
Abstract
The regenerative capacity of the endometrium has been attributed to resident stem/progenitor cells. A number of stem/progenitor markers have been reported for human endometrial stem/progenitor cells; however, the lack of convenient markers in the mouse has made experimental investigation into endometrial regeneration difficult. We recently identified endometrial epithelial, endothelial, and immune cells, which express a reporter for the stem/progenitor marker, mouse telomerase reverse transcriptase (mTert). In this study, we investigate the expression pattern of a green fluorescent protein (GFP) reporter for mTert promoter activity (mTert-GFP) in endometrial regeneration following a menses-like event. mTert-GFP expression marks subepithelial populations of T cells and mature macrophages and may play a role in immune cell regulated repair. Clusters of mTert-GFP-positive epithelial cells were identified close to areas of reepithelialization and possibly highlight a role for mTert in the repair and regeneration of the endometrial epithelium.
Introduction
Human endometrium is a highly dynamic and regenerative tissue undergoing ∼400 cycles of proliferation, differentiation, shedding, and repair throughout a woman's reproductive lifetime. Postmenstrual repair is a tightly controlled process that relies on the coordination of multiple mechanisms to initially reduce blood loss, stabilize the tissue following tissue shedding, and promote repair and subsequent regeneration of the endometrium [1]. This cyclical repair and regeneration of the endometrium is essential for a successful pregnancy, but the dysregulation of this process may lead to endometrial pathologies such as cancer or even endometriosis [2].
The regenerative capacity of the human endometrium has been attributed to stem/progenitor cells [2]. Endometrial mesenchymal stem cells can be easily isolated from human endometrium using a single marker, sushi domain containing 2 (SUSD2) [3], while a novel marker for endometrial epithelial progenitors, N-cadherin, has recently been identified [4]. Genetic markers for mouse endometrial stem/progenitor cells remain largely elusive, thereby limiting our understanding of endometrial regeneration.
We recently identified mouse telomerase reverse transcriptase (mTert) as a marker of stem/progenitor activity in the intact mouse endometrium [5]. mTert is a stem cell marker in the intestine [6] and mTert-green fluorescent protein (GFP) expression identifies rare populations of epithelial and endothelial cells as well as a CD45+ leukocyte population in the mouse endometrium. Ovariectomy results in a decrease in mTert-GFP endometrial epithelial cells, suggesting that mTert expression is linked to estrogen-driven regenerative activity. Menstruation, postmenstrual repair, and early regeneration of the endometrium involve extensive reepithelialization, while circulating ovarian steroid hormones, estradiol and progesterone, are low [7,8]. In this study, we investigate the expression pattern of mTert-GFP+ cells during reepithelialization and remodeling of the endometrium in a mouse model of menses.
Methods
mTert reporter mice
mTert-GFP reporter mice on a C57BL/6J background were bred in the Monash Medical Centre Animal Facility and housed on a 12-h light/12-h dark cycle with access to normal chow ad libitum. Approval for all procedures used was obtained from the Monash Medical Centre Animal Ethics Committee A.
Mouse model of menstruation
mTert-GFP reporter mice of 8–12 weeks of age underwent a mouse model of menstruation as published previously [9]. Briefly, mice were ovariectomized (day 0) and then subjected to β-estradiol injections (100 μL, 1 μg/mL, days 7–9), a progesterone-secreting pellet was placed under the skin (500 ng/day, days 13–19), and the mice received maintenance doses of estradiol (100 μL, 50 ng/mL, days 13–15). On day 15, artificial decidualization was performed by transvaginal injection of 20 μL of sesame oil into the uterine horns using a nonsurgical embryo transfer device. Progesterone pellets were removed on day 19 to induce artificial menstruation. Uterine horns were collected on day 19 at the following time points: 0 h before breakdown (n = 8), 8 h during breakdown (n = 11), 24 h during repair (n = 11), and 48 h during remodeling (n = 11).
Flow cytometry
Fresh mTert-GFP uteri were dissected, finely minced, and enzymatically digested in 2 mL of 10 mg/mL collagenase Type I (Worthington Biochemical) in phosphate-buffered saline (PBS) with 10 mg/mL deoxyribonuclease type I (Worthington) at 37°C for 30 min on a rotator. Uteri from two wild-type C57BL/6J mice were used as GFP-negative control. Dissociated tissues were then filtered sequentially through 70 and 40 μm cell strainer. Resulting cell suspensions were diluted with 5 mL of 5% fetal bovine serum (FBS):PBS and centrifuged at 400 g for 5 min at 4°C. Cell pellets were then washed with a further 5 mL of 1% FBS:PBS and centrifuged for another 5 min. Cell pellets were resuspended in 100 μL 5% FBS:PBS.
Controls
One hundred microliter of seven mTert-GFP uteri was pooled for use as unstained GFP controls. Two C57BL/6J uteri were pooled to form the “Wild-Type” unstained/isotype controls.
Cells were incubated in the dark at 4°C for 1 h with the conjugated primary and isotype control antibodies listed in Supplementary Table S1 (Supplementary Data are available online at
Immunofluorescence analysis
Upon dissection, uteri were cleaned of fat and immersed in 4% w/v paraformaldehyde in PBS overnight at 4°C, and then cryoprotected in 30% w/v sucrose in PBS overnight at 4°C. Tissues were frozen in an optimal cutting temperature medium (Sakura Finetek, Netherlands) and then cryosectioned at 8 μm thick. Sections were permeabilized in 0.2% Triton X-100 in PBS for 15 min, blocked in DAKO blocking buffer for 1 h, and then stained for GFP and epithelial/leukocyte markers for 1 h at room temperature (Supplementary Table S2). In the case of unconjugated primary antibodies, sections were incubated with secondary antibodies for 1 h at room temperature (Supplementary Table S2). Nuclei were counterstained with 5 μg/mL Hoechst 33258 for 3 min. Images were captured on an Olympus FV1200 confocal microscope (Tokyo, Japan) using a 20× or 40× objective lens and adjusted for brightness and contrast in a linear manner using FIJI software [10].
Quantification of mTert-GFP populations in the endometrium
Longitudinal sections of mTert-GFP uteri from four time points were examined for mTert-GFP+ cells in the epithelial and stromal cell compartments. Luminal and glandular epithelial mTert-GFP+ cells were confirmed through colocalization with epithelial cell adhesion molecule (EpCAM), while intraepithelial leukocytes were identified by CD45 staining and not included in epithelial cell counts. mTert-GFP+ cells in the epithelium were rare. Therefore, the number of mTert-GFP+ epithelial cells was not counted as a proportion of the total epithelium. Instead, a minimum of eight fields of view were imaged per section (n = 3–5/time point). Cells with the following combinations of markers were counted manually in each 20 × image using FIJI: EpCAM+, EpCAM+Ki67+, mTert-GFP+EpCAM+, and mTert-GFP+EpCAM+Ki67+.
To examine immune populations, mTert-GFP+CD3+ T cells and mTert-GFP+F4/80+ macrophages were manually counted in FIJI. A minimum of eight fields of view per section (n = 5–6/time point) were imaged and the distance between each dual-positive cell and the luminal epithelium was averaged from three points of the cell to account for the folds observed in the luminal epithelium at repair and remodeling time points. Data were plotted as a dot plot, where one dot is one average cell measurement. Uterine natural killer (uNK) cells were manually counted in FIJI. Total cell nuclei were calculated by counting the number of nuclei using the “threshold,” “watershed,” and “analyze particle” functions. A minimum of four fields of view per section (n = 4–5/time point) were imaged. The percentage of uNK cells in the total cell population and the percentage mTert-GFP+DBA-lectin+ cells in the total uNK cell population were calculated. Data were plotted with one point representing the mean percentage for each animal.
Statistical analysis
All statistical analyses were performed in Graphpad Prism 7.01. Raw data were subjected to D'Agostino-Pearson normality testing.
For those data where all time points passed normality testing, parametric one-way analysis of variance was performed using Tukey's multiple post hoc testing. Significance was accepted where P ≤ 0.05 and data are represented graphically as mean ± the standard error of the mean; data are displayed as triangular dot plots. Those data where one, or none of the time points, failed to pass normality testing were subjected to nonparametric Kruskal Wallis testing with Dunn's multiple comparison post hoc testing. Significance was accepted where P ≤ 0.05 and data are represented graphically as median; data are displayed as circular dot plots.
Results
mTert reporter activity remains constant during endometrial repair and remodeling
In agreement with our previous study, mTert-GFP+ cells in the uterus were rare [5]. Histological analysis of the endometrium revealed mTert-GFP+ cells scattered throughout the stroma close to the luminal epithelium (Fig. 1A, repair time point) and rare mTert-GFP+ cells in the epithelium (Fig. 1A, inset arrowheads).

Uterine mTert-GFP cells in the menses-like mouse model.
Flow cytometry analysis revealed that pooled mTert-GFP uteri contained, on average, 0.2% mTert-GFP+ cells in the total cell population (mTert-GFP+ cells green and mTert-GFP- cells yellow, Fig. 1B). The number of mTert-GFP+ cells in the total cell population did not significantly differ throughout our chosen time points (Fig. 1C; Table 1).
Summary of mTert-Green Fluorescent Protein Cell Populations As Investigated by Flow Cytometry
All values are shown as the percentage mean ± SEM of the total cell type population, for example, mTert-GFP+CD45+ of total CD45+ population.
No. of mTert-GFP+ cells in total live cell population.
SEM, standard error of the mean; GFP, green fluorescent protein; EpCAM, epithelial cell adhesion molecule; mTert, mouse telomerase reverse transcriptase.
mTert-GFP marks CD45+ leukocytes in the stroma during endometrial regeneration
Previously, we have shown that mTert promoter activity is predominantly localized to leukocytes in the stromal compartment of the mouse endometrium [5]; therefore, our first analysis was to determine which types of leukocytes express mTert-GFP during endometrial regeneration. mTert-GFP+ leukocytes localized to the endometrial stroma (Fig. 2A), particularly in the decidualized cell mass (upper panels white arrowheads) before breakdown of the endometrium at menses. At this time point, the majority of mTert-GFP+ cells observed histologically were CD45+ leukocytes. Upon endometrial breakdown and the initiation of repair mechanisms, mTert-GFP+CD45+ leukocytes localized to the stroma underlying the repairing epithelium (Fig. 2A, lower panels, yellow arrowheads) and were also observed in the residual basal layer (Fig. 2A, repair, white arrowheads).
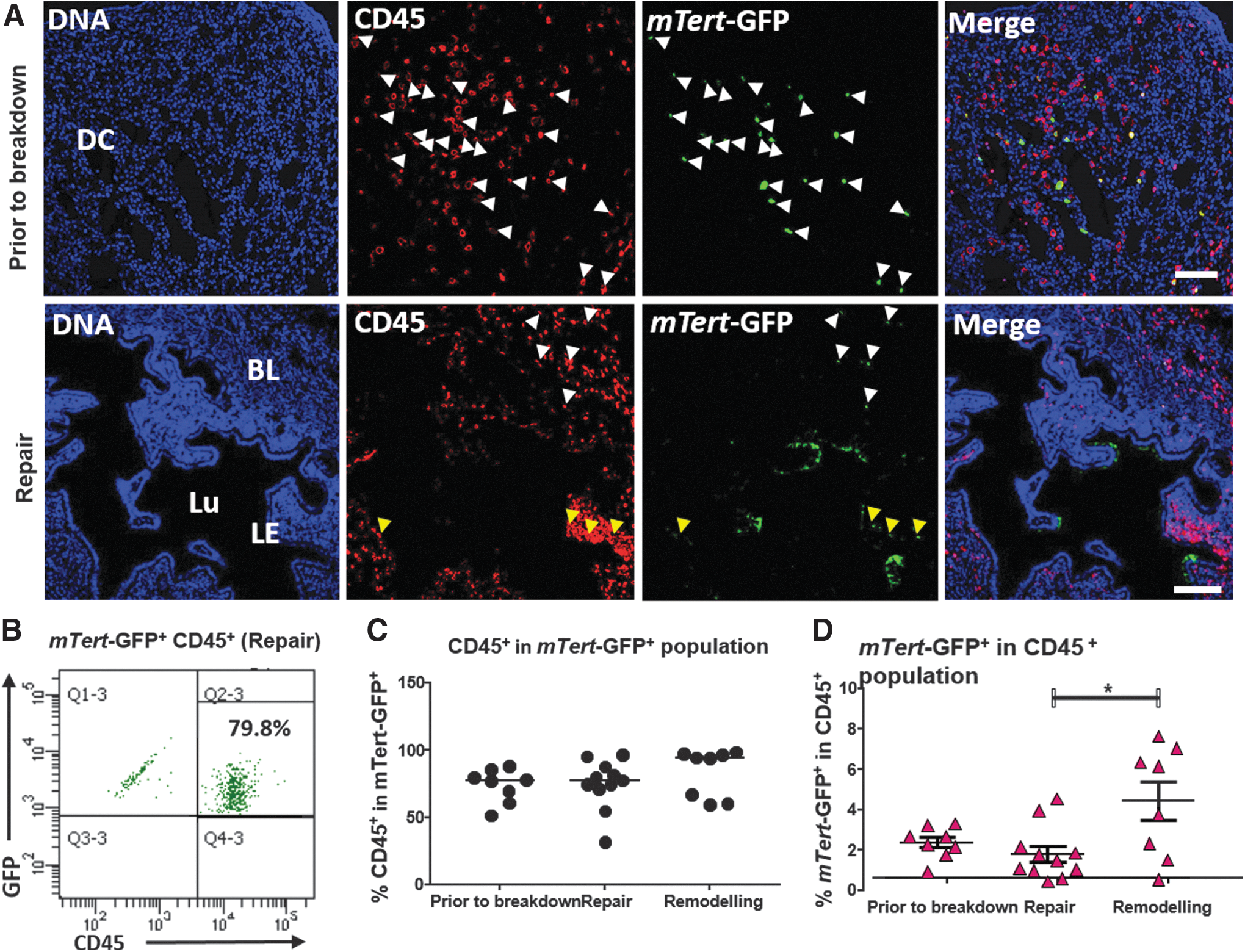
Coexpression of the pan-leukocyte marker CD45 and mTert-GFP in the menses-like model.
Flow cytometry analysis for mTert-GFP+CD45+ cells indicated that the majority of mTert-GFP+ cells were CD45+ leukocytes (Fig. 2B). Total mTert-GFP+ populations across three time points investigated constituted 73.53% CD45+ leukocytes before breakdown, 74.76% during repair, and 83.26% during remodeling (median values). While there were no significant differences in the percentage of mTert-GFP+ leukocytes (Fig. 2C) across the regeneration time points, the percentage of GFP+ leukocytes in the total CD45+ population did significantly increase during the remodeling phase (mean 4.42% ± 0.97 vs. 1.80% ± 0.4 during repair phase, Fig. 2D; Table 1).
mTert-GFP marks myeloid and lymphoid immune cell populations within the regenerating endometrium
As with previous studies in the intestine [6,11], characterization of the mTert-GFP+CD45+ population revealed that mTert-GFP expression marks both myeloid and lymphoid lineages during all stages of regeneration (Table 2; Fig. 3A, E). Interestingly, CD3+ T cells were the predominant cell type in the mTert-GFP+CD45+ population at all three time points investigated: 30.55 ± 5.5%, 44.63 ± 4.5%, and 65.01 ± 6.31%, respectively (Table 2). In addition, the percentage of mTert-GFP+CD3+ cells in the total CD3 cell population significantly increased during repair and remodeling, with 10 ± 1.63% of the T cell population also expressing mTert-GFP at the remodeling time point (Table 1; Fig. 3C). The overall percentage of CD3+ cells in total cell population significantly increased during the remodeling phase (P < 0.05; data not shown). The temporal and spatial distribution of phenotypical immune cells during repair and remodeling have been previously reported [12]. Therefore, to investigate whether mTert-GFP+ CD3+ T cells are activated and recruited to sites of endometrial remodeling to potentially aid in reepithelialization and endometrial regeneration, immunofluorescence for CD3 and mTert-GFP was performed and the distance of dual-labeled cells from the luminal epithelium was measured. Dual-positive cells were rare, as evidenced by the low number of cells counted and measured in 12 fields of view per animal per time point. Those identified typically fell into two groups (Fig. 3D) irrespective of the time point, those within 25 μm of the luminal epithelium and those greater than 150 μm from the luminal epithelium.
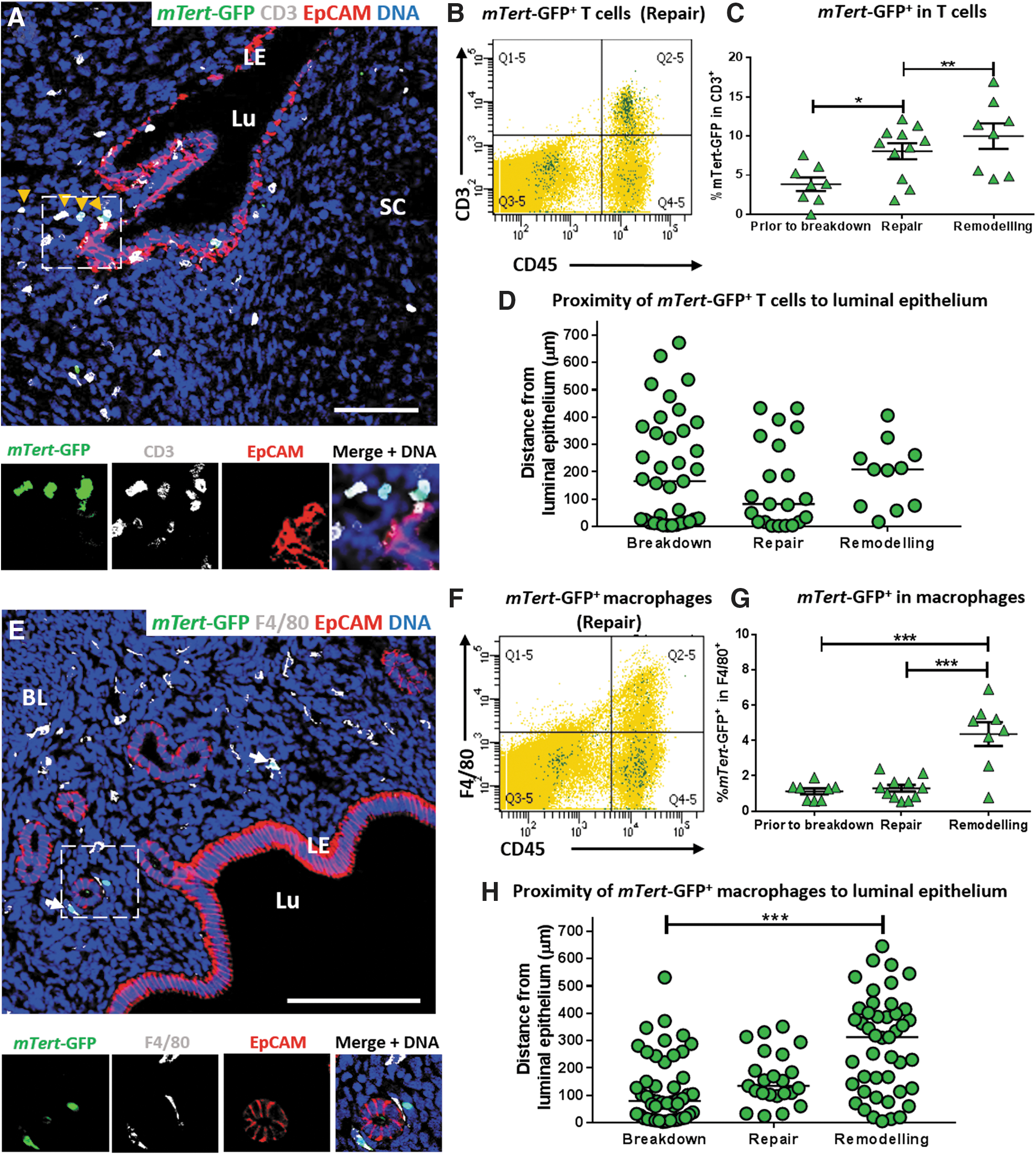
mTert-GFP+ T cells and macrophages in the mouse menses-like model.
mTert-GFP+ Immune Cell Types in the mTert-GFP+CD45+ Population As Investigated by Flow Cytometry
All values are shown as the percentage mean ± SEM.
mTert-GFP+ expression in the myeloid lineage was investigated by flow cytometry for F4/80+ macrophages (Fig. 3F), Ly6G+ neutrophils, and CD11b+ immature monocytes (Fig. 4A, B). mTert-GFP+F4/80+ macrophages accounted for 13%–31% of the mTert-GFP + leukocytes, depending upon the stage of regeneration (Table 2). Activation of mTert-GFP was observed during the remodeling phase, where mTert+F4/80+ cells accounted for 4.36 ± 0.67% of the total F4/80 population, significantly increased in comparison to the repair time point (Fig. 3G). Unlike dual-positive T cells, dual-labeled macrophages were evenly distributed throughout the regenerating stroma (Fig. 3H).
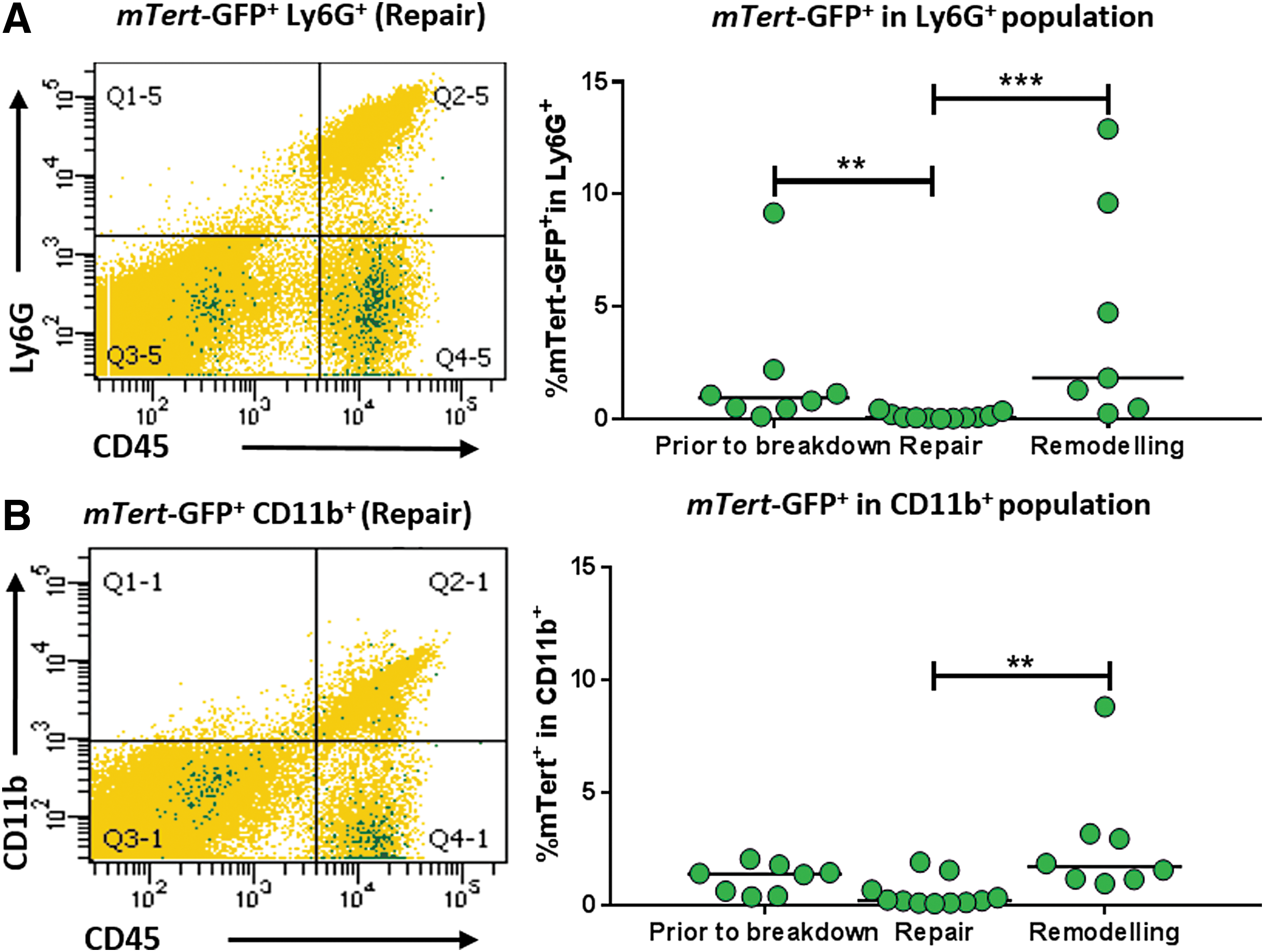
Flow cytometric analysis of mTert-GFP+ neutrophils and macrophage subpopulations in the uterus during the menses-like model
mTert-GFP was expressed by very low numbers of neutrophils and monocytes (Fig. 4A, B; Table 1) irrespective of time point. There was a significant decrease in the percentage of mTert+Ly6G+ neutrophils during the repair phase (compared to before breakdown); however, this coincided with an increase in the percentage of total neutrophils (data not shown). A significant increase in the percentage of mTert-GFP+CD11b+ monocytes was observed during the remodeling phase, but no change to the CD11b+ population was observed (data not shown).
A number of large lymphocytes were observed in the decidualized and shedding endometrium (Fig. 2A, CD45+). Dolichos biflorus agglutinin (DBA)-lectin staining revealed that these were uNK cells (Supplementary Fig. S1, red staining). The percentage of uNK cells in total cell number varied across the four time points investigated (median values: before breakdown 0.7%, breakdown 3.97%, repair 0.85%, and remodeling 0%). DBA-lectin+ cells were localized to the shedding endometrium and were rarely found in the residual basal layer. A small percentage of uNK cells were mTert-GFP+DBA-lectin+ (median values: before breakdown 0.23%, breakdown 1.145%, repair 0.24%, and remodeling 0%); dual-positive cells were primarily located in decidualized tissue or shed tissue.
mTert-GFP marks rare epithelial cell populations within the regenerating endometrium
We have previously shown that mTert-GFP marks both glandular and epithelial cell populations in the intact, cycling mouse endometrium [5]. Flow cytometric analyses indicated that mTert-GFP+EpCAM+ cells represent <0.1% of the total EpCAM+ population (Fig. 5A; Table 1), indicating that these cells are extremely rare. No significant differences in mTert-GFP+EpCAM+ cell percentage were observed during endometrial regeneration (Fig. 5A); however, flow cytometry analysis was unable to discern between luminal and glandular epithelial cells.

Characterization of mTert-GFP+ epithelial cells.
mTert-GFP+EpCAM+ cells were identified as clusters in areas of the luminal epithelium undergoing reepithelialization; however, no glandular epithelial cells were identified by immunofluorescent microscopy. Due to the rarity of mTert-GFP+ epithelial cells and their focal distribution, 2–3 clusters per animal, the whole luminal epithelium was not quantified. Instead, closer examination of these clusters was performed by counting the number of mTert-GFP+ and mTert-GFP- epithelial cells per eight fields of view. Of 17,902 luminal epithelial cells counted across four time points, only 205 were mTert-GFP+ (1.1%). No mTert-GFP+ epithelial cells were identified in the luminal epithelium before breakdown (Fig. 5B), although a significant increase in the percentage of mTert-GFP+ luminal epithelial cells was observed during the repair phase.
A previous study had indicated that CD44 marked progenitor cells in the mouse endometrium [13]. We have previously shown that CD44 did not colocalize with luminal epithelial mTert-GFP expression in the cycling mouse endometrium [5]. Triple immunofluorescence for mTert-GFP, CD44, and CD45, to rule out intraepithelial leukocytes, was performed on tissue sections during repair and regeneration. CD44 did not colocalize with luminal epithelial mTert-GFP+ cells (Fig. 5C) during regeneration; however, intraepithelial CD44+CD45+ cells were identified (orange arrowheads, inset). N-cadherin has been reported as an epithelial progenitor marker in human endometrium [4]; however, no N-cadherin expression was detected in the mouse endometrium during regeneration (Supplementary Fig. S2).
To determine whether mTert-GFP expressing luminal epithelial cells were proliferating, immunofluorescence for mTert-GFP+Ki67+ colocalization was performed. Dual-positive cells were identified during breakdown, repair, and remodeling phases (Table 3; Fig. 5D), surrounded by Ki67+ epithelial cells (Fig. 5D, repair). The number of mTert-GFP +Ki67+ and mTert-GFP-Ki67+ cells in each cluster was counted (Fig. 5E). Of the 2370 Ki67+ cells identified across four time points, a total of 17 cells were identified as dual positive for Ki67 and mTert-GFP (Table 3), with most dual-positive cells observed at the repair time point.
Proliferative and mTert-Green Fluorescent Protein Status of Luminal Epithelial Cells As Investigated by Histological Analysis
The total number of cells is shown for single- and dual-positive cells. The percentage of epithelial cells that are dual positive is shown in brackets.
Discussion
We investigated uterine mTert reporter expression in a mouse menses model and found that it marks a heterogeneous population of immune cells and clusters of endometrial epithelial cells. An influx of immune cells increased mTert-GFP expression in the endometrium during the repair and remodeling phases of the model. Although extensive reepithelialization occurred, there was not an accompanying expansion of mTert-GFP epithelial cells.
Resident immune cell populations express mTert during repair
As with our previous study of cycling and hormonally deprived endometrium, CD45+ leukocytes were the most abundant mTert-GFP+ population in the menses-like model, irrespective of regeneration time point. The mTert-GFP+CD45+ population contained both myeloid and lymphocyte lineages at all time points, in agreement with a previous study of mTert expression in the mouse [5].
Flow cytometry revealed fluctuating populations of immune cell types across all three time points, with CD3+ cells being the most common cell type in the mTert-GFP+CD45+ population irrespective of time point. In the case of the remodeling time point, where the mean percentages of the four cell types totaled >100%, it is likely that some overlap in cell types occurred, for example, CD3+CD11b+ or F4/80+Ly6G+. Our flow cell suspensions were incubated with each marker separately and therefore we are unable to report the degree of overlap in these experiments.
Our flow cytometry data revealed changes to the number of mTert-GFP+ immune cells; however, to determine whether mTert-GFP+ immune cells were likely to be contributing to repair, their proximity to the repairing epithelium was also investigated. mTert-GFP+F4/80+ cells were identified close to the repairing epithelium (within 100 μm) at all time points, but a significant shift in distribution was observed during the remodeling phase where a number of dual-positive cells were identified deep in the basal layer.
Previous studies assessing the immune cell types present at menses using the same mouse model have concentrated on myeloid cell types, which are typically involved in the inflammatory processes regulating menstruation [12,14 –16]. As in previous reports, an influx of neutrophils and immature monocytes was observed upon endometrial breakdown and repair, respectively [12,14]. Proinflammatory stimuli can promote TERT expression in human macrophages in vitro [17], whereas deletion of TERT results in cellular senescence suggesting that TERT maintains resident populations of macrophages. Thus, the detection of mTert-GFP+ macrophages in the menses/repair model may indicate a population of resident macrophages that is activated as part of the remodeling process. A previous study has reported spatial and temporal distribution of three mononuclear phagocyte subsets during repair and remodeling [12]. In this study, tissue-resident F4/80+Gr-1− macrophages remained deep in the basal layer of the endometrium and were not involved in repair. A subset of monocyte-derived F4/80+Gr-1+ monocytes was identified in subepithelial locations where reepithelialization was occurring. The data in this study support this previous evidence, suggesting that two subtypes of mTert+ macrophages may be involved in regeneration of the endometrium, one during breakdown and repair and another subset that is activated during remodeling.
Lymphoid contributions to endometrial repair have not been previously investigated. CD3+ T cells were located close to areas of breakdown and repair and the total CD3+ T cell population increased during remodeling. Interestingly, CD3+ T cells were the predominant immune cell type in the mTert-GFP+CD45+ population and dual-labeled cells were localized to the subluminal epithelium (within 100 μm) once menses had been initiated (repair phase), suggesting that T cells may contribute to reepithelialization by aiding tissue repair. Fewer dual-positive cells were observed in the residual endometrium in the remodeling phase, in contrast to our flow cytometry data where a significant increase in the percentage of mTert-GFP+CD3+ in the total CD3+ population was observed. This disparity may be attributed to using total uterine cell suspensions for flow cytometry, whereas dual-labeled cells in only the endometrium were counted for the proximity experiments and therefore the cells in the shed tissue were not quantified. Murine T cells do not usually express telomerase activity while at rest, but telomerase can be induced in an antigen-specific manner coincident with clonal expansion [18]. Our data showing increases in T cell mTert-GFP expression suggest that endometrial T cells are activated during the endometrial repair and remodeling phases and therefore may contribute to the regeneration of the tissue; however, further functional assays would be required to validate these findings.
uNK cells play an integral role in endometrial decidualization and early pregnancy [19]. In our model, DBA-lectin staining revealed that uNK cells were localized to the decidualized cell mass or the shedding endometrium and were rarely seen in the residual basal layer. This suggests that uNK cells do not contribute directly to repair of the residual endometrium as they are predominantly shed during menses. However, there is evidence to suggest that the shedding endometrium secretes factors that aid in reepithelialization [20]. mTert+DBA-lectin+ uNK cells observed at breakdown and repair phases may identify a population of uNK cells that were activated before progesterone withdrawal to maintain decidualization and support blood vessel maturation.
The endometrium is unique in its ability to undergo an inflammation-driven menstruation and then “heal” without scarring (reviewed by Salamonsen [21]). Regulatory T cells have been implicated as a key regulator of neutrophil and macrophage activity in the repair of other body tissues, including the lung and skeletal muscle [22 –24]. As members of the innate immune system, macrophages and neutrophils likely play the integral role of tissue destruction and clearance, but it would appear that T cells of the adaptive immune response, in particular, may also play a role in this tightly controlled process.
Luminal epithelial progenitors support reepithelialization of the endometrium
Reepithelialization of the human endometrium occurs in a piecemeal process [25]. It is controlled in such a way that the severity of damage and bleeding from sloughing of the functionalis is limited [25,26]. New luminal epithelial cells are postulated to arise from the stumps of basalis glands [27,28], and subsequent studies have shown that basal glands contain cells with stem/progenitor properties [4,29]. N-cadherin+ epithelial progenitors [4] and SSEA-1+ progenitors [29] are part of a glandular epithelial hierarchy [4] and play a key role in the formation of new epithelium during endometrial regeneration. N-cadherin expression was not detected in the mouse endometrium during regeneration, suggesting that it does not mark epithelial stem/progenitor cells in the mouse endometrium. Label-retaining cells have been identified in the glandular epithelium in a similar mouse model of menstruation [30], where glandular epithelial proliferation was maximal at 48 h after progesterone withdrawal, equivalent to the remodeling phase in this study. mTert-GFP+ cells have previously been shown not to overlap with label-retaining cells in the epithelium of intact mice [5], although both are located in the luminal epithelium, and therefore may mark a more progenitor-like cell. In this study, epithelial derived mTert-GFP+ cells were extremely rare, accounting for <0.15% of all epithelial cells in the endometrium when quantified by flow cytometry. Due to the rarity of these cells, it was not possible to isolate these cells to perform clonogenic assays to test their stem-like properties.
No mTert-GFP+ glandular epithelial cells were identified by microscopy in this study. In this model, the endometrium repairs in the absence of estrogen [8,9]. It is possible that glandular epithelial mTert expression is dependent on estrogen support, which correlates with human studies where glandular epithelial proliferation and migration occur later in the repairing process when circulating estrogen levels are increasing [26].
Despite the evidence of glandular epithelial stem/progenitor cells in human endometrium, evidence for other complementary mechanisms of reepithelialization has been put forward. The glandular epithelium of menstrual phase endometrium does not actively proliferate, despite evidence of reepithelialization by scanning electron microscopy [26]. It has been suggested that residual luminal epithelium may proliferate and migrate to denuded stromal surfaces, a mechanism that has been observed using a mouse model of menses [9]. Residual luminal epithelial cells have been observed to actively proliferate at the leading edge of cells, next to denuded stroma. Furthermore, the leading edge appears to be rolling across the denuded surface. In this study, residual epithelial cells were also observed to proliferate, as denoted by immunostaining for Ki67, but these cells were interspersed with mTert-GFP+Ki67− luminal epithelial cells. Of the ∼18,000 luminal epithelial cells visualized and counted, ∼1% were mTert-GFP+ during our regeneration window. mTert-GFP+ epithelial cells were present in areas of reepithelialization, but were rarely proliferating, as indicated by rarity of Ki67+mTert+ cells (0.09% mTert-GFP +Ki67+ luminal epithelial cells).
Our recent study indicated that mTert-GFP+ epithelial cells are an intrinsic component of the uterus. It has been reported that stem cells from the bone marrow transdifferentiate into endometrial epithelium [31 –34], although we highlighted the likelihood that intraepithelial immune cells have been misidentified as bone marrow-derived epithelial cells [35]. In this study, we performed triple immunostaining for GFP, EpCAM, and CD45 to detect GFP+CD45+ intraepithelial immune cells. The mTert-GFP+ cells we detected in the epithelial layer during the menses-like model were EpCAM+ and CD45−, confirming that they were an intrinsic endometrial epithelial population rather than infiltrating bone marrow-derived immune cells.
Luminal epithelial mTert-GFP+ cells were observed in hotspots close to sites of repairing epithelium. We suggest a model where luminal epithelial progenitor cells are interspersed along the entire epithelium, so that upon endometrial shedding, telomerase activity is activated to support reepithelialization of the tissue. mTert+ progenitors may undergo asymmetrical division to form a transit-amplifying cell, which in turn proliferates to form new Ki67+ epithelial cells that then migrate to repair the epithelium (summarized in Fig. 6). It is also possible that mTert is marking cells undergoing telomere salvage following extensive proliferation, a mechanism that has been previously reported in luminal epithelial cells in the mammary gland [36].

Potential mechanism of reepithelialization. mTert-GFP+ progenitor cells (green) may undergo asymmetric proliferation to form a transit amplifying cell (red) that rapidly divides to form new luminal epithelial cells.
It has been postulated that, in addition to the residual or glandular epithelium contributing to the new luminal epithelium, mesenchymal–epithelial cell transition (MET) may also play a role in reepithelialization, as evidenced in both human and mouse models of menstruation [9,25,26,37]. Transient cells close to the denuded edge were observed to express both mesenchymal and epithelial markers [9,26,37]. We did identify rare mTert-GFP+ cells in the stromal cell compartment by immunohistochemistry, which were CD45 negative, but these are most likely endothelial cells, as previously published [5]. We do have access to antibodies for SUSD2 to identify mesenchymal stem cells in human tissues; however, these antibodies are not specific for mouse antigens and therefore we were unable to look for any SUSD2 expression in our mouse tissues. While it is possible that stromal fibroblasts do undergo MET to form new luminal epithelial cells, we did not identify any potential stromal-derived mTert-GFP+ progenitors, suggesting that mTert is not a suitable marker for identifying mesenchymal progenitors.
Conclusions
mTert-GFP expression identifies rare leukocyte and epithelial populations in the mouse endometrium during endometrial repair and regeneration. mTert facilitates stem cell renewal in other tissues and our findings point to a role for mTert in the cyclical regeneration of epithelial compartments of the endometrium. mTert-GFP expression in immune cells may also provide insight into resident and activated innate and adaptive populations involved in the regulation of regeneration. Further insight into how stem/progenitor cells and immune cells contribute to endometrial repair and regeneration after menstruation will help improve our understanding of endometrial disorders such as heavy menstrual bleeding, Asherman's syndrome, and endometriosis.
Footnotes
Acknowledgments
This work was supported by an NHMRC project grant (1085435) to C.E.G. and J.A.D. and the Victorian Government's Operational Infrastructure Support. The authors acknowledge the technical support of Monash Health Translational Precinct Platforms; Histology and Micro Imaging.
Conference Presentation: Data presented, in part (poster), at the 48th Annual Scientific Meeting of the Society for Reproductive Biology, August 2017, Perth, Australia. Data presented, in part (poster), at the International Society for Stem Cell Research, Melbourne, Australia, June 2018. Data presented, in part (oral presentation), at the 51st annual meeting of the Society for the Study of Reproduction, New Orleans, America, July 2018.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.