
Abstract
Hypopharyngeal carcinoma is a common malignant tumor of the head and neck with a very poor prognosis; the median survival time for curatively treated patients was 17.2 months in India. However, cell-based gene therapy holds promise to improve patient outcomes. In this study, we investigated whether human bone marrow mesenchymal stem cells (BMSCs) possess potential homing capacity for hypopharyngeal carcinoma. To monitor the efficiency of BMSC transplantation therapy through reporter gene imaging, we employed a hybrid baculovirus vector containing the Luc-P2A-eGFP fusion or sodium iodide symporter (NIS) sequence under the control of the cytomegalovirus promoter. To enhance the transfection efficiency, baculovirus vectors (Bac-CMV-Luc-P2A-eGFP-ITR and Bac-CMV-NIS-ITR) were flanked by inverted terminal repeats (ITRs), which are key elements of adeno-associated viruses. The infection efficiency of Bac-CMV-Luc-P2A-eGFP-ITR in BMSCs was as high as 92.84 ± 1.14% with no obvious toxic effects at a multiplicity of infection of 400. Moreover, Bac-CMV-NIS-ITR-infected BMSCs showed highly efficient radioactive iodide (125I) uptake; these high uptake levels were maintained for at least 2 h. Transwell migration assays further demonstrated the chemotaxis of BMSCs to hypopharyngeal carcinoma cells (FaDu cells) in vitro. BMSCs modified by firefly luciferase report gene or NIS were injected into nude mice with hypopharyngeal carcinoma, and changes in the localization of the BMSCs were successfully tracked with bioluminescent imaging and micro–single-photon emission computed tomography imaging. These data indicate the potential utility of BMSCs as a promising targeted-delivery vehicle for hypopharyngeal carcinoma gene therapy. Importantly, BMSCs may represent a promising targeting vector for general tumor radionuclide therapy.
Introduction
Head and neck cancers are one of the most common malignancies in developing countries, where they impose an increasing financial burden on cancer patients and their families [1]. Hypopharyngeal carcinoma usually occurs at the pyriform sinus, the posterior wall of the laryngeal pharynx, and the postcricoid area [2]. Among the head and neck cancers, hypopharyngeal carcinoma is associated with the worst prognosis as early detection is difficult to achieve. Because of its biological characteristics, lymph node metastasis of hypopharyngeal carcinoma occurs early. There has been a notable progress in surgical techniques and chemoradiotherapies for hypopharyngeal carcinoma; however, the 5-year survival rate is only ∼40% [3]. Therefore, this cancer requires a multimodal therapeutic strategy [4,5]; however, even with multiple treatments, the outcome of patients with hypopharyngeal carcinoma remains poor.
Recently, gene therapy has emerged as a revolutionary therapeutic approach with a mechanism distinct from that of conventional drugs. Owing to the intrinsic biological capabilities of stem cells, such as homing, trafficking potential, and regenerative capacity, they can serve as vehicles for targeted gene delivery; this potential provides opportunities for gene therapy to overcome a large number of unconquered medical challenges [6]. The main technical challenge associated with the efficient use of stem cells in gene therapy is the safe delivery of therapeutic genes in a targeted manner to specific cells. Viral vectors are widely used for gene delivery; however, their immunogenicity, potential for insertional mutagenesis, and carcinogenicity limit their clinical application [7].
Baculovirus is a promising viral vector for gene delivery owing to its advantages of low cytotoxicity, large capacity (>100 kb), and easy mode of preparation [8,9]. However, the efficiency of transduction of baculovirus-mediated gene delivery needs further improvement. To date, no study has examined the application of bone marrow-derived mesenchymal stem cells (BMSCs) as a targeted carrier for gene therapy of hypopharyngeal carcinoma.
Two noninvasive and serial methods have been recently developed to visualize and track transplanted cells in vivo: bioluminescent imaging (BLI) and radionuclide imaging. BLI enables the real-time monitoring of target cells through detection of light generated by the reaction of target cells transfected with luciferin genes with their bioluminescent substrates [10,11]. In addition, sodium iodide symporter (NIS) mediates the uptake of radionuclides, such as 131I, 123I, 94mTcO4, and 125I, making it suitable for radionuclide imaging and radionuclide therapy. Therefore, the NIS gene has been recognized as a versatile new reporter gene and potential therapeutic gene [12 –16].
Therefore, in this study, we used both methods, BLI and radionuclide imaging with NIS, to reveal the dynamics of the distribution and tropism of BMSCs transplanted in a mouse model of hypopharyngeal carcinoma. Furthermore, we investigated the feasibility of a novel hybrid baculovirus-adenovirus-associated viral vector-mediated gene as a new strategy for modifying BMSCs.
Materials and Methods
Isolation, culture, and differentiation of BMSCs
Human BMSCs were purchased from the Chinese Academy of Sciences Cell Bank (Shanghai, China) and cultured in human BMSC growth medium (Cat. HUXMA-90011; Cyagen Biosciences, Inc., China) at 37°C in 5% CO2. Cells of passage 4 through 6 were used for experiments. To confirm the differentiation ability of the BMSCs, adipogenic (Supplementary Fig. S1.) and osteogenic (Supplementary Fig. S2.) differentiation was induced as previously described [17,18].
Construction and preparation of recombinant baculovirus-adeno-associated vectors
The baculoviral plasmid pFBGFPR was kindly provided by the School of Biomedical Sciences (Faculty of Medicine, The Chinese University of Hong Kong, Hong Kong, China), and pFBNIS was obtained from the laboratory of Rui Jin Hospital. Two reporter genes encoding firefly luciferase (ffLuc) and enhanced green fluorescent protein (EGFP), respectively, were fused as an open reading frame (ORF) through the P2A sequence. The fusion ORF was then subcloned into the baculoviral plasmid under the control of a cytomegalovirus (CMV) promoter flanked by inverted terminal repeat (ITR) sequences from adeno-associated virus (AAV). The resulting product was dubbed pFB-CMV-Luc-P2A-eGFP-ITR. According to the protocol of the Bac-to-Bac baculovirus expression system (Invitrogen), we prepared recombinant baculovirus-adeno-associated vectors (BA-AAV) (Bac-CMV-Luc-P2A-eGFP-ITR and Bac-CMV-NIS-ITR). Recombinant baculovirus-adeno-associated viral vectors were amplified to high titers by infecting Sf9 cells, and the virus titer was determined by the endpoint dilution [19] method and quantitative real-time PCR [20].
Baculoviral infection of BMSCs
BMSCs were incubated with Bac-CMV-Luc-P2A-eGFP-ITR (BMSCs-Bac-Luc-P2A-eGFP) at various multiplicities of infection (MOI), ranging from 0 to 1,600, at 27°C for 4 h with mild shaking, using phosphate-buffered saline (PBS) as the surrounding solution. Transduction efficiency was determined using flow cytometry (Beckman Coulter, Miami, FL) by detecting EGFP.
In vitro cytotoxicity assay
Following virus infection, the viability of BMSCs was assessed using the Cell Counting Kit-8 (CCK-8) assay (Beyotime Inst. Biotech, China). After incubating overnight in 96-well plates, BMSCs (5,000 cells/well) were infected with Bac-CMV-Luc-P2A-eGFP-ITR at different MOIs (0–1,600) using PBS as the surrounding solution for 4 h. In the control group, uninfected BMSCs were cultured with PBS for 4 h. After 4 h, cells were incubated with complete medium for 24 h. CCK-8 solution (10 μL) was added, and the absorbance was measured 1 h later at 450 nm using a microplate reader (Thermo Fisher Scientific).
125I uptake and inhibition
BMSCs (3 × 104 cells/well) were incubated overnight in 24-well plates and infected with Bac-CMV-NIS-ITR at MOI = 400 (BMSCs-Bac-NIS); uninfected BMSCs were used as a control group. Following infection, BMSCs-Bac-NIS were incubated in a radioiodine working solution containing 500 μL of Dulbecco's modified Eagle's medium, 3.7 kBq Na125I (Shanghai GMS Pharmaceutical Co., Ltd., China), and 10 μmol/L sodium iodide (NaI) per well. The radioiodine working solution was aspirated at different time points (15, 30, 60, and 120 min) to terminate radioactive iodide uptake. In the uptake inhibition assay, NaClO4 (30 μM) was added to the radioiodine working solution. After washing three times with ice-cold PBS, the BMSCs were incubated with NaOH (1 M) for 15 min and the radioactivity was measured in counts per minute using an automatic gamma counter (Shanghai Hesuo Rihuan Photoelectric Instrument Co., Ltd., China).
Migration of BMSCs in vitro
Cell suspensions were prepared in culture medium containing 1 × 105 cells/mL for 24-well chambers. BMSCs or BMSCs-Bac-Luc-eGFP (200 μL) were added to the upper chamber of transwell plates with 8-μm pore membranes (Corning Costar). Hypopharyngeal carcinoma FaDu cells (1 × 105 cells/mL) or cell-free medium (500 μL) as the control was placed in the lower chamber with 8-μm pore membranes. After 24 h of incubation, cells on the upper side of the membrane were removed with a cotton stick. The migrated cells attached to the lower side of the membrane were stained with 0.5% crystal violet and dissolved in 33% acetic acid. Equal volumes (100°μL) of the mixture were placed in a 96-well plate and the absorbance was measured at 570 nm to quantify the amount of migration.
Establishment of the hypopharyngeal carcinoma mouse model
All animal studies were carried out according to the ethical principles of animal experimentation under the approval of the Institutional Animal Care and Use Committee of Ruijin Hospital. FaDu cells (5 × 106 cells in 100 μL of PBS) were injected into the left axilla or right hindlimb of female nude mice (BALB/c nu/nu, 5–6 weeks old; Jie Si Jie Laboratory Animal Corp., China). After ∼1 week, the tumors reached 0.5 cm in diameter, and then the mice were used for the subsequent experiments.
BLI in vitro
To verify the ability of BLI to track BMSCs to the target cells, we first conducted BLI in vitro with a luciferin reporter assay. BMSCs-Bac-Luc-eGFP were seeded into a black 96-well plate at various densities [5 × 103, 1/2 × (5 × 103), 1/4 × (5 × 103) … 1/32 × (5 × 103) cells/well] with four wells per group. Then, 100 μL of a 2 × solution (0.3 mg/mL) of
BMSC distribution in tumor-free nude mice and hypopharyngeal carcinoma model mice
We next used BLI to evaluate the distribution of the BMSCs in vivo. BMSCs-Bac-Luc-eGFP (1 × 105) were respectively injected into tumor-free nude mice and the hypopharyngeal carcinoma model mice through the tail vein (n = 3 per group). As a control, the nude mice were injected with 1 × 105 uninfected BMSCs through the tail vein (n = 3).
According to previous studies [21,22], BLI of tumor-free nude mice was performed at 4, 8, and 24 h after injection. To trace the BMSCs in the hypopharyngeal tumor model, BLI was performed at 1, 2, 5, and 7 days after BMSCs-Bac-Luc-eGFP injection.
Micro–single-photon emission computed tomography/computed tomography imaging
Single-photon emission computed tomography/computed tomography (SPECT/CT) was also used for tracking the BMSCs based on radioactive iodine uptake. BMSCs-Bac-NIS (1 × 105) were injected into the tail veins of the hypopharyngeal tumor-bearing mice (n = 3) twice on days 0 and 2; these mice were designated tumor-BMSCs-Bac-NIS mice. Uninfected BMSCs were also injected into the model mice as a control. On day 5 after injection of 125I (74 MBq) through the tail vein, the mice were anesthetized with 4% isoflurane to facilitate scanning with Mediso nanoScan SPECT/CT (Bioscan). Axial, coronal, and sagittal raw images were obtained, and then reconstructed using InVivoScopeH 1.43 software (Bioscan).
Immunofluorescence analysis
The mice were sacrificed after BLI and micro-SPECT/CT imaging, and the tumors were removed and snap frozen for immunofluorescence staining. The slides from frozen tumor tissues were incubated with polyclonal rabbit anti-NIS antibody (Proteintech) at 4°C overnight, and then with tetramethylrhodamine (TRITC) anti-rabbit secondary antibody for 1 h (diluted 1:100; YEASEN Biology, China). Nuclear staining of the samples was then performed with 4′,6-diamidino-2-phenylindole (DAPI). The slides were rinsed several times to remove all free DAPI, and then viewed under a fluorescent confocal microscope (Carl Zeiss, Inc., NY).
Statistical analyses
All data are expressed as mean ± standard deviation from at least three independent experiments. The unpaired t-test was used for determining the significance of differences between groups; P < 0.05 was considered to indicate statistically significant differences. GraphPad Prism 7.00 (GraphPad Software, Inc.) was used for all statistical analyses.
Results
Construction and amplification of recombinant Bac-CMV-Luc-P2A-eGFP-ITR and Bac-CMV-NIS-ITR
The plasmid pFB-CMV-Luc-P2A-eGFP-ITR was constructed (Fig. 1). BV-AAV vectors were amplified, and the titer was determined to be 109 plaque forming unit (PFU)/mL.

Schematic representation of the plasmid pFB-CMV-Luc-P2A-eGFP-ITR. The fusion open reading frame (Luc-P2A-eGFP) was subcloned into the baculoviral plasmid under a CMV promoter flanked by a pair of ITRs. GFP, green fluorescent protein; CMV, cytomegalovirus; ITR, inverted terminal repeat.
Infection efficiency of BMSCs with Bac-CMV-Luc-P2A-eGFP-ITR
As illustrated in Fig. 2A and B, BMSCs were effectively infected with Bac-CMV-Luc-P2A-eGFP-ITR at an MOI of 400. Flow cytometry demonstrated a rapid increase in the transfection rate with an increase in the MOI value from 0 to 1,600; however, the growth rate decreased gradually. At an MOI of 200 and 400, the transfection efficiencies were (82.33 ± 2.27%) and (92.84 ± 1.14%), respectively. When the MOI value was 800, the transfection efficiency reached its peak (97.08 ± 0.92%), and decreased slightly at an MOI of 1,600 (94.9 ± 0.63%) (Fig. 2C).
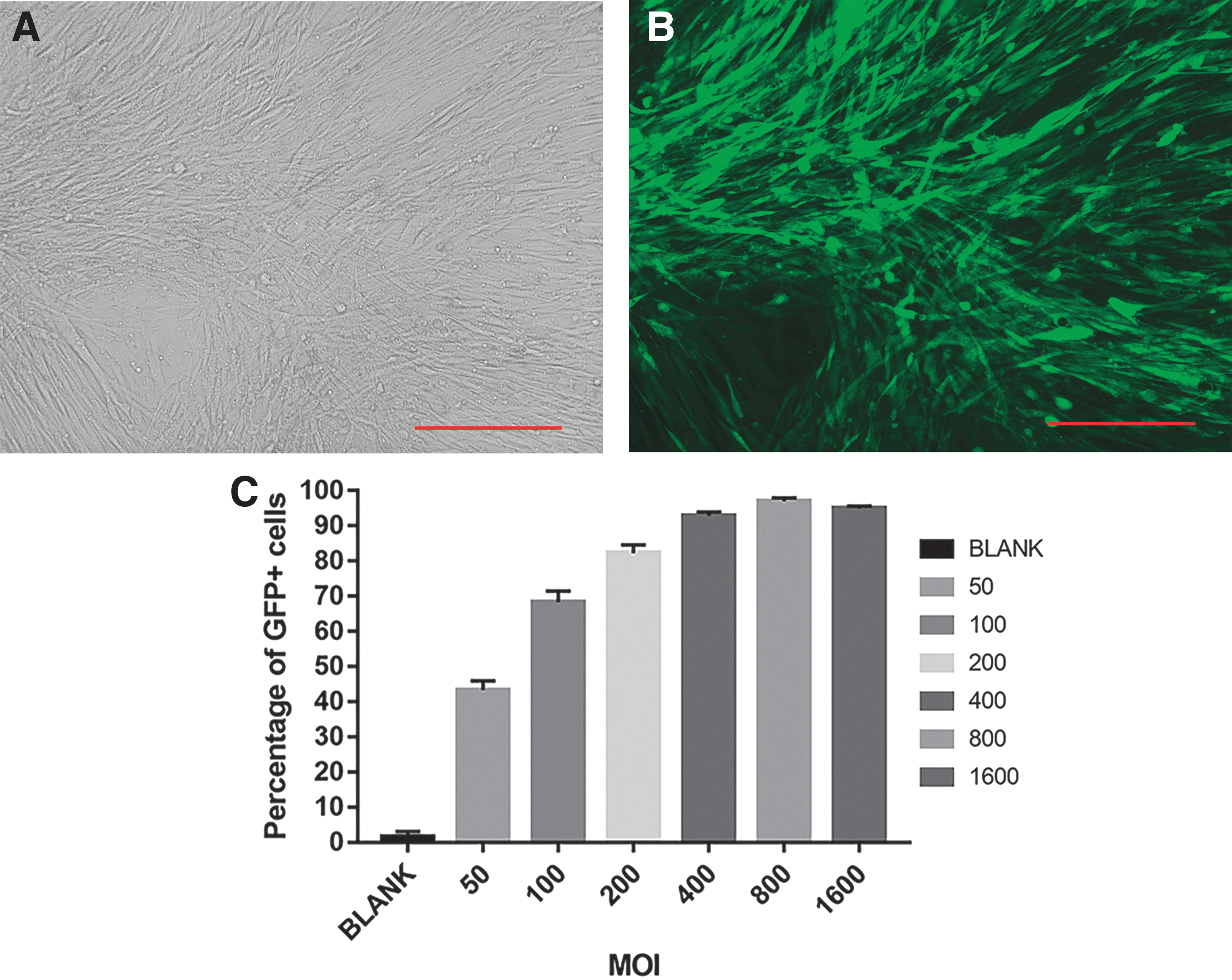
Baculovirus-adeno-associated viral infection of BMSCs.
Cytotoxicity assay
At an MOI of 100, 200, 400, 800, and 1,600, P values of unpaired t-test were 0.27, 0.09, 0.06, 0.04, and 0.0006, respectively. The CCK-8 assay showed that the baculovirus-derived vector had no significant toxic effects on BMSCs at an MOI <400; however, substantial toxic effects were detected at an MOI ≥800 (Fig. 3).
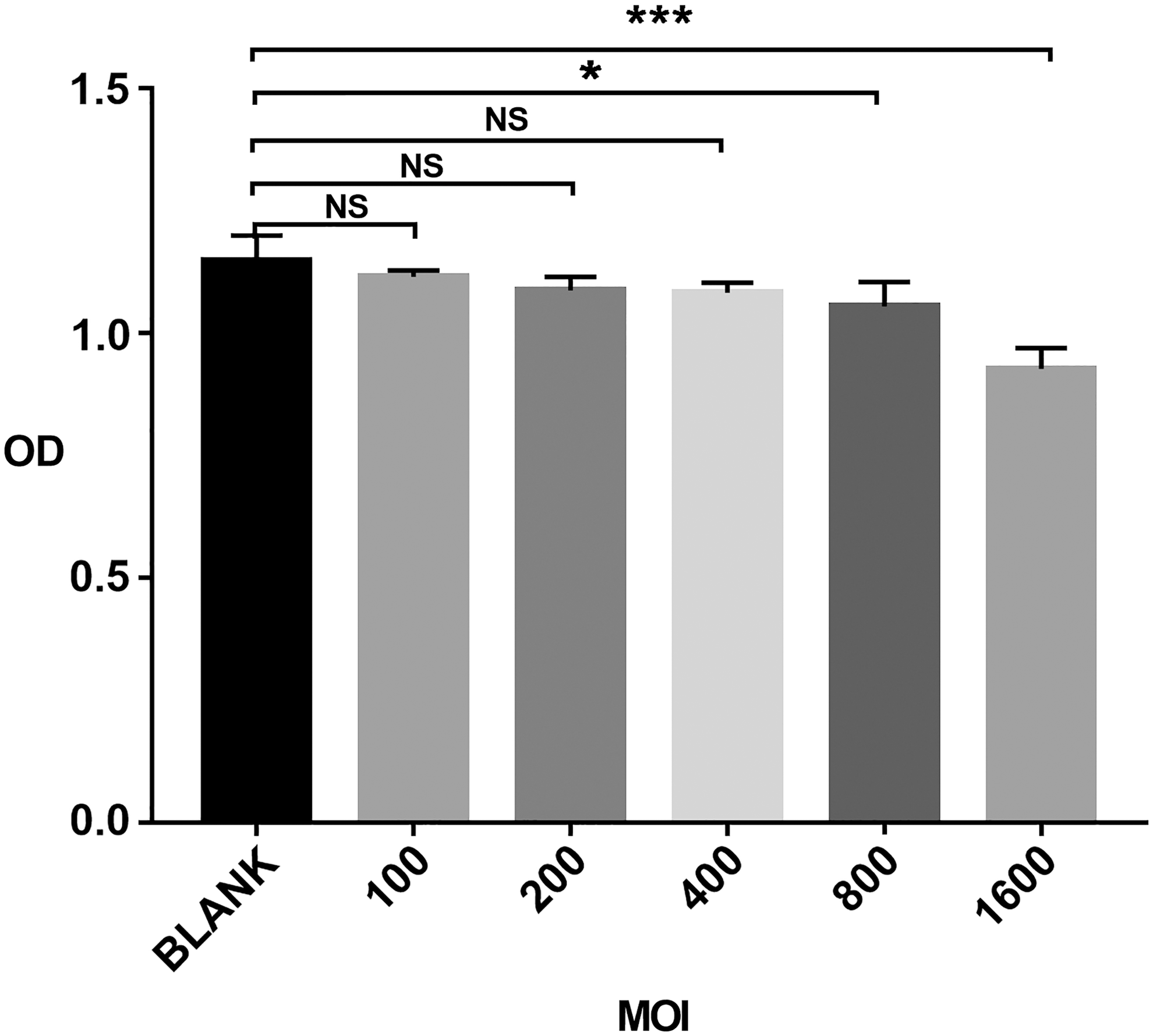
Toxic effects of the baculovirus-derived vector on BMSCs. BMSCs (5,000 cells/well) were infected with Bac-CMV-Luc-P2A-eGFP-ITR at an MOI of 0–1,600 for 4 h. In the control group, uninfected BMSCs were cultured with phosphate-buffered saline for 4 h. On the second day, cell viability was determined using the Cell Counting Kit-8 assay. Results are expressed as means ± SD (n = 4); *P < 0.05, ***P < 0.001, NS, not significant; SD, standard deviation.
Radioiodine uptake of BMSCs-Bac-NIS in vitro
As shown in Fig. 4A,125 I uptake of BMSCs-Bac-NIS increased rapidly over time. High levels of 125I uptake were observed at about 15 min, peaking at nearly 60 min; high levels were then maintained during the 2 h of testing. The uptake levels of the experimental group and control group differed by about 10-folds at the same time points. The 125I uptake of BMSCs-Bac-NIS was significantly blocked by treatment with the (Na+, K+)-ATPase inhibitor NaClO4 (30 μM) (Fig. 4B), indicating that NaClO4 efficiently inhibited the NIS-mediated radioiodine uptake of BMSCs.

125I uptake and inhibition of BMSCs-Bac-NIS in vitro.
In vitro migration of BMSCs
Transwell migration assay showed dramatic increases in the migration of BMSCs cocultured with FaDu cells. Moreover, BMSCs displayed significant tumor-targeting ability after infection with BV-AAV, displaying no significant difference in tumor-targeting ability in comparison with the uninfected BMSCs incubated with FaDu cells. By contrast, few migrated cells were observed in the control group (Fig. 5). Thus, the BMSCs maintained their tumor tropism capabilities toward FaDu cells, even after infection with the baculovirus-derived vector.

Migration capacity of BMSCs assessed through the Transwell migration assay.
Detection of BMSCs-ffLuc in vitro
Luciferase activity decreased in a cell number-dependent manner in BMSCs-Bac-Luc-P2A-eGFP in vitro; the lowest number of BMSCs-ffLuc detected was 625 (Fig. 6). No BLI signal was detected in the control group.
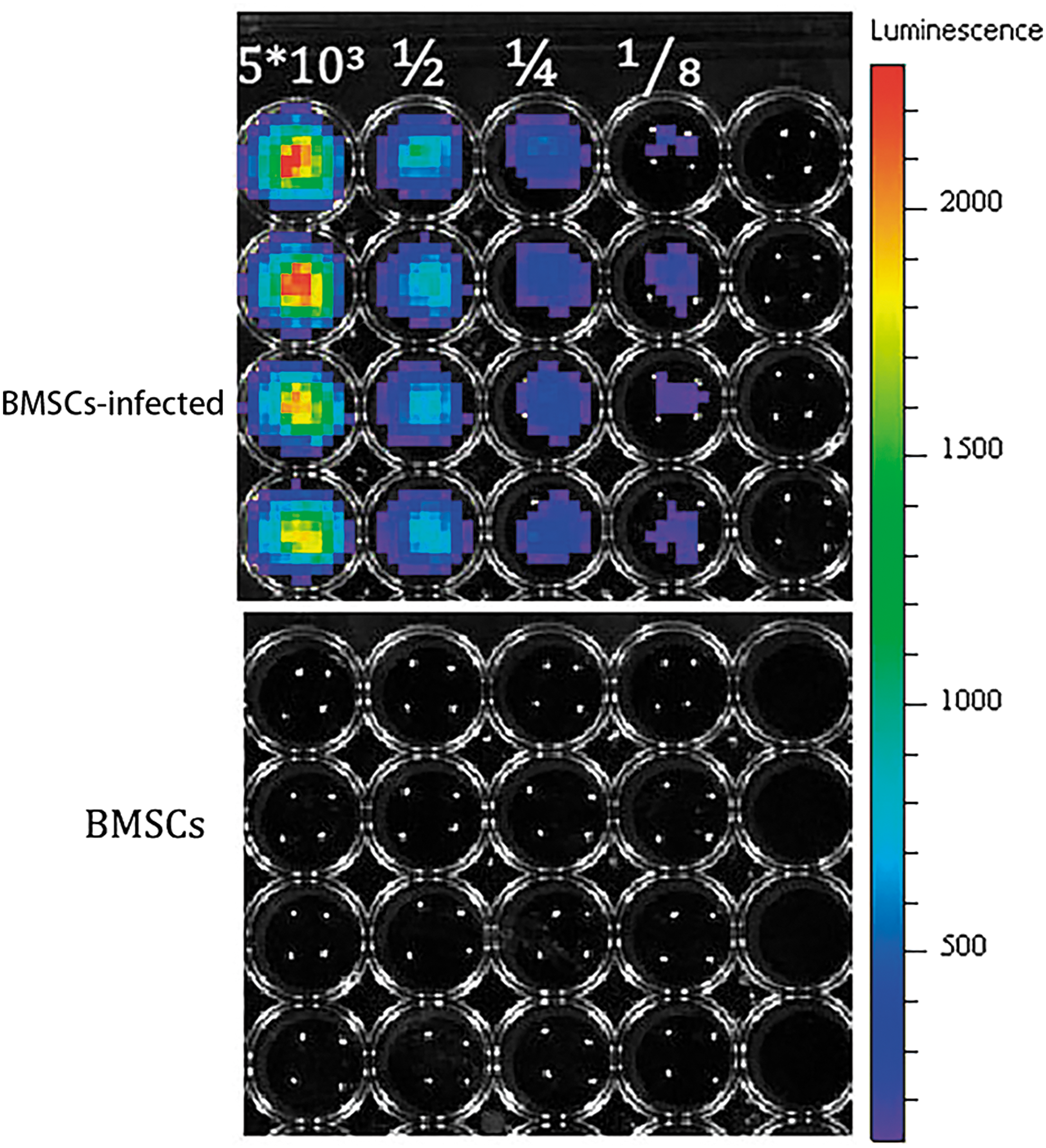
In vitro BLI of BMSCs using the IVIS@ Lumina II system. BMSCs-Bac-Luc-eGFP and uninfected BMSCs, respectively, were cultured with Luciferin (0.3 mg/mL) for 2 min, and then were detected by BLI.
Biodistribution of BMSCs in nontumor BALB/C nude mice
Bioluminescence signals were detected by 4 h postinjection through the tail vein in nontumor nude mice. The signal was enhanced in the lung 8 h following injection and was found to disappear completely at 24 h after injection (Fig. 7). Interestingly, some of the ffLuc signals were retained at the site of the wound caused by injection in the tail at 24 h after injection.
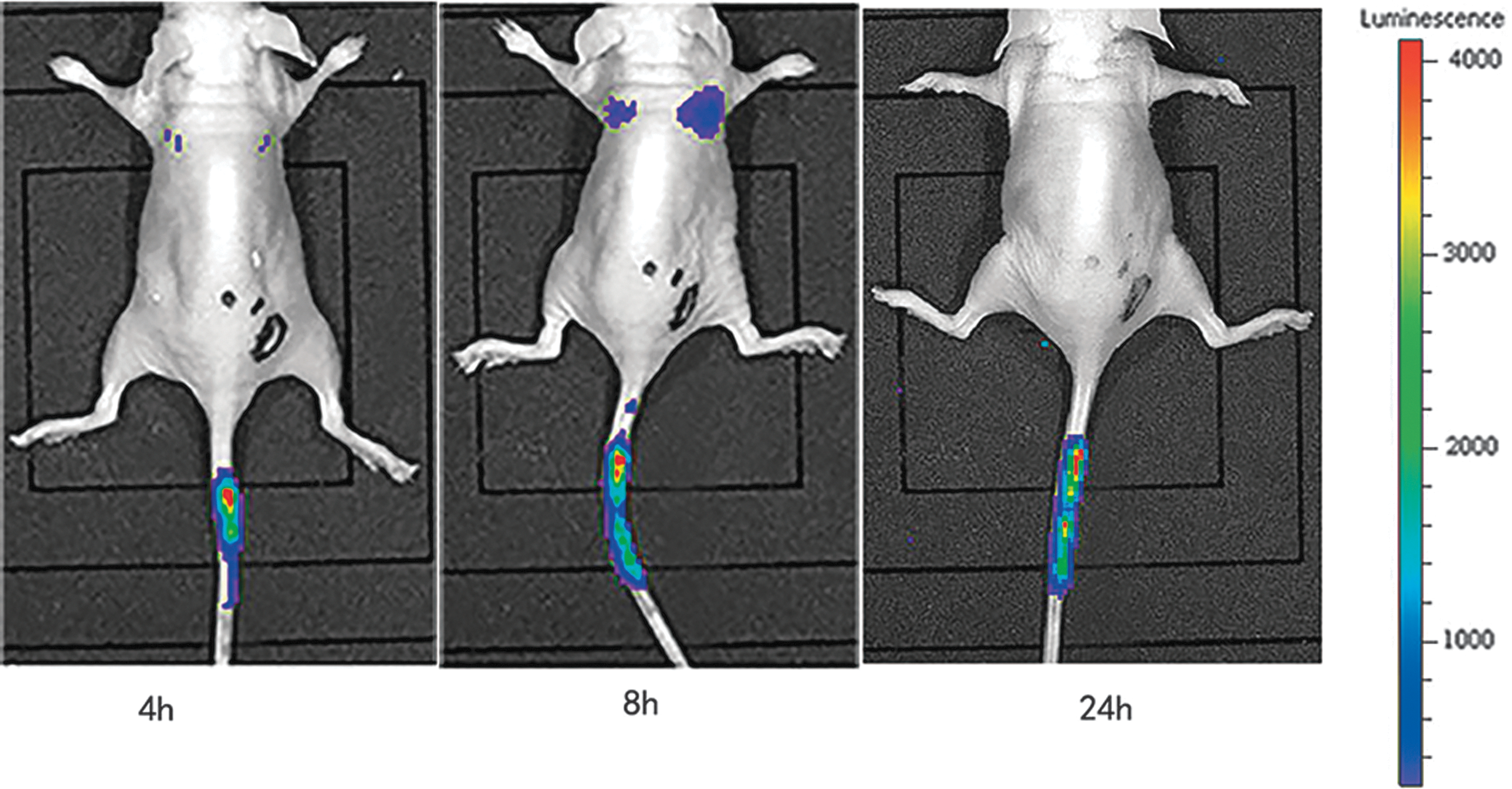
Biodistribution of BMSCs in nontumor BALB/c nude mice. BLI of tumor-free nude mice was performed at 4, 8, and 24 h after injection of BMSCs-Bac-Luc-eGFP into the tail vein.
Tumor tropism and immunostaining of BMSCs in hypopharyngeal carcinoma model mice
As shown in Fig. 8A, the bioluminescent signal accumulated in the lungs of the hypopharyngeal carcinoma model mice on the first day after injection, demonstrating that initial localization of BMSCs occurs in the lung. Visualization of the BLI signal from the tumor site further confirmed that BMSCs were first detectable on day 2 posttransplantation. However, the BLI signal was also detected in the lungs, but at a decreased intensity compared with that detected on the first day; the intensity of this signal then continued to decrease over the next 3 days. On the fifth day after injection, the signal could be found only in the tumor site. One week after injection, no BLI signal was observed in the tumor area or in any other part of the body. No BLI signal was detected in any other organ, such as in the liver, heart, or kidney, throughout the assessment period. These results suggested that BMSCs are capable of specifically targeting hypopharyngeal tumors as a carrier. Immunofluorescence analysis further revealed that the incorporation of the BMSCs-Bac-Luc-P2A-eGFP increased at the tumor site (Fig. 8B).
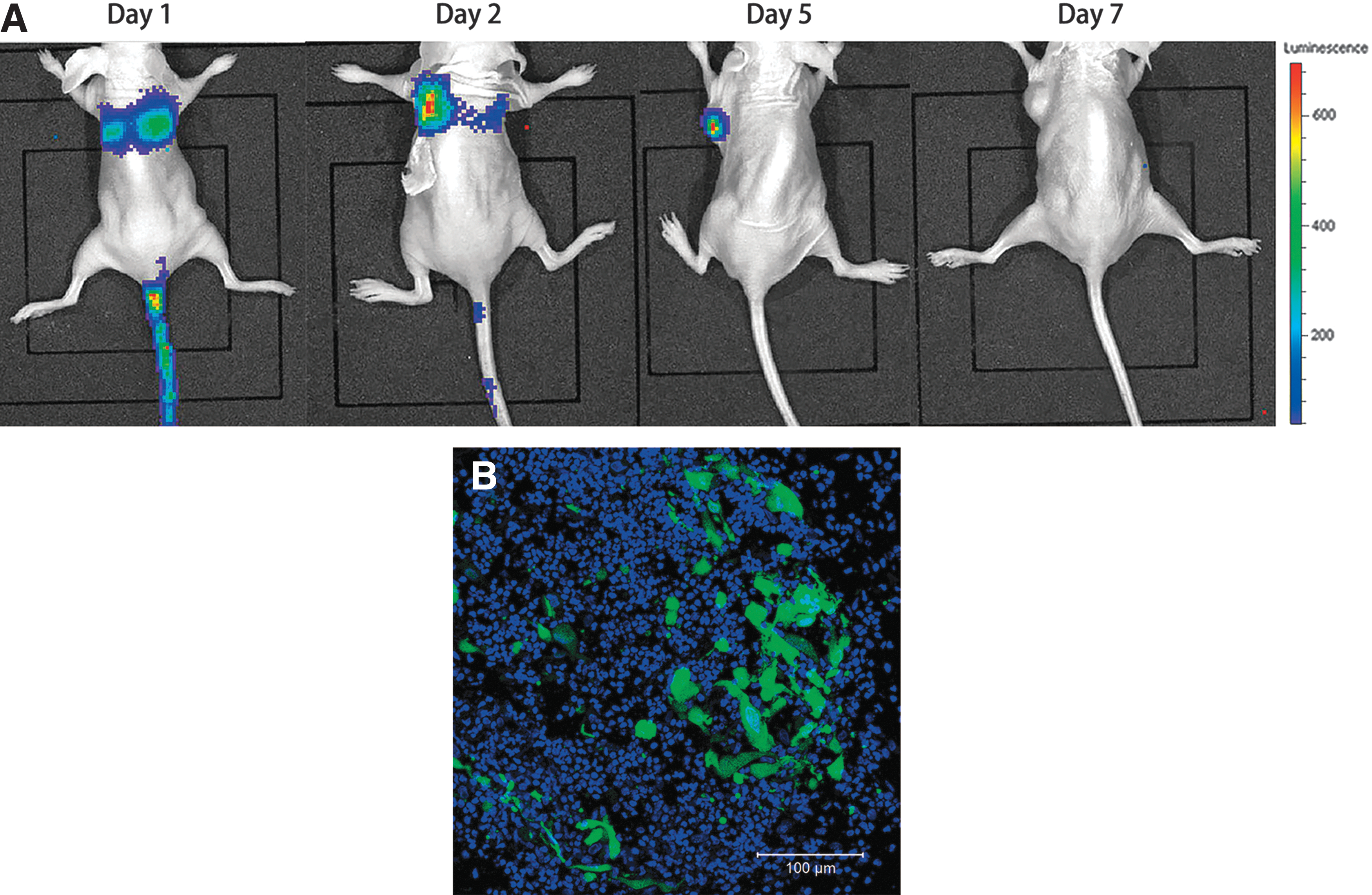
Biodistribution of BMSCs-ffLuc in hypopharyngeal tumor models at various time points over a 1-week period.
Micro-SPECT/CT imaging and immunostaining of hypopharyngeal tumors in vivo
No radioactive uptake was detected by SPECT/CT 1 h after 125I injection in the control group; however, in the tumor-BMSCs-Bac-NIS group, obvious radioiodine uptake was observed at the tumor site (Fig. 9A, B). In addition, high radioactivity levels were detected in some highly endogenous NIS-expressing organs, including the salivary glands, thyroid gland, and stomach. Furthermore, urine radioactivity levels were markedly high, as confirmed by urinary elimination of 125I. Immunofluorescence performed on tumor sections revealed positive NIS staining (Fig. 9C, D). As shown in Fig. 9D, some of the BMSCs that accumulated at the tumor site were in the cell division phase, further indicating their capacity to migrate to the tumor and proliferate in the cancer microenvironment.
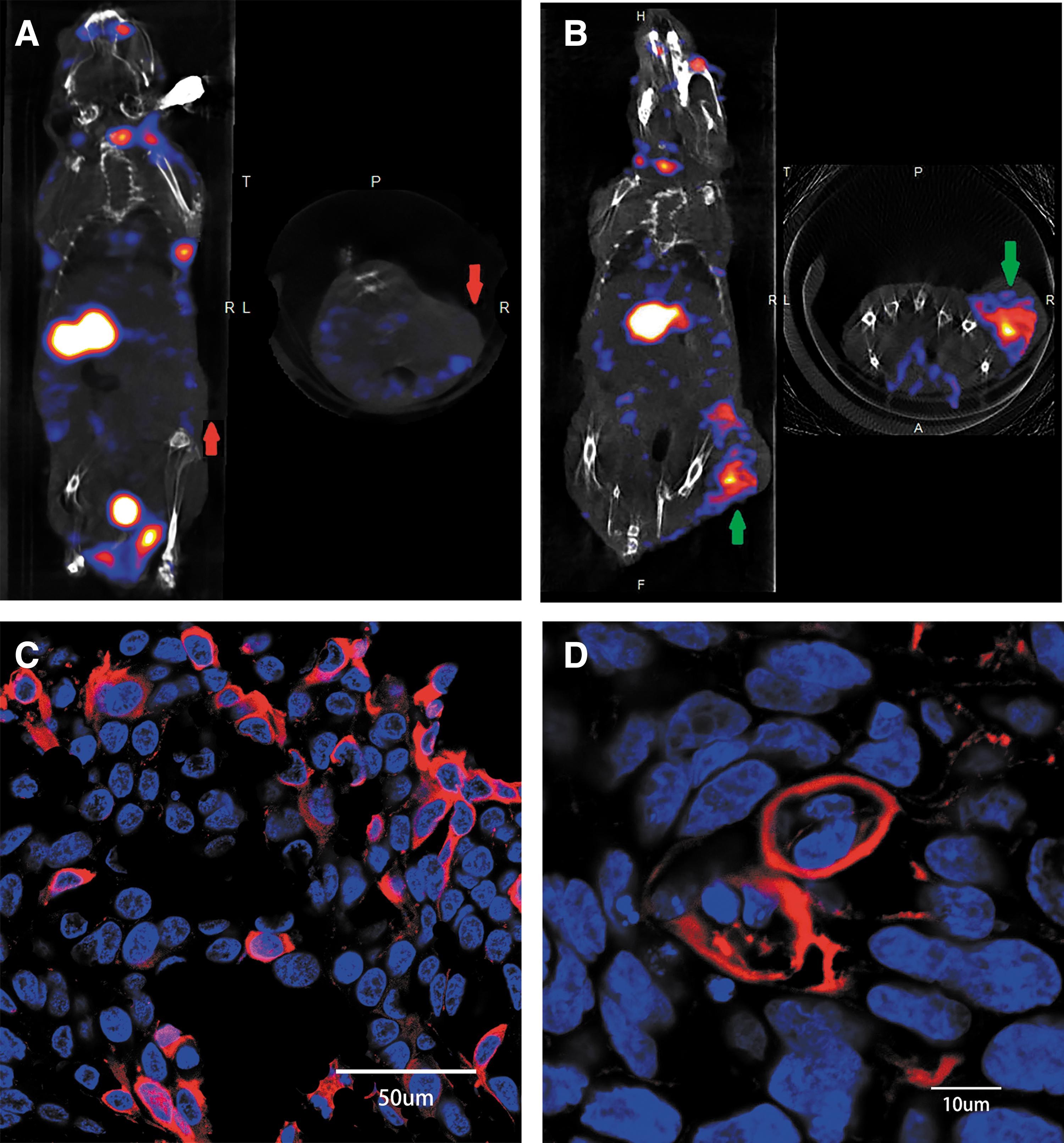
Micro–single-photon emission computed tomography/computed tomography imaging of hypopharyngeal-tumor-bearing model mice and immunofluorescence staining.
Discussion
MSCs have attracted substantial attention owing to two main characteristics that make them ideal vectors for the delivery of therapeutic molecules, with potential applications as carriers for cancer gene therapy. The first characteristic is their low immunogenicity, which is attributable to the low expression levels of major histocompatibility complex (MHC) class I and costimulatory molecules, and the lack of MHC class II expression [23]. The other significant feature of MSCs is their high affinity for tumor tissues, which supports their application in gene therapy for cancer [24 –26].
However, the homing abilities, biodistribution, and localization of BMSCs in a hypopharyngeal cancer model are poorly understood. Herein, we provide direct evidence for the tropism of human BMSCs in vivo using a hypopharyngeal cancer model based on BLI. In nontumor nude mice BMSCs-Bac-Luc-eGFP were injected through the tail vein. Bioluminescence signals were detected at 4 h postinjection, and the signal was enhanced in the lung 8 h following injection. At 24 h after injection, no signal was found in the lung; however, some of the ffLuc signals were retained at the site of the wound caused by injection in the tail, suggesting that BMSCs are capable of homing to sites of injury.
Specifically, we found that BMSCs could effectively home to hypopharyngeal cancer tissues, without any signal detected in the liver, heart, brain, and other essential organs. The BMSCs that accumulated in the lungs gradually disappeared after 2 days of BMSC transplantation. These findings suggest that BMSCs serve as a promising targeted-delivery vehicle for hypopharyngeal carcinoma gene therapy. The potential of MSCs as a promising targeted-delivery vehicle for cancer therapy warrants further validation using MSCs from different sources and different tumor types.
Interestingly, some of the BMSCs that accumulated at the tumor site were in the cell division phase, indicating their capacity to migrate to the tumor and proliferate in the cancer microenvironment. However, at more than 7 days after injection, no ffLuc signal was detected at the tumor site. This suggested that despite the proliferation potential of BMSCs, they did not become a permanent constituent of the tumor site.
The mechanism and factors contributing to the ability of BMSCs to migrate to the tumor microenvironment have not yet been clearly elucidated. Interaction between cytokines or chemokines produced from tumor tissues and the corresponding receptors secreted by MSCs may play an important role in this process [27,28]. Tumors continuously secrete cytokines, chemokines, and other inflammatory mediators into the tumor microenvironment, which potentially serve as ligands for MSC receptors [29 –31]. Growth factors, Toll-like receptors, chemokines, and chemokine receptors are present on MSC surface.
The mechanism underlying MSC migration toward tumors is similar to that underlying the migration of leukocytes through integrin and adhesion molecules [32,33]. Cytokines and inflammatory mediators, such as epidermal growth factor, fibroblast growth factor, interleukin (IL)-6, IL-8, and granulocyte colony-stimulating factor aid in MSC migration [34,35]. Numerous studies have shown that MSCs accumulate at tumor sites, differentiate into pericytes, and secrete cytokines or chemokines to stimulate tumor growth and vascularization [36,37]. However, some studies have suggested that MSCs are involved in the downregulation of Akt, NF-κB, and Wnt signaling pathways, which may contribute to their antitumor potential [38]. Importantly, MSCs are widely recognized as an attractive carrier for targeted antitumor gene therapy [39].
The development of BMSCs as vectors for the delivery of therapeutic agents necessitates appropriate genetic modification of these cells. Currently, the clinical applications of the most common methods of gene delivery are limited owing to challenges, such as low transfection efficiencies of nonviral vectors [40], relatively small transgene size and requirement of helper viruses for AAV activation [41], and the potential tumorigenicity and insertion mutagenesis of lentiviruses or retroviruses [42]. By contrast, the baculovirus genome, along with large DNA inserts, is relatively large (38 kb) and does not insert into the DNA of the host cells, thus avoiding the risk of insertional mutagenesis [43].
Additionally, numerous studies have confirmed that the baculovirus transduction efficiency for stem cells is not very high. Although the addition of sodium butyrate has been shown to increase the transfection efficiency [44,45], its possible cytotoxicity limits its clinical application [46]. Many studies have indicated that the transfection efficiency of mammalian cells may be improved by modifying a plasmid vector with ITRs, which are key elements of AAV, as they allow for virus replication, encapsidation, and long-term maintenance and expression in target cells [47 –51].
BA-AAV have been indicated to efficiently infect primary cells [52] and prolong transient gene expression in human neuronal and glial cell lines [53]. Consistent with these findings, in the present study, a recombinant baculovirus vector containing two AAV ITRs was efficiently transfected into BMSCs (97.08%) without requiring sodium butyrate, demonstrating superior efficiency compared with the transgene expression level (75.8%) reported in a previous study [54]. Therefore, BV-AAV vectors represent a novel, highly efficient, and noncytotoxic method for the functionalization of MSCs.
The viral 2A peptide-coding sequence is small (18–22 amino acids), but enables the expression of discrete protein products within a single vector through a novel “cleavage” event within the sequence to achieve a high efficiency of multigene coexpression [55 –57]. Based on the large packaging capacity of baculoviruses, we designed and constructed a P2A peptide-linked multicistronic vector; this vector was used to express the two discrete proteins, EGFP and firefly luciferase. Therefore, our results further demonstrate that the P2A peptide is an ideal tool for achieving multigene coexpression from a single ORF.
In recent years, NIS has been widely used as both a reporter gene and a therapeutic gene for radionuclide imaging and therapy [58,59]. However, this is the first report of the radionuclide imaging of NIS carried by BMSCs in hypopharyngeal carcinoma in vivo. Radioiodine uptake rapidly increased with time in BMSCs-Bac-NIS, reaching a peak at ∼60 min in vitro. However, 125I uptake did not decrease over time, in contrast to a previous report [60]. This finding supports that the imaging of BMSCs at targeted sites can be monitored by SPECT at the early phase. More importantly, BMSC-mediated NIS expression represents a promising approach for both radionuclide imaging and targeted radionuclide therapy of hypopharyngeal carcinoma.
One of the limitations of this study is the mode of injection used for the establishment of the hypopharyngeal carcinoma mouse model. In the heterotopic model, subcutaneous injection has the advantages of convenient vaccination and easy observation; however, its biological behavior often limits tumor growth and infiltration, and it is difficult to observe important lymphatic and hematogenous metastatic behaviors. In situ implantation of tumors can provide a suitable microenvironment for tumor growth and metastasis to maintain the original tumor tissue structure. However, the procedure is difficult to perform, and instead of real tumor cell transfer, it easily leads to the artificial diffusion of tumor cells. The role of nude mice as an animal model for medical research is enormous. However, as the immune system of nude mice differs from the human immune system, it is difficult to comment on the safety of BMSCs for therapy in humans.
In summary, our novel recombinant baculovirus vector containing ITRs and a P2A showed high efficiency for multi-gene co-expression in BMSCs. BLI and micro-SPECT/CT imaging of transplanted BMSCs in a hypopharyngeal carcinoma mouse model demonstrated a strong potential for the use of BMSCs infected by this novel recombinant baculovirus as promising targeted carriers for radionuclide therapy in hypopharyngeal carcinoma.
Footnotes
Acknowledgments
This research was supported by the Project of Shanghai Municipal Commission of Health and Family Planning (grant no. 201540387), The National Key Research and Development Program of China (grant no. 2016YFC0902700), and Shanghai Shenkang Hospital Development Center Foundation (grant no. 16CR2029B).
Author Disclosure Statement
No competing financial interests exist.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.