
Abstract
Adult tissues are thought to harbor two populations of “dormant” and “actively dividing” stem cells. Quiescent stem cells undergo rare asymmetric cell divisions (ACDs) through which they self-renew and give rise to tissue-committed “progenitors” of distinct fate and “progenitors” in turn undergo symmetric cell divisions (SCDs) and clonal expansion. However, quiescent stem cells have not been demonstrated in adult tissues such as skin, testis, liver, and brain. After surgical removal of part of liver and pancreas—adult differentiated cells divide and regenerate and a possible role of stem cells remains doubtful. Long-term repopulating hematopoietic stem cells are quiescent in nature but ACD has not been convincingly demonstrated even among them. Attempts by various groups to identify a common stemness program that ensures self-renewal among different kinds of stem cells have also remained futile. Uncontrolled self-renewal and compromised differentiation of stem cells possibly initiate leukemia/cancer, but the identity of leukemic stem cells and whether cancer stem cells arise by epithelial–mesenchymal transition (EMT) in solid tumors are all open-ended questions that need greater clarity. Acceptance of the presence of very small embryonic-like stem cells (VSELs) in adult tissues could clarify several of these existing dilemmas in the field. Data are compiled showing that VSELs undergo ACD in the hematopoietic system, testis, ovary, uterus, and pancreas, whereas tissue-committed progenitors undergo SCD and clonal expansion. VSELs possess similar overlapping stemness program as in embryonic stem cells, embryonic carcinoma cells, embryonic germ cells, induced pluripotent stem cells, and primordial germ cells. VSELs and leukemic and cancer cells express overlapping embryonic markers. Uncontrolled proliferation of VSELs and compromised differentiation possibly initiate leukemia. Process of EMT and initiation of solid tumor from VSELs (located among the epithelial cells) are indeed two distinct and parallel events. To conclude, VSELs provide explanation to several confounding aspects of adult stem cell biology.
Introduction
P
Two articles published simultaneously in 2017 in New England Journal of Medicine raise serious questions on the potential of both ASCs and iPS cells. Kuriyan et al. [3] reported that patients with age-related macular degeneration became blind when injected autologous adipose tissue-derived stem cells. A clinical trial was stopped after iPS-derived retinal progenitors were transplanted in a patient with age-related macular degeneration for safety reasons [4]. It was stressed that much more basic research is required before rushing to the clinics [5].
Despite studies showing that allogeneic iPS-derived retinal pigment epithelial (RPE) cells show immune response and graft failure in animal models [6,7], Japanese scientists have obtained permission to undertake a clinical study involving transplantation of a thin sheet of 1 million cardiac cells derived from allogeneic iPS cells to regenerate the heart [8], but they still need to demonstrate their claims [9]. The animal study based on which they got approval to undertake the clinical study shows that the transplanted cells do not integrate in the heart but only provide a trophic support. Mesenchymal cells are the ideal cells to provide trophic support. Then why do they prefer iPS cells to provide trophic support to the damaged cardiac tissue? This was recently discussed [10]. A recent study using RPE cells obtained by differentiation of hES cells also showed no efficacy on transplantation in patients with macular degeneration [11].
Articles published in 2018 are discussing the very definition of ASCs and have raised doubts regarding the existence and role (if any) of stem cells during regeneration of tissues such as the liver and pancreas [12]. This existing confusion in the field points to the fact how little we understand basic stem cell biology and it will be a long time before stem cells move from bench to bedside. The present review article is an attempt to re-examine and discuss in depth the hallmark property of asymmetric cell division (ACD) of stem cells through which they self-renew and give rise to “progenitors” and whether the normal tissue stem cells have any role in cancer initiation.
Do Adult Tissues Harbor “Dormant” and “Active” Stem Cells?
Earlier it was suggested that adult tissues harbor two populations of quiescent and actively dividing stem cells [13,14]. However, recent emerging literature expresses doubt on this concept of ASC biology [12,15] since quiescent stem cells have not been reported in skin, gut epithelium, intestine, liver, lung, mammary gland, and testes. Confusion exists on the presence of quiescent stem cells in various adult tissues and whether they have a role during regeneration to regain homeostasis after severe loss of function.
Skin epidermal cells have huge power to expand in culture and have been successfully used to treat burns wound. However, the true identity of epidermal quiescent stem cells is not yet known. Similarly, the liver possesses huge potential to regenerate after hepatectomy, but possibly the existing/surviving differentiated cells divide and regenerate and the liver does not harbor stem cells. Whether stem cells exist in adult tissues such as the pancreas and kidney also remains doubtful. Actively dividing stem cells undergoing symmetric cell divisions (SCDs) have been reported in the testes, stomach glands, and intestinal crypts, but quiescent stem cells remain elusive in these tissues. Clevers and Watts [12] suggested that ASCs should be defined based on their function rather than their phenotype because of the disparity that exists among different tissues and the variable role played by stem cells to regenerate damaged tissues.
The only liquid organ—hematopoietic system—possesses the classical hematopoietic stem cells (HSCs) that are rare, quiescent, and remain unidirectional. The majority of long-term HSCs (LT-HSCs) are in G0 (resting, quiescent) state but get regularly recruited in cell cycle under steady state and possibly switch to active state under stress conditions. Maintenance and restoration of bone marrow-derived blood cells is achieved by a discrete, transplantable entity—the HSC and this approach of bone marrow transplantation (BMT) is a method of standard care in the clinics. However, we are well aware that a purified population of HSCs is never transplanted during BMT. Rather, the mononuclear cells get transplanted, which are expected to contain HSCs. It is rather intriguing that for describing the stem cell properties, quiescence, and for the coexistence of LT-HSCs and short-term HSCs (ST-HSCs), a very old article by Cheshier et al. [16] was quoted by Clevers and Watt [12]. Has no further significant advancement been made in the field since 1999 and how do LT-HSCs and ST-HSCs differ from each other? The available evidence that HSCs undergo ACDs to self-renew is also not very strong since in the diagrammatic representation in the reviews [12,17], the size of the HSCs that self-renew and their immediate progeny appears similar although they supposedly arise by ACD. Whereas it is expected that the two cells that arise by ACD should be of unequal size, have distinct fate, and also should exhibit distinct proliferation kinetics [18].
Based on the above discussion, it seems that stem cell biology is still not well understood in any of the adult tissues, including the hematopoietic system. We need further clarity on how to define ASCs and differentiate them from lineage-restricted progenitors. The term “lineages” is loosely used to describe different types of blood cells that arise by differentiation of HSCs, whereas it strictly implies to the three germ layers, including the ectoderm, mesoderm, and endoderm. Based on the outcome of global efforts to use autologous HSCs to regenerate other tissues, it is very evident that HSCs have failed to cross boundaries and show any potential to differentiate into various lineages—rather they are lineage restricted and have the ability to only differentiate into different types of blood cells. Thus, it is best to describe them as tissue-committed progenitors rather than stem cells.
Integrating Very Small Embryonic-Like Stem Cells with ASCs to Better Understand Stem Cell Biology
It has been proposed that primordial germ cells (PGCs) survive in few numbers in various adult tissues as very small embryonic-like stem cells (VSELs) [19]. VSELs are relatively quiescent in nature [20 –22] and serve as a backup pool to give rise to ASCs throughout life, thus maintaining homeostasis. These stem cells were recently reviewed [16,23 –26] and differentiate into all the three germ layers, germ cells, and also give rise to the HSCs in vitro [27 –31]. They remain poorly accepted by the scientific community because of the controversies raised while attempts were made to study them by flow cytometry methods [32]. The queries raised were explained [33] and now methods have evolved to demonstrate their presence even after eliminating the use of flow cytometry [34]. Emerging data also show that it may be possible to expand them in vitro [19,27].
Thus, VSELs express pluripotent markers, show ability to differentiate into three germ layers, and can also be expanded in vitro. It is high time to integrate them into the existing understanding of ASC biology. VSELs have been reported in several adult tissues, including the liver and pancreas [35,36], and have a crucial role to play during regeneration after partial pancreatectomy [37,38]. They remain elusive due to their scarce nature and very small size. Thus, rather than adult somatic cells having a role in regeneration of tissues such as the liver and pancreas as discussed by Clevers and Watt [12], it is the VSELs that bring about regeneration.
Bhartiya [39] discussed that the true stem cells in adult tissues are the relatively quiescent and pluripotent VSELs that give rise to tissue-committed and lineage-restricted progenitors, including HSCs, spermatogonial stem cells (SSCs), neural stem cells (NSCs), and mesenchymal stem/stromal cells. This newly emerging understanding calls for a huge paradigm shift in the current understanding of stem cell biology in adult tissues and, as discussed by Smadja [40], their presence could explain controversial data that have emerged over the last two decades showing the ability of HSCs to cross boundaries and differentiate into multiple cell types.
“Stemness” Signature of Stem Cells
Several groups attempted to identify a core transcriptional program of “stemness” by comparing microarray data on embryonic stem (ES) and ASCs and cancer cells, but a set of common “stemness” genes still remains elusive. It was expected that a common stemness program will be shared by different types of stem cells especially genes regulating self-renewal, which is a hallmark feature of the stem cells. The stemness program is similar in mouse and human ES cells and overlaps with embryonic carcinoma (EC) cells. However, the ASCs tend to cluster away from this stemness program and rather have their own distinct subset of genes [41]. Earlier attempts in 2002–2003 by leading stem cell groups to detect a “stemness” gene profile by comparing transcriptome of ES and ASCs have remained futile [42]. Various reasons may have led to this confusion in the field such as (1) different stem cells may have distinct mechanisms for self-renewal, (2) technical variations in stem cell isolation, (3) purity of cells used for microarray, or (4) data analysis methods may vary. Shin et al. [43] have carried out global transcriptome analysis on 20 purified VSELs (Sca-1+Lin−CD45−) and hES cells and reported a distinct overlap between them. Interestingly, this group also reported none of the stemness genes expressed in complementary DNA isolated from HSCs (Sca-1+Lin−CD45+) and bone marrow (BM) mononuclear cells.
Thus, it is likely that various groups failed to find a common stem cell program since they were comparing apples and oranges. Rather, the pluripotent stem cell program remains conserved across different kinds of stem cells, including mouse ES cells, human EC and ES cells, PGCs (embryonic germ cells), VSELs, and cancer cells. HSCs, NSCs, and others are indeed adult tissue-committed stem cells and it will be more appropriate to term them as tissue-committed “progenitors” [39].
ASCs and Property of Self-Renewal
The lifelong, mutation-free survival of ASCs is due to the unique property of ACD, through which the stem cells self-renew and also give rise to “progenitor” cells that undergo rapid SCDs and clonal expansion followed by differentiation into tissue-specific cell types to maintain tissue homeostasis. As a result, stem cells remain relatively quiescent and give rise by ACD to progenitors that undergo SCD and clonal expansion to expand and produce billions of sperm/blood cells in the testes and the hematopoietic system (Fig. 1). The two cells undergoing ACD are expected to be of unequal size and have distinct gene expression pattern and distinct proliferation kinetics [18]. A careful search of published literature suggests that convincing evidence supporting ACD (hallmark feature of stem cells) is still lacking for various ASCs, including the most extensively studied HSCs and SSCs. During hematopoiesis, initial separation of lymphomyeloid and erythromyeloid lineages is realized through ACD but whether self-renewing ACD occurs among HSCs has not yet been convincingly demonstrated. The LT-HSCs, ST-HSCs, and the progenitors are presumably of similar size [12,17,44]. LT-HSCs can be distinguished and purified on the basis of their markers as Lin−/Sca-1+/Ckit+/CD34−/Slamf1+(CD150+ and CD48+) in mice and as Lin−/CD34−/CD38−/CD90+/CD45RA− in humans along with low retention of DNA dyes. Differential expression of CD34, CD48, CD49b, CD150, Flt3, Sca-1, and other markers such as CD127 and FcgR1, as well as staining with DNA dyes, has achieved separation of LT-HSCs and ST-HSCs [45,46]. Few studies have shown that HSCs undergo ACD based on the two cells being of different fates (unequal expression of few markers). But if the definition of ACD becomes more restrictive—cells of different fates should also be qualitatively different and exhibit different proliferation kinetics–such ACD has not yet been demonstrated for HSCs. A cell undergoing ACD is expected to exhibit asymmetric distribution of proteins that is assigned intrinsically rather than by extrinsic signals. Table 1 lists various studies reporting ACD among HSCs using different markers [18,47 –51], however as evident, a consensus still needs to emerge. Similarly, recent efforts by Shinohara's group to identify a marker that selectively detects ACD in testicular stem cells failed as CDY1 dye was also found to stain stem cells undergoing SCD [53].

ACD, asymmetric cell division; eGFP, enhanced green fluorescent protein; HSCs, hematopoietic stem cells; SCD, symmetric cell division.
Our group has recently reported ACD in the stem cells isolated from adult tissues, including mouse BM and testis, and in sheep ovary [54 –56]. A remarkable similarity of ACD in various tissues is intriguing. It becomes apparent that VSELs undergo ACD to give rise to HSCs, SSCs, and so on. Thus, in the true sense, VSELs are the stem cells in various adult tissues that undergo self-renewal and undergo ACD to give rise to the HSCs/SSCs/NSCs, which are tissue-specific progenitors or transit-amplifying cells that further undergo SCD and clonal expansion before differentiation into tissue-specific cell types. Pluripotent VSELs express nuclear OCT-4A, whereas we have observed that in various tissues, the ASCs express cytoplasmic OCT-4B under steady state [23]. This observation suggests that the ASCs are indeed immediate progenitors (no longer pluripotent) that arise from the pluripotent VSELs. Similar nuclear and cytoplasmic OCT-4 localization (reflecting spliced variants OCT-4A and OCT-4B) in pluripotent and nonpluripotent human PGCs has also been reported by others [56]. They proposed that OCT-4A in PGCs either translocates to the cytoplasm or is attenuated there possibly for degradation and that the significance of cytoplasmic OCT-4 is unknown, but is notably coincident with major global epigenetic changes. We have recently demonstrated ACD among mouse BM stem/progenitors cells by showing difference in size of the two dividing cells and differential expression of nuclear OCT-4A in the smaller VSEL and NUMB (a differentiation marker) in the slightly bigger cell [54].
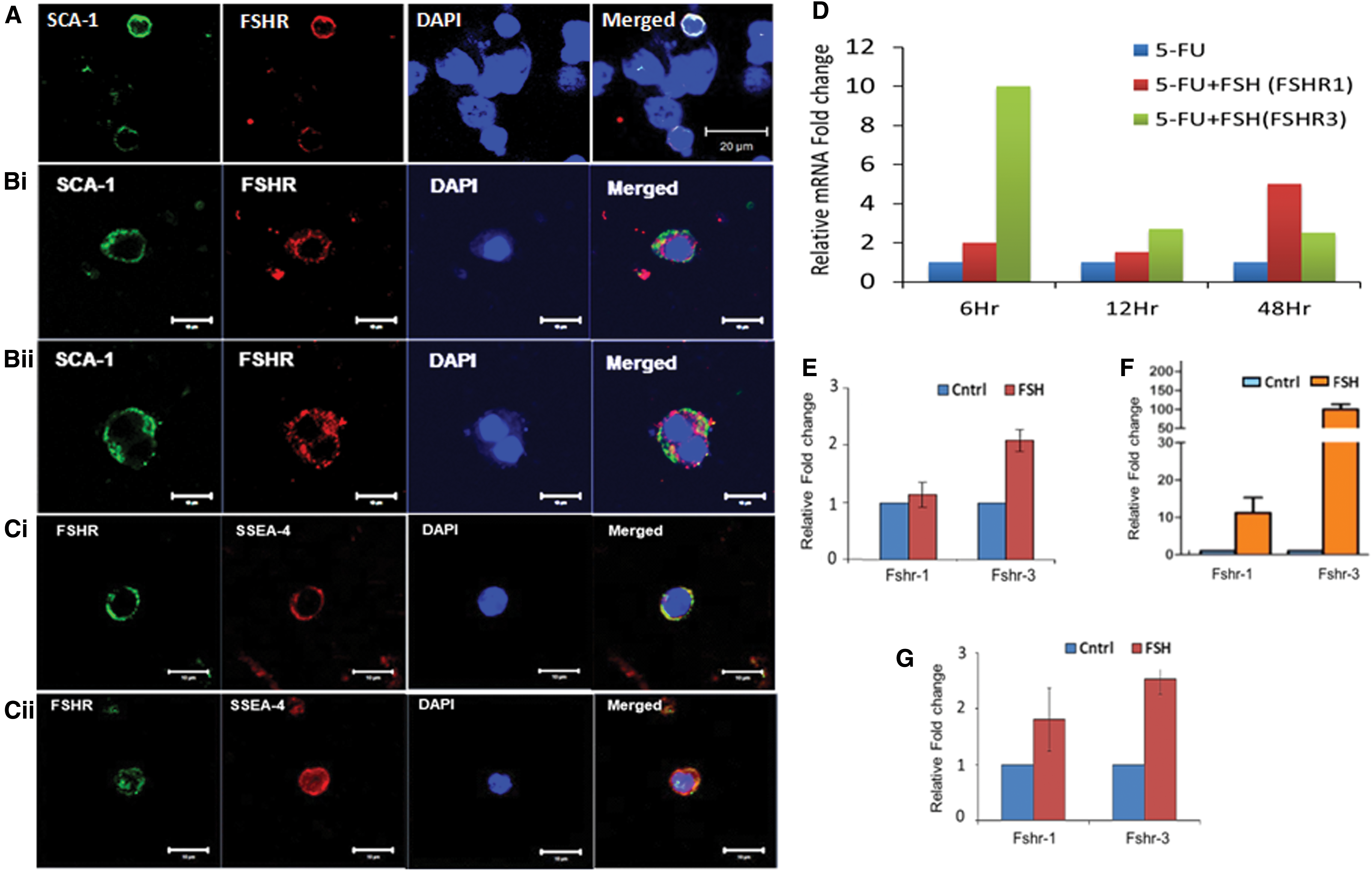
Stem/progenitor cells residing in various tissues, including the ovary, testis, and BM, express receptors for pituitary gonadotropin follicle stimulating hormone (FSH) [56,58,59]. Similar expression of sex and pituitary gonadotropin hormone receptors has been reported on hematopoietic stem/progenitor cells by other groups as well [60,61]. Our results demonstrate for the first time that the stem/progenitor cells are regulated by FSH to undergo ACD/SCD and clonal expansion (Fig. 2). Further FSH action is mediated via alternately spliced growth factor type one isoform FSHR3 [56,58] (Fig. 2D–G) via MAPK/ERK pathway rather than the canonical FSHR1 via cAMP as reviewed earlier [62].
ACD, SCD, and clonal expansion of stem cells in the mouse BM
Different approaches have been used in the past to study replicative behavior of HSCs, including serum-free single-cell culture followed by transplantation of cultured cells into lethally irradiated mice or time-lapse imaging of single cells in culture. Factors that may induce self-renewal of stem cells are also not well known. Nakauchi et al. [63] reported that stem cell factor (SCF) + interleukin 3 (IL-3)-3, SCF + IL-6, or SCF + IL-11 + FLT3 ligand (FL) were not very effective to enhance self-renewal of HSCs. It has also been reported that the VSELs survive in the BM of mice exposed to total body irradiation, whereas HSCs are destroyed [64].
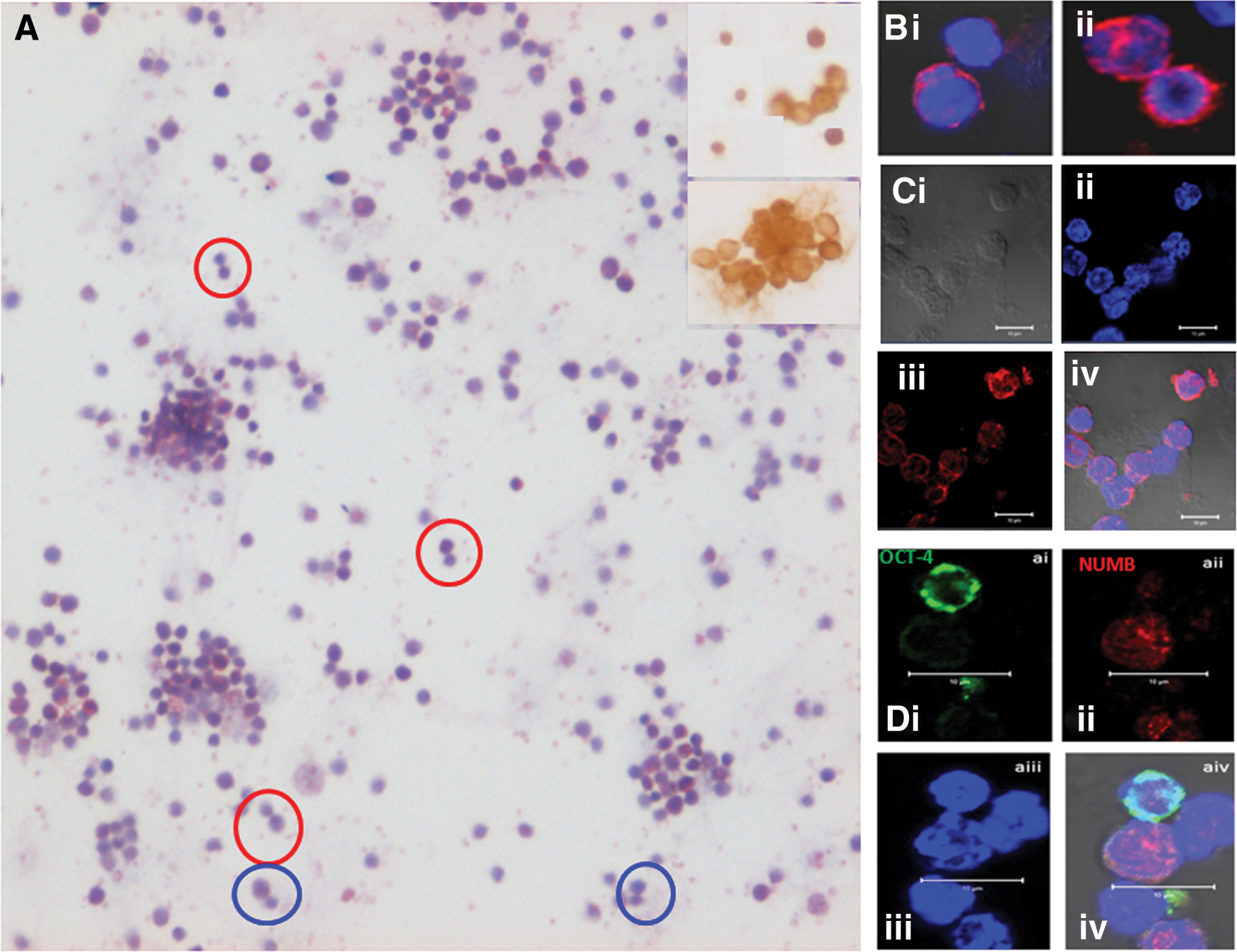
We studied the proliferative activity of stem/progenitors in the BM of chemoablated mouse. As reported earlier [54], mice were treated with 5-fluorouracil (5-FU, 150 mg/kg) that depleted the BM of a large number of HSCs, whereas the VSELs survived and were increased in numbers on day 4 with increased BrdU uptake—in an attempt to regain tissue homeostasis. BM cells were collected on day 4 after 5-FU treatment and used to make smears after lyses of red blood cells. Care was taken to always spin the cells at 1,000 g since VSELs pellet down at a higher speed. The smear was enriched for stem cells, and various stem cell divisions, including ACD, SCD, and clonal expansion, were clearly visualized in hematoxylin and eosin (H&E)-stained cells (Fig. 3A). The stem cells were positive for OCT-4 that showed nuclear to cytoplasmic expression (Fig. 3A, insert). CD45− VSELs underwent ACD to give rise to CD45+ HSCs (Fig. 3Bi) and SCD was evident among CD45+ HSCs (Fig. 3ii). Also, the CD45+ cells underwent clonal expansion with incomplete cytokinesis (Fig. 3Ci–iv). Coexpression studies for OCT-4 (pluripotent marker) and NUMB (differentiation marker) showed dividing doublets produced as a result of ACD (Fig. 3Di–iv). Differential expression of OCT-4A and NUMB in the stem cells has been published earlier [54]. The VSELs and HSCs have different proliferation kinetics and the differential expression of OCT-4 and NUMB was assigned intrinsically in response to 5-FU treatment. Murine SCA-1+ stem cells expressed receptors for follicle stimulating hormone (FSHR) and FSH treatment augmented stem cells possibly via FSHR3 (Fig. 2).
Asymmetric stem cell divisions in the mouse testes
Mice were treated with busulfan (25 mg/kg) that resulted in loss of sperm and germ cells in the testes, whereas the quiescent stem cells survive [56,65]. Chemoablated testes were enzymatically digested and cells from seminiferous tubules were cultured overnight in the presence of FSH [55]. ACD of small-sized VSELs gives rise to slightly bigger SSCs that, in turn, underwent SCD and clonal expansion to form spheres (Fig. 4A). FSH receptors were expressed on murine SCA-1 positive stem/progenitor testicular cells and quantitative real time-polymerase chain reaction data showed that this activation of stem cells by FSH was associated with selective upregulation of FSHR3 rather than the canonical FSHR1 (Fig. 2).
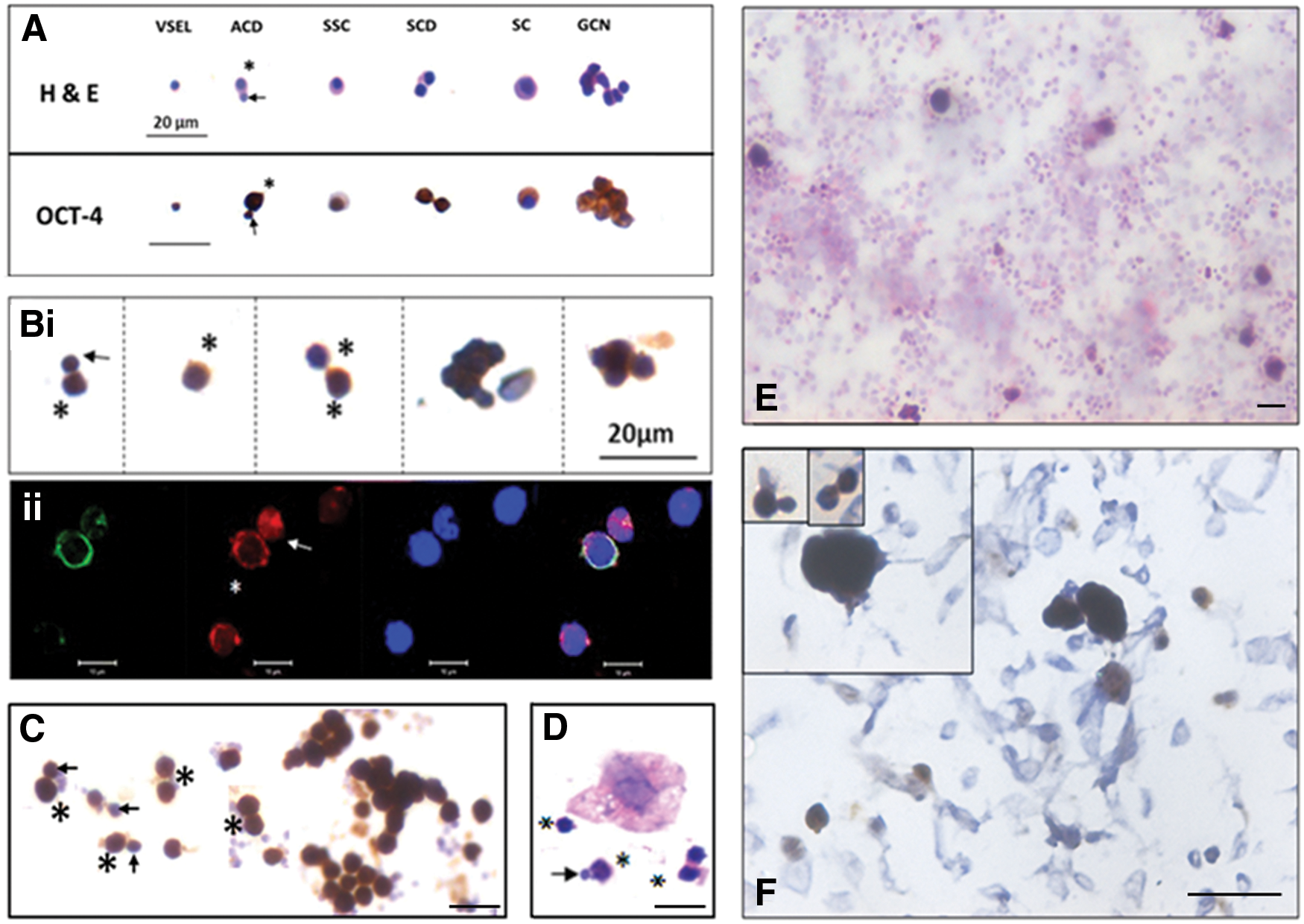
Asymmetric and symmetric divisions and clonal expansion in the testis, ovary, uterus, and pancreas.
Asymmetric stem cell divisions in sheep ovary
Two populations of stem cells, including small-sized, spherical VSELs and slightly bigger ovarian stem cells (OSCs), which are equivalent to SSCs in the testis, get enriched from sheep ovaries by gently scraping the ovary surface epithelium (OSE) [55,58,66]. Overnight culture in the presence of FSH showed stem cells undergoing ACD, SCD, and clonal expansion to form germ cell nests (Fig. 4Bi, E). Stem cells/germ cell nests expressed FSHR, whereas epithelial cells were distinctly negative for FSHR (Fig. 4F). This was associated with selective increase of FSHR3 (Fig. 2). The two cells undergoing ACD were of distinct size and VSELs expressed nuclear OCT-4A, whereas OSCs expressed NUMB (Fig. 4Bii).
Asymmetric stem cell divisions in the mouse uterus
Bilaterally ovariectomized mice were treated with FSH and uterine stem cells were enriched after enzymatic digestion. For this, the enzymatically digested cell suspension was first spun at 1,200 rpm, through which a majority of somatic cells pellet down. The supernatant was later spun at 3,000 rpm, through which stem cells settle down and are used to make smears. Events, including ACD, SCD, and clonal expansion to form clusters, were easily visualized (Fig. 4C). Figure 5 shows higher magnification images of uterine stem/progenitor cells undergoing ACD, SCD, and clonal expansion with incomplete cytokinesis.
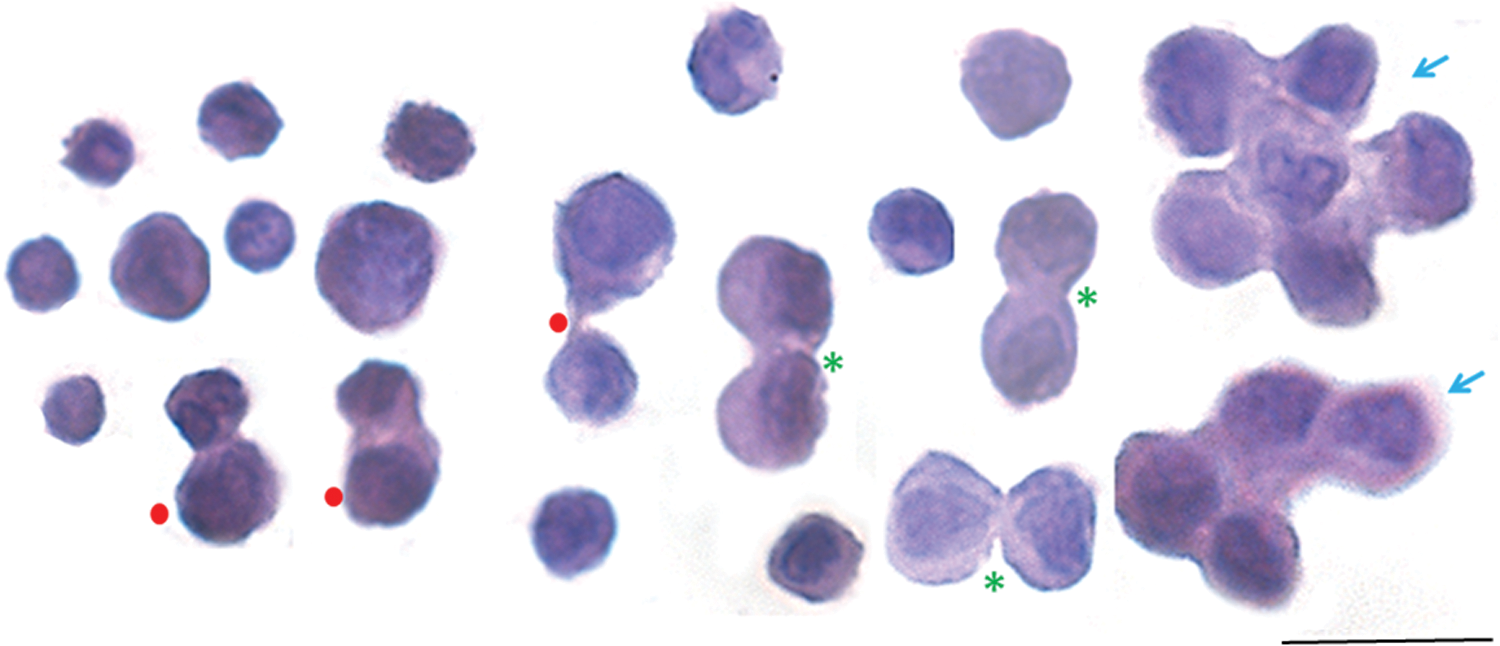
Higher magnification visualization of asymmetric and symmetric divisions and clonal expansion among mouse uterine stem cells. Cell smears were prepared from mouse uterine tissue after treatment with FSH (5-IU per day) and stained with H&E. Cell suspension obtained after enzymatic digestion was first spun at 1,000 rpm when the majority of somatic cells settle down. Stem cells were enriched by spinning the supernatant at a higher speed of 3,000 rpm. Spherical cells of two distinct sizes were clearly visible. Dividing doublets comprising cells of unequal (red solid circles) and equal (green asterisk) size were clearly visible along with cell spheres formed by clonal expansion with cytoplasmic continuity and incomplete cytokinesis (blue arrow). It is a composite image prepared by pasting together dividing cells from different fields. Scale bar = 20 μm. Color images available online at
Asymmetric stem cell divisions in the mouse pancreas
Adult mouse pancreas was processed to prepare smears as described above for the uterus. H&E-stained smears of cells obtained by spinning at 3,000 rpm are shown in Figure 4D, which shows both ACD and SCD [37].
Based on the results summarized above, VSELs and the tissue-specific progenitors (ASCs) that arise by ACD are of unequal size, show different gene expression, and also have different proliferation kinetics with VSELs mostly being in the G0 phase of cell cycle, whereas the progenitors undergo active proliferation. The stem/progenitor cells in adult tissues express FSHR and are activated by FSH via FSHR3 to undergo self-renewal. These results have huge implications at several fronts as discussed below. To conclude, “true” stem cells in adult tissues that undergo ACD are the VSELs, and in various tissues, they express FSHR and are activated by FSH treatment to undergo self-renewal. ASCs in the true sense are tissue-specific progenitors that arise by ACDs of VSELs and labeling them as “stem cells” is indeed a mistake that needs urgent correction.
What Exactly Are Cancer Stem Cells?
Understanding normal tissue stem cell biology is crucial to better understand cancer stem cells (CSCs). It is very likely that rather than the prevailing views of dedifferentiation/reprogramming of somatic cells to CSCs expressing pluripotent markers, cancers may arise due to uncontrolled proliferation or “self-renewal” of VSELs in adult tissues along with compromised differentiation [34,67]. CSCs may arise by the disturbance in ACD (self-renewal and differentiation) of the VSELs. CSCs undergo extensive self-renewal and lose the property to differentiate.
Identifying leukemia stem cells
The concept of CSCs was proposed four decades ago and created a lot of excitement as it could explain both initiation and recurrence [68,69]. It was suggested that similar to regeneration and maintenance of lifelong homeostasis, stem cells may also have a role in tumor growth. However, their existence and identity still remains a challenge [70]. Understanding normal stem cell biology is crucial to better understand CSCs. Virchow and Connheim in the IX century proposed that adult tissues contain dormant embryonic remnants “lost” during developmental organogenesis, which may give rise to malignancies [71]. Rather than the prevailing views of dedifferentiation/reprogramming of somatic cells, it may be that the tissue-resident VSELs possibly get transformed into CSCs, as was suggested for the first time by Ratajczak et al. [72] and recently discussed [34]. Thus, cancer could initiate as a result of blocked differentiation and uncontrolled proliferation of VSELs (embryonic remnants in adult tissues) and not due to dedifferentiation/reprogramming of ASCs in the body. VSELs and CSCs share several characteristics, including (1) both survive oncotherapy due to quiescent nature, (2) ability to migrate/mobilize (metastasis/restore homeostasis), (3) property of self-renewal, and (4) expression of embryonic markers (OCT-4A, NANOG, SOX2) and CXCR4, FSHR, and CD133.
Al-Hajj and Clarke [73] discussed that both HSCs and acute myeloid leukemia (AML) cells are CD34+CD38−, but HSCs could not give rise to AML since AML cells are Thy1−, whereas HSCs are Thy1+.They suggested that possibly AML arises from early downstream progenitors lacking Thy1 or Thy1 gets lost as a consequence of neoplastic transformation. VSELs are Thy1− and express pluripotent markers [36]. Recent publications have discussed a possible role of OCT-4 in leukemogenesis [74,75]. Thus, in addition to HSCs or their downstream progenitors, it is the more primitive nuclear OCT-4A+ VSELs that possibly initiate some types of leukemia. We have earlier shown using both human cord blood [76] and mouse BM [59] that VSELs express nuclear OCT-4A, whereas the immediate progenitors express cytoplasmic OCT-4B. It becomes crucial to understand the changes that cause VSELs to exit quiescence and undergo excessive proliferation leading to leukemia initiation. The niche definitely has a crucial role to play; altered VSEL behavior is due to an altered niche, and heterogeneity among leukemia patients is because of the extent to which the niche gets affected.
Identifying CSCs
A lot of confusion exists on the basic understanding of CSCs in solid tumors as well. Various epithelial tumors such as breast, prostate, ovarian, cervical, and pancreatic cancers express EMT markers [77] and it was initially suggested that epithelial cells acquire properties of CSCs through the activation of EMT program [78 –80]. Once mobile, cancer cells undergo metastases and travel through the blood stream to distant sites where they establish new tumors. Experimental induction of EMT in mammary epithelial cells generated cells with a CD44high/CD24low phenotype, considered to be a putative marker for breast CSCs [78]. Nieto et al. [81,82] cautioned that the contribution of EMT to tumor initiation requires more research. At the tumor-invasive front, cells are mesenchymal in nature, whereas the main tumor bulk remains largely epithelial. It is likely that CSC development and EMT occur in parallel rather than through the activation of the same pathway. Brabletz et al. [83] have also discussed the existing controversies in the field of CSCs and associated complexities. Similar to breast cancer, other solid tumors, including ovarian cancers, also arise in the epithelial cells along with EMT. It is well known that >90% of ovarian cancers are of epithelial origin and EMT may be involved in cancer initiation [84].
Rather than EMT leading to formation of CSCs in various kinds of epithelial tumors, including breast, prostate, ovarian, cervical, and pancreatic cancers, we suggest that these events possibly occur in parallel. Our earlier work on OSE cells have shown that the ovarian epithelial cells are interspersed with two populations of stem/progenitor cells, including VSELs and immediate progenitor OSCs [55,58,67,85]. These VSELs/OSCs that reside among the epithelial cells could initiate cancers (Fig. 1B) and this has been shown by other groups as well [86,87]. Rather than the epithelial cells, ovarian stem/progenitor cells and ovarian CSCs express FSHR (Fig. 4) and pluripotent markers including OCT-4 [88,89]. Under normal conditions, the epithelial cells undergo EMT followed by MET (mesenchymal–epithelial transition) to form pregranulosa cells that surround the oocytes arising by differentiation of stem cells leading to primordial follicle assembly. In cases of ovarian cancer, the VSELs get transformed into CSCs, whereas the epithelial cells (that provide a niche to the OSCs) rather than differentiating into granulosa cells form myofibroblasts (a distinct niche to the CSCs) by EMT that exist at the front of tumors (Fig. 1B). Similarly, Virant-Klun's group has also proposed a model describing how VSELs in ovarian cancers may have a role in promoting metastasis of ovarian cancer [87].
To conclude, VSELs are the possible primitive and pluripotent stem cells in the adult tissues that possibly get transformed into CSCs in both the hematopoietic system and also in various solid tumors. EMT does not lead to transformation of epithelial cells to CSCs, but EMT and tumor initiation from the VSELs occur simultaneously.
Conclusions, Challenges, Future Directions, and Perspective
Conclusions
VSELs are the most primitive and pluripotent stem cells that exist in various tissues, including the liver [35] and pancreas [37]. Thus, rather than suggesting that adult differentiated cells bring about liver regeneration [6], endogenous VSELs in adult tissues serve as a backup pool for tissue-committed stem cells and thereby maintain homeostasis and get activated to regenerate under stress.
VSELs share a similar stemness program to ES cells, EC cells, embryonic germ cells, PGCs, and iPS cells. It is expected that ASCs fall out of this as they do not undergo self-renewal and rather are lineage-restricted and tissue-committed stem cells.
VSELs are quiescent in nature with minimal cytoplasm and few organelles, including spherical mitochondria. Nuclear OCT-4A-positive VSELs give rise to slightly bigger “progenitors” with cytoplasmic OCT-4 B and low nucleocytoplasmic ratio. Chemoablation leads to enrichment of VSELs/progenitors and treating with FSH leads to ACD, SCD, and clonal expansion.
VSELs possibly get transformed into LSCs/CSCs and initiate leukemia/solid tumors since they share similar properties and marker expression. EMT occurs in parallel to tumor initiation and results in formation of myofibroblasts, which exist at the tumor front and facilitate metastasis. Tumor occurs due to uncontrolled proliferation of VSELs.
Challenges
A global consensus needs to emerge on the presence of VSELs in adult tissues. They have been struggling to get acknowledged over two decades [90]. They can easily be enriched from BM/peripheral blood/cord blood [76,91,92] without using flow cytometry. They can also be enriched from any solid tissues without using flow cytometry [23,91,92]. Flow cytometry is used to characterize these stem cells taking precaution to always spin the cells at 3,000 rpm rather than 1,000–1,200 rpm.
Future directions
Scientific community needs to replicate our findings and confirm the ability of VSELs in adult tissues to undergo self-renewal and ACD. How VSELs/progenitors and their niche cells get altered with age needs to be understood better.
How do the VSELs exit from quiescence to initiate tumors needs further research. Need to study epigenetic status of VSELs isolated from normal and tumor tissues.
Perspective
As suggested earlier by Parker [93], the scientific community needs to brainstorm the existence of VSELs. VSELs have not yet been studied for their translational potential as they were invariably discarded while processing autologous mononuclear cells from BM for cell therapy. They get affected with age resulting in increased incidence of various age-related diseases, including cancers.
Footnotes
Acknowledgments
The authors acknowledge the Indian Council of Medical Research, government of India, New Delhi, for financial support. They also acknowledge the help of Vaibhav Shinde with the art work. NIRRH article number: OTH/620/03-2018.
Author Disclosure Statement
No competing financial interests exist.