frog development show that increased cartilage sizes and increased cell division may be due to the effects of vibration while SMG resulted in abnormally shaped cartilages, particularly those that were neural crest derived [15]. This study exposed embryos for 4 days from neurulation through to tadpole stages and concluded that the underlying mechanism was either disruption in neural crest cell migration or chondrocyte differentiation.
The potential effect of a long exposure of SMG or vibration (for 96 h starting at 10 hpf) on neural crest migration was explored in zebrafish by examining whether pigmentation was altered after exposure [11]. This study showed that there were significantly fewer melanophores after SMG exposure compared to fish exposed to vibrations. These effects were very short term, and recovery was observed within 1 day after exposure. Further research is needed to determine whether there was a delay in the migration of the melanophores or a delay in melanophore differentiation.
The origin of the zebrafish skull is both neural crest and mesodermal; however, the boundary between these two regions has shifted anteriorly in zebrafish [31] compared to other vertebrates (Fig. 2D [31]). We previously showed that some bones of the adult skull (eg, opercle and parasphenoid) were abnormal after medium exposure to SMG [11]. These bones displayed delayed ossification and/or buckling. Delayed ossification likely contributes to the buckling of bones. In this study, we show that delayed ossification is also present in the short exposure group, but is not present after vibration exposure. Delayed ossification could be the result of delayed neural crest migration, delayed differentiation of osteo- and chondroblasts or delayed matrix production.
True microgravity has been shown to reduce the differentiation of embryonic stem cells [9] and reduce the differentiation capacity of cells from the ectoderm, mesoderm, and endoderm lineage [9,32 –34], although this response is tissue specific. We have shown that both mesodermally and neural crest derived bones were affected by SMG exposure. Thus it is possible that osteoblast differentiation may have been affected in the current study, and this led to delays in ossification. Research examining the effects on differentiation markers and extracellular matrix markers are needed to fully elucidate these results.
Collectively, these results indicate that there may be a delay in cell migration and/or a delay in cell differentiation after SMG exposure and that these effects are less extreme than in vibration exposure controls. A more detailed analysis of these underlying mechanisms is necessary. Zebrafish provide an excellent system in which to elucidate these effects due to the transparency of their embryos, the array of transgenic lines with skeletal markers, and the ease with which to expose them to SMG.
Footnotes
Acknowledgments
This study was funded by Canadian Space Agency (9F007-07123) and the Natural Sciences and Engineering Research Council of Canada (Discovery Grant 328376) to TFO.
Author Disclosure Statement
Authors declare no competing financial interests. Authors have no commercial associations. The funders had no role in the study design, data collection, and analysis.