
Abstract
Uterine remodeling during pregnancy is a fundamental, dynamic process required for successful propagation of eutherian species. The uterus can increase in size up to 40-fold during pregnancy, which is largely attributed to expansion of the myometrium by hyperplasia and hypertrophy. After pregnancy, the uterus repairs the remodeled or “damaged” tissue during uterine involution (INV). Little is known about this repair process, particularly the role of mesenchymal stem/progenitor cells. The objective of this study was to identify and characterize putative mesenchymal stem/progenitor cells in the murine myometrium using a combination of label retention and mesenchymal stem cell (MSC) marker expression and a pregnancy and uterine INV model. Tet-off transgenic mice with the Cre-lox system were used to specifically label mesenchymal cells (ie, myometrial and endometrial stromal cells) within the uterus while avoiding other cell types (eg, epithelial, immune, and endothelial cells) to identify slowly dividing cells and assess their stem cell qualities. We identified myometrial label-retaining cells (LRCs) that persisted for at least 3 months, expressed CD146 and CD140b (MSC markers), and proliferated at a higher rate during uterine INV compared with nonlabeled cells. The LRCs did not appear to express either estrogen receptor alpha or progesterone receptor, nor did the number of LRCs change at different estrous stages or in response to exogenous estradiol or progesterone administration, suggesting that LRCs were not involved in normal estrous cycling. The results from this study provide important insight into putative stem/progenitor cells in the myometrium and their possible role in uterine physiology.
Introduction
T
After parturition, the uterus returns to its nongravid state in a process termed uterine involution (INV). Very few studies have assessed the cellular mechanisms of myometrial INV, which are thought to include apoptosis, proliferation [4], and autophagy [5]. Even more unclear is the role of stem/progenitor cells in myometrial INV, particularly in the replacement of apoptotic cells and terminally differentiated or hypertrophied cells. To replace these cells, it is thought that proliferation and differentiation are reactivated, most likely by stem and/or progenitor cells, since terminally differentiated myometrial cells do not proliferate [6]. We have previously shown some of the strongest evidence for a myometrial stem cell population in murine uteri in the Ctnnb1 cKO mouse, which undergoes a switch from myogenesis to adipogenesis as the mouse develops postnatally [7].
Adult stem cells are responsible for tissue homeostasis in many tissues [8,9]. Classically, stem cells have been characterized by their relatively quiescent nature, their ability to both self-renew and generate lineage-specific daughter cells, and ultimately by their ability to repair or replace lost tissues. Several methods exist for identifying or enriching for putative somatic stem cells, including expression of validated stem cell markers, in vitro and in vivo stem cell assays, and label retention [10,11]. Mesenchymal stem cells (MSCs) have been identified in various tissues by expression of specific surface antigens and lack of expression of others [12]. In human tissues, classical markers of MSCs include CD73, CD90 and CD105. Additional markers, including CD146 and CD140b, which can mark perivascular cells, can also serve to enrich for putative MSCs in various tissues [13,14]. In human endometrial stroma, putative MSCs were initially recognized by their colony-forming [15] and multilineage differentiation [16] potential, and shown to be isolated by coexpression of CD146 and CD140b [17] and later by singular expression of SUSD2 [18]. Because the endometrial stroma and the myometrium originate from the same embryonic tissue, the Müllerian duct mesenchyme (MDM), it is plausible that these markers may be used to identify putative MSCs in the myometrium.
Generally, stem cells divide relatively infrequently and can retain nuclear labels such as the nucleotide analog, bromodeoxyuridine (BrdU), while other more frequently dividing cells lose label over time. BrdU label retention has been used to identify label-retaining cells (LRCs) in various tissues [19 –22]. Our laboratory recently identified myometrial BrdU-LRCs that are hormonally responsive [23]. However, no system has been reported to isolate viable BrdU-LRCs for assays to confirm their stemness. Therefore, a transgenic approach was developed to circumvent this limitation [24]. In the “tet-off” transgenic mouse model, a histone H2BJ-GFP fusion protein serves as the label, and this becomes stably incorporated into the nucleosomes of dividing cells during the “labeling” period. H2BJ-GFP expression is then stopped, and the label is diluted out during the “chase” period in dividing cells but is retained in more quiescent cells. This model has been used to detect LRCs in the female reproductive tract [25,26], specifically by our laboratory to characterize LRCs in the ovarian surface epithelium [27]. In this study, we have used a version of this mouse model that allows for both tissue-specific and temporally controlled expression of H2BJ-GFP by incorporating the Cre-lox system. Using Amhr2-Cre expressing mice, H2BJ-GFP expression was restricted to the MDM-derived myometrium and endometrial stroma. With this model, we have identified and characterized LRCs in the adult mouse myometrium as putative MSCs.
Materials and Methods
Animals
All protocols involving animal experiments were approved by the Institutional Animal Care and Use Committee at Michigan State University. CD1 mice were purchased from Charles River and maintained in house. TRE-H2bj-GFP [Tg(tetOHIST1H2BJ/GFP)47Efu] mice [24] and Rosa-flox-stop-tTA [Gt(ROSA)26Sortm1(tTA)Roos/J] mice [28] were purchased from The Jackson Laboratory (Bar Harbor, ME). Amhr2-Cre [Amhr2tm3(cre)Bhr] mice were kindly provided by Dr. Behringer [29]. Triple transgenic offspring, Amhr2-Cre;Rosa26-tTA;TRE-H2bj-GFP, were generated and are hereafter referred to as LRC mice. LRC mice and indicated controls received doxycycline HCl (dox, 2 mg/mL; Research Products International, Mt. Prospect, IL) in drinking water, which was refreshed two times per week for the duration of the chase period. Uteri were collected from CD1 mice at P 0.5, 5, 10, and 25 and from LRC and control females at the following time points: No chase (P 28), and 1, 2, 3, and 6 months of chase. Tissues were also collected after hormone treatment in ovariectomized mice or during pseudopregnancy and uterine INV (ie, postpartum uterine repair) as described below. The following genotypes served as controls: (1) LRC mice that were maintained on dox from conception (transplacental/translactational exposure via dams) to time of tissue collection to determine “leakiness” of the system and (2) Amhr2-Cre;Rosa26-tTA mice that received dox with LRC mice to determine autofluorescence in the tissue.
Estrous cycle-staging and hormone manipulation
To assess estrous cycle stage, 40 μL of 10% trypan blue in PBS was lavaged into the vaginal canal of 3 months chase-LRC mice, then placed on a glass slide and cover slipped. Stages were determined by the presence/absence of leukocytes, nucleated epithelial cells, and cornified epithelial cells [30]. Serum was then collected from each mouse on specified days of the estrous cycle by retro-orbital bleed method, and assayed for E2 and P4 (University of Virginia Ligand Core).
For hormone treatments, LRC mice at 3 months chase were first ovariectomized and allowed to clear endogenous hormones for 1–2 weeks. Mice were then given one of the following hormone treatments [31] in sesame oil s.c. for 3 consecutive days: (1) vehicle (Veh, sesame oil alone), (2) estrogen (E2, 125 ng; Sigma-Aldrich, St. Louis, MO), (3) progesterone (P4, 2 mg; Sigma-Aldrich), and (4) E2+P4 (125 ng and 2 mg, respectively). Twenty-four hours after the last treatment, uteri were collected, and one horn was processed for gelatin embedding and the other was processed for flow cytometry.
Pseudopregnancy and INV
Pseudopregnancy followed by uterine INV was induced in LRC females as previously described [32] to assess proliferation during myometrial expansion (pseudopregnancy) and uterine repair (INV). In brief, females were placed with vasectomized CD1 males and upon the observation of a vaginal plug were designated day of pseudopregnancy (DOPP) 0.5. On DOPP 4, sesame oil was injected into the lumen of both uterine horns to induce endometrial decidualization, which leads to expansion of the myometrium similar to natural pregnancy. Seventy-two hours postoil-induced decidualization (DOPP 7.5), INV was induced by bolus injection of the progesterone receptor antagonist, RU486 (8 mg/kg s.c. in sesame oil; Sigma-Aldrich), for 2 days. During this time, the uterus returned to its nongravid state. One hour before euthanasia, mice were given a single i.p. injection of 5-ethynyl-2′-deoxyuridine (EdU, 50 mg/kg; Thermo Fisher Scientific, Waltham, MA). Mice were euthanized, and uteri were collected at DOPP 0.5, 4, and 7.5 and at 48 h during uterine INV. One uterine horn was processed for frozen gelatin embedding and the other for flow cytometry.
Flow cytometry
At the designated time points, uteri were harvested from animals, and one horn was split open lengthwise. The endometrium was carefully scraped out under a dissecting microscope leaving most of the myometrium intact. The tissue was cut into 1-mm pieces and digested at 37°C in digestion media (DMEM/F12, 10% FBS, 2 mg/mL Collagenase Type II, 1 mg/mL DNase I, 5 mM MgCl2) with agitation for 1–1.5 h to release myometrial cells from the serosa. The resulting cell suspension was passed through a 100-μm cell strainer, washed with PBS, and pelleted. The cell pellet was resuspended in 5 mL ACK Lysing Buffer (Thermo Fisher Scientific) to remove red blood cells, then washed with PBS and pelleted again. Cells were resuspended in flow cytometry buffer (1% BSA in PBS) and analyzed on an Accuri C6 flow cytometer (BD Biosciences, San Jose, CA). Events were gated initially by forward and side scatter, then for singlets (side scatter area × height) and finally for GFP fluorescence using FlowJo software (FlowJo, Ashland, OR).
For analysis of EdU expression, dissociated cells were treated with the Click-iT Plus EdU Alexa Fluor 647 Flow Cytometry Assay Kit (Thermo Fisher Scientific) per manufacturer's instructions. In brief, cells were washed in flow cytometry buffer, pelleted, then incubated in Click-iT fixative. Cells were washed again in flow cytometry buffer, then incubated in Click-iT saponin-based permeabilization and wash reagent before adding Click-iT Plus reaction cocktail for an additional incubation. Cells received a final wash in Click-iT saponin-based permeabilization and wash reagent, were resuspended in the same reagent, and analyzed by flow cytometry. Events were gated as above with the addition of gating for EdU expression. Fluorescence minus one was used as the gating control for EdU, and mice lacking the H2bj-GFP transgene served as the gating control for GFP.
Gelatin embedding and frozen tissue preparation
After collection, uteri were fixed in 4% paraformaldehyde for 10–45 min on ice depending on age of mice, washed three times in ice-cold PBS, and then incubated overnight at 4°C in 15% sucrose buffered in PBS. The next day, tissues were incubated at 37°C for 1 h in a gelatin mix (15% sucrose, 7.5% gelatin in PBS), embedded on a cold block, frozen at −50°C to −65°C in isopentane, and stored at −80°C until sectioning. Tissues were cryosectioned at 8 μm and thaw mounted. Gelatin was removed from the tissue sections by incubating slides in 37°C PBS. After gelatin removal, slides were either used for immunofluorescence staining or immediately counterstained with 4′,6-diamidino-2-phenylindole (DAPI; BioLegend, San Diego, CA), cover slipped with Fluro-Gel [Electron Microscopy Sciences (EMS), Hatfield, PA], and viewed for direct GFP fluorescence.
Immunofluorescence
The following primary antibodies were used: biotinylated-CD140b (1:50; Cat. No. 13-1402-80; Thermo Fisher Scientific ), CD146 (1:100; Cat. No. 04-1147; Millipore, Burlington, MA), CD31 (1:100; Cat. No. RB-10333; Thermo Fisher Scientific), estrogen receptor alpha (Erα) (1:200; Cat. No. 32063; Abcam, Cambridge, MA), progesterone receptor (PR) (1:100; Cat. No. A0098; Agilent Technologies), Ki-67 (1:100; Cat. No. RM-9106; Thermo Fisher Scientific), and CD44 (1:250; Cat. No. ab189524; Abcam). Secondary antibodies used were as follows: streptavidin-Cy3 (Cat. No. SA1010; Thermo Fisher Scientific) and goat antirabbit AlexaFluor 568 or 647 (Thermo Fisher Scientific) all at 1:1,000. After removal of gelatin tissues were incubated for 1 h at room temperature (RT) in blocking solution (0.1% triton × 100, 0.1% BSA, and 10% normal goat serum in PBS), followed by incubation with primary antibody diluted in blocking solution overnight at 4°C. Tissues were then washed with PBS, and secondary antibody diluted in blocking solution was applied in the dark for 45 min at RT. Tissues were washed in PBS followed by water, then counterstained with DAPI (BioLegend), and cover slipped with Fluro-Gel (EMS). For costaining with biotinylated-CD140b and CD146, after gelatin removal, tissues were incubated with Endogenous Biotin-Blocking Kit (Thermo Fisher Scientific) per manufacturer's instructions, then continued with blocking solution in above protocol. Omission of primary antibodies served as a negative control.
Statistical analyses
One-way ANOVA (GraphPad Prism, La Jolla, CA) was used for analysis of the percentage of GFP+ cells, as shown in Supplementary Fig. S1 (Supplementary Data are available online at
Results
CD146 and CD140b are spatially and temporally expressed during myometrial differentiation
CD146 and CD140b coexpressing cells, which are enriched for putative MSCs, are located in the perivascular space in human endometrial stroma [17]. The myometrium and the endometrial stroma originate from the same embryonic tissue, the MDM [32,35]. We therefore examined whether CD146 and CD140b were similarly expressed in perivascular cells in the myometrium and could potentially mark putative MSCs in that tissue. We determined the spatiotemporal expression patterns of CD146 and CD140b in murine uteri during postnatal differentiation of the MDM into the endometrial stroma and myometrium. At birth [postnatal day (P) 0.5], the mesenchyme was undifferentiated, and CD146 and CD140b were expressed ubiquitously throughout the mesenchyme and absent from the single layer of luminal epithelium (Fig. 1A–C). By P 5, the mesenchyme had begun differentiating into the endometrial stroma and the inner circular and outer longitudinal layers of the myometrium, which are demarcated by the white line (Fig. 1D–F). At this time, although CD140b remained ubiquitously expressed, CD146 became more restricted to the differentiating myometrium. At P 10, the inner circular and outer longitudinal layers of the myometrium were clearly differentiated and continued to show high expression of CD146 (Fig. 1G–I). Within the endometrial stroma, CD146 appeared to become restricted to the vasculature. CD140b expression was decreased in the outer myometrial layer but was still abundantly expressed in the inner layer and the endometrial stroma. At this early age, perivascular expression of CD146 and CD140b was already evident, particularly in the endometrial stroma. By P 25, the histoarchitecture of the adult uterus was complete (Fig. 1J–L). CD140b continued to be expressed in the endometrial stroma but was restricted to the perivascular cells in the two myometrial layers. In the myometrium of the adult, sexually mature mouse, CD146 and CD140b were coexpressed by perivascular cells (Fig. 1M–O). These results indicate that the diffuse expression of CD146 and CD140b on P 0.5–5 coincided with the undifferentiated state of the mesenchyme. Then as the tissue differentiated, CD146 and CD140b became more restricted to the perivascular space, a well-established MSC niche in many tissues [12 –14].

CD146 and CD140b are spatially and temporally expressed during postnatal uterine maturation. Representative cross-sections of mouse uteri (n = 3/time point) showing expression of CD146 and CD140b at postnatal (P) and adult ages.
Murine myometrium contains LRCs
After having identified the perivascular niche for the CD146 and CD140b coexpressing cells, we employed another mouse model to further characterize these perivascular cells as putative MSCs. In LRC mice (Fig. 2A), tetracycline transactivator (tTA) was constitutively expressed in MDM cells and bound the tetracycline response element (TRE) inducing expression of H2BJ-GFP, which became stably incorporated into nucleosomes during DNA replication, thereby labeling MDM-derived cells (stroma and myometrium). Mice were allowed to incorporate H2BJ-GFP label beginning during embryonic Müllerian duct development (E 12.5) and continuing through postnatal uterine maturation up to postnatal (P) day 28 (Fig. 2B, Label). To stop labeling and initiate the chase period (Fig. 2B, Chase), dox was administered to mice in their drinking water beginning at P 28 until collection. This resulted in dilution of residual GFP in frequently dividing cells but retention in more quiescent cells allowing for identification of LRCs.
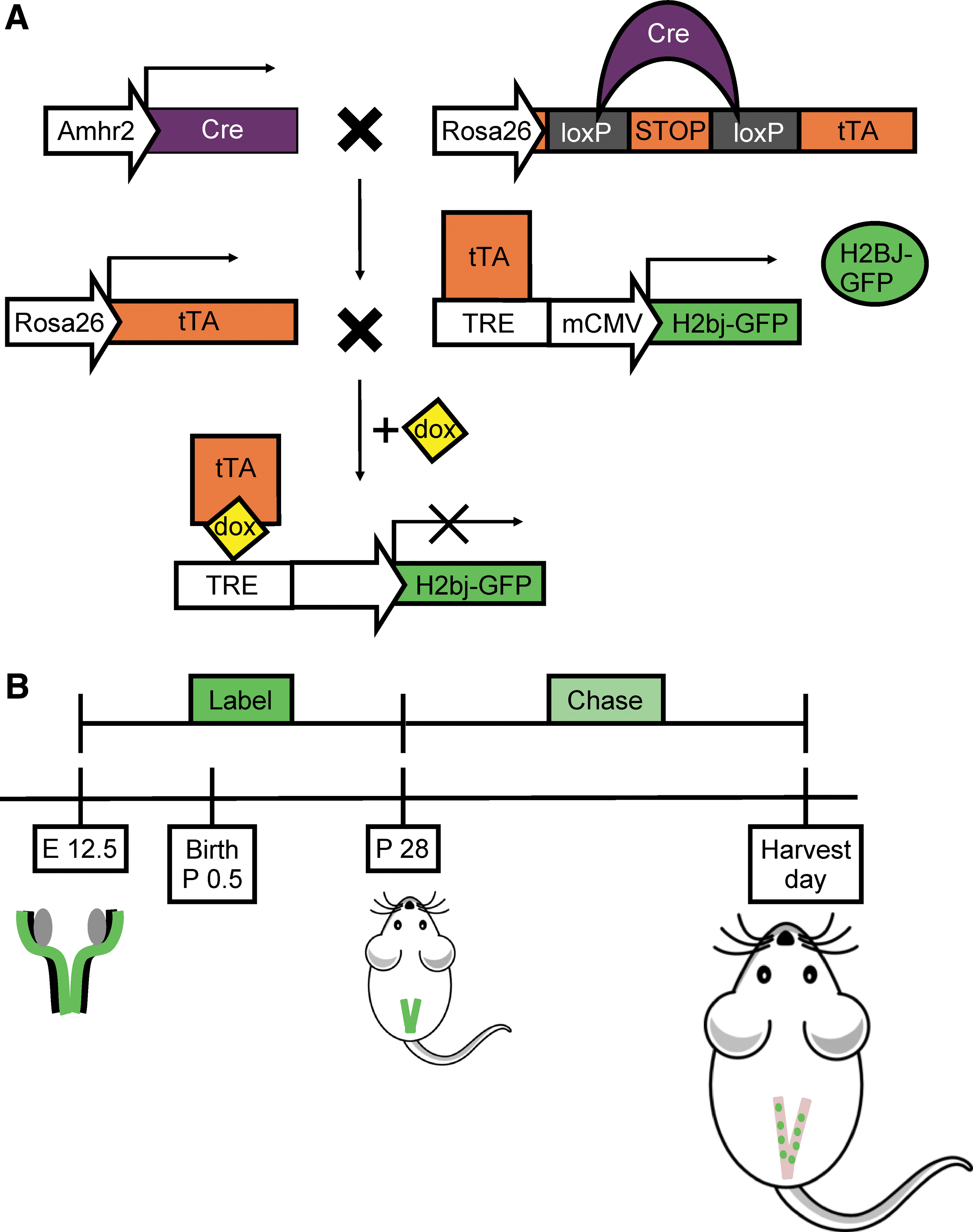
“Tet-off” LRC transgenic mouse model.
To determine the location of LRCs and the extent of label retention within the myometrium, uteri were collected from LRC mice at the following time points: No chase (P 28) and at 1, 2, 3, and 6 months of chase. At the no chase time point, almost all of the myometrial cells (mean = 94%) were GFP+ indicating efficient labeling (Fig. 3A, B, I), and a distinct population of GFP-bright (GFPbr, ∼47%) cells were identified representing about half of the total GFP+ cells. After 1 month of chase ∼60% remained labeled (Supplementary Fig. S1A, B and Fig. 3I), and at 2 months ∼25% were labeled (Supplementary Fig. S1C, D and Fig. 3I). At 3 months (Fig. 3C, D) of chase, the dilution of label appeared to level off at ∼25% of the cells, and the GFPbr had dropped to ∼1% of the total cells (Fig. 3I). By 6 months (Fig. 3E, F, I) of chase, the percentage of LRCs was not different from the leaky GFP expression seen in control mice (LRC mice on consistent dox to prevent H2B-GFP expression) (Fig. 3G–I). These results indicate that (1) in general, there was substantial cellular turnover in the myometrium during the estrous cycle as seen by dilution of label, and (2) despite the relatively high turnover, the myometrium contained slowly dividing cells as indicated by the persistence of LRCs for several months.
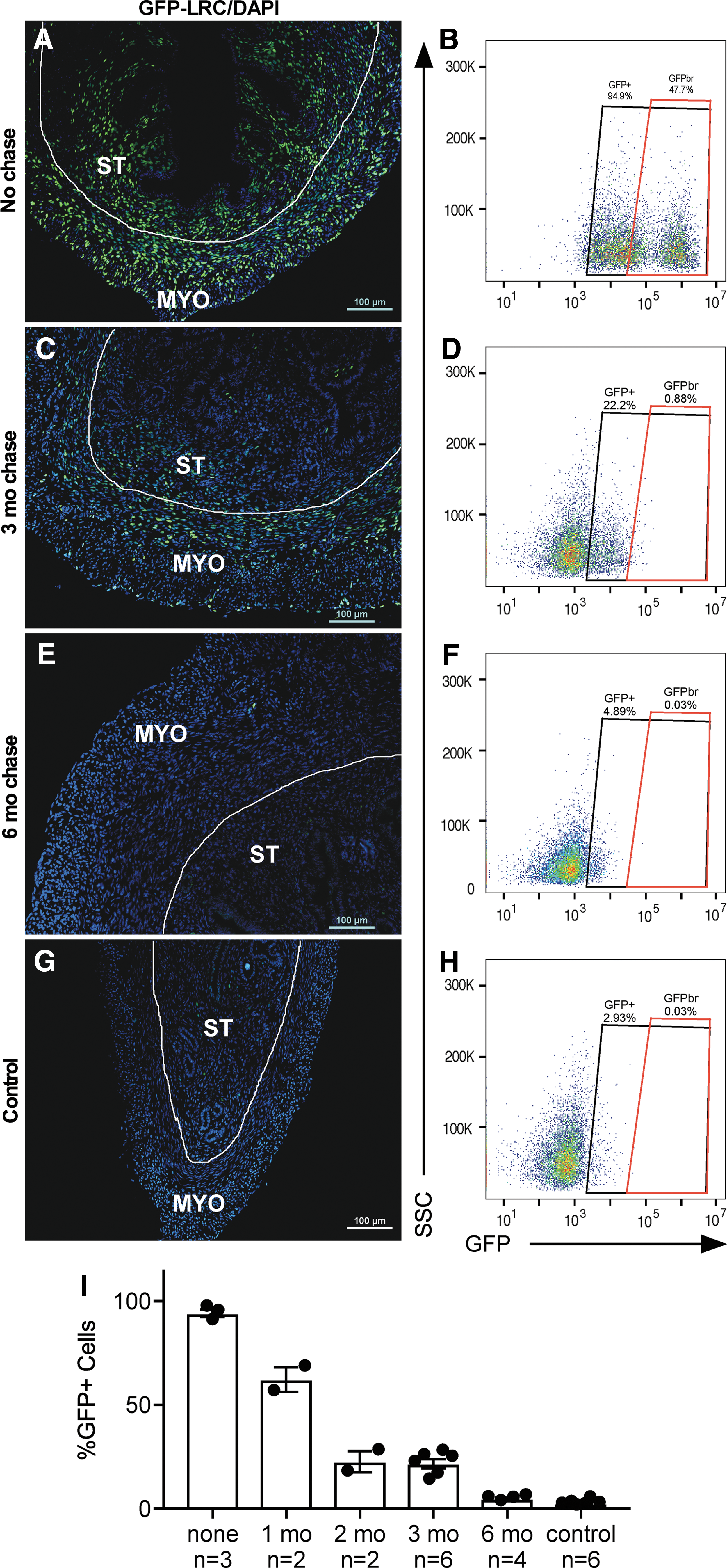
Timeline of label retention in myometrial cells. Representative images [sample size indicated in graph
Distinct populations of LRCs exist within the myometrium
Upon closer examination of the LRCs within the myometrium at 3 months chase, an array of LRCs were observed. Specifically, unique bright LRCs, which are cells that had divided less as compared with dim LRCs, were identified. The bright LRCs were few in number and tended to be located in the perivascular space as seen by surrounding CD31+ endothelium, whereas dimmer LRCs were much more abundant and located more randomly (Fig. 4A–C). Bright-perivascular LRCs also did not express ERα or PR. However, dim, nonperivascular LRCs did express PR. The percentage of LRCs (dim and bright) did not vary across the estrous cycle (Supplementary Fig. S2A, P = 0.769) or by endogenous levels of E2 and P4 (Supplementary Fig. S2B; P = 0.145 and P = 0.158, respectively). Similarly, the percentage of LRCs did not vary significantly in ovariectomized mice in response to exogenous hormone treatment (Supplementary Fig. S2C, P = 0.158). Bright-perivascular LRCs coexpressed the MSC markers, CD146 and CD140b (Fig. 5A–D, arrowhead). Interestingly, bright-perivascular LRCs could be identified in the stroma and also coexpressed CD146 and CD140b (Fig. 5A–D, arrow). Studies on human myometrium have shown that CD44 in combination with Stro-1 enrich for putative myometrial stem/progenitor cells [36]. We show that bright-perivascular LRCs do not appear to express CD44 (Supplementary Fig. S3A–C). However, when images were overexposed to visualize dimmer LRCs, CD44 was often expressed by very dim LRCs that tended to be located near brighter LRCs (Supplementary Fig. S3D–F). These dim CD44+ LRCs may represent more differentiated cells that underwent more divisions resulting in loss of label. Taken together, these results suggest that bright-perivascular LRCs in the myometrium are relatively quiescent and less differentiated, and therefore may be a mesenchymal stem/progenitor population.
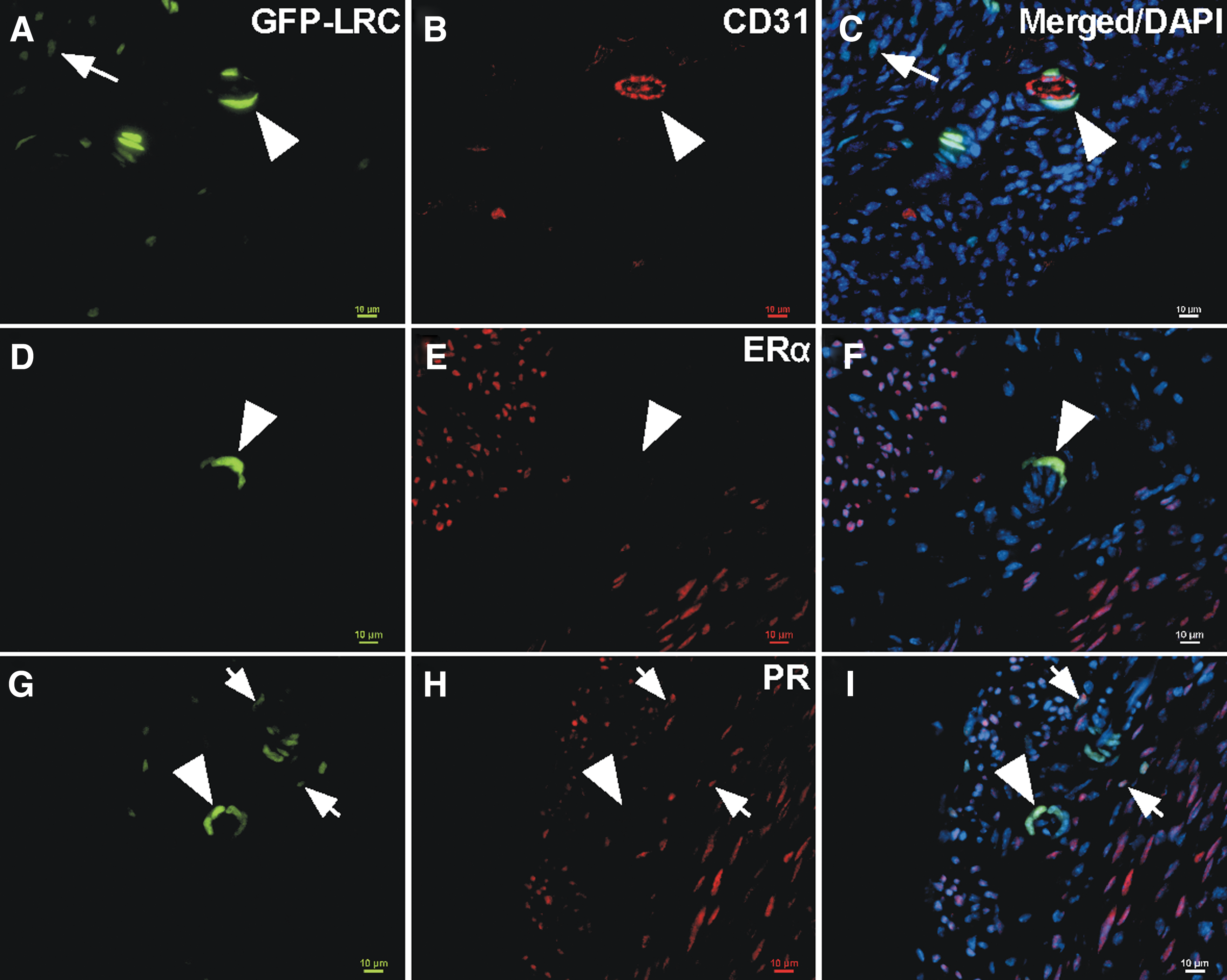
Characterization of bright LRCs. Representative images of uterine cross-sections showing dim and bright LRCs.
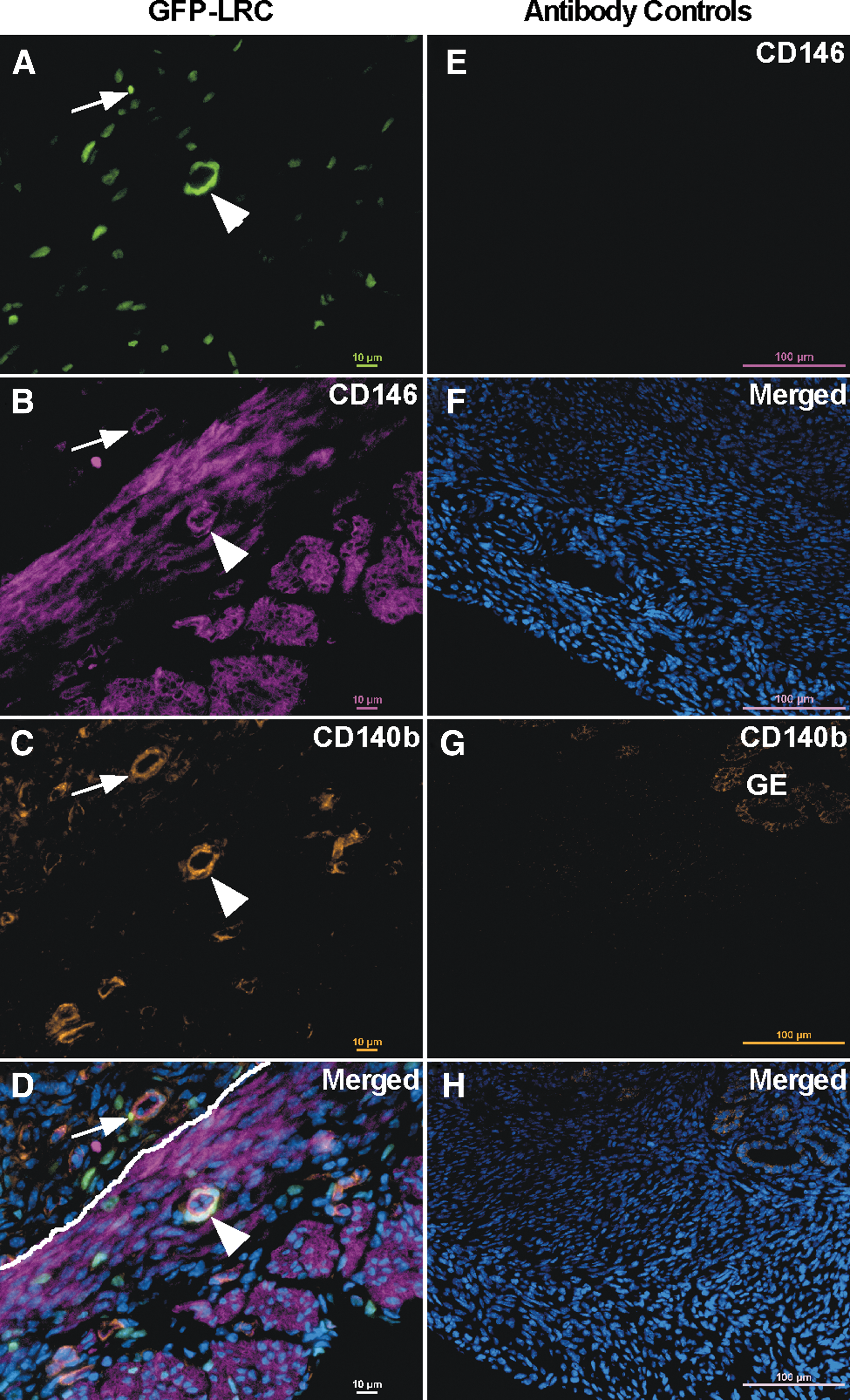
Bright perivascular cells express MSC markers. Representative images (n = 6 at 3 months chase) of uterine cross-sections showing expression of CD146 and CD140b in bright-perivascular LRCs.
Distinct LRC populations are activated in response to pseudopregnancy and uterine INV
The contribution of LRCs to two important uterine processes, pregnancy and uterine INV (ie, postpartum uterine repair), was determined using a time-controlled model for these events (Fig. 6A) [32]. Histological images show the dramatic changes that occurred as the uterus transitioned from the estrous cycling state (Fig. 6B) through decidualization (Fig. 6C) and during uterine INV (Fig. 6D). Similar to what was seen during the estrous cycle, we did not see a significant difference in the overall percentage of LRCs at the different times during pseudopregnancy and uterine INV (Supplementary Fig. S2D, P = 0.429). To assess proliferation of LRCs, mice were given a bolus injection of EdU 1 h before harvesting tissues at each time point (Fig. 6A, boxed days), and the cells were analyzed by flow cytometry. There was a significant increase in the percentage of EdU+ LRCs from DOPP 0.5 to 7.5 (Fig. 6E, P < 0.0001). At DOPP 7.5, there was the highest percentage of EdU+ LRCs (∼12%), which corresponded with myometrial expansion to accommodate the growing decidual tissue (Fig. 6C). During uterine INV (Fig. 6D), the decidual tissue degenerated, and the uterus as a whole underwent repair, which was complete within 48–72 h. The percentage of EdU+ LRCs decreased from DOPP 7.5 to 48 h into INV; however, there was a significantly higher proportion of EdU+ cells at INV 48 h compared with DOPP 0.5 and 4 (Fig. 6E, P ≤ 0.001 and 0.04, respectively). Interestingly, when comparing the LRC and non-LRC populations, there was a significantly higher percentage of EdU+ cells at DOPP 0.5, DOPP 4, and INV 48 h (Fig. 6F, P ≤ 0.05, 0.05, and 0.0001, respectively) but not at DOPP 7.5. These results suggest that LRCs contributed more to proliferation at DOPP 0.5, DOPP 4, and INV 48 h; however, when substantial expansion of the tissue was required (DOPP 7.5) both non-LRCs and LRCs contributed equally.
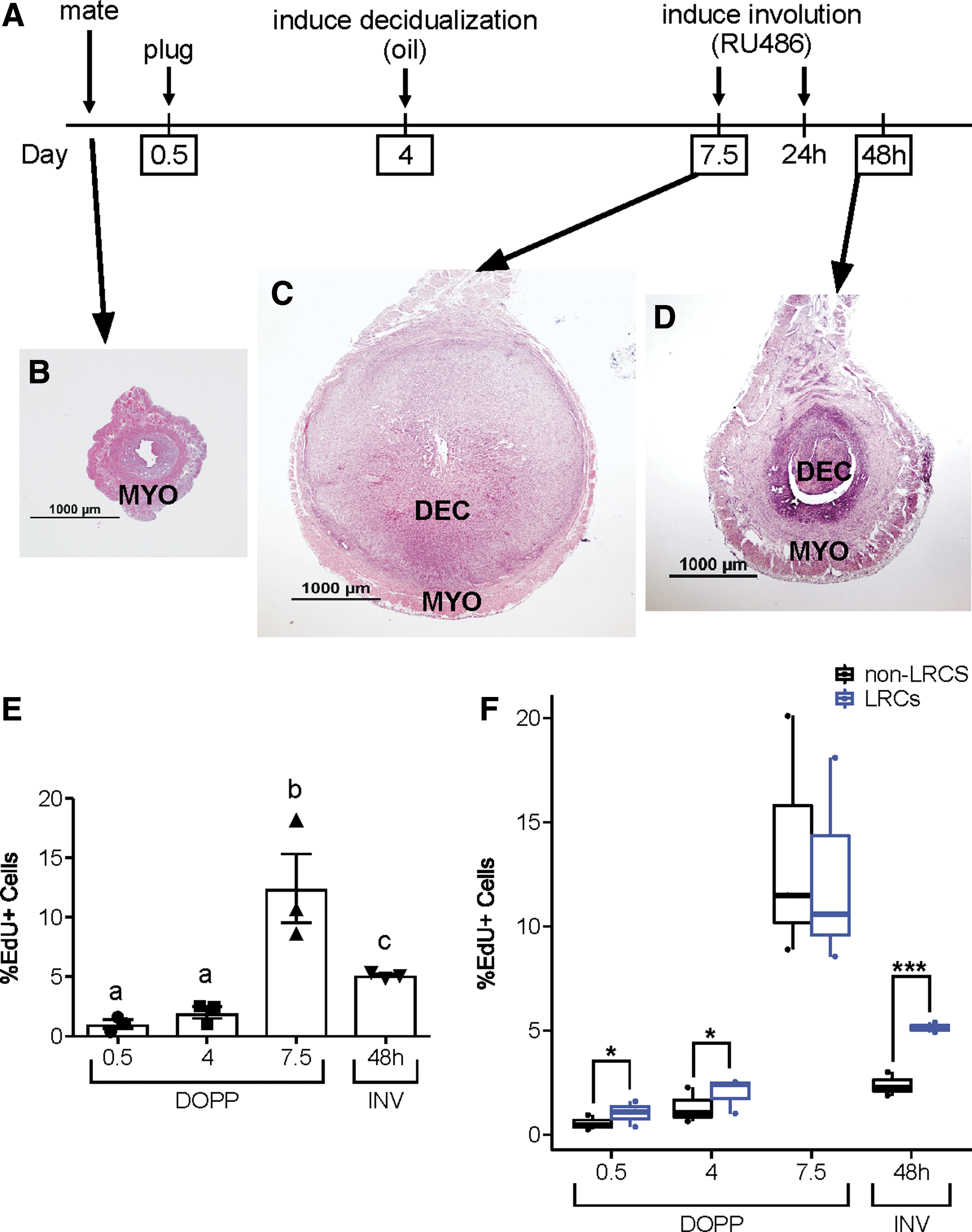
Proliferation of LRCs during pseudopregnancy and uterine INV.
To further evaluate the contribution of LRCs to pseudopregnancy and INV, tissues were immunostained for the proliferation marker Ki-67 in the distinct bright-perivascular LRCs compared with dim/nonperivascular LRCs. During early pseudopregnancy (DOPP 0.5 and 4), LRCs that expressed Ki-67 were predominantly dim or nonperivascular (Fig. 7A–F, dashed circles) and were not bright-perivascular LRCs (solid circles). At DOPP 7.5, when the highest proliferation was shown to occur by EdU incorporation (Fig. 6E), there was also an abundance of Ki-67-expressing LRCs both dim and bright. Interestingly, at INV 48 h bright-perivascular LRCs were now shown to express Ki-67 (Fig. 7J–L, solid circles), as compared with DOPP 0.5 and 4 when they did not. These results suggest that both dim and bright LRCs contributed to myometrial expansion during pseudopregnancy (DOPP 7.5), but that bright-perivascular LRCs preferentially proliferated during uterine INV (ie, postpartum uterine repair) in contrast to the early time points during pseudopregnancy.
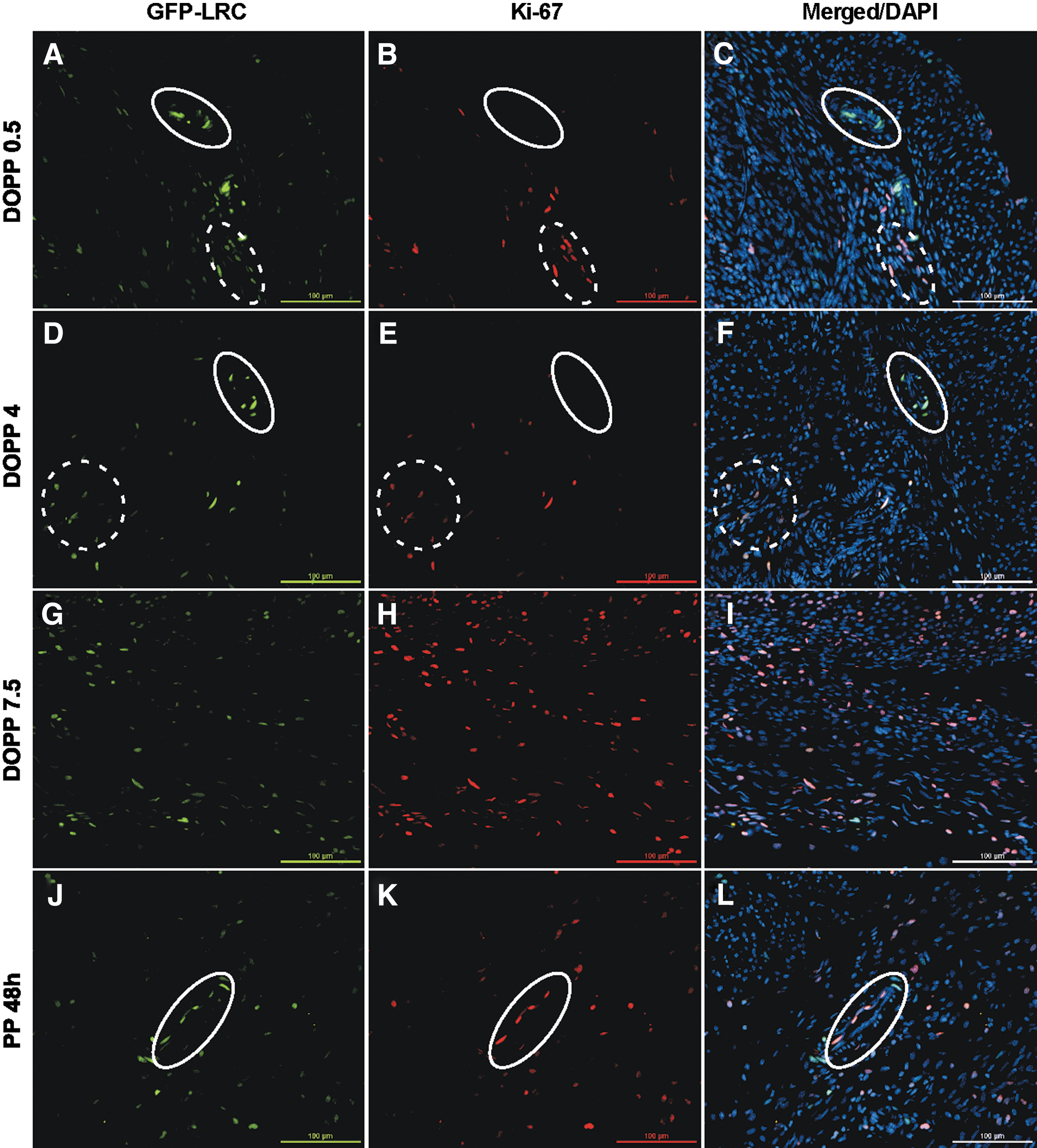
Bright-perivascular LRCs preferentially proliferate during uterine INV. Representative (n = 3/time point) images of uterine cross-sections showing expression of Ki-67 in different LRC populations corresponding to the pseudopregnancy and uterine INV timeline in Fig. 6.
Discussion
In some species, the uterus increases in size up to 40-fold during pregnancy, which is largely attributed to expansion of the myometrium by hyperplasia and hypertrophy [3,4]. After pregnancy, the uterus undergoes INV, the process of returning to its nongravid state, which includes repair of the tissue that was remodeled or “damaged” during pregnancy. Little is known about this repair process in the myometrium, particularly in regard to the role that stem/progenitor cells may play. Previous reports suggest the existence of stem/progenitor cells in human myometrium and their possible contribution to uterine fibroids (benign tumors of the myometrium) [23,36 –42]. In a prior study from our laboratory, we used BrdU label retention to identify myometrial LRCs and assessed their response to gonadotropin treatment [23]. In this study, we show for the first time the presence of myometrial LRCs after embryonic and postnatal labeling and their activity in the adult during pseudopregnancy and uterine INV.
We began by looking for cells that coexpressed CD146 and CD140b during differentiation of the postnatal uterine mesenchyme into the endometrial stroma and myometrium. CD146 and CD140b, among other surface markers, have been used to identify and isolate MSCs [reviewed in Refs. 14,43]. These markers have been commonly used to mark perivascular cells in various tissues, including the endometrial stroma in humans [17], and in many tissues, MSCs are known to localize to the perivascular space [reviewed in Refs. 12 –14]. Interestingly, we found both CD146 and CD140b to be ubiquitously expressed in the mesenchyme at P0.5, which is when the uterus was still undifferentiated. Then as maturation and differentiation progressed, the expression patterns of CD146 and CD140b became more restricted, so that coexpressing cells in the adult uterus were predominantly located in the perivascular space. Because expression of certain surface markers does not definitively identify stem cells in a tissue, we employed a second technique, label retention, to further characterize putative myometrial MSCs.
Label retention, although not a definitive marker of stem cells, has often been used in various tissues to identify slowly dividing cells with stem cell qualities. Historically, the label used was BrdU; however due to the caveats associated with this label, including toxicity and the inability to recover live BrdU-labeled LRCs, we used a transgenic approach [24]. Using “tet-off” transgenic mice in combination with the Cre-lox system, we were able to specifically label the mesenchymal cells within the uterus while avoiding other cell types (eg, epithelial, immune, and endothelial cells). In addition, we were able to label cells embryonically during Müllerian duct development when there is presumably a high proportion of stem cells, to increase the likelihood of identifying true MSCs as opposed to transient amplifying cells. Based on the expression patterns of CD146 and CD140b (Fig. 1), we continued the labeling period out to P 28 to capture potential stem cells during postnatal mesenchyme differentiation and maturation into the adult uterus. At P 28, the chase period was initiated to allow dilution of label. Importantly from P 28–42, the uterus substantially increases in size as the mouse reaches sexual maturity and produces E2 and P4. Because the adult uterine histoarchitecture is established by P 15–25 [44,45], we hypothesized that the uterine growth that occurred from P 28 to 42 would be predominantly due to transient amplifying cells and other nonstem cells. The goal was to dilute out label during this uterine expansion and subsequent estrous cycles, thereby allowing us to identify and characterize LRCs and assess their stem cell qualities. Our results show that we were able to successfully dilute label by ∼40% with 1 month of chase (Fig. 1M, 1 month) and ∼80% with 2 months of chase (Fig. 1M, 2 months). The remaining LRCs after 3 months of chase could predominantly be classified as dim and/or nonperivascular or as bright and perivascular LRCs.
Upon further examination, the bright-perivascular LRCs did not express ERα or PR (Fig. 4), which is consistent with what has been shown for putative human myometrial stem/progenitor cells [39,40] as well as other stem-like cells in the female reproductive tract [25,46,47]. Interestingly, Janzen et al. showed that although endometrial epithelial stem-like cells do not express ERα and PR, they were hormonally responsive suggesting a paracrine signaling mechanism with niche cells [46]. In our study, the surrounding dim/non-LRCs expressed ERα and PR, indicating that they may be responsive to hormones and could possibly signal to bright-perivascular LRCs. Unlike the aforementioned study, E2 and P4 treatment did not change the overall LRC population (Supplementary Fig. S2), but this could be due to proliferation of brighter LRCs into dim LRCs combined with proliferation of dim LRCs into non-LRCs, leading to equilibration of the LRC population. Importantly, bright-perivascular LRCs were positive for CD146 and CD140b (Fig. 5), which have been used to isolate mesenchymal stem-like cells in tissues, including bone marrow [48 –50], adipose [51,52], and endometrial stroma [17]. In addition, decreased expression of CD146 has been associated with differentiation [49] and senescence [53] of MSCs. A study by Espagnolle et al. suggested that CD146(high) bone marrow-derived MSCs, although they exhibit similar colony-forming and multilineage differentiation potential to CD146(-/low) MSCs, are uniquely suited for myogenic differentiation [54]. Bright-perivascular LRCs did not express CD44, a marker that along with Stro-1 has been shown to be expressed by putative human myometrial stem/progenitor cells [36]. CD44 was, however, expressed by some very dim LRCs that were often next to brighter LRCs (Supplementary Fig. S3). We speculate that these CD44+ dim LRCs might be transient amplifying cells that were derived from the brighter adjacent LRCs and underwent subsequent divisions resulting in dilution of label. Therefore, because bright-perivascular LRCs expressed CD146 and CD140b, and not ERα and PR, and were among the brightest cells in the tissue, revealing their quiescent nature, we speculate that they are less differentiated and are good candidates for further testing of stem cell qualities by in vitro and in vivo analyses.
In the pseudopregnancy model, we showed that the peak of myometrial proliferation occurred at DOPP 7.5 (Fig. 6), to accommodate the expanding decidual tissue, and that non-LRCs and LRCs contributed equally to this myometrial growth. Further, it appeared that both dim and bright LRCs participated in that expansion as they both expressed Ki-67 at that time (Fig. 7). Because of the rapid expansion of this tissue from DOPP 4 to 7.5, it is likely that both stem/progenitor cells (possibly bright LRCs) and more differentiated transient amplifying cells (possibly dim and non-LRCs) proliferated during that time to expand the tissue. Outside of DOPP 7.5, the LRCs contributed to the proliferating cell population more than non-LRCs. This was particularly true for INV 48 h. When the LRC population was further evaluated by Ki-67 expression, we observed that bright-perivascular LRCs appeared to proliferate at INV 48 h but not at earlier time points, which was opposite for dim/nonperivascular cells that expressed more Ki-67 at the earlier time points (Fig. 7). It is tempting to speculate that the dim LRCs are transient amplifying cells and proliferated during early and mid-pseudopregnancy, whereas the bright-perivascular LRCs may be stem/progenitor cells whose proliferation was reserved for repair during INV. This would make sense if the myometrium needs to replace the terminally differentiated cells during postpartum repair (INV) by contribution from stem/progenitor cells.
We show based on label retention, MSC marker expression, and activity during pseudopregnancy and uterine INV that the bright-perivascular LRCs identified in this study are good candidates for being myometrial stem/progenitor cells. It is important to note however that these techniques, although suggestive of stem cell nature, do not fully confirm it. These cells therefore require further characterization and functional testing to confirm their possible stem/progenitor cell identity.
Conclusion
Using a combination of MSC markers and label retention, we identified a population of bright-perivascular myometrial LRCs that expressed CD146 and CD140b. These cells did not appear to be directly responsive to E2 and P4 since the classical receptors for these hormones were not detected. The bright-perivascular LRCs persisted in the myometrium for at least 3 months, and were shown to proliferate during pregnancy and postpartum INV in a model that simulates these two important uterine functions (Fig. 8). The results from this study provide important insight into putative stem/progenitor cells in the myometrium and their possible role in uterine physiology.
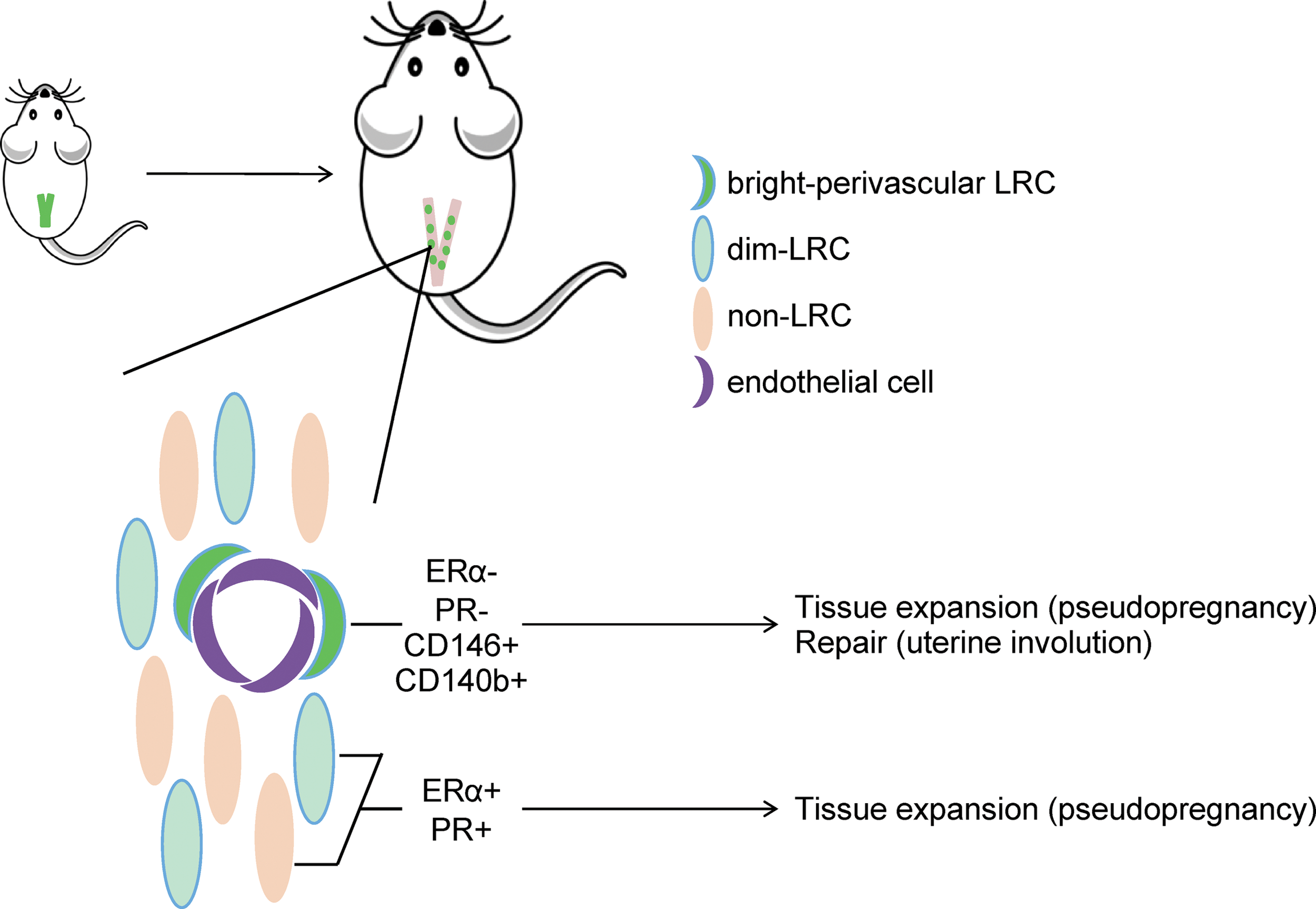
Graphical representation of major conclusions. Using a transgenic label-retaining mouse model, LRCs were identified in the myometrium (uterine smooth muscle) and persisted for at least 3 months. Two distinct populations of LRCs were found, bright-perivascular LRCs surrounding endothelial cells and randomly distributed, dim LRCs. Bright-perivascular LRCs expressed the MSC and perivascular cell markers, CD146 and CD140b, and contributed to both tissue expansion and uterine INV in a model of pregnancy and postpartum repair. Color images available online at
Footnotes
Acknowledgments
Funding for this study was provided by the NIH: R01 OD012206 and F32 HD081856. We thank the University of Virginia Center for Research in Reproduction Ligand Assay and Analysis Core, supported by the Eunice Kennedy Shriver NICHD/NIH P50 grant (HD28934), for performing the serum E2 and P4 hormone assays.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.