
Abstract
Human induced pluripotent stem cell-derived cardiomyocytes (hiPSC-CMs) are potential sources for cardiac regeneration and drug development. hiPSC-CMs express all the cardiac ion channels and the unique cardiac Ca2+-signaling phenotype. In this study, we tested for expression of acid sensing ion channels (ASICs) in spontaneously beating cardiomyocytes derived from three different hiPSC lines (IMR-90, iPSC-K3, and Ukki011-A). Rapid application of solutions buffered at pH 6.7, 6.0, or 5.0 triggered rapidly activating and slowly inactivating voltage-independent inward current that reversed at voltages positive to ENa, was suppressed by 5 μM amiloride and withdrawal of [Na+]o, like neuronal ASIC currents. ASIC currents were expressed at much lower percentages and densities in undifferentiated hiPSC and in dermal fibroblasts. ASIC1 mRNA and protein were measured in first 60 days but decreased in 100 days postdifferentiation hiPSC cultures. Hyperacidification (pH 5 and 6) also triggered large Ca2+ transients in intact hiPSC-CMs that were neither ruthenium red nor amiloride-sensitive, but were absent in whole cell-clamped hiPSC-CMs. Neither ASIC1 current nor its protein was detected in rat adult cardiomyocytes, but hyperacidification did activate smaller and slowly activating currents with drug sensitivity similar to TRPV channels. Considering ASIC expression in developing but not adult myocardium, a role in heart development is likely.
Introduction
Since the advent of cellular reprogramming of adult somatic cells and development of cardiomyocytes from human induced pluripotent stem cells (hiPSCs) some 10 years ago [1 –4], comprehensive electrophysiological studies have confirmed the expression of almost all of the cardiac ion channels and carriers in human induced pluripotent stem cell-derived cardiomyocytes (hiPSC-CMs) as well as the calcium signaling events specific to mammalian cardiomyocytes [5 –8]. There are, however, no reports as yet that hiPSC-CM might express other ionic channels not functionally expressed in adult mammalian myocardium. It was, therefore somewhat unexpected when in our study acidic solutions activated acid-sensitive ionic channel (ASIC) currents on rapid application of low pH solutions on hiPSC-CMs.
ASIC family of proteins are proton-gated cation channels [9] that are abundantly expressed in the central and the peripheral nervous system, which have high permeability to monovalent cations [10 –12] and been implicated in a variety of neuronal signaling events, including pain perception [13,14]. Six members of ASIC gene family have been, thus far, identified that include ASIC1a, ASIC1b, ASIC2a, ASIC2b, ASIC3, and ASIC4 [15 –17]. The ASIC family members are generally distinguished, based on their pharmacology, kinetics, ionic selectivity, and the acid concentration levels that activate them. For instance, ASIC1a and ASIC1b are characterized by rapid kinetics of inactivation, ASIC2a shows slow kinetics of inactivation, and ASIC3 has a rapid phase of inactivation followed by a noninactivating plateau phase. PcTx1 and APETx, isolated from spider and sea anemone venoms, have been identified as selective high-affinity blocker of ASIC1a and ASIC3, respectively [18 –21].
In this study, for the first time, we report on time course of development of ASICs mRNA and the proteins encoded by them in the first 100 days of postcardiac differentiation in three different hiPSC lines. Molecular (mRNA, immunocytochemistry) and functional (electrophysiological measurements of ionic currents) studies showed significant levels of ASIC expression in 15–60 day cultures that decreased in older 100 days postcardiac differentiation cultures. Electrophysiologically, modest protonation (pH 6.7 to 6.0) activated a voltage-independent inward current carried primarily by Na+ that was suppressed by amiloride, in CMs from all three hiPSC lines, similar to proton-gated monovalent currents described in dorsal root ganglion and CNS neurons [10,22,23]. We failed to find immunoflorescence evidence for ASIC proteins and their encoded ionic currents, with ASIC1 kinetics and pharmacology, in adult or neonatal rat cardiomyocytes. Our findings did, however, show that hyperprotonation (pH 5 and 6) activates small and slowly activating and inactivating currents, insensitive to amiloride, which could be mistaken for ASIC-type channels. Independent of presence or expression of these small currents, hyperprotonation triggered large and slowly activating and decaying calcium transients that were insensitive to 5–10 μM concentrations of amiloride, IP3, and TRPV1 channel blockers, suggestive of a novel protons-activating Ca2+-signaling pathway.
Materials and Methods
Culture of human iPSs
The human iPSc lines IMR-90 and UKKi011-A (NP0040,
Method 1: The undifferentiated hiPSCs were maintained on mitomycin-C-treated murine embryonic fibroblasts (MEFs) that were isolated from embryonic day 14 embryos of a CF1 mouse strain. Mitomycin-C-treated MEFs were plated at a density of 0.2 × 106 cells/6 cm plate 1 day before seeding hiPSCs. hiPSCs were cultured in DMEM/F12 medium supplemented with GlutaMAX, 20% knockout serum replacement, 1% nonessential amino acids (NAA), 0.1 mM β-mercaptoethanol (βME; Invitrogen, Darmstadt, Germany), and 50 ng/mL FGF-2 (PeproTech, Hamburg, Germany). Method 2: Undifferentiated hiPSCs were maintained in Essential 8 medium (Gibco) on Matrigel (BD Biosciences) or Vitronectin- (Thermo Fisher Scientific, Waltham, MA) coated culture dishes at 37°C with 5% CO2. hiPSC cell culture medium was changed every 2 days and passaged by manual dissection of cell colonies or by dissociation with Versene (Thermo Fisher Scientific) every 4–6 days.
Differentiation of hiPSCs to cardiomyocytes
Cardiac differentiation was carried out by two standard techniques. Method 1: Coculture of END2 and hiPSCs [1]. hiPSC colonies were dissociated into clumps by using collagenase IV (1 mg/mL in DMEM/F-12 at 37°C for 5–10 min; Sigma-Aldrich) and then cocultured with murine visceral endoderm-like cell line END2. END2 cells were mitotically inactivated for 3 h with 10 μg/mL mitomycin C (Sigma-Aldrich Chemie GmbH, Munich, Germany) and 1.2 × 106 cells were plated on 60 mm dishes coated with 0.1% gelatin 1 day before coculture with hiPSC. The differentiation was carried out in 1% knockout-DMEM containing 1 mM L-glutamine, 1% NAA, 0.1 mM βME, and 1% penicillin/streptomycin (100 U/mL and 100 μg/mL, respectively). The coculture was left undisturbed at 37°C, 5% CO2 for 5 days. First medium change was performed on day 5 and later on days 9, 12, and 15 of differentiation. Spontaneously contracting clusters were dissociated into single hiPSC-CM for electrophysiological experiments. For long-term cultures, the medium was changed every 3–5 days and beating cardiac clusters were isolated by microdissection on day 30 or day 100 of differentiation for isolation of total RNA.
Method 2: Directed differentiation using small molecules. The cardiac differentiation was followed by the standard protocol from literature [26]. In brief, dissociated hiPSCs were plated on Matrigel-coated 6-well or 12-well plates and confluent cultures treated for 24 h with 8–12 μM CHIR99021 (a GSK-3 inhibitor, Cat. No. C-6556; LC Laboratories) in RPMI 1640 medium (Cat. No. 11875119; Fisher Scientific) supplemented with B-27 without insulin (Cat. No. A18956-01; Thermo Fisher Scientific). Exactly 24 h after addition of CHIR99021, the medium was changed to RPMI 1640 containing 1 × B-27 supplement without insulin. Seventy-two hours after addition of CHIR99021, Wnt signaling inhibitor IWP2 (5 μM) was added in the same medium for 48 h, then changed to RPMI/B-27 without insulin. RPMI/B-27 with insulin medium was applied when the spontaneously beating myocytes appeared and was replaced every 3 days until analysis. Spontaneously contracting clusters were then dissociated into single cardiomyocytes for electrophysiological experiments.
Preparation of hiPSC-CM for patch-clamp experiments
Spontaneous beating hiPSC-CM colonies were microdissected mechanically at day 15–60 of differentiation. Collagenase, Type IV (Worthington biochemical corporation) was used at 1 mg/mL in low Ca2+ solution to dissociate the beating clusters, and single hiPSC-CM were then plated on 25 mm glass coverslips coated with fibronectin (2.5 μg/mL). Cells were incubated in IMDM supplemented with 20% fetal bovine serum in the incubator (37°C, 5% CO2) for 3 days before their use in electrophysiological experiments.
Adult rat ventricular myocytes preparation
Protocols for obtaining rat cardiac tissue were approved and supervised by Institutional Animal Care and Use Committees (IACUC) of Medical University of South Carolina and the Department of Veterans Affairs according to national legislation and guidelines as we described before [27,28]. Ventricular cardiomyocytes were isolated from adult male Sprague–Dawley rats weighing 200–300 g at age of 8–12 weeks (Envigo). Rats were deeply anesthetized with isoflurane (3%, vaporized) and hearts were rapidly excised, cannulated, and installed on a Langendorff perfusion system and enzymatically digested by a mixture of a mixture of collagenase NB8 (SERVA) and protease type XIV (Sigma). For detailed protocol, see our previous publication [28].
Electrophysiological recordings of ASIC currents
hiPSC-CMs or adult ventricular myocytes were voltage-clamped in the whole-cell configuration with an Dagan amplifier (Axon Instruments) interfaced with a Digidata 1320A (Axon Instruments). Borosilicate glass capillaries were pulled using a Sutter P-87 micropipette puller with an electrical resistance of 4–5 MΩ. Changes in extracellular pH were induced by a rapid shift of the outlets of a microperfusion system from pH 7.4 to 6. The voltage-clamped cells were superfused with Tyrode's solution and dialyzed with a Cs+-based, moderately Ca2+-buffered pipette solution containing (in mM) the following: 110 Cs+-Aspartate, 15 NaCl, 20 TEACl, 5 Mg-ATP, 0.2 EGTA (2EGTA for adult ventricular cells) and 0.1 CaCl2, 10 glucose, and 10 HEPES (titrated to pH 7.2 with CsOH; measured osmolality: 290 mOsm). The continuously superfused extracellular Tyrode's solution contained (in mM) 137 NaCl, 5.4 KCl, 2 CaCl2, 1 MgCl2, 10 glucose, and 10 HEPES (titrated to pH 7.4 with NaOH). Electrophysiological experiments were performed at 25°C–32°C.
Ca2+ signal measurements with TIRF imaging system
hiPSC-CMs were incubated routinely with 1 μM of Fluo-4AM for 30 min and then imaged using a total internal reflection fluorescence imaging microscope (Leica Microsystems, Buffalo Grove, IL). The system was fitted with a 63 × oil-immersion objective lens and an Andor iXon3 camera with 512 × 512 pixels [25]. Ca2+ imaging was measured using an argon ion laser with excitation at 488 nm and fluorescence emission at wavelengths >515 nm. Ca2+ transients were recorded at 60 Hz with a depth of penetration of 110–150 nm.
Reverse transcription-polymerase chain reaction
The reverse transcription-polymerase chain reaction (RT-PCR) was used to detect mRNAs of ASIC subfamily. In brief, Total RNA was isolated from undifferentiated hiPSCs, hiPSC-CM, hiPSC-derived neural progenitor cell (NPC), hiPSC-NPC-derived neurons, and human dermal fibroblasts using TRIzol (Invitrogen). DNase I-pretreated RNA was reverse-transcribed by Superscript II RTase (Invitrogen) using random hexamers for priming. The cDNA was amplified using DreamTaq Green PCR Master Mix (Thermo Fisher Scientific) and the resulting products analyzed by agarose gel electrophoresis. PCR conditions: samples were denatured at 94°C for 2 min, then cycled at 94°C for 35 s, 64°C for 45 s, and 72°C for 45 s for 30 cycles, which was followed by final extension at 68°C for 5 min.
Quantitative RT-PCR
Total RNAs of different stages K3 and IMR-90 hiPSC-derived cardiomyocytes were extracted with TRIzol reagent (Life technologies) and reverse transcript to cDNA immediately. The cDNAs were synthesized by Verso cDNA Synthesis Kit (Thermo Scientific). Quantitative PCR was performed using SYBR™ Green PCR Master Mix (Thermo Scientific) in the CFX96 Connect Real-Time PCR Detection System (Bio-Rad). The thermal cycling parameters for PCR were 95°C for 10 min, followed by 40 cycles of 95°C for 15 s, and 60°C for 1 min.
Primers for Fig. 3A:
ASIC1-F: ACTTCCGCAGCTTCAAACCC
ASIC1-R: GCCCGAGTTGAACGTGTAGC
ASIC2-F: CGCCACTTCGAGGGAATCAGC
ASIC2-R: TTTGCCATCCTCGCCTGAGTT
Primers for Fig. 3B-D:
ASIC1a-F: ACATAGGGGAGAACATCCTGGT
ASIC1a-R: ACAGGTGTCATCAGCAGCTC
ASIC2a-F: GCACTACAAACCCAAGCAGTT
ASIC2a-R: TTTTGTAAACACTGTGGTGAAGTCT
ASIC2b-F: TTCGAGGGAATCAGCGCC
ASIC2b-R: CCCATATTTTGTAAACACGGAGGA
ASIC3-F: 5′-GAGCTGCTTGGTGACATTGG-3′
ASIC3-R: 5′-GAGTGCTGTCGGTTCCAGAA-3′
Immunohistochemistry
Brain and heart tissues for immunohistochemical analyses were obtained from adult male Sprague–Dawley rats weighing 250–300 g at age of 12–14 weeks (Charles River). Rats were anesthetized with the intraperitoneal injection of pentobarbital (60 mg/kg body weight) and perfused with phosphate-buffered saline (PBS), pH 7.4. Organs were explanted, fixed in 4% buffered paraformaldehyde (PFA) overnight at 4°C and rapidly frozen in Tissue-Tek O.C.T. mounting medium (Sakura). Before staining with primary antibodies, 10 μm tissue sections were permeabilized in PBS containing 0.5 M ammonium chloride and 0.2% Triton X-100 for 25 min at room temperature (RT) followed by antigen retrieval in PBS containing 0.05% Trypsin and 0.01% CaCl2 for 7 min at 37°C. After washing with PBS, sections were blocked with Background Buster (Innovex Biosciences) for 30 min at RT and incubated with primary antibodies overnight at 4°C in PBS containing 0.07% Triton X-100, 0.2% Tween 20, 2% bovine serum albumin (BSA), and 0.001% trypsin inhibitor.
The following primary antibodies were used: guinea pig polyclonal anti-ASIC1 (1:200, Cat. No. AGP-053; Alomone Labs), mouse monoclonal anti-α-actinin (1:800, Cat. No. A-7811, Clone: EA-53; Sigma), and rabbit polyclonal anti-MAP2 (1:200, Cat. No. ab32454; Abcam). Next day, tissue sections were washed with PBS and incubated for 2 h at RT in PBS containing 0.07% Triton X-100, 0.2% Tween 20, 2% BSA, and the following corresponding secondary antibodies: Alexa Fluor 555-conjugated anti-guinea pig IgG (1:200) in combination with Alexa Fluor 488-conjugated anti-mouse IgG (1:400) or Alexa Fluor 488-conjugated anti-rabbit IgG (1:200) (all from Thermo Fisher Scientific). Control sections were processed in the same way as described above with the exception that primary antibodies were omitted. Tissue sections were washed in PBS and counterstained with Hoechst 33342 (1 μg/mL, Molecular Probes) for 10 min at RT before mounting in Fluorescence Mounting Medium (Dako). Images were captured by AxioVision 4.5 software on an Axiovert 200M fluorescence microscope (Zeiss) coupled with a high-resolution digital camera.
Immunocytochemistry
CMs used for this analysis were generated from hiPSC line UKKi011-A (
Statistical analysis
Patch-clamp data were analyzed using Clampfit 10.6 (Axon Instruments) and Origin 8.0 (OriginLab). Average values are presented in histograms and in the text as the mean ± the standard error of the mean for “n” cells. The distribution of data in individual cells is shown in separate panels. One-way analysis of variance followed by Tukey's test or t-test was used to determine statistical significance. Significant findings are labeled with one (*p < 0.05) or two stars (**p < 0.01).
Results
Electrophysiological evidence for proton-gated currents
In voltage clamped 15–60 days old hiPSC-CMs generated by the END2-protocol, rapid application of acidic solutions pH 6.7, 6.0, or 5.0 for 2–4 s evoked a large rapidly activating and slowly inactivating inward current (Fig. 1A, inset traces), with kinetics similar to that described for ASICs, reported in neurons [10,22,23]. Note that the amplitude of the proton-gated current, measured at −50 mVs, was dependent on extracellular H+ concentration, where the average activated current was 19.85 ± 7.0 pA/pF (n = 9) at pH 5.0 and 12.02 ± 4.8 pA/pF (n = 9) at pH 6.0. Rapid changes of pH from 7.4 to 6.7 often activated little or no significant inward current in most cells examined, suggesting pH 6.7 to be the approximate threshold for activation of ASIC current in hiPSC-CMs. Quantification of peak ASIC-type current at holding potentials between −80 to +70 mVs (Fig. 1B) showed a linear dependence on holding potential, suggesting voltage-independence of the current, consistent with the idea that the magnitude of activated current depended on the electrochemical driving force for Na+, ENa, as also reported for DRG neurons [10].

pH dependence, voltage independence, and external Na+ and Ca2+ dependence and pharmacological suppression of proton-gated inward currents in hiPSC-CMs.
To characterize the ionic and pharmacological sensitivities of this current, we replaced the extracellular Na+ with Tetra-ethyl-ammonium (TEA+) and found that replacement of Na+ by TEA+ almost fully suppressed the current, revealing often a slowly activating component of the current (Fig. 1C). The strong Na+ dependence of the current and the marked decrease of ASIC current at potentials positive to +70 mV, near ENa, suggest that the proton-gated inward current is carried primarily by Na+ (Fig. 1C).
Similar to the findings of Konnerth et al. [10], 1987 in DRG neurons, divalent cations (Cd2+ and Ca2+) suppressed the ASIC current, such that removal of Ca2+ enhanced the current (Fig. 1C), and addition of 1 mM Cd2+ to the activating acidic solutions suppressed the current (Fig. 1F). These findings are also consistent with previous reports that Ca2+ suppresses proton-activated current [30], and 1 mM Cd2+ inhibits ASIC2a and ASIC3 homomeric channels and ASIC1a/2a, 1a/3, and 2a/3 heteromeric channels, but not ASIC1a homomeric channel [31].
Figure 1D and E show that 5–10 μM amiloride, a broad-spectrum inhibitor of ASIC channels, also significantly suppresses the proton-gated current (∼50% suppression at 5 μM amiloride) in hiPSC-CM, consistent with previous findings in neurons [22,32] and suggest that the proton-gated channel in hiPSC-CMs is an amiloride-sensitive Na+ channel. The inhibitory effect of amiloride was only partially reversed after a 3-min washout of the drug.
Functional expression of ASIC
Following the discovery of proton-gated currents in dorsal root ganglion neurons in 1987 [10], we attempted to measure the expression of proton-gated currents in CMs, but failed to detect ASIC-like currents in neonatal or adult rat, adult guinea pig, and adult cat CMs (Morad, unpublished observations, 1989). The expression of ASIC currents in hiPSC-CMs was therefore unexpected. To trace the possible ancestral origins of ASIC currents, we tested for ASIC expression in undifferentiated hiPSC and in dermal fibroblasts (data not shown). Figure 2A shows that ∼17% of undifferentiated hiPSC expressed the proton-gated currents on exposure of the cells to pH 6.0. Generally, two kinetically different types of currents were found in undifferentiated hiPSCs. The variability in ASIC currents kinetics most likely reflects the level of expression of different subtypes of this gene family of channels [20]. In another set of experiments, we tested for expression of ASIC currents in dermal fibroblast and found variable expression of the current (0.3–5 pA/pF) and its kinetics (data not shown).

Time course of ASIC current development in CMs differentiated from hiPSC lines-K3, IMR-90, and NP0040.
To determine whether procedures used to differentiate hiPSC lines into cardiomyocytes (coculture with END2 vs. Wnt-signaling differentiation) contribute to the level of expression of ASIC, we tested for expression of ASIC in K3 lines where Wnt signaling was used to differentiate hiPSCs into cardiomyocytes. Consistent with our mRNA studies (Fig. 3C), we found a strong expression of ASIC currents in K3-line-derived cardiomyocytes, Fig. 2C, the magnitude of which decreased in older 100 day hiPSC-CM cultures.
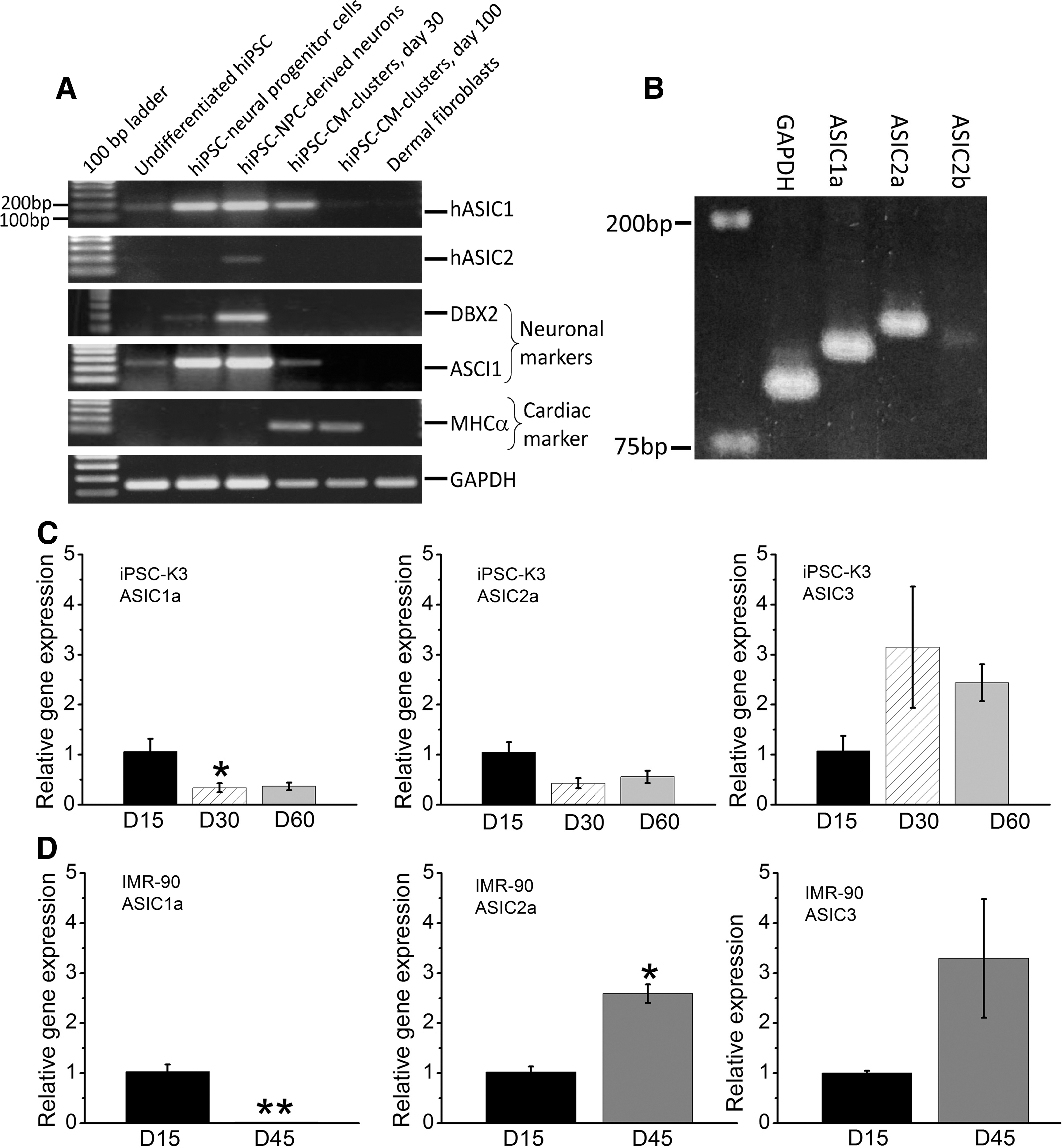
mRNA expression of ASIC family in hiPSC differentiated cardiomyocytes.
We also tested for the expression of proton-gated currents in hiPSC-CMs differentiated from two other lines (K3 and NP0040). Figure 2A compares the fraction of cells (in %) expressing ASIC currents activated by pH 6.0 at −50 mVs, in undifferentiated hiPSCs and hiPSC-CMs differentiated from different hiPSC lines. In ∼80%–90% of hiPSC-CM, rapid applications of pH 6.0 solutions activated robust rapidly developing inward currents at −50 mVs holding potential. The density of ASIC currents in undifferentiated hiPSC was significantly lower, ranging about 1 pA/pF, compared to 5.15 ± 0.66 pA/pF in hiPSC-CMs (Fig. 2B–D). To determine the time course of development of ASIC expression in hiPSC-CMs cultures, we compared the current densities of ASIC in 15, 30, 60, and 100 days in K3 line, at both pH 5 and 6, and found gradual decreasing levels of expression in older cultures (Fig. 2C). Similarly, hiPSC-CMs derived from the NP0040 (Fig. 2D) and IMR-90 iPSC line (Fig. 2B) showed a similar pattern of decreasing expression of ASIC currents in older cultures, suggesting possible developmental aspects to the expression of proton-gated currents.
Expression of ASIC mRNA and protein in hiPSC-CMs
Our electrophysiological data provides evidence for expression of proton-gated channels with properties similar to ASIC family of channels in hiPSC-CM, with kinetics of activation and inactivation suggestive of multiple subtypes of ASIC channels. To determine the molecular identities of the ASIC currents, the mRNA expression levels of ASIC1 and ASIC2 were measured with RT-PCR using primers specific for these transcripts. Human iPSC-derived NPCs and neurons differentiated from them were used as positive controls for ASIC expression. Figure 3A shows that ASIC1 transcripts were highly expressed both in hiPSC-NPCs and hiPSC-NPC-derived neurons. ASIC1 mRNA was also expressed in early cultures of hiPSC-CM (differentiation day 30, IMR-90 line), but at much lower levels in 100 day cultures (Fig. 3A, C, D), consistent with the electrophysiological measurements showing lower percentage of older hiPSC-CMs express the proton-gated currents (Fig. 2). ASIC1 transcripts were also expressed at low levels in undifferentiated hiPSC but not in human dermal fibroblasts (Fig. 3A), where ASIC currents were occasionally detected. ASIC2 mRNA was detected only in hiPSC-NPC-derived neurons, but not in any other cell types (Fig. 3A), suggesting that ASIC1 mRNA most likely encodes the proton-gated currents in early cultures of hiPSC-CMs from IMR-90 line. In another series of RT-PCR experiments, we used different sets of primers for ASIC1a, ASIC2a, and ASIC2b. They revealed the presence of also ASIC1a, ASIC2a, and ASIC2b transcript in 55 days old hiPSC-CMs derived from iPSC-K3 line (Fig. 3B). The electrophysiological and molecular studies thus confirm the presence of ASIC gene family in both hiPSCs and CMs differentiated from them.
Quantitative PCR data confirmed that ASIC1a, ASIC2a, and ASIC3 members of ASIC family are expressed in hiPSC-CMs, Fig. 3C and D. It appears that ASIC1a mRNA levels measured at days 30, 45, or 60 were significantly lower compared to day 15 in K3 and IMR-90 hiPSC-CMs. ASIC2a mRNA levels at days 30 and 60 show a decreasing trend compared with day 15 in K3 hiPSC-CMs. In IMR-90 hiPSC-CM line, however, ASIC2a mRNA expression at day 45 were significantly higher, suggesting different patterns of ASIC family member expression in different hiPSC lines. ASIC3 expression tends to increase in both the long-time cultured K3 and IMR-90 hiPSC-derived CMs. However, it should be noted that Cycle threshold (Ct) values of quantitative PCR for ASIC3 is considerably high (30–33) in IMR hiPSC-CMs, suggesting that ASIC3 expression level in IMR hiPSC-CMs is low. We also tested for ASIC protein expression in hiPSC-CMs. Figure 4 shows immunocytochemical staining evidence for presence of ASIC1 proteins in days 20 and 62 cultures of hiPSC-CMs from the NP0040 hiPSC line, using SH-SY5Y neuroblastoma cells as a positive control. The quantitative PCR and immunostaining measurements are consistent with our electrophysiological findings suggesting the presence of ASICs in hiPSC-CMs.
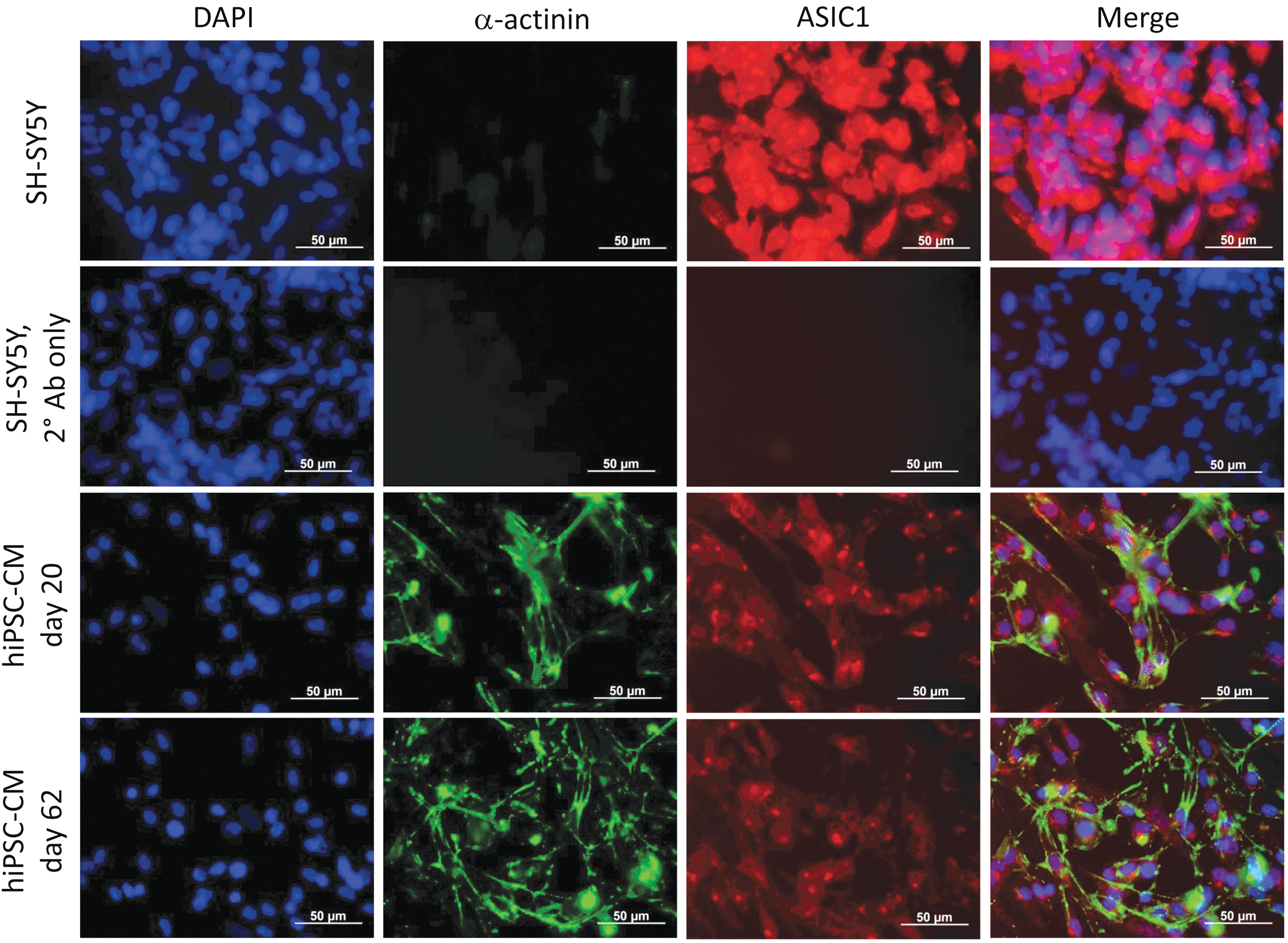
Assessment of ASIC1 expression in human iPSC-CMs using immunocytochemistry. Representative immunofluorescence panels show colocalization of ASIC1 (red) with the cardiomyocyte marker a-actinin (green) in human iPSC-derived cardiomyocytes both at day 20 and at day 62 of directed differentiation. The neuroblastoma SH-SY5Y cells were used as positive control and exhibit more intense ASIC1 signal than CMs. Cells incubated only with fluorescently labeled secondary antibodies did not produce any specific signals. Nuclei were counterstained with DAPI (blue). CMs were differentiated from NP0040 iPSC line. Scale bars: 50 μm. Color images are available online.
Cytosolic Ca2+ transients activated by hyperacidification in hiPSC-CMs
Since rapid acidifications and activation of large inward currents are likely to depolarize the cells, we examined whether such depolarizations would activate I Ca and thereby trigger intracellular calcium stores through a calcium-induced calcium release (CICR) mechanism. Figure 5 shows that in intact cells, incubated with Ca2+ indicator Fluo-4AM, rapid application of pH 5.0 solutions for 2 s triggered Ca2+ transients with two components of Ca2+ release. The first component was larger and occurred with application of acid pulse, while the second component was smaller and developed at the end of 2 s long pH 5.0 pulse. Interestingly, 1 mM Cd2+ suppressed the first component by only ∼38%, but completely abolished the second pH 5 triggered Ca2+ transient component, suggesting that activation of ICa and CICR significantly modulate the pH-triggered Ca2+ transients. Zero Ca2+ plus EGTA suppressive effects, on the contrary, were smaller at ∼17%, suggesting a minor effect of ICa and CICR in modulating the first component of pH-triggered Ca2+ transients (Fig. 5D, F). There was nevertheless a trend toward abbreviation of pH 5.0-triggered Ca2+ transients by Cd2+ and zero Ca2+ solutions, related possibly to ASIC current-mediated depolarization of the membrane and activation of CICR.
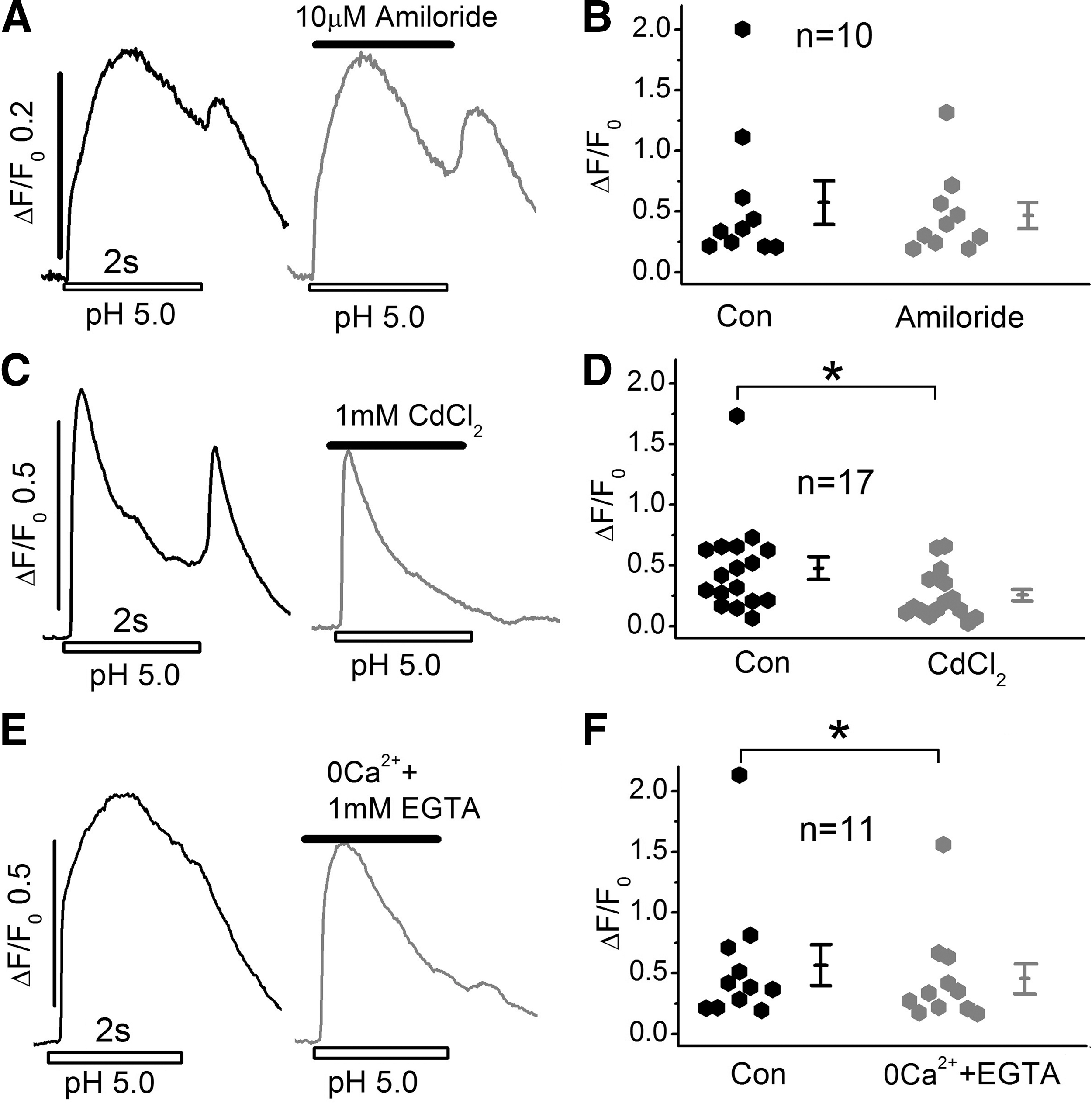
Ca2+ transients triggered by rapidly exogenous application of acidic Tyrode's solution in Fluo4-AM incubated hiPSC-CMs.
In another set of experiments, we tested for the possible role of IP3 signaling and TRPV channels in triggering or modulating the pH-triggered Ca2+ transients, but found no significant effects of IP3-receptor blocker, 2-APB (20 μM), or TRPV channel blocker BCTC (10 μM).
The finding that 10 μM amiloride failed to alter the time course or amplitude of these Ca2+ transients (Fig. 5A, B), also suggests that these transients are not triggered directly by influx of Na+ on activation of ASIC currents (compare Fig. 5A, B with Fig. 1D, E). The almost complete suppression of ASIC current by amiloride, but only minor suppression of Ca2+ transients and our consistent finding of absence of pH-triggered Ca2+ transients in whole-cell patch-clamped hiPSC-CMs, where the cytosol is dialyzed with solutions buffered with 10 mM HEPES, 0.1 mM EGTA, and 0.1 mM Fluo4, may suggest that extracellular acidification and influx of protons may directly signal the release of Ca2+ from intracellular stores by a novel mechanism.
Expression of ASIC in neonatal and adult mammalian cardiomyocytes
Following our initial reports of expression of acid-activated currents in chick DRG neurons [10,11], we attempted to measure proton-gated currents in adult Guinea pig and rat cardiomyocytes, but failed to measure detectable levels of proton-gated currents in either heart, but found significant acid-activated currents in bovine adrenal gland chromaffin cells [33] and neuroblastoma cell lines [22]. A recent report, however, suggests possible expression of “ASIC like current” in cultured adult rat cardiomyocytes that was suppressed by very high concentrations (100 μM) of amiloride [34]. Therefore, we reinvestigated the possible expression of ASIC currents in freshly isolated or cultured adult rat cardiomyocytes, but failed to activate significant ASIC currents on rapid applications of pH 5 solutions in adult rat cardiomyocytes having the appropriate ASIC current kinetics (Fig. 6).
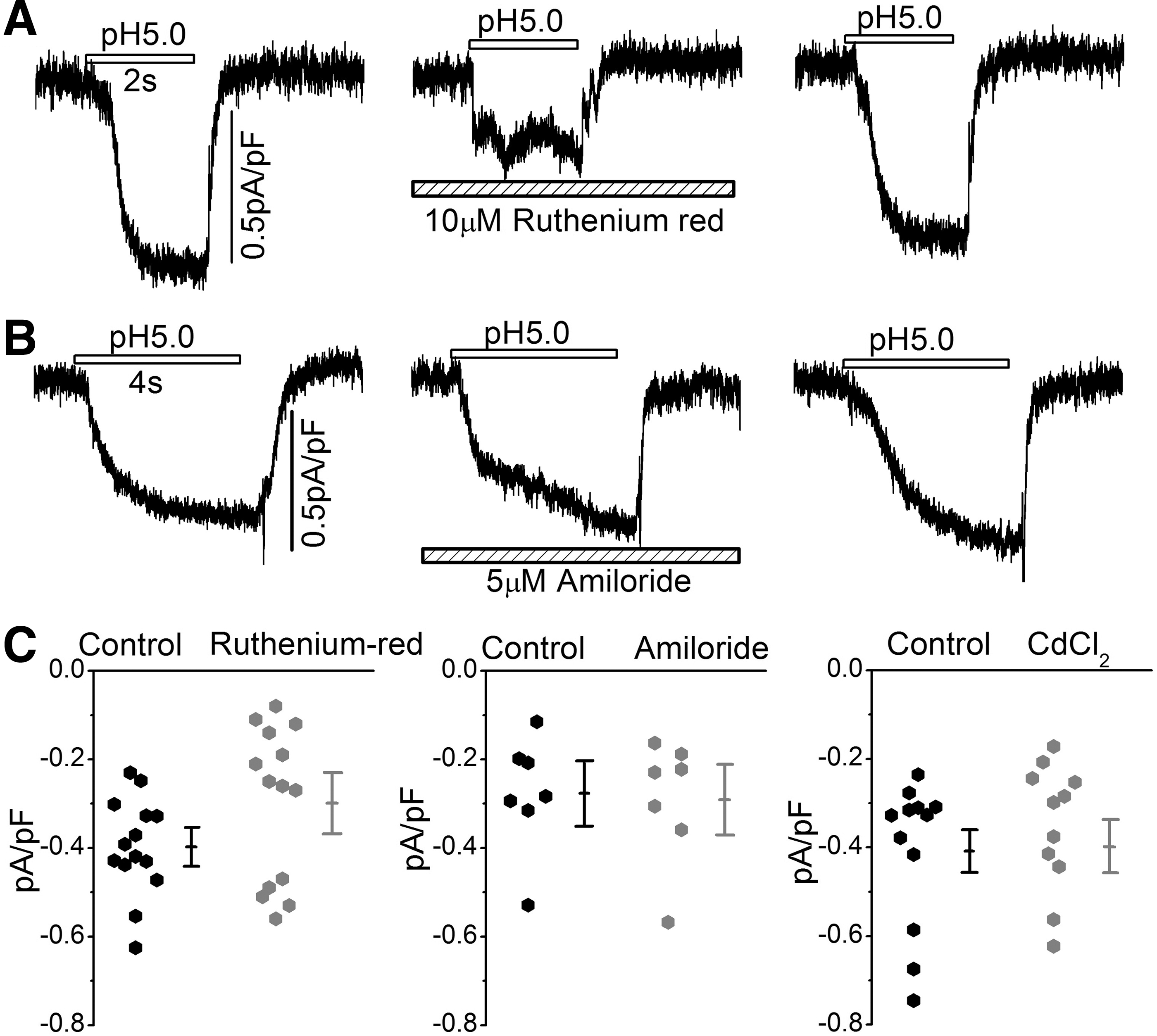
Acidosis-induced currents recorded from adult rat ventricular cardiomyocytes.
Hyperacidification did, however, activate slowly developing currents with kinetics somewhat similar to those reported for TRPV channels, and similar to the currents reported by Hu et.al, recently [34]. It should be noted that there were not only significant differences between the kinetics of activation and inactivation of acid-activated currents in freshly isolated rat heart cells and human iPSC-CMs, but also there was significant differences in the drug sensitivities of the two types of currents. Figure 6A and B show that while 10 μM ruthenium red suppressed some of the acid-activated non-ASIC-like current in adult rat cardiomyocytes, 10 μM amiloride and 1 mM CdCl2 failed to suppress the acid-activated currents in adult cardiomyocytes, as was the case for ASIC currents expressed in hiPSC-CMs, where 5 μM amiloride and 1.0 mM Cd2+ were sufficient to significantly suppress the current (Fig. 1C, D). Note also that while ASIC currents ranged about 6 pA/pF at −50 mV holding potentials in hiPSC-CMs, the non-ASIC acid-activated current ranged about 0.3–0.4 pA/pF in freshly isolated adult rat cardiomyocytes (Fig. 6C). Similarly, we also failed to activate ASIC-type currents in neonatal rat cardiomyocytes (data not shown).
ASIC1 proteins expression in the adult rat heart
To further examine whether the acid-induced currents of Fig. 6A and B reflect the possible expression of ASIC proteins in adult cardiomyocytes, we tested for the expression of ASIC1 in adult heart cryoslices and compared it to those of rat brain slices. Figure 7 shows that while there was colocalization of ASIC1 with the neuronal microtubule marker, associated protein 2 (Map2), in the adult rat brain, there was no ASIC1 expression in the adult rat heart muscle cells that stained positive for the cardiomyocyte marker sarcomeric α-actinin (Actn2). Tissue specimens incubated only with fluorescently labeled secondary antibodies was used as negative control producing no specific signals. These findings are consistent with our electrophysiological results suggesting the absence of ASIC in adult heart cells.
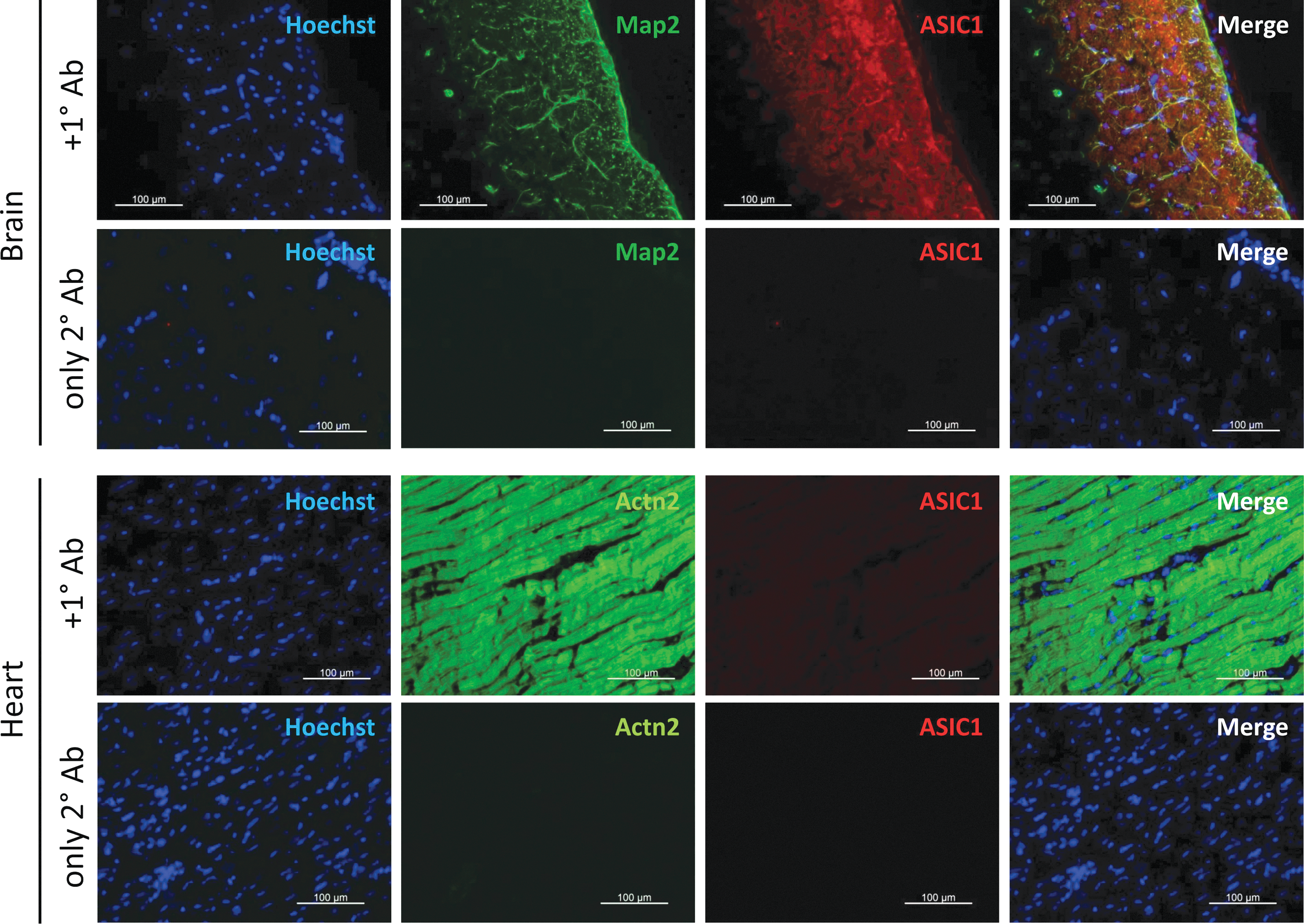
Immunohistochemical detection of ASIC1 in adult rat brain and heart tissue cryoslices. Representative immunofluorescence panels show colocalization of ASIC1 with the neuronal marker microtubule-associated protein 2 (Map2) in the adult rat brain but no ASIC1 expression in the adult rat heart muscle cells, which stained positive for the cardiomyocyte marker sarcomeric α-actinin (Actn2). Tissue specimens incubated only with fluorescently labeled secondary antibodies gave no specific signals. Nuclei were counterstained with Hoechst 33342. Scale bars: 100 μm. Color images are available online.
Discussion
The major finding of this study is that cardiomyocytes derived from human iPSCs express significant levels (∼5 pA/pF) of proton-gated ASIC current in the first 15–60 days' postcardiac differentiation in cultures. The acid-activated currents had the same biophysical and pharmacological properties as the ASIC currents reported in neuronal tissues. This is a novel finding because adult or neonatal rat cardiomyocytes do not express this gene family of channels even though there is strong expression of ASIC current in cardiac sensory neurons [35,36]. The expression of ASIC1 mRNA or its encoded proton-gated current depended on the age of the myocytes in culture, such that the density of the current or mRNA levels was high in 15–60 day cultures but decreased significantly in older cultures, becoming almost undetectable in 100 days, suggesting developmental aspects to the expression of this channel family in the heart. In this respect, we failed to detect ASIC protein or ASIC-like current in freshly isolated rat cardiomyocytes or cultured neonatal cardiomyocytes isolated from 3 to 4 day old pups, even though we consistently measured small (<1 pA/pF) acid-activated slow- or noninactivating TRPV-type currents from adult ventricular cardiomyocytes, which look like proton-gated currents.
Expression of ASIC gene family in hiPSC-CMs and its cellular ancestry
We consistently found significant expression of proton-gated channels in CMs derived from all three hiPSC lines (Fig. 3). There was significant variability in the magnitude and kinetics of proton-gated currents suggesting cellular heterogeneity possibly related to different members of this gene family, culture age postdifferentiation, or expression of other types of acid-activated ion channels, such as TRPV currents in our culture. Clearly, the functional data must be carefully scrutinized and supported by molecular/imaging data (mRNA or immunofluorescence) before determining whether the proton-gated currents are generated by ASIC or TRPV family of channels. In this respect, presence of ASIC1a, ASIC2a, and ASIC3 is suggested based on both mRNA data and the kinetics of activated currents (Figs. 1 and 3).
The absence of ASIC mRNA in dermal fibroblasts, but its expression in hiPSC suggests that the reprogramming maybe responsible for the functional expression of ASIC current. In this respect in a series of experiments on mouse embryonic stem cell-derived cardiomyocytes, we found no proton-gated ASIC-type currents in either 13 or 21 day cultures, even though there was robust expression of ICa and “funny current” (If) in some 20 cells examined, consistent with the idea that reprogramming may be, in part, a stimulant to expression of ASIC proteins/currents in hiPSC-CMs (Zhang, Foley, and Morad, 2019, unpublished data). It is not as yet clear whether the higher density of ASIC current in early developmental stages of hiPSC-CMs is related to a specific signaling pathway or other maturation factors. Although molecular evidence supports the idea that differences in the kinetics of inactivation of the current may arise from expression of different ASIC family members in hiPSC-CMs, the heterogeneity in the magnitude of the current in hiPSC-CMs and in undifferentiated hiPSC-lines makes it difficult to accurately quantify the kinetics of the proton-gated current with sufficient reliability as to associate them with specific members of this gene family.
Acid-activated calcium transients and ASIC currents
In intact hiPSC-CMs, we consistently found that rapid hyper-acidification (pH 5 and 6) triggered cytosolic rise of intracellular calcium that was not directly activated by ASIC currents, because pharmacological interventions that blocked ASIC currents did not significantly suppress them (Fig. 5A, B). Acid-activated Ca2+-transients had generally two components. The second component of these transients appeared always to develop at the end of 2 s long acidification pulse (Fig. 5C), and was completely blocked by 1 mM Cd2+ or zero Ca2+/EGTA (Fig. 5C, E). The triggering of Ca2+ release at the end of acidification pulse and its block by Cd2+ and EGTA suggests that depolarization of membrane caused by large ASIC currents activates the L-type calcium channels, which is unable to transport Ca2+ through the channel or release calcium from the ryanodine receptors of SR, because they are suppressed by large concentrations of protons, which when washed away allow sufficient influx of Ca2+ activating CICR and the second component of Ca2+ transients. This possibility is supported by absence of hyperacidification-induced Ca2+ transients in whole-cell clamped myocytes where cells are dialyzed with Fluo4, EGTA, and HEPES, buffering significant changes of both calcium and protons.
In this scheme, the first component of hyperacidification is caused by a novel signaling pathway that releases calcium from other intracellular compartments that either binds or store calcium (endoplasmic reticulum, nuclear envelop, mitochondria, and sarcolemma). The role of IP3 receptor or TRPV1 channels in this signaling pathway could not be confirmed as IP3 and TRPV1 channel blockers failed to suppress consistently these acid-activated Ca2+ transients.
Physiological implications of ASIC expression in the heart
ASIC family of channels are uniquely expressed in almost every type of neurons and have been implicated in pain perception and signaling as well as modulating synaptic transmission. Cardiac sensory neurons also strongly express ASIC, which may be responsible for the strong pain felt on coronary occlusions caused by local or global ischemia. Our data clearly suggest that adult mammalian myocardium does not express functional ASIC proteins. Molecular and functional studies, however, suggest that mammalian embryonic myocardium and hiPSC-derived cardiomyocytes express ASIC mRNA and functional channels encoded by them. A search of the GEO profiles dataset [37,38] found data demonstrating ASIC1 mRNA expression in mouse embryonic stem cells and at lower levels in the mouse heart tube at dpc 8.5. The levels of mRNA continued to be expressed at E9.5 and E12.5 with decreasing mRNA expression throughout later stages of embryonic development (GDS5003) [39]. ASIC1 transcripts were also shown to be expressed at high levels in human embryonic stem cells, CMs derived from them and in fetal ventricular cardiomyocytes. By contrast adult ventricular cardiomyocytes expressed only low levels of ASIC mRNA (GDS5603) [40]. Our preliminary immunohistochemistry studies in human neonatal heart also suggest significant expression of ASIC1 protein. Thus, the significant expression of ASIC-associated mRNA and functional protein suggest a developmental role for ASIC family in the heart.
The finding that acidification in addition to activation of large Na+ current triggers also the release of cytosolic calcium may provide a clue as to the developmental role of ASICs in initiation of spontaneous beating of the heart, before development of ionic channels in mammalian myocardium. Mouse embryonic cardiomyocytes have been shown to beat spontaneously at E-6.5 even when placed in isotonic KCl solutions [41], suggesting that their beating precedes the expression of ICa and CICR. Acidification and activation of Na+ transporting channels may in conjunction serve to activate mitochondria to release the calcium necessary for spontaneous beating in the early stages of cardiac development. This would be consistent with significant expression of ASIC in early stages of cardiac development but not in the adult heart. Although direct experimental data are at present unavailable to test this hypothesis, the well-known effects of hypoxia (likely producing acidosis) on development have been long recognized.
Footnotes
Acknowledgments
We thank Drs. Hua Wei and Yanli Xia, Ms. Taylor Rienzo and Christy Couch, and Dr. Kurt Pfannkuche for generation and dissociation of hiPSC-CMs and Drs. Foley and Yunkai Dai for providing mouse embryonic stem cell-derived cardiomyocytes. We thank Ms. Andrea Hoffman for technical support in cultivation of hiPSC-NPCs and generation of neurons from them, Dr Naohiro Yamaguchi for helping designing PCR primers, and Ms. Alessa Großert and Prof. Nicole Teusch for providing SH-SY5Y cells. We thank the support of NIH RO1 HL016152 to M.M. and grants awarded by Köln Fortune Program and Deutsche Forschungsgemeinschaft (SA 1382/7–1) to T.Š.
Author Disclosure Statement
No competing financial interests exist.