
Abstract
The histone chaperone facilitates chromatin transactions (FACT) is associated with nuclear processes, including DNA transcription, replication, and repair. We previously showed that FACT is transiently recruited to pluripotency-associated target genes by newly bound Oct4. In this study, we tested the effects of FACT depletion by knockout or chemical inhibition on the induction and maintenance of pluripotency. Clustered regularly interspaced short palindromic repeat (CRISPR)-mediated deletion of the FACT subunit Spt16 did not affect the viability or proliferation of fibroblasts but blocked their ability to form induced pluripotent stem cells. Similarly, a small molecule inhibitor of FACT blocked the induction of pluripotency at an early step in reprogramming, without affecting the viability, proliferation, undifferentiated state, or the expression of core pluripotency genes. Notably, trypsinization and passage of pluripotent cells transiently reintroduced a requirement for FACT. Although FACT has been considered to be an essential transcription elongation factor, these results contribute to the emerging view that it instead promotes transitions between stable chromatin states, including during reprogramming to pluripotency.
Introduction
T
Although abundant in yeast and undifferentiated stem-like and malignant mammalian cells, and broadly considered to be an essential transcription elongation factor, FACT is nearly undetectable in most adult mammalian tissues [6,7]. Furthermore, it is dispensable for viability and growth in many cell types, but is required for efficient transformation and growth of malignant cells [6]. Although FACT physically associates with highly transcribed genes [1], FACT is recruited to specific target genes during induction of new gene expression in both yeast and metazoans [2,3,8 –11]. These results suggest that FACT also has a role in nontranscribed promoter regions during transcription activation, and may localize to transcribing gene bodies primarily to restore chromatin.
FACT inhibitors have been developed [12], and consistent with a less general role supporting transcription, they are selectively toxic to malignant cells in a variety of situations such as glioblastoma [13 –15]. These inhibitors also block induction of new viral gene expression and, therefore, may function as effective antivirals [16]. Taken together, these results point away from a role for FACT as a general transcription factor and instead suggest that it has functions during activation of specific silent genes.
FACT physically interacts with the sequence-specific transcription factor Oct4 [17,18], a key activity involved in maintenance [19] and establishment [20] of pluripotency. Oct4 also interacts with multiple other cofactors, including Jmjd1c and NuRD [17]. Upon initial binding to the Pou5f1 (Oct4) gene, Oct4 first recruits Jmjd1c to mediate loss of H3K9me2, and subsequently recruits FACT in a step concomitant with local histone H3 depletion [21]. FACT association peaks 10 h after induction of Oct4 and 4 h after detectable Oct4 enrichment at Pou5f1. FACT association is transient, with signal quickly gained and lost over the course of 4 h.
ChIPseq experiments identify multiple pluripotency-associated genes bound in this manner, including Pou5f1, Nanog, and Sox2 [21]. Blocking H3 demethylation pharmacologically blocks FACT recruitment and H3 depletion. These findings indicate that FACT mediates Oct4 functions at newly bound but not at established Oct4 targets to mediate nucleosome eviction in support of new gene expression and, therefore, that FACT may be required for establishing pluripotency but dispensable for its maintenance.
In this study, we targeted FACT using clustered regularly interspaced short palindromic repeat (CRISPR)/Cas9-mediated knockout or chemical inhibition. We found that loss or inhibition of the FACT subunit Spt16 did not affect viability or proliferation of primary mouse embryonic fibroblasts (MEFs), but severely impaired their ability to undergo reprogramming to pluripotency. To determine whether this effect was due to the inability to establish a pluripotent state or to maintain it once established, we acutely inhibited FACT using the compound CBL0100 [12] either in MEFs undergoing reprogramming, or in established pluripotent cells.
We found that inhibiting FACT blocked the ability to induce pluripotency during a specific window early in the reprogramming process. However, FACT was dispensable for both the viability and the maintenance of an undifferentiated state in established embryonic stem cells (ESCs) and in induced pluripotent stem cells (iPSCs). Notably, however, passage of these cells by trypsinization transiently reintroduced a requirement for FACT. Cumulatively, the data point to a specific requirement for FACT during the reprogramming process, suggesting that it has an important role during transitions between differentiated and pluripotent transcriptional states but is less important for proliferation.
Materials and Methods
Cell culture
3T3 cells and primary MEFs were cultured in Dulbecco's modified Eagle's medium plus 10% fetal bovine calf serum (Sigma), 100 U/mL penicillin, 100 μg/mL streptomycin, and 2 mM
Clustered regularly interspaced short palindromic repeat
Mouse Supt16 guide RNAs (gRNAs) were designed to target Exon 1 or 6. gRNAs were cloned into lenti-CRISPR-v2-mCherry, based on lenti-CRISPR-v2 [23] but modified to express mCherry. gRNA sequences (with PAM) were as follows: Spt16Exon1, AGAGATTGTACAGTAACTGGCGG; Spt16Exon6, CCACCACTCTGAATGATAGGAGG; luciferase, ACAACTTTACCGACCGCGCCCGG. Constructs were transfected into 293T cells and viral supernatant was collected at 48 h post-transfection. Virus was concentrated with Lenti-X-Concentrator (Clontech), according to the vendor instructions. Viruses were used with 6 μg/mL polybrene to infect target MEFs.
iPSC reprogramming
Early-passage MEFs (P3) were infected with viral particles expressing mouse Oct4, Sox2, Klf4, and c-Myc. The mSTEMCCA lentiviral construct [24] was used as published [21]. Images were taken using a Nikon A1R confocal microscope and a 40× objective.
Reverse transcriptase-quantitative polymerase chain reaction
Sox2, Nanog, and Oct4 (Pou5f1) gene expression was analyzed by reverse transcriptase-quantitative polymerase chain reaction (RT-qPCR) as previously published [25]. Primers were identical to those used previously [25].
Results and Discussion
Supt16 knockout does not affect fibroblast survival or growth but blocks reprogramming to pluripotency
To study the role of FACT in reprogramming, we knocked out Supt16, which encodes the FACT subunit Spt16. We did not manipulate the other FACT subunit, SSRP1, as it is known to have FACT-independent functions [26]. We used a lentiviral construct encoding Cas9, a firefly luciferase-specific control gRNA, or either of two Supt16-specific gRNAs that target exon 1 or exon 6. mCherry encoded by the vector allowed identification and sorting of productively infected cells.
Using 3T3-immortalized MEFs, Spt16 protein levels were nearly eliminated by infection with either Supt16-specific construct compared to control gRNAs (Fig. 1A). Immortalized cells lacking Spt16 remained viable and morphologically normal, similar to luciferase controls and uninfected mCherry- cells in the same culture (Fig. 1B). They also proliferated normally, as sorted mCherry+ cells doubled at the same rates after infection with either Supt16 knockout or control luciferase constructs (Fig. 1C). Furthermore, the ratio of mCherry+ to mCherry- cells did not change over multiple passages (not shown).

CRISPR-mediated reduction of the FACT subunit Spt16 allows for proliferation but blocks formation of iPSCs.
These results indicate that FACT is dispensable for viability and growth of MEFs under standard culture conditions. This finding is surprising given the widely held view that FACT is an essential transcription elongation/replication factor, but is consistent with studies of FACT levels in cultured human cells [27] and genetic characterization of FACT mutants in zebrafish and plants [27,28], indicating that FACT does not have an essential function in differentiated cells.
To test the effect of loss of FACT on reprogramming, we transduced primary, early-passage Oct4-GFP MEFs [22] with CRISPR/gRNA lentiviruses, together with lentiviruses encoding the four mouse Yamanaka factors Oct4, Klf4, Sox2, and c-Myc [24]. Using this system, Oct4-GFP MEFs become GFP+ upon successful reprogramming, whereas mCherry fluorescence can be used to identify control or Spt16 CRISPR-infected cells. Spt16 expression was again efficiently ablated in primary cells (Fig. 1D) with no effect on growth, as sorted mCherry+ cells did not display differences in growth rates over three additional passages (Fig. 1E).
Although treatment with Spt16-specific CRISPR lentiviruses had little effect on the emergence of GFP+, mCherry- colonies (Fig. 1F), productive infection strikingly eliminated GFP+ iPSC colony emergence, as no GFP+, mCherry+ colonies were observed (Fig. 1G). Example images are shown in Fig. 1H. This result was confirmed by picking iPSC colonies, plating in differentiation media to eliminate GFP fluorescence, and verifying the absence of mCherry fluorescence (not shown). A separate set of Oct4-GFP MEFs were infected in parallel with only the CRISPR constructs. These confluent cells were imaged after 10 days for mCherry expression to confirm equivalent initial infection (Fig. 1I). These results show that FACT is not needed for viability or proliferation of differentiated fibroblasts, but is either required during reprogramming or for subsequent viability or proliferation of iPSCs.
Acute FACT inhibition blocks iPSC formation during a specific window early in the reprogramming process
To distinguish between a requirement for establishment versus maintenance of pluripotency, we repeated the iPSC formation assays using Oct4-GFP MEFs in the presence of a small molecule FACT inhibitor, CBL0100 [12]. CBL0100 reversibly inhibits FACT function. In ESC culture conditions, high doses of CBL0100 (>15 nM) were toxic to nonmitotic feeder fibroblasts. A concentration of 10 nM did not have an effect and was, therefore, used in subsequent experiments. As with the genetic knockout of FACT, continuous administration of CBL0100 throughout the iPSC protocol timecourse eliminated the formation of GFP+ colonies with pluripotent iPSC morphology, whereas treatment with dimethyl sulfoxide (DMSO) vehicle control resulted in ∼100 GFP+ colonies within 10 days of infection (Fig. 2A).
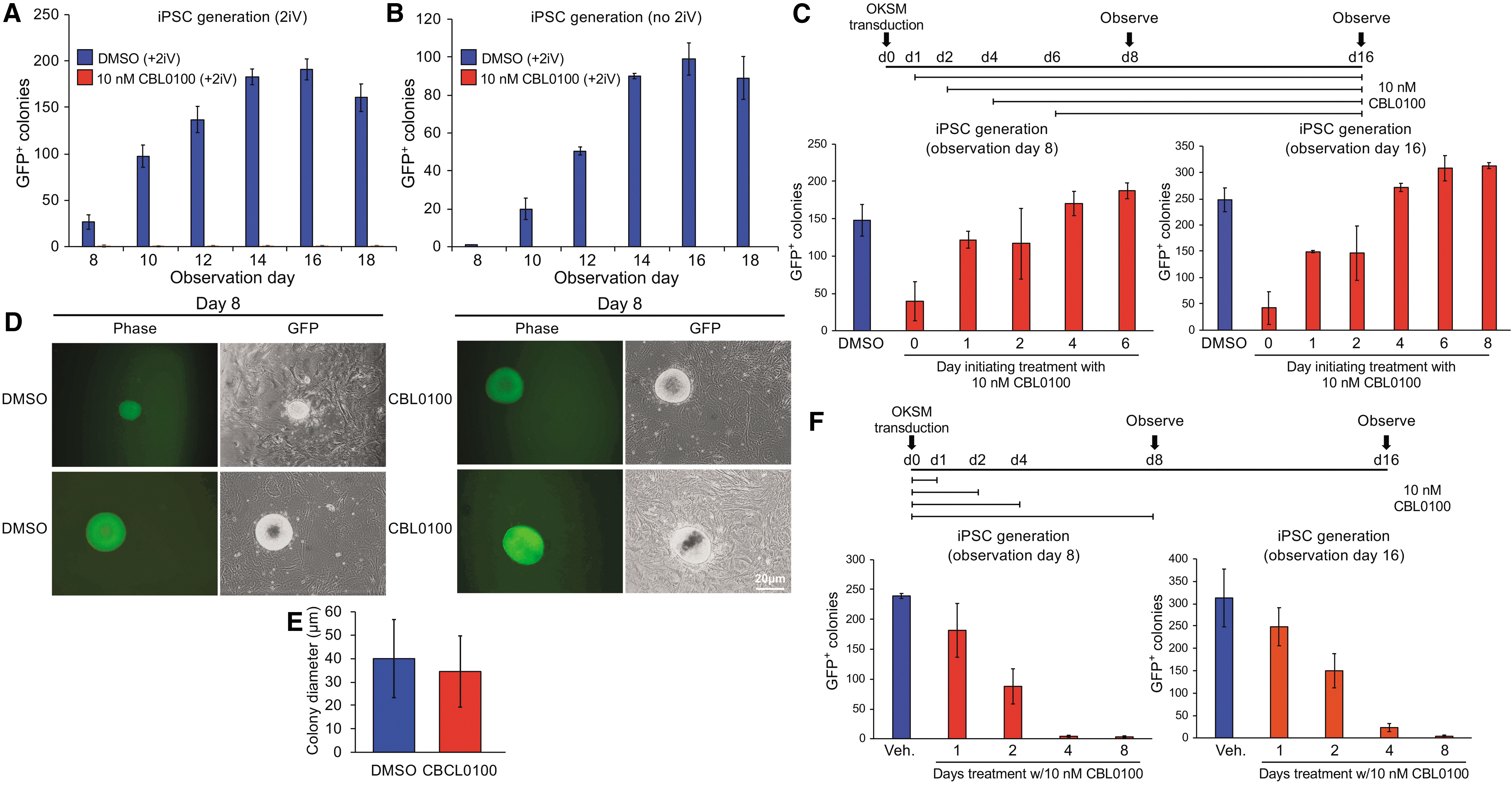
Chemical inhibition of FACT blocks emergence of iPSC colonies from primary MEFs.
Our standard (“2iV”) culture conditions for ESCs and iPSCs include GSKβ and MEK inhibitors, and
To determine whether FACT is required during a specific window of iPSC reprogramming, we administered CBL0100 at different times after infection with the Yamanaka reprogramming gene cocktail. Surprisingly, CBL0100 administration starting at 4 days postinfection had no effect on the emergence of GFP+ iPSC colonies (Fig. 2C), indicating that FACT is only required during the first 4 days of reprogramming. Examples of iPSC colonies generated at day 8, with FACT inhibitor treatment beginning at 6 days, are shown in Fig. 2D. The average size of generated colonies was also the same with and without inhibitor, showing that proliferation rates were unaffected by CBL0100 (Fig. 2E). Similar results were obtained at 16 days (not shown).
In a reciprocal experiment, we administered CBL0100 on the day of infection and determined the length of sustained treatment required to inhibit colony formation. Four days of treatment was sufficient to completely block colony formation, whereas 1 day of treatment had only minimal effects (Fig. 2F). Collectively, the data indicate that FACT is required during a 72-h window beginning 1 day after infection with lentiviral vectors expressing the Yamanaka reprogramming factors to successfully reprogram cells to pluripotency. Multiple epigenetic and transcriptional changes critical to the reprogramming process are known to occur within this window [31 –33]. This result, therefore, suggests that the requirement for FACT in reprogramming may reflect a role in controlling transitions between chromatin states, including the activation of repressed genes associated with pluripotency. Additional work will be required to thoroughly test this model.
FACT is dispensable for maintenance of pluripotency in the absence of passage
The finding that iPSCs could be generated with FACT inhibition late in the reprogramming process suggested FACT is dispensable for the viability and maintenance of pluripotent cells. To test this more carefully, we treated ESCs established in culture with CBL0100. No differences in growth rate (Fig. 3A) or morphology (Fig. 3B) were observed. After 5 days in culture with CBL0100, we prepared cDNAs to study the expression of genes encoding the pluripotency-associated transcription factors Sox2, Oct4, and Nanog. These were expressed at normal levels compared to vehicle controls (Fig. 3C). Similar results were obtained using a single trypsinized and plated GFP+ iPSC colony (Fig. 3D–F). These results indicate that FACT inhibition does not affect the proliferation of pluripotent cells maintained in culture. These findings are surprising given that FACT associates with ∼1,000 target sites in ESCs [21], but are consistent with the observation that FACT is dispensable for viability in established human ESCs [34].

Inhibition of FACT does not affect either the growth or undifferentiated state of established pluripotent cells.
The earlier experiments were performed by plating cells to allow adherence and treating with the inhibitor after 24 h in culture. Notably, treatment with CBL0100 before this time point, during a window 0–16 h after plating, eliminated pluripotent colony formation (Fig. 4A). The underlying feeder cells were unaffected (Fig. 4B). Similar results were obtained using a single GFP+ iPSC colony that was picked, trypsinized, and replated (Fig. 4C). The toxicity associated with passage in the presence of CBL0100 could be largely eliminated by plating 2.5 × 105 ESCs in a six-well dish rather than the standard 5 × 104 cells (data not shown). This change could result from dilution, degradation, or accelerated metabolism of the drug. These results indicate that pluripotent cells required FACT transiently during a 16 h window to recover from the effects of passaging.
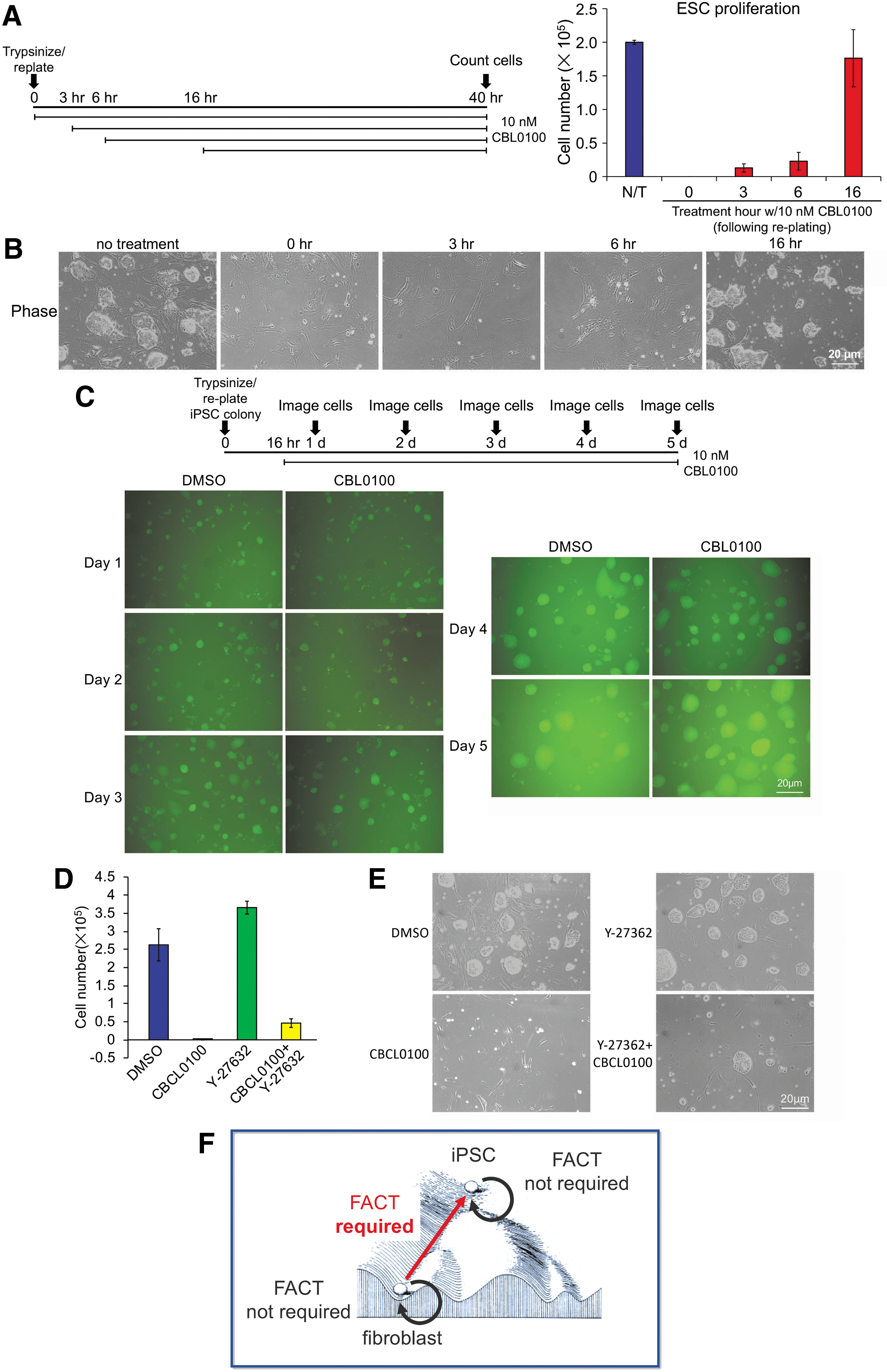
Passaging pluripotent cells introduces a transient requirement for FACT.
Inhibition of Rho-associated kinase (ROCK), which regulates the cytoskeleton, cell polarity, and apoptosis, has been shown to improve ESC colony generation after passage [35,36]. We used ROCK inhibition with a range of Y-27632 concentrations together with CBL0100 to determine if the effects of FACT inhibition could be reversed. No effect was observed at lower concentrations (not shown), and minimal effects at higher (10 μM) concentrations (Fig. 4D,E). This result suggests that the requirement for FACT in a brief window after passage is unlikely to be mediated through pathways associated with ROCK.
Knockdown of either FACT subunit has been reported to be deleterious to mouse ESCs in prolonged culture [37]. Our finding that FACT is required to sustain ESC maintenance immediately after passage of mouse ESCs or iPSCs can explain this prior result. The transient toxicity associated with FACT inhibition after passage may indicate a requirement for FACT in responding to or recovering from the effects of trypsinization or replating to reestablish growth. Cumulatively, the data indicate that FACT is dispensable for proliferation and for maintenance of differentiated and pluripotent states, but is required at an early stage in the reprogramming process, and for recovery from the stress caused by passaging (Fig. 4F).
A recent study focused primarily on Caenorhabditis elegans development also concluded that FACT is dispensable for maintenance and growth of pluripotent cells; however, this study reported that iPSC formation from telomerase reverse transcriptase (TERT)-immortalized human fibroblasts was moderately enhanced by FACT knockdown [38]. Multiple experimental system differences could explain these different outcomes. These include: the use of telomerase reverse transcriptase (TERT)-immortalized human fibroblasts versus primary mouse fibroblasts, the use of pre-integrated, dox-inducible human reprogramming factors, the lack of feeder cells, transient FACT siRNA transfection and partial mRNA ablation rather than deletion or durable inhibition, and lastly scoring SSEA+ colonies rather than colonies expressing endogenous Oct4. Given our result that the role of FACT is confined to a short time window after passage, perhaps the most likely explanation is the use of transient siRNA transfection in the study by Kolundzic et al. Without analysis of FACT protein levels, it is difficult to assess the level or timing of depletion in this study, but ultimately the discrepancy may reveal the enigmatic functions of FACT, which can both destabilize and assemble nucleosomes in vitro.
Current models suggest that FACT assists the disassembly of nucleosomes to allow transcription factor access to blocked sites during activation of transcription and to promote progression of polymerases during transcription elongation. Recent results showing that FACT is primarily recruited to chromatin that has been disrupted by transcription [5], and multiple reports, including data presented in this study, show that FACT's roles in disassembling and restoring chromatin need to be reevaluated. FACT's ability to assemble nucleosomes could be important for providing barriers to fate switching, and its ability to destabilize nucleosomes could be important for making transitions to new transcription profiles. Our results not only stress the latter role but also highlight the need for further work to establish the range of processes affected by FACT.
Footnotes
Acknowledgments
We thank D. Stillman and K.V. Gurova for critical reading of the article. We thank T. Dahlem, C. Davey, and the University of Utah Health Sciences Center Mutation Generation and Detection Core Facility for assistance with CRISPR lentiviral constructs, as well as C. Rodesch and the Fluorescence Microscopy Core facility for help with imaging, and J. Marvin and the Flow Cytometry Core facility for help with cell sorting. The mSTEMCCA 4-in-1 lentiviral construct was obtained from G. Mostoslavsky. This work was supported by an Endowed Chair (Watkins Endowed Chair, to D.T.) and NIH grants R01GM064649 to T.F. and R01GM122778 to D.T.
Author Disclosure Statement
No competing financial interests exist.