
Abstract
Islet regeneration is stimulated after transplantation of human umbilical cord blood (UCB) hematopoietic progenitor cells with high aldehyde dehydrogenase (ALDH)-activity into NOD/SCID mice with streptozotocin (STZ)-induced β cell ablation. ALDHhi progenitor cells represent a rare subset within UCB that will require expansion without the loss of islet regenerative functions for use in cell therapies. ALDHhi cells efficiently expand (>70-fold) under serum-free conditions; however, high ALDH-activity is rapidly diminished during culture coinciding with emergence of a committed megakaryocyte phenotype CD41+/CD42+/CD38+. ALDH-activity is also the rate-limiting step in retinoic acid (RA) production, a potent driver of hematopoietic differentiation. We have previously shown that inhibition of RA production during 9-day cultures, using diethylaminobenzaldehyde (DEAB) treatment, enhanced the expansion of ALDHhi cells (>20-fold) with vascular regenerative paracrine functions. Herein, we sought to determine if DEAB-treatment also expanded ALDHhi cells that retain islet regenerative function following intrapancreatic transplantation into hyperglycemic mice. After DEAB-treatment, expanded ALDHhi cell subset was enriched for CD34+/CD38− expression and demonstrated enhanced myeloid multipotency in vitro compared to the ALDHlo cell subset. Unfortunately, DEAB-treated ALDHhi cells did not support islet regeneration after transplantation. Conversely, expanded ALDHlo cells from DEAB-treated conditions reduced hyperglycemia, and increased islet number and cell proliferation in STZ-induced hyperglycemic NOD/SCID mice. DEAB-treated ALDHlo cells were largely committed to a CD41+/CD42+ megakaryocyte phenotype. Collectively, this study provides preliminary evidence that committed cells of the megakaryocyte-lineage support endogenous islet regeneration and/or function, and the retention of high ALDH-activity did not coincide with islet regenerative function after expansion under serum-free culture conditions.
Introduction
T
There is accumulating evidence that demonstrates human pancreas contains resident stem/progenitor cell populations [4 –6] and DM may be effectively treated by the restoration of physiologically functional β cell mass [5,7]. As a result, cell-based therapies have become a focus of translational research aiming to restore depleted β cell mass be either replacement or regeneration strategies. Recent phase I/II clinical investigations to replace (NCT02239354) or regenerate (NCT00690066) β cell mass are currently underway or have reported early results [8,9].
In 2010, the Joslin Medalist study reported measurable C-peptides levels and actively proliferating β cell mass in the pancreata of individuals with Type I DM for >50 years [10]. This encouraged the idea that transplantation of appropriate cell types and/or stimuli may boost endogenous β cell regeneration in the islets of a diabetic individual [6]. We initially demonstrated human bone marrow (BM) and umbilical cord blood (UCB)-derived progenitor cells selected for high aldehyde dehydrogenase (ALDH) activity could stimulate endogenous β cell regeneration and reduce hyperglycemia in streptozotocin (STZ)-treated NOD/SCID mice [11,12]. We have recently demonstrated the secretome of blood-derived progenitor cells such as multipotent stromal cells or hematopoietic progenitor cells (HPC) can stimulate human β cell proliferation in vitro [13,14]. It has also been demonstrated that mature hematopoietic cells, such as platelets, may have β cell supportive functions during the clinical treatment of Type 1 DM autoimmunity [15]. Collectively, identifying phenotypic characteristics islet-regenerative cell populations and the effectors used to orchestrate robust islet regeneration is crucial to effective treating or reversing DM.
UCB represents an accessible source of hematopoietic, endothelial, and mesenchymal progenitor cells and collection imposes no risk to either the newborn or mother. The worldwide establishment of public and private cord blood-banking registries has generated a renewable pool with increased diversity for human leukocyte antigen (HLA)-matching for use in therapeutic applications. We view UCB as a favorable source of progenitor cells compared to autologous bone marrow, as accumulating evidence demonstrates loss of progenitor cell number and function when cells are harvested from the microenvironment of chronic diseases [16 –19]. Cells harvested from healthy allogenic sources (ie, UCB) are untouched by damaging metabolites that circulate throughout the body of diabetic individuals [18]. Further research is warranted to determine the full potential of UCB cell progeny for therapeutic strategies to treat DM and common cardiovascular comorbidities. The selection of UCB ALDHhi cells isolates a HPC-enriched cell population with multifaceted regenerative functions [12,20]. Unfortunately, UCB ALDHhi cells represent a rare subpopulation (<0.5% of total UCB cells; ∼1.0 × 105–5.0 × 105/100 mL of UCB) that will require ex vivo expansion to increase regenerative cell numbers for translational applications.
UCB ALDHhi cells efficiently expand under serum-free culture conditions (>70-fold after 9 days; >1.0 × 107 viable cells); however, a primitive cell phenotype and regenerative function are rapidly lost [21 –23]. We have recently identified that UCB ALDHhi cells preferably differentiate toward the megakaryocyte lineage as culture time was increased [22]. Specifically, we detected increased expression of megakaryocyte-specific lineage markers, including CD41 (ITGA2) and CD42 (GP1B), by global proteomic analyses and validated by multiparametric flow cytometry. UCB ALDHhi cells became enriched for a megakaryoblast phenotype (CD41+/CD42−) during ex vivo expansion and subsequently acquired a committed megakaryocyte phenotype (CD41+/CD42+); coinciding with loss of high ALDH-activity (<10%) and vascular regenerative function in vivo. The loss of islet regenerative function in ALDHhi cells occurs early following ex vivo expansion, as ALDHhi cell reselected after 6 days of culture do not stimulate islet regeneration following intrapancreatic injection in hyperglycemic NOD/SCID mice. Thus, prevention of ALDHhi cell differentiation, via small-molecule supplementation, may be required to expand UCB hematopoietic progeny capable of robustly stimulating islet regeneration.
ALDH is rate limiting enzyme in the production of retinoic acid (RA), a potent driver of HPC differentiation via the activation of the RAR/RXR nuclear receptor complex (RAR/RXR) [24]. Activation of the RAR/RXR complex will increase the production of downstream products that initiate hematopoietic differentiation and/or cell proliferation [25 –27]. This imposes a paradoxical challenge to efficiently expand ALDHhi HPC while limiting the intrinsic propensity for hematopoietic differentiation in vitro. Reversible inhibition of ALDH1A-activity, using diethylaminobenzaldehyde (DEAB) supplementation, limited HPC differentiation during 9-day culture and generated an ALDHhi cell-enriched population (>20% of day 9 progeny; >20-fold increase of ALDHhi cells) that retained vascular regenerative function in vivo [22]. Herein, we investigated the islet regenerative capacity of expanded UCB ALDHhi cells reselected for high versus low ALDH-activity following 9 days culture under previously described DEAB-treated culture conditions. We hypothesized expanded ALDHhi cells reselected from DEAB-treated conditions would demonstrate islet regenerative function following intrapancreatic transplantation into STZ-induced hyperglycemic NOD/SCID mice. Surprisingly, this study identified that only a CD41+ megakaryocyte-lineage enriched ALDHlo cell population harvested from DEAB-treated conditions supported islet regeneration, albeit modestly. Collectively, this study provides evidence that the retention of high ALDH-activity, via DEAB-treated conditions, does not coincide with islet regenerative function.
Materials and Methods
UCB ALDHhi cell isolation and expansion
Human UCB samples were obtained with informed consent by venipuncture of the umbilical vein following Ceasarian section at Victoria Hospital Birthing Centre, London, ON, Canada (REB No. 12934). The Human Studies Research Ethics Board (HSREB) at Western University approved all procedures. Within 24 h of collection, UCB samples were initially depleted of mature myeloid, lymphoid and red blood cells using RosetteSep Human Cord Blood Progenitor Cell Enrichment Cocktail (Stem Cell Technologies, Vancouver, BC, Canada). Lineage-depleted (Lin−) mononuclear cells were isolated by Hypaque Ficoll centrifugation and incubated with Aldefluor regent before purification by FACS (FACS Aria III; BD Biosciences, Mississauga, ON, Canada). UCB ALDHhi cells were selected based on low side scatter and high ALDH-activity with >98% purity (n = 5). Purified UCB ALDHhi cells were cultured on fibronectin-coated plates for 9 days under untreated (X-vivo 15 + 10 ng/mL SCF, TPO, FLT-3L) or DEAB-treated (Untreated+10 μM DEAB on days 3–6), as previously described (Fig. 1A) [21 –23].

Expansion of UCB ALDHhi cells generated cells with low ALDH-activity and a megkaryocyte phenotype.
Flow cytometry
UCB ALDHhi cells expanded under untreated or DEAB-treated conditions were harvested at day 9 and enumerated by trypan blue exclusion using hemocytometer counts. Cells were labeled with Aldefluor to assess ALDH-activity and co-stained with 7-amino-actinomycin D to determine viability. Fresh UCB Lin− MNC and expanded UCB ALDHhi cell progeny were co-labeled with anti-human antibodies for CD41 (ITGA2), and CD42 (GP1B; megakaryocyte). Expanded cells were assessed for a primitive HPC phenotype using anti-human CD34 and hematopoietic lineage differentiation was determined using anti-human CD38 (cyclic ADP ribose hydrolase). Cell surface marker expression for fresh cells, and expanded ALDHhi versus ALDHlo subsets was determined using an LSRII flow cytometer (BD Biosciences) at the London Regional Flow Cytometry Facility. All analyses were performed using FlowJo software v8.2.
Hematopoietic colony formation
Day 9 expanded cells were harvested from untreated or DEAB-treated conditions and reselected for high or low ALDH-activity before being seeded in semi-solid methylcellulose media (H4434; Stem Cell Technologies) at 500 cells/mL in 12-well plates, performed in duplicate (Fig. 3C). Hematopoietic colonies were enumerated after 12–14 days in vitro and scored based on morphological characteristics of burst forming unit erythocyte (BFU-E), colony forming unit granulocyte (CFU-G), colony forming unit macrophage (CFU-M), colony forming unit granulocyte/macrophage (CFU-GM), or colony forming unit mixed (CFU-Mixed) [20]. CFU/100 cells were calculated by number of colonies formed per 100 cells seeded in the assay. Furthermore, the frequency of lineage-specific colony formation was calculated by dividing the number of a lineage-specific colony (ie, BFU-E) by the total number of colonies enumerated.
Transplantation of expanded ALDHlo and ALDHhi cells
Pancreatic β cell ablation was induced in NOD/SCID mice aged 8- to 10-weeks (Jackson Laboratories, Bar Harbor, ME;
Immunohistochemistry and immunofluorescent analyses
Pancreata were harvested, embedded in OCT, and cryosectioned (12 μm) to obtain three sections per slide ≥200 μm apart. Cryosections were fixed in 10% formalin (Sigma) and blocked with 5% serum before immunohistochemical staining for insulin to determine islet number, size, and β cell area as previously described [23]. Additionally, immunofluorescent co-staining for insulin with EdU, CD31, Nkx6.1, and/or HLA-A,B,C was performed. EdU+ proliferating cells were detected using the Click-iT imaging kit (Alexafluor 488; Life Technologies). Intra-islet capillary density was enumerated post-transplantation using rat anti-mouse CD31 (1:100; BD Biosciences) detected with FITC-labeled rabbit anti-rat secondary antibodies (1:200; Vector). Nxk6.1+ cell nuclei were enumerated using anti-rabbit (1:2000; AbCam) and detected using goat anti-rabbit FITC-labeled secondary antibodies (1:200; Vector). Human cell engraftment was enumerated by staining pancreas sections with mouse anti-human HLA-A,B,C (1:100; AbCam) detected with horse anti-mouse FITC-labeled secondary antibodies (1:200; Vector). DAPI was used as counterstain to label nuclei. Quantification and analyses were performed using Fiji (Image J v2.0).
Quantification of islet number, size, and β cell mass
Islet number was quantified across four random images per tissue section (12 images per mouse) by enumerating the presence of insulin+ islet structures containing more than 20 nuclei. Islet size was determined by measuring the circumference of each islet identified in the previous quantification using Fiji (Image J v2.0). Lastly, β cell mass of each mouse was calculated:
Statistical analyses
Statistical analysis for flow cytometry and colony analyses was performed by one-way ANOVA with Tukey's multiple comparisons tests. Analysis of significance was performed by one-way ANOVA with Tukey's multiple comparisons tests for all in vivo experiments and enumeration of histological analyses. Outliers were identified using Grubb's test; P < 0.05.
Results
High ALDH-activity does not select for the megakaryocyte lineage in fresh UCB cells
We have previously demonstrated that purified UCB Lin− ALDHhi cells represent a heterogeneous progenitor cell population with multifaceted regenerative functions [12,20]. However, the robust expansion of UCB Lin− ALDHhi cells (>70-fold) under serum-free conditions containing SCF, Flt-3L, and TPO generated a megakaryocyte-enriched population with diminished ALDH-activity and progenitor cell surface marker expression [22]. To understand the propensity of UCB Lin− ALDHhi cells to differentiate toward the megakaryocyte lineage, we sought to determine whether the initial selection of primitive cells based on high ALDH-activity also enriched for committed precursors within the megakaryocyte lineage. Therefore, we first compared fresh UCB Lin− MNC, or FACS-purified ALDHhi and ALDHlo cells for megakaryoblast (CD41+/CD42−) and megakaryocyte (CD41+/CD42+) cell surface marker expression (Fig. 1A). The frequency of cells with CD41+/CD42− megakaryoblast phenotype was equivalent between freshly isolated UCB Lin− MNC (28.8% ± 6.12%), or purified ALDHhi (26.9% ± 2.1%) versus ALDHlo cell subpopulations (29.1% ± 6.4%; Fig. 1B). Moreover, the frequency of committed CD41+/CD42+ megakaryocytes was also equivalent within the fresh UCB Lin− cell subsets (Fig. 1C). Thus, selection of UCB Lin− cells based on high ALDH-activity did not enrich for cells of the megakaryocyte lineage; although ∼60% of UCB ALDHhi cells expressed early megakaryocyte marker CD41.
The expanded ALDHhi cell subpopulation was enriched for a megakaryoblast phenotype and the ALDHlo cell subpopulation was enriched for a megakaryocyte phenotype
The ex vivo expansion of UCB Lin− ALDHhi cells under DEAB-treated culture conditions robustly increased the total number of viable ALDHhi cells for preclinical studies (>20-fold) [22]. Despite the retainment of ALDHhi cells under DEAB-treated conditions, the frequency of CD41+/CD42+ megakaryocytes in the total cell population was comparable to untreated conditions. It is currently unreported whether ALDHhi cells reselected from DEAB-treated conditions are enriched for specific subpopulations of the megakaryocyte lineage, compared to untreated conditions. Following ex vivo expansion of UCB Lin− ALDHhi cells under untreated or DEAB-supplemented conditions, cell progeny that retained high ALDH-activity was enriched (*P < 0.05) for early megakaryoblast (CD41+/CD42−) phenotype, compared to the ALDHlo cell subpopulation (Fig. 1D) from either condition. Conversely, the ALDHlo cell subpopulation generated under untreated and DEAB-treated conditions was significantly enriched (**P < 0.01) for a committed CD41+/CD42+ megakaryocyte phenotype, compared to the ALDHhi cell subpopulation (Fig. 1E). Collectively, DEAB-treated conditions generated CD41+/CD42− megakaryoblasts that retained ALDHhi progenitor cell characteristics and reduced the commitment of UCB Lin− ALDHhi cells toward an ALDHlo CD41+/CD42+ megakaryocyte phenotype.
Reselection of ALDHhi cells enriched for primitive phenotypes and increased multipotent hematopoietic colony formation in vitro
Multipotent HPC with colony formation function in vitro and SCID-repopulating capacity in vivo can be enriched from fresh UCB samples by the selection of CD34+/CD38− or ALDHhi cells [28,29]. To determine whether multipotent HPC remained after 9-days culture, we assessed expanded UCB Lin− ALDHhi cells for a primitive progenitor cell phenotype and for clonal colony formation in vitro. Following ex vivo expansion of UCB Lin− ALDHhi cells under untreated or DEAB-supplemented conditions, reselected ALDHhi cells were significantly enriched (**P < 0.01) for primitive CD34+/CD38 cell surface marker expression (20.8% ± 7.3% or 15.1% ± 3.9%), compared to the ALDHlo cell subpopulation (1.7% ± 0.7% or 1.7% ± 0.5%; Fig. 2A). The frequency of CD34+/CD38 cells in the ALDHhi subset was comparable between culture conditions (Fig. 2A). Regardless, we have previously reported that the enrichment of ALDHhi cells under DEAB-treated conditions induced a more than twofold increase in the number of ALDHhi/CD34+/CD38 cells, compared to untreated conditions [22]. Importantly, DEAB-treated conditions induced a significant decrease in frequency of ALDHhi or ALDHlo cells with CD38+ expression (Fig. 2B), a marker of hematopoietic maturation [27]. To determine hematopoietic colony forming capacity in vitro, ALDHhi and ALDHlo cells were reselected after expansion under untreated and DEAB-treated conditions and seeded in methylcellulose media. Hematopoietic colony formation was manually enumerated after 14 days (Fig. 2C). Notably, ALDHhi cells from either untreated or DEAB-treated conditions were significantly enriched for colony forming cells compared to ALDHlo cells (Fig. 2D). ALDHhi cells from DEAB-treated cultures also demonstrated a higher frequency (P < 0.05) of colony formation (1 CFU in 10.5 cells, P < 0.05) compared to ALDHhi cells expanded under untreated conditions (1 CFU in 30.0 cells). The frequency of myeloid colonies formed was found to be similar to cells expanded under untreated or DEAB-treated conditions (Fig. 2E). Within the ALDHhi or ALDHlo subsets only ALDHhi cells demonstrated myeloid multipotency by the formation of CFU-mixed colonies, whereas ALDHlo cells largely contained progenitors restricted to the macrophage (CFU-M) or granulocyte (CFU-G) lineages (Fig. 2E). Collectively, these results demonstrate DEAB-treated conditions reduced the acquisition of mature marker CD38 and augmented the expansion of ALDHhi cells that retained multipotent hematopoietic colony forming capacity in vitro.
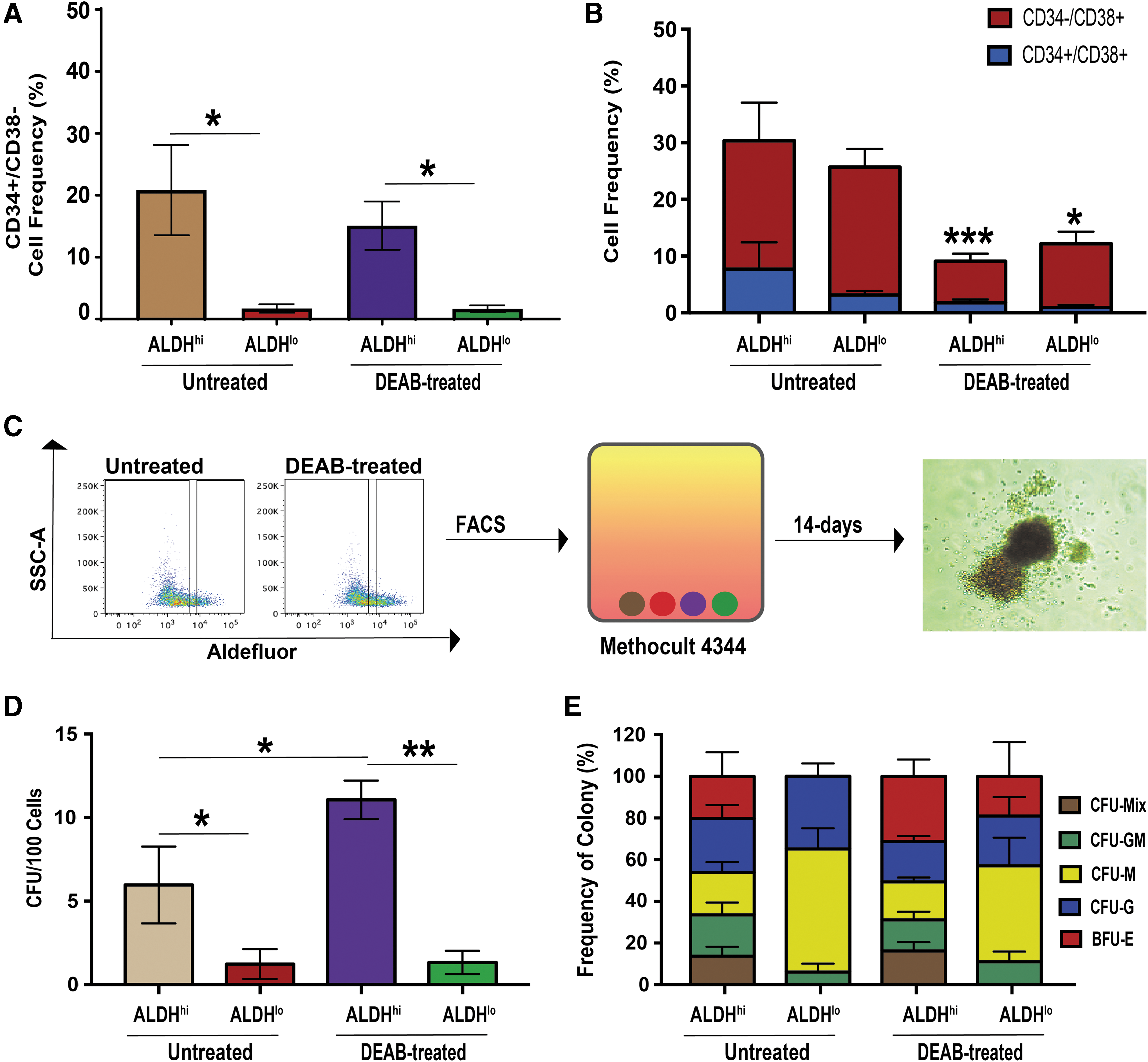
Reselection of expanded cells with high ALDH-activity enriched for cells retaining primitive (CD34+/CD38−) phenotype and multipotent colony forming capacity in vitro.
Intrapancreatic transplantation of ALDHlo cells from DEAB-treated conditions reduced hyperglycemia in STZ-treated NOD/SCID mice
We have shown that ex vivo expansion of UCB Lin− ALDHhi cells rapidly diminishes islet regenerative function as culture time was increased [14]. Therefore, we sought to utilize previously reported culture conditions that increase the number (twofold) of ALDHhi cells following 9-day expansion, compared to untreated conditions [22]. On day 10, STZ-treated, hyperglycemic (>15 mM) NOD/SCID mice received intrapancreatic transplantation (Fig. 3A) of PBS or 5.0 × 105 bulk cells harvested from either untreated or DEAB-treated conditions. Similar to PBS-injected controls, mice transplanted with untreated bulk cells remained hyperglycemic up to 42 days (Fig. 3B, C). Compared to PBS-injected controls, transplantation of DEAB-treated bulk cells reduced hyperglycemia on day 17 (P < 0.05); however, a sustained reduction of hyperglycemia was not observed at later time points out to day 42 (P = 0.17; Fig. 3B, C). In a recent publication, we also demonstrated that sustained reduction in hyperglycemia was only achieved when 6-day expanded cells were reselected for high ALDH-activity before intravenous transplantation [23]. Therefore, we reselected our expanded cell populations for high versus low ALDH-activity before direct intrapancreatic transplantation. Hyperglycemic mice received an intrapancreatic transplantation of 2.0 × 105 ALDHhi or ALDHlo cells harvested from either untreated or DEAB-treated conditions. ALDHhi cells harvested from either untreated or DEAB-treated conditions at 9 days were unable to reduce hyperglycemia, compared to PBS-injected controls (Fig. 3D, E). Interestingly, the ALDHlo cell subset from DEAB-treated conditions significantly reduced hyperglycemia at days 14, 17, and 28 (P < 0.05; Fig. 3D). More importantly, hyperglycemia was significantly reduced by measurement of area under the curve (P < 0.05; Fig. 3E), compared to PBS-injected controls. After performing glucose tolerance tests 24-h before euthanasia, mice transplanted with bulk, ALDHhi, or ALDHlo cells from either untreated or DEAB-treated conditions did not demonstrate an improved physiological response to i.p. injection of a glucose bolus (Supplementary Fig. S1). Thus, DEAB-treated conditions generated ALDHlo cells that modestly reduced resting glycemia levels in STZ-treated NOD/SCID mice following intrapancreatic transplantation. However, the reduction in hyperglycemia was modest, transient, and mice were unable to respond to glucose bolus in a physiological manner at a later time point.
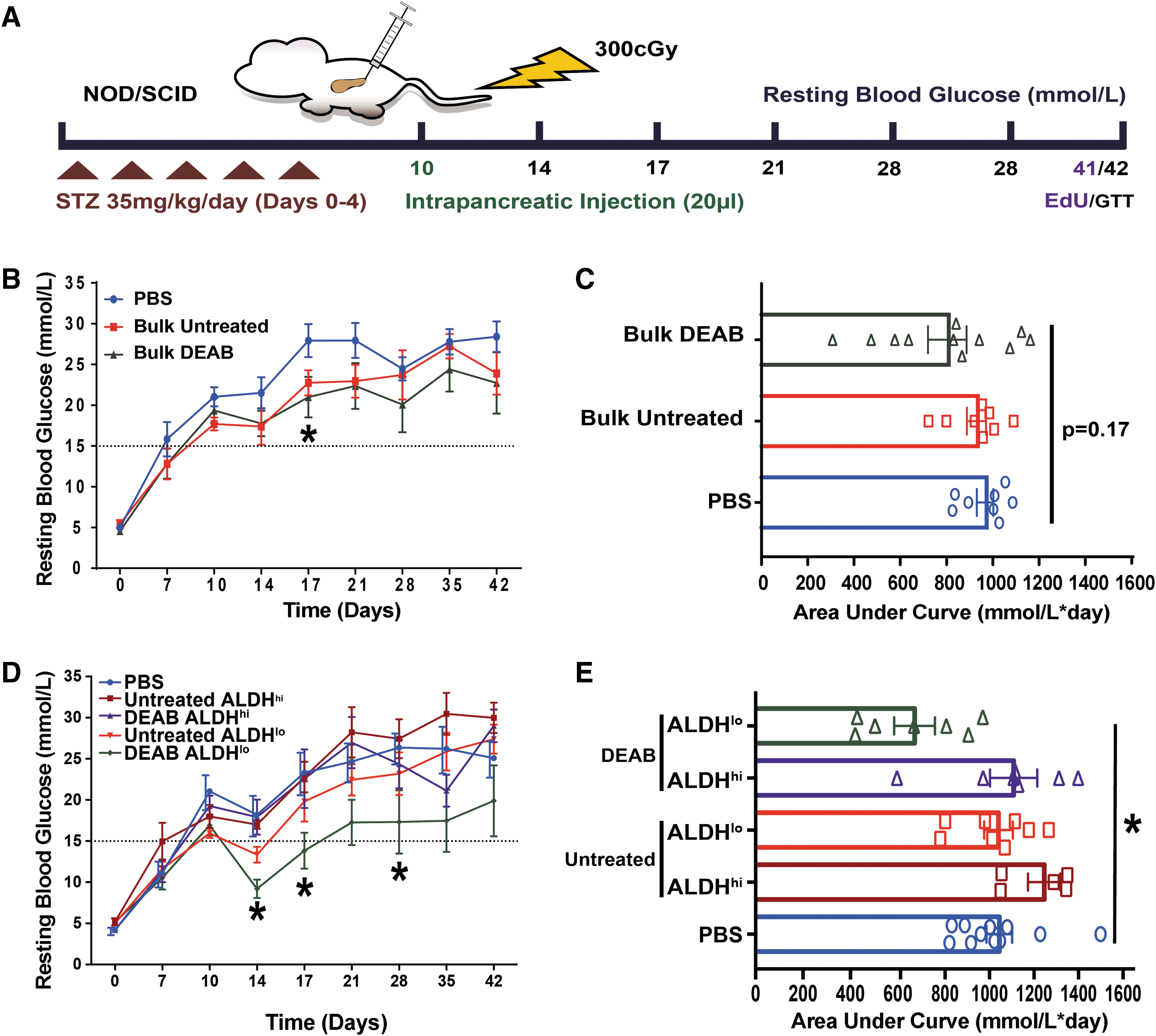
Intrapancreatic transplantation of reselected ALDHlo cells from DEAB-treated conditions reduced hyperglycemia in STZ-treated NOD/SCID mice.
Intrapancreatic transplantation of expanded ALDHlo cells from DEAB-treated conditions increased islet number
To investigate the mechanisms by which expanded ALDHlo cells generated from DEAB-treated conditions modestly reduced hyperglycemia in NOD/SCID mice, pancreas sections were analyzed for islet size, number, and total β-cell mass (Fig. 4A–D). Histological analysis revealed a significant increase in the number of insulin+ islets in mice receiving intrapancreatic transplantation of DEAB-treated ALDHlo cells, compared to PBS-injected mice (*P < 0.05; Fig. 4B). In contrast, islet circumference remained comparable between all transplantation conditions (Fig. 4C). Ultimately, mice transplanted with DEAB-treated ALDHlo cells demonstrated an increased trend in total β cell mass, compared to PBS-injected controls (P = 0.14; Fig. 4D). Thus, expanded DEAB-treated ALDHlo cells did not robustly increase β cell mass and transplanted mice demonstrated a failed physiological response to a glucose challenge, despite modestly improved resting glycemic levels.
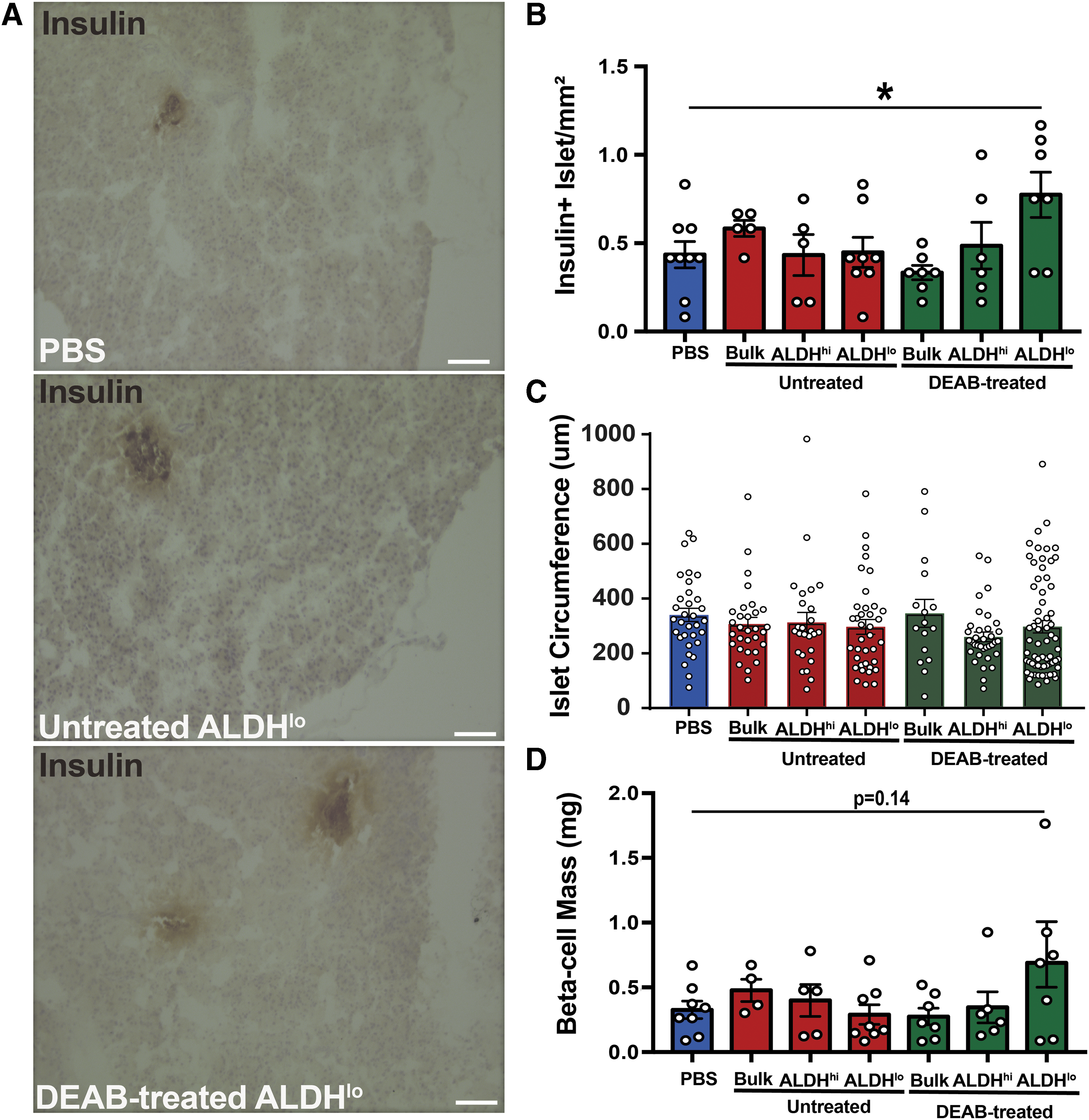
Mice transplanted with ALDHlo cells from DEAB-treated conditions showed modestly increased islet number and beta-cell mass.
Expanded ALDHlo cells increased intra-islet cell proliferation, but did not increase intra-islet vascularity or Nxk6.1+ expression
To better understand regenerative processes induced by intrapancreatic ALDHlo cell xenotransplantation, we assessed intra-islet cell proliferation in mice pulsed with EdU for 24 h before sacrifice at day 42. Pancreata transplanted with ALDHlo cells harvested from DEAB-treated conditions demonstrated an increase in the number of proliferating (EdU+) cells within islets at day 42 (Fig. 5A). Although increased proliferation was observed, intra-islet vascular density (CD31+ cells/Islet Area; Fig. 5B) and β cell identity marker expression (Nkx6.1; Fig. 5C) were equivalent to PBS-injected mice. Thus, indicating proliferating cells were likely not of an endothelial or β cell phenotype. HLA-A,B,C+ human cells were not detected in the pancreas of transplanted mice at day 42, despite detectable engraftment at day 14 (Supplementary Fig. S2). Collectively, ALDHlo cells from DEAB-treated conditions transiently engrafted the murine pancreas and modestly increased islet cell proliferation without significantly increasing islet-vascularity or functional β cell mass.
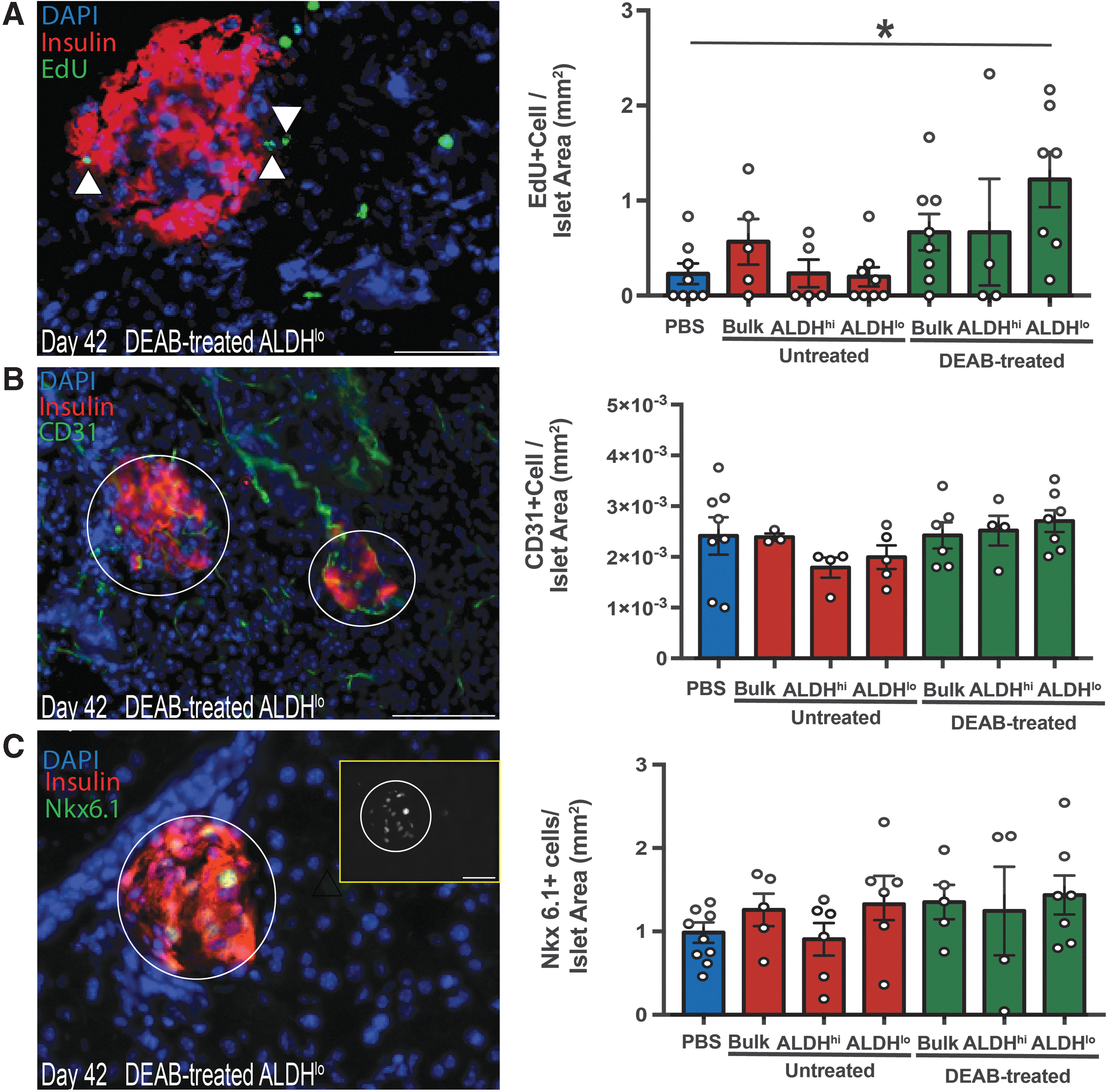
Intrapancreatic transplantation of ALDHlo cells from DEAB-treated conditions increased intra-islet cell proliferation.
Discussion
Restoration of β cell mass for the treatment of diabetes has recently become an intense focus of translational research, yet many of the mechanisms facilitating the regeneration of endogenous β cell mass remain elusive. For example, identifying hematopoietic cell subtypes that stimulate islet regenerative mechanisms has largely been unexplored. We have demonstrated selection of UCB Lin− ALDHhi cells enriched for a HPC population capable of reducing hyperglycemia in STZ-injected NOD/SCID mice [12]. Unfortunately, UCB ALDHhi cells are rare and expansion will be required to collect enough cells to potentially treat human subjects. Our recent work and this study demonstrate the challenge in expanding these cells with concurrent retention of islet regenerative capacity [14]. The selection of UCB Lin− ALDHhi cells biases HPC differentiation by active RA-induced differentiation under serum-free conditions that promoted the acquisition of a megakaryocyte phenotype (CD41+/CD42+) and loss of regenerative function (Fig. 6). On the other hand, we have recently reported that DEAB-induced inhibition of RA-production enhanced the expansion of HPC that retained vascular regenerative paracrine functions in vivo [22]. With these properties in mind, we initially hypothesized that reversible inhibition of ALDH-activity could enhance the expansion of UCB-derived hematopoietic progeny with islet regenerative function by preventing differentiation from an ALDHhi cell phenotype. Herein, we show the robust islet regenerative function of UCB Lin− ALDHhi cells is lost during ex vivo culture; however, beneficial functions were partially retained by inhibiting RA-induced differentiation and paradoxically reselecting for CD41+ enriched ALDHlo cells.
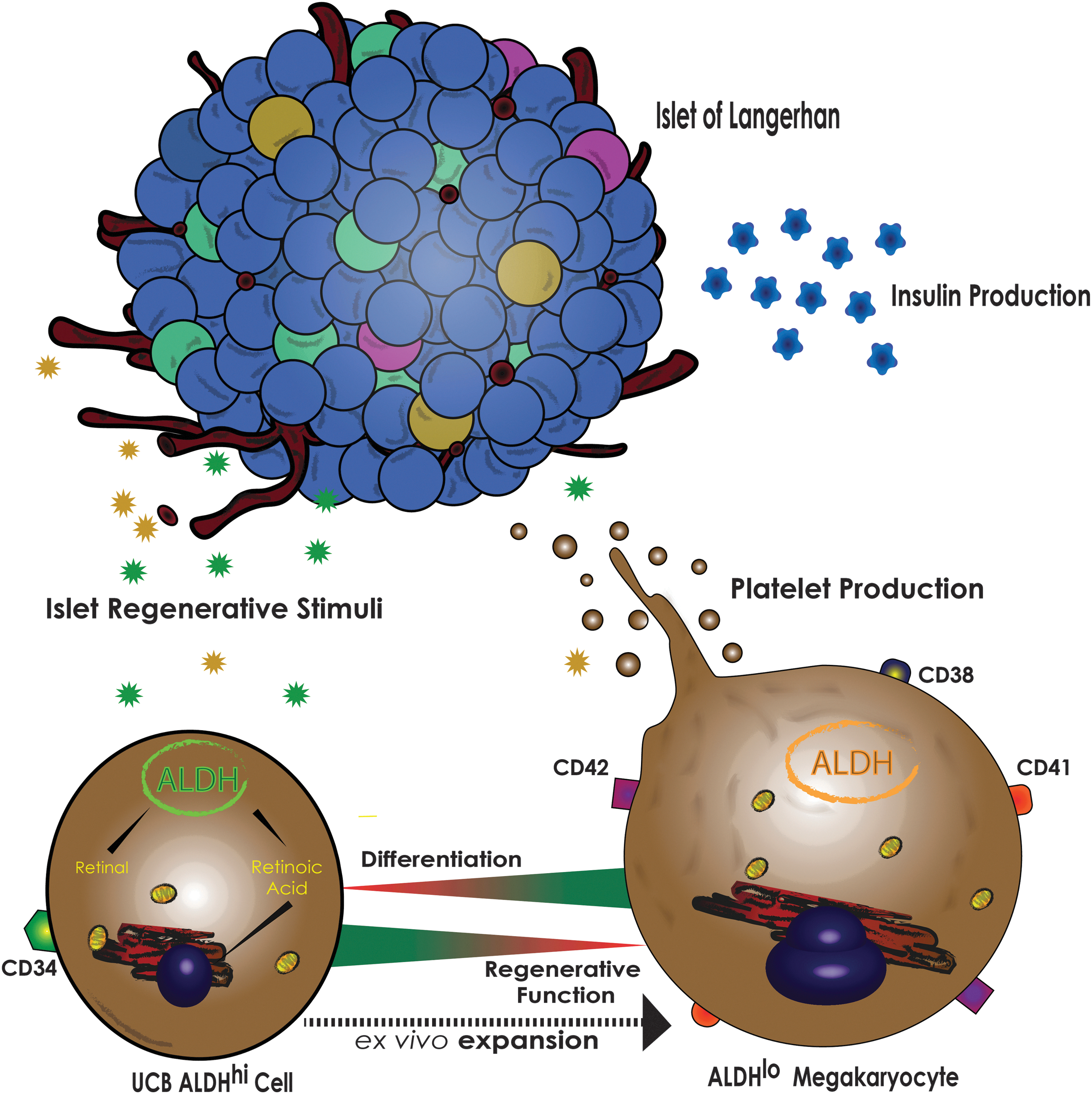
UCB Lin- ALDHhi cells acquire megakaryocyte phenotype during expansion that coincides with reduced islet regenerative function. UCB ALDHhi cells efficiently expand under serum-free conditions; however, UCB ALDHhi cells will rapidly lose an ALDHhi phenotype and acquire a committed megakaryocyte phenotype. The differentiation of UCB ALDHhi cells coincides with the loss of islet regenerative function.
The islet regenerative function of uncultured UCB Lin− ALDHhi cells was rapidly lost under clinically applicable, hematopoietic selective serum-free culture conditions. With this in mind, we first sought to determine whether the megakaryocyte content of uncultured UCB Lin− ALDHhi cells could provide help elucidate the phenotype of an islet regenerative cell population, as expanded cells are largely committed to the megakaryocyte lineage. CD41+/CD42+ cell content was equivalent between uncultured UCB Lin− cells, or Lin− ALDHhi or Lin− ALDHlo cell subsets. These results provided indirect evidence that CD41+/CD42+ megakaryocyte content likely played a minimal role in the islet regenerative function of transplanted uncultured UCB Lin− ALDHhi cells and the enrichment of a megakaryocyte phenotype is a result of subsequent culture conditions. Interestingly, Gentry et al. previously reported that the selection of ALDHhi cells enriched for megakaryocyte progenitors by >2000-fold, compared to the ALDHlo subpopulation [30]. We attributed our conflicting results observed in this study to our use of a lineage-depletion cocktail, which does not deplete cells of the megakaryocyte lineage from UCB MNC. Furthermore, megakaryoblast and megakaryocyte phenotype in UCB cells was assessed by flow cytometry in our study, as opposed to the use of a clonogenic assay that would assess the megakaryocyte potential of selected subpopulations. Nonetheless, our results corroborate the findings by Gentry et al., such that UCB ALDHhi cells robustly generated cells of the megakaryocyte lineage in vitro.
Megakaryocytes are large cells (>50 μm) that primarily reside within the bone marrow [31] or lungs [32] and are derived from a common megakaryocyte/erythroid myeloid progenitor cell [33]. Considering our culture conditions are supplemented with TPO, a known driver of megakaryopoiesis and paradoxically required for effective progenitor cell expansion, it is likely that the expansion of CD41+/CD42+ cells from ALDHhi cells can be attributed to active TPO-signaling. Future research would need to determine the role exogenous TPO and/or extracellular matrix molecules have on UCB ALDHhi cell differentiation toward a CD41+/CD42+ phenotype, an area that has been extensively studied with other HPC subpopulations [34 –36]. Moreover, TPO produced by cells of the megakaryocyte lineage could signal in an autocrine fashion to stimulate the expression of CD41 and CD42 [36]. Considering that >60% of freshly isolated UCB Lin− ALDHhi cells express CD41, we propose that UCB Lin− ALDHhi cells are primed to respond to drivers of differentiation, including the RA- and TPO-signaling pathways. Collectively, the expansion of ALDHhi cells and modulation of megakaryopoiesis in vitro is complex and future studies would need to consider several parameters including (1) progenitor cell purification, (2) culture conditions, and (3) culture duration.
For nearly 2 decades, high ALDH-activity has served as an enzymatic marker to identify and enrich for hematopoietic progenitor subtypes in fetal and adult tissue [37 –39]. Our recent publication demonstrated DEAB supplementation and subsequent inhibition of RA-production enriched for ALDHhi cells progeny (>2-fold) during UCB HPC expansion [24], leading to a >20-fold increase in viable ALDHhi cell number. Supporting our previous studies, we report that CD34+/CD38− cell surface expression was enriched within the ALDHhi cell subset following expansion [23]; moreover, reselected ALDHhi cells demonstrated enhanced multipotent hematopoietic colony formation, compared to ALDHlo cells [23]. Importantly, ALDHhi cells reselected from DEAB-treated conditions showed enhanced hematopoietic colony formation, compared to untreated conditions. Considering CD34+/CD38− enrichment within the ALDHhi subset was comparable between untreated and DEAB-treated conditions, our results support previous reports that high ALDH-activity further enriches for multipotent HPC from the CD34+/CD38− subpopulation [29]. These results indicate that the loss of high ALDH-activity coincides with the loss of multipotent progenitor capacity and hematopoietic colony formation can be retained by temporarily inhibiting RA-production in vitro. The loss of ALDH-activity has been associated with HPC differentiation and here we add that the ALDHlo cell subsets were enriched for a CD41+/CD42+ megakaryocyte phenotype (>50%) following ex vivo expansion. On the other hand, ALDHhi cells were enriched for CD41+/CD42− megakaryoblasts compared to ALDHlo cells. Therefore, we propose a model where high ALDH-activity is gradually diminished during culture as uncommitted multipotent myeloid progenitors lose a primitive CD34+/CD38− phenotype and transition toward a CD41+/CD42− megakaryoblast phenotype, before commitment toward a CD41+/CD42+ megakaryocyte phenotype and acquisition of CD38 expression (Fig. 6).
In the current study, we observed a modest improvement of glycemia levels in STZ-treated NOD/SCID mice receiving intrapancreatic transplantation of expanded cells harvested from DEAB-treated conditions. However, our histological analysis revealed minimal regeneration of β cell mass had occurred following direct transplantation of bulk expanded cells. These results lead us to predict the reselection of cells based on high versus low ALDH-activity would be required to purify regenerative cells with enhanced engraftment and/or migration. Intrapancreatic transplantation of expanded cells reselected for high versus low-activity surprisingly revealed that only ALDHlo cells harvested from DEAB-treated conditions (DEAB ALDHlo) reduced hyperglycemia. The capacity of ALDHlo cells harvested from DEAB-treated conditions to reduce hyperglycemia and improve islet function after intrapancreatic transplantation was highly variable, with 3 of 7 mice showing notable recovery. Although we observed increased islet number within the pancreas of mice transplanted with DEAB ALDHlo cells, the size of murine islets and total β cell mass were comparable between all conditions. Our previous reports demonstrated that transplanted fresh UCB ALDHhi cells transiently engrafted the pancreas, yet demonstrated sustained regeneration of β cell mass [12]. Similarly, we detected engraftment of transplanted expanded cell progeny at day 14 but not day 42; suggesting that expanded ALDHlo cell engraftment is also transient while islet regenerative function is modest. Nonetheless, DEAB-treated ALDHlo cells modestly reduced hyperglycemia, by increasing islet number through unidentified mechanisms. The results of this study contrast our previous HPC transplantation studies that reported increased β cell mass as a result of increased islet size not islet number [23]. We observed an increased rate of proliferating cells within murine islets at day 42 following direct transplantation of DEAB ALDHlo cells, however, it was concluded that EdU+ cells were not of an endothelial (CD31+) or β cell (Nkx6.1) phenotype. Collectively, we provided preliminary evidence that transplanted DEAB ALDHlo cells temporarily engraft the pancreas to support β cell function via unidentified and likely multifactorial mechanisms.
Highly vascularized networks within islets of Langerhans facilitate the release and transport of blood glucose regulating hormones, including insulin and glucagon, through systemic circulation [40]. Seneviratne et al. demonstrated increased intra-islet vascularity islet size and β cell mass after intravenous transplantation of 6-day expanded ALDHhi HPC [23]. Considering DEAB-treated cells robustly stimulate angiogenesis in the ischemic hindlimb, we investigated whether islet vascularity was increased following intrapancreatic transplantation; in return increasing the efficiency of systemic insulin delivery. Immunofluorescent analysis revealed intra-islet vascularity was equivalent between all transplantation conditions. This was anticipated, considering islet size was not increased following DEAB-treated ALDHlo cell transplantation. These collective results suggest that intra-islet vascularity may increase only when robust recovery β cell mass occurs. Previous studies have demonstrated a “symbiotic” relationship between endothelial cells and pancreatic β cells [41,42]. Specifically, the proliferation and survival of β cells was dependent on paracrine signaling from the neighboring vascular network. In this study, DEAB ALDHlo cell transplantation increased islet number and not size. Therefore, we suggest the modest reduction of hyperglycemia was likely a result of increasing the number of endogenous islets capable of releasing insulin into systemic circulation. We propose that DEAB ALDHlo cells likely provided unidentified supportive stimuli to surviving islets following STZ-induced β cell ablation. Nonetheless, identification of these stimuli will require future proteomic studies using cell populations with more robust and consistent islet regenerative function in vivo.
In summary, we provide preliminary evidence that expanding ALDHhi cells while retaining islet regenerative function is complex and the loss of islet regenerative function is likely multifactorial. Specifically, we have identified several important findings that may benefit future research aiming to expand HPC ex vivo for therapeutic applications. These include results such as the following: (1) FACS purification of UCB Lin− MNC based on ALDH-activity does not enrich for megakaryocytes, (2) inhibition of RA-production generates multipotent ALDHhi cells in vitro, and (3) retainment of a primitive ALDHhi phenotype does not coincide with islet regenerative function following intrapancreatic transplantation. Collectively, these results indicate that cellular therapies using expanded HPC to accelerate endogenous islet regeneration require further studies to prevent the loss of islet proliferative and vascularizing function.
Footnotes
Acknowledgments
We thank Kristin Chadwick at the London Regional Flow Cytometry Facility for cell sorting. This work was supported by an operating grant from the Canadian Institute of Health Research (CIHR) (MOP# 378189) and Juvenile Diabetes Research Foundation (2-SRA-2015-60-Q-R).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.