
Abstract
Epigenetic modifications play an important role in neural development. Trimethylated histone H3 at lysine 27 (H3K27me3) is a repressive epigenetic marker that mediates tissue development. In this study, we demonstrate that H3K27me3 and histone methyl transferase Ezh2 regulated the development of dopaminergic (DA) neurons in vitro and in vivo. We found that H3K27me3 increased during differentiation of ventral midbrain-derived neural stem cells (VM-NSCs). However, histone demethylase selective inhibitor GSK-J1 increased H3K27me3 level and decreased the expression of tyrosine hydroxylase. Treated with Ezh2-selective inhibitor EPZ005687 repressed the trimethylation of H3K27 and enhanced differentiation of DA neurons in VM-NSCs cultures. Furthermore, Ezh2 inhibition promoted the expression of DA neurons developmental-related factors by modifying H3K27 trimethylation on the relevant promoter regions. Moreover, the effect of Ezh2 inhibition-mediated DA neurons differentiation was blocked by the expression of shRNA specific for Nurr1. In vivo, Ezh2 decreased and resulted in a reduction of H3K27me3 in developing midbrain. Deletion of Ezh2 by RNA interference approach promoted differentiation of DA neurons during midbrain development. Overexpression of Ezh2 enhanced cell self-renewal and did not affect differentiation of DA neurons.
Introduction
Midbrain dopaminergic (DA) neurons play an important role in the regulation of voluntary movement, cognition, and emotional behavior [1]. Degeneration of DA neurons in substantia nigra parscompacta affects the motor features and underlies Parkinson's disease (PD) [2]. Ultimately, ventral midbrain-derived neural stem cells (VM-NSCs) are an appropriate cell source for learning the development of DA neurons and it is a candidate cell for PD transplant therapy [3]. A lot of efforts have been put into gene modification or multisteps that generate genuine DA neurons in vivo and engineering of stem cells in vitro [2,4]. The basic research to study the precise molecular mechanisms underlying the generation and survival of DA neurons underlies the better therapies for PD [5].
Trimethylated histone H3 at lysine 27 (H3K27me3) is a repressive epigenetic marker [6 –8]. Pieces of evidence show that H3K27me3 is involved in midbrain DA neurons development and maintenance [9 –14]. During aging, H3K27me3 increases with downregulated histone demethylases Jmjd3 and UTX in midbrain [15]. UTX and JMJD3 are the only two histone demethylases which demethylates H3K27me3 to H3K27me2 or H3K27me1, resulting in a gene derepression [16,17]. Importantly, deletion of Jmjd3 causes an increased ratio of M1/M2 microglia markers and DA neurons death in 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP) models [15,18]. In contrast, knockout histone methyltransferase Ezh2 causes a reduction of H3K27me3 and results in a midbrain to forebrain identity switch in the early stage of development [12]. Ezh2, the core component of polycomb repressive complex 2 (PRC2), trimethylates lysine residues of the core histone H3K27 and represses the expression of nearby genes through the formation of heterochromatic regions [19,20]. However, the midbrain patterning-related factors are not affected in the midbrain lacking Ezh2 [12]. Moreover, increase of H3K27me3 level appears at Bdnf promoter and the expression of brain derived neurotrophic factor (BDNF) reduces by chronic morphine. Consistent with these results, the binding of PRC2 proteins Suz12 and Ezh2 to BDNF promoter is increased in ventral tegmental area [14]. Besides, H3K27me3 increased during differentiation of VM-NSCs in vitro [11,13]. Otherwise, ascorbic acid and other compounds such as Apicidin decrease the level of H3K27me3 and promote VM-NSCs differentiation into DA neurons. The contribution of H3K27me3 and Ezh2 to the development of DA neurons remains unclear.
Our investigation revealed that pharmacological inhibition of Ezh2 enhanced differentiation of DA neurons and increased the expression of DA neurons developmental-related regulators in VM-NSCs. The decline of Ezh2 mediated the reduction of H3K27me3 and promoted differentiation of DA neurons in the developing midbrain. We also showed that upregulated Nurr1 was an important downstream target for the differentiation of DA neurons through the inhibition of Ezh2. These findings suggest the important role of Ezh2 in the development of DA neurons.
Materials and Methods
Animal welfare and ethical statement
All Sprague–Dawley rats were purchased from SLAC Laboratory Animals Co. Ltd. (Shanghai, China). Every rat was kept in an individual cage with random assignment and housed in conditional environment (12 h light and 12 h dark cycle, temperature 20–24°C). The related experiments were guarded and approved by the Institutional Animal Care and Use Committee of Shanghai Institute of Material Medica. And experimental procedures were performed following the rules of the Association for Assessment and Accreditation of Laboratory Animal Care International.
VM-derived NSC cultures
Primary VM cells were prepared from a described protocol with some modifications [21]. VM cells were dissected from E11 Sprague–Dawley rat embryos. After dissection, cells were cultured in Dulbecco's modified Eagle's medium (DMEM)/F12 supplemented with B27 as described [22] and stimulated to proliferation with 20 ng/mL recombinant epidermal growth factor, 20 ng/mL basic fibroblast growth factor, and 1% N2 for 3 days. Then these cells were induced to differentiation in DMEM/F12 medium supplemented with B27 for experiments.
Immunocytochemistry assay
The cells were washed by cold phosphate-buffered saline (PBS) and fixed with paraformaldehyde (PFA, 4%) for 15 min. The cells were blocked and permeabilized for 1 h with blocking buffer (10% normal goat serum supplemented with 0.1% Triton X-100 in PBS) at room temperature before incubation with the primary antibody [mouse anti-BrdU, 1:1,000 (Sigma); mouse antityrosine hydroxylase (TH), 1:500 (Millipore); mouse anti-Nurr1, 1:1,000 (Abcam); rabbit anti-Tuj-1, 1:500 (Cell Signaling Technology); rabbit anti-Map2, 1:500 (Abcam); rabbit anti-GFAP, 1:1,000 (Abcam)] at 4°C overnight. On the second day, the cells were incubated with Alexa Fluor 488-conjugated secondary antibodies (1:500) at room temperature for 1 h after being washed with PBS.
Western blot
Protein extraction was prepared as described [22]. Twenty micrograms of protein samples ran on a 10% sodium dodecyl sulfate–polyacrylamide gel and transferred onto a polyvinylidene difluoride membrane (0.45 or 0.22 μm). The target membranes were incubated with primary antibodies in blocking buffer at 4°C overnight and then incubated with the secondary antibodies conjugated with horseradish peroxidase (HRP) (1:5,000). The immunoreactive proteins were visualized with an enhanced chemiluminescent kit. Protein bands were quantified with ImageJ. Primary antibodies were directed against the following: rabbit anti-H3K27me3 antibody (1:200; CST), rabbit anti-Jmjd3 antibody (1:1,000; Proteintech), rabbit anti-UTX antibody (1:1,000; Proteintech), rabbit anti-Ezh2 antibody (1:1,000), mouse anti-TH antibody (1:100; Millipore), and mouse anti-Nurr1 antibody (1:100; Abcam).
Chromatin immunoprecipitation assay
ChIP Kit was purchased from Cell Signaling Technology. Cells were treated with 4% PFA for 15 min at room temperature, then added glycine into medium to stop the crosslink. Ultrasonic treatment isolated chromatin to get shear DNA. About 50 μL solution was reserved as input, and the rest of the solution was immunoprecipitated with H3K27me3-specific antibodies. The purified DNA was analyzed by quantitative real-time polymerase chain reaction to identify H3K27me3 promoter occupancy.
Virus infection
The adeno-associated viruses (AAVs) shRNA, overexpression sets for Ezh2, and lentiviral shRNA sets for Nurr1 were purchased from Obio Technology Corp., Ltd. (Shanghai). For adeno-associated viral transfection, E11 embryos were observed under a microscope and microinjected with 1 μL into the midbrain. Then E18 embryos were assessed for western blotting and immunohistochemistry assays. VM-NSCs were treated with lentivirus for 48 h and used for further experiments. The shRNA sets are as follows: Ezh2, 5′-CCGGCAGCAGAAGAACTGAAAGA-3′, 5′-TCTTTCAGTTCTTCTGCTGTTTTTG-3′; Nurr1, 5′-CCGGGGTCTTGCACAGGTTGCAA-3′, 5′-TTGCAACCTGTGCAAGACC-3′.
Cell counting
Number of cells was determined by counting the number of biomarker positive cells per field at 40 × magnification from 10 random fields in three samples per VM-NSCs or at 10 × magnification from 10 random fields in three samples per brain slices using an Olympus fluorescent microscope (Olympus). The number of cells shown in the figures represent the average number of cells and standard error of the mean (SEM) from 10 random fields in three samples per VM-NSCs.
Statistical analysis
The data obtained are the means ± SEM from three or more independent experiments. Statistical analysis was performed by using one-way analysis of variance (ANOVA) and the Dunnett test. Statistical significance (*P < 0.05, **P < 0.01, and ***P < 0.001) was determined with GraphPad Prism 5 software.
Results
H3K27me3 increases during differentiation of VM-NSCs and upregulated H3K27me3 causes VM-NSCs to lose differentiation potential to DA neurons
To investigate the variation of H3K27me3 during differentiation of VM-NSCs, we measured its change with western blot. The level of H3K27me3 gradually increased with a time-dependent manner (Fig. 1A). The level of H3K27me3 is mainly determined by histone demethylases UTX, Jmjd3 [23], and histone methyltransferase Ezh2 [24]. Consistent with H3K27me3 change, immunoblotting analysis revealed that Jmjd3 and UTX were reduced (Fig. 1B, C). Besides, histone methyltransferase Ezh2 marginally changed during differentiation of VM-NSCs (Fig. 1D). To investigate the significance of the increase of H3K27me3 level, we used histone demethylases inhibitor GSK-J1 [25,26] and Ezh2-selective inhibitor EPZ005687 [27,28] to assess whether increase of H3K27me3 level could affect the differentiation potential of VM-NSCs to DA neurons. Actually, TH expression level increased during normal differentiation (Fig. 2A, D). In contrast, 0.5 and 1 μM GSK-J1 resulted in an increase of H3K27me3 level, followed by reduced protein level of TH in VM-NSCs cultures (Fig. 2B, D, E, G). EPZ005687 caused a reduction of H3K27me3 in a dose-dependent manner and resulted in an increase in TH level (Fig. 2C, D, F, G). These results suggest that the increase of H3K27me3 level during differentiation of VM-NSCs delays its differentiation potential to DA neurons.
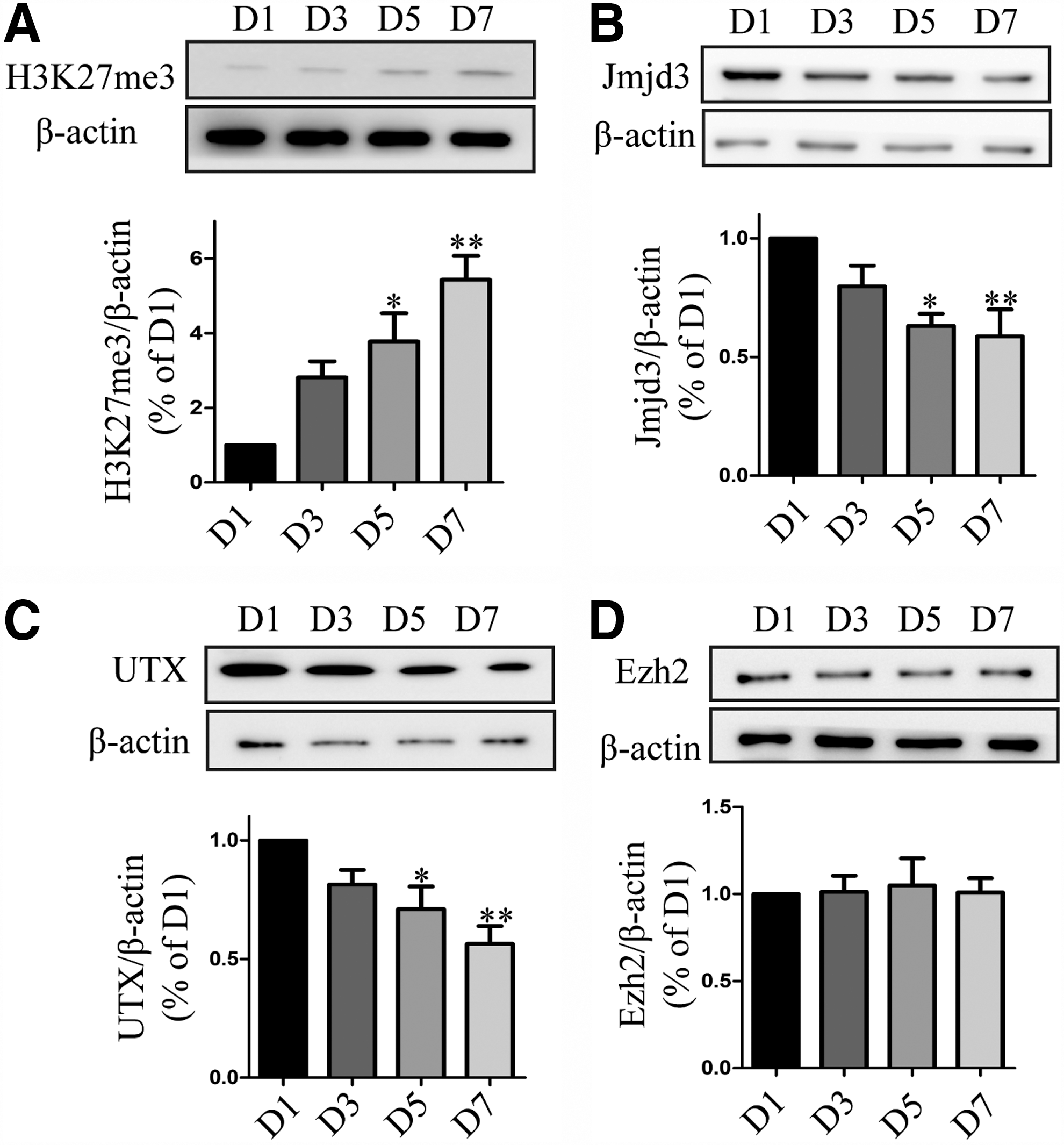
H3K27me3 increases during differentiation of VM-NSCs. VM-NSCs were dissected from E11 rats and expanded in the presence of 20 ng/mL EGF, 20 ng/mL bFGF, 2% B27, and 1% N2 in DMEM/F12 medium for 3 days. Expanded cells were differentiated in DMEM/F12 with 2% B27 medium without EGF, bFGF, and N2.
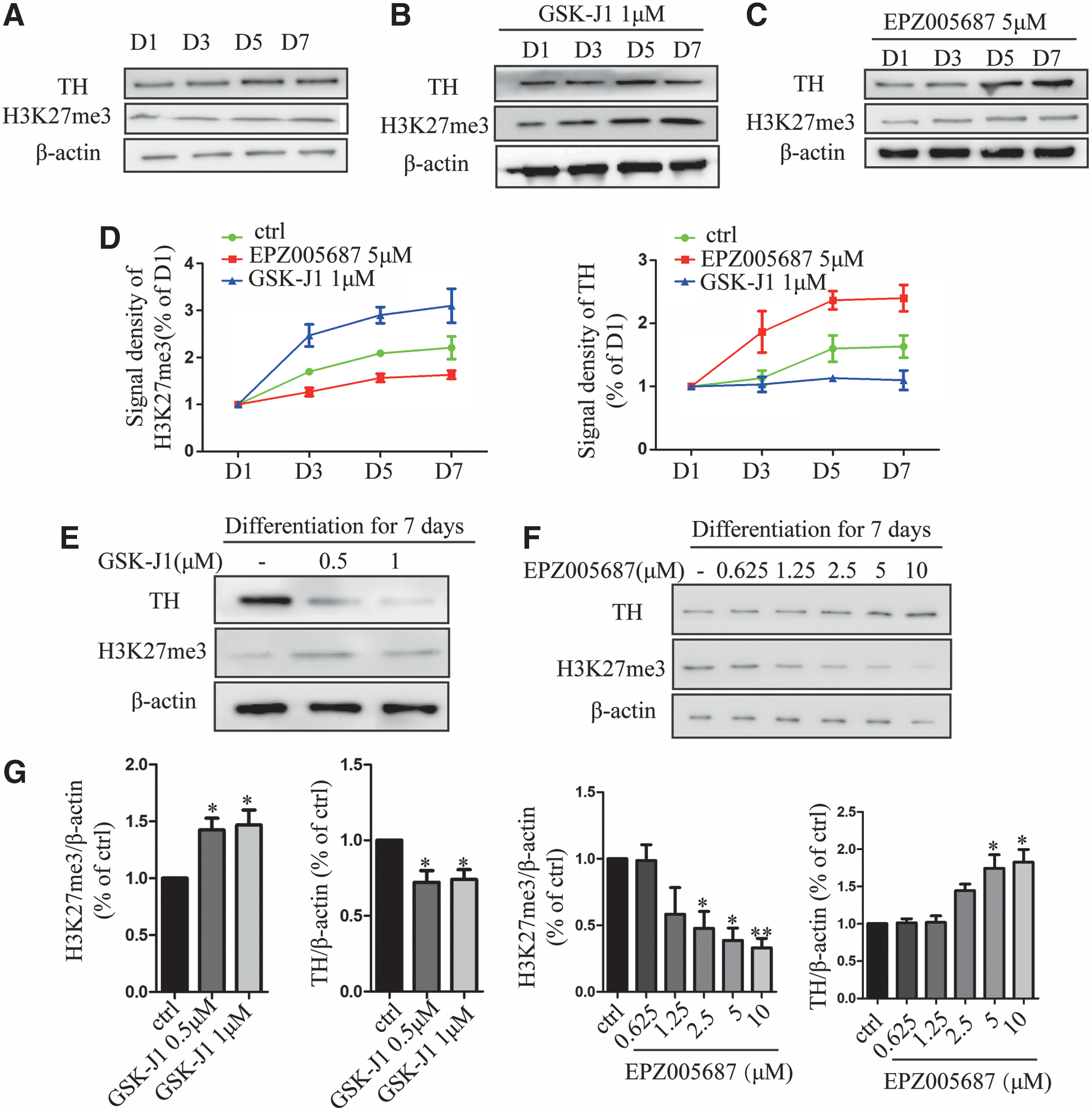
Upregulated H3K27me3 delays the potential of differentiation of VM-NSCs into DA neurons.
Inhibition of Ezh2 promotes the expression of DA neurons developmental-related factors by modifying H3K27me3 occupation on promoters
To further investigate the effect of Ezh2 inhibition on differentiation of DA neurons, we added 2.5 and 5 μM EPZ005687 for 7 days to the expanded VM-NSCs and measured the number of TH+ and Map2+ cells. We found that EPZ005687 treatment increased the proportion of TH+/Map2+ cells (Fig. 3A). Virtually, cells in the EPZ005687-treated cultures also expressed other DA neural markers, including Nurr1 and FoxA2 (Fig. 3B, C).
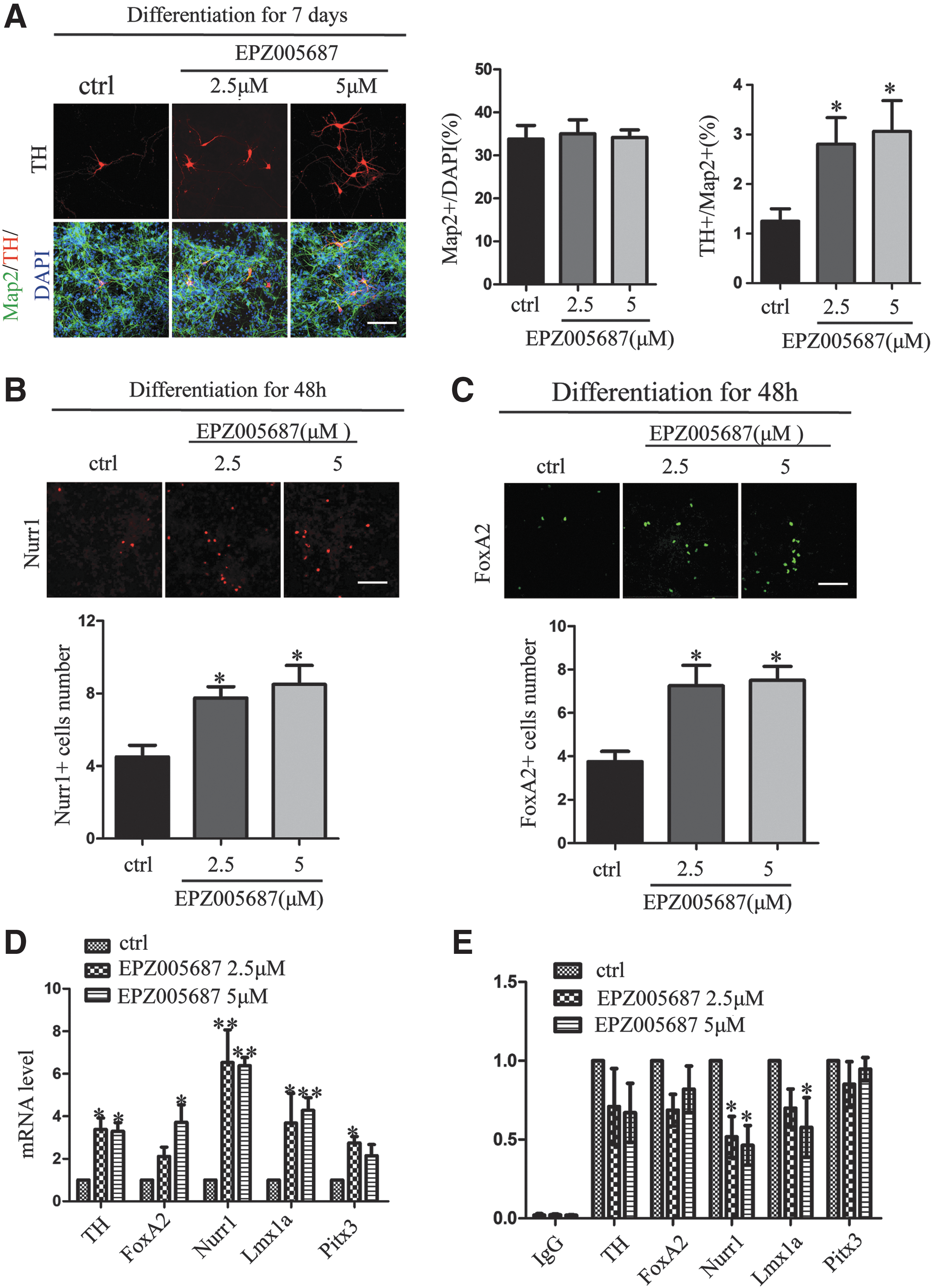
Inhibition of Ezh2 enhances differentiation of DA neurons in VM-NSCs cultures.
Previous studies have proved that the deletion of Ezh2 in cortical progenitors upregulated 1,080 transcripts and downregulated 12 transcripts, and these transcripts were involved in a wide range of functional classes, including neurogenesis and neural differentiation [29]. In our case, we focused on the expression of DA neurons developmental-related factors. The mRNA level of FoxA2, Lmx1a, Nurr1, and Pitx3 was found to have a significant increase in EPZ005687-treated VM-NSC cultures (Fig. 3D). By ChIP assay, we revealed that EPZ005687 reduced the level of H3K27me3 occupation on these promoters, especially Nurr1 (Fig. 3E). Collectively, our data suggest that inhibition of Ezh2 exerted a significant effect on differentiation of DA neurons.
Inhibition of Ezh2 facilitates the differentiation of Nurr1+ precursors in vitro
H3K27me3 and Ezh2 have a critical role in stemness maintenance [30]. To investigate whether the Ezh2 inhibition-mediated differentiation of DA neurons is associated with the alteration of the balance between the self-renewal and differentiation of VM-NSCs, we detected the proliferative ability of VM-NSCs with BrdU uptaking assay. Then 30 μg/mL BrdU was added to VM-NSCs for 24 h on the first day of differentiation of VM-NSCs and then the number of BrdU+ cells was measured. It showed that EPZ005687 repressed proliferation of VM-NSCs (Fig. 4A). Besides, Ezh2 is involved in the differentiation potential of neural stem/progenitor cells in vivo and in vitro [31,32]. We costained Ki67, a nuclear protein that is associated with cellular proliferation, with Nurr1 that is involved in the cell fate of DA neurons. EPZ005687 repressed the expression of Ki67 while the number of Nurr1+ cells was increased (Fig. 4B). In contrast, GSK-J1 treatment increased the ratio of Ki67 and the expression level of Nurr1 was reduced (Fig. 4B). To further investigate the potential of differentiation of VM-NSCs into neurons, astrocytes, and oligodendrocytes, we detected the expression of specific makers (Tuj-1, GFAP, and PDGFR-α) and found no significant differences between the control and EPZ005687 treatment group (Fig. 4C).
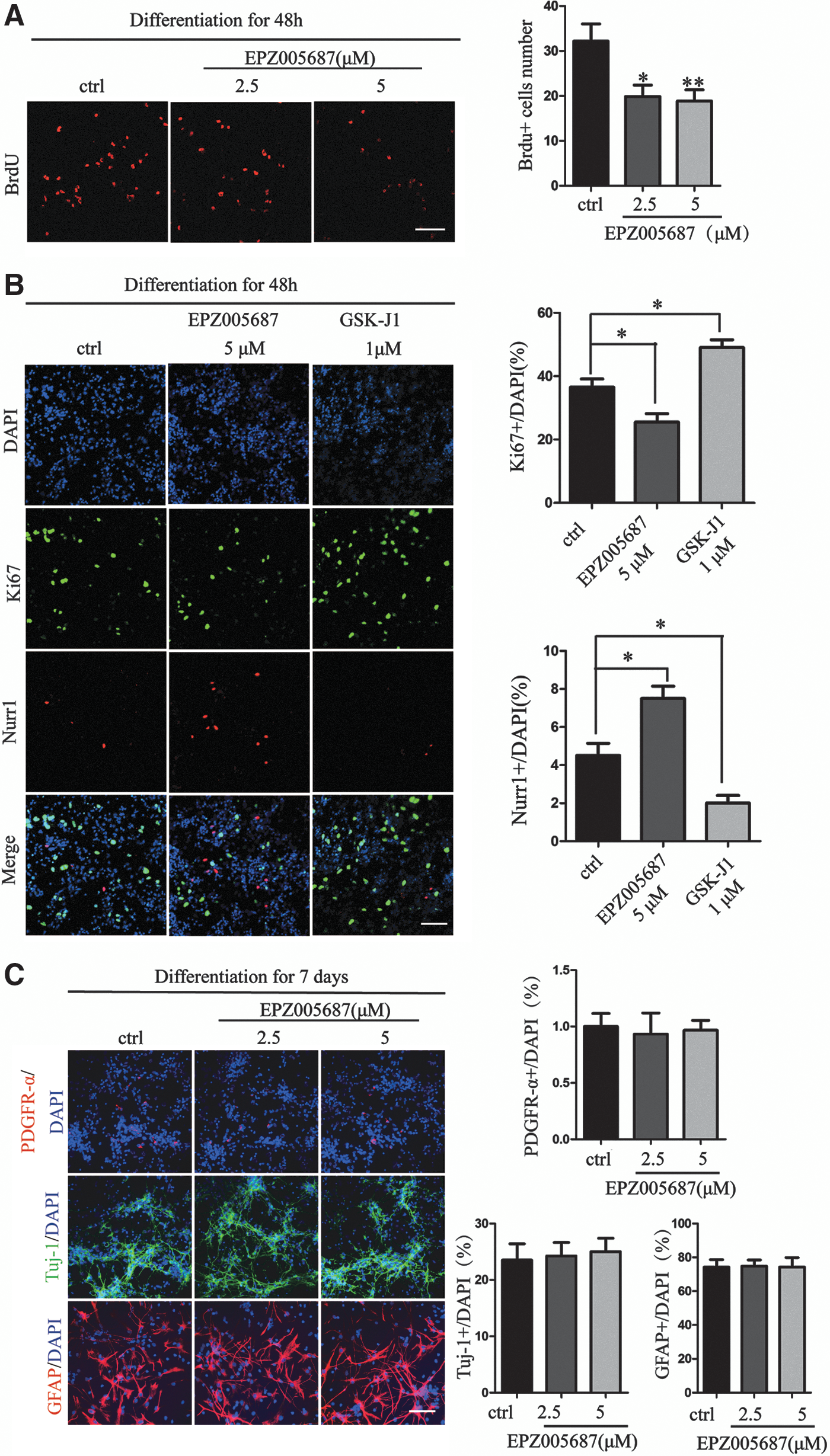
EPZ005687 represses the proliferation of VM-NSCs.
It has been reported that H3K27me3 could regulate the expression of Nurr1 in adult midbrain [14]. To determine whether Nurr1 plays a role in Ezh2 inhibition-mediated differentiation of DA neurons, we used Nurr1 RNA inference (RNAi) with lentivirus approach in parallel with 5 μM EPZ005687 treatment. Knockdown of Nurr1 decreased the density of Nurr1 (Fig. 5A) and the ratio of Nurr1+/DAPI cells was significantly reduced in VM-NSCs cultures. Consistently, we found a decreased ratio in TH+ cells in the knockdown of Nurr1 cultures. Cells depleted of Nurr1 lost their differentiation potential to DA neurons, and EPZ005687 treatment could not reverse the effect of Nurr1 depletion (Fig. 5B). These results together suggested that Nurr1 played a central role in Ezh2 inhibition-mediated differentiation of DA neurons.
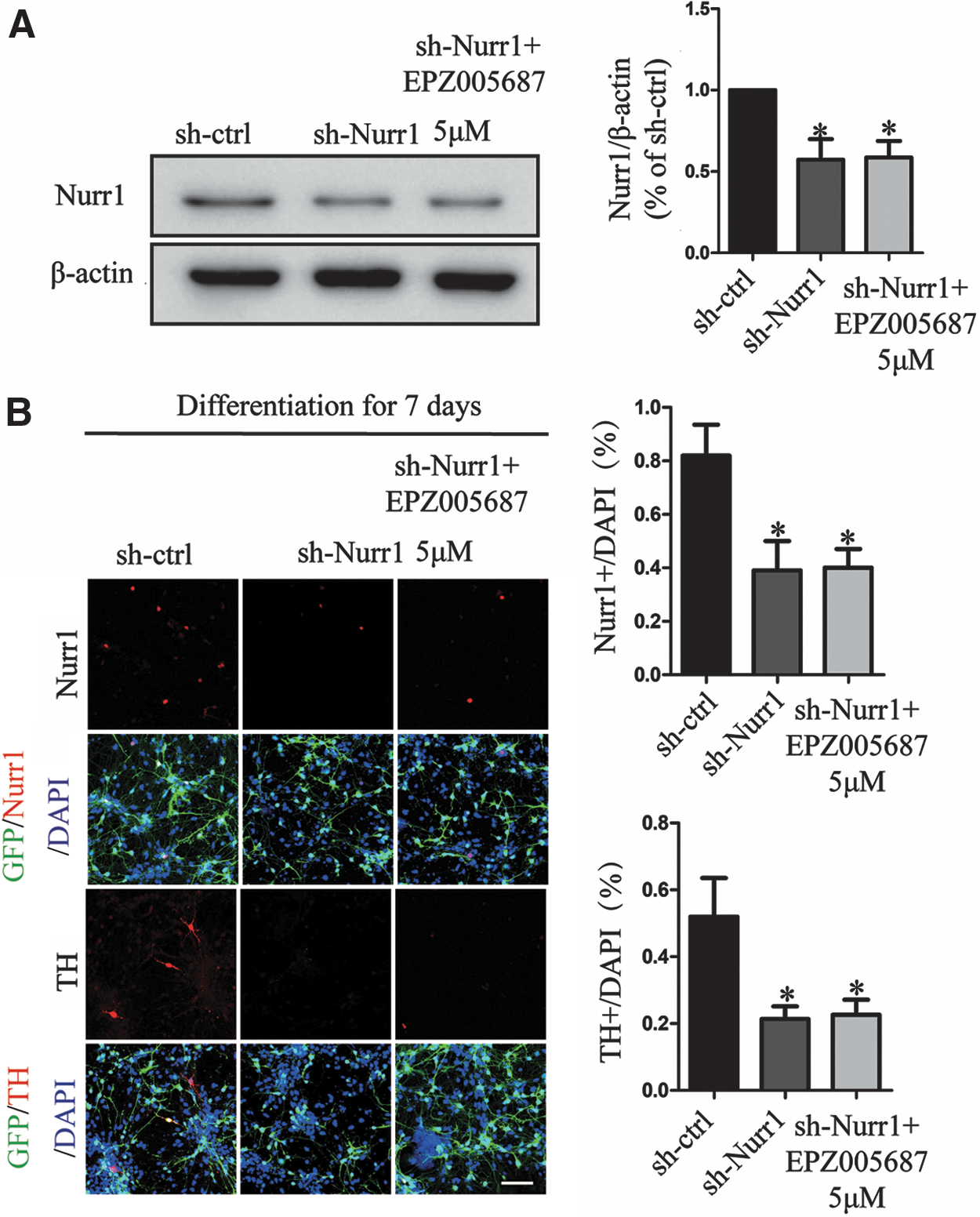
Inhibition of Ezh2-upregulated Nurr1 triggers differentiation of DA neurons. Nurr1 lentiviral shRNA-treated VM-NSCs for 48 h and EPZ005687 incubated with cells persistent for 7 days.
The decline of Ezh2 reduces H3K27me3 in developing midbrain
To further investigate the variation of H3K27me3 in developing midbrain, we measured its change with western blot during E11–E18. We found that H3K27me3 decreased in a time-dependent manner in the developing midbrain (E11: 1 ± 0; E13: 0.694 ± 0.049; E16: 0.490 ± 0.038; E18: 0.0284 ± 0.0438) (Fig. 6A). We measured the protein expression level of Jmjd3 and UTX, the signal density of these two demethylases had marginally changed (Fig. 6B, C). Consistent with the change of H3K27me3 level, Ezh2 was found decreasing during E11–E18 (Fig. 6D, E).
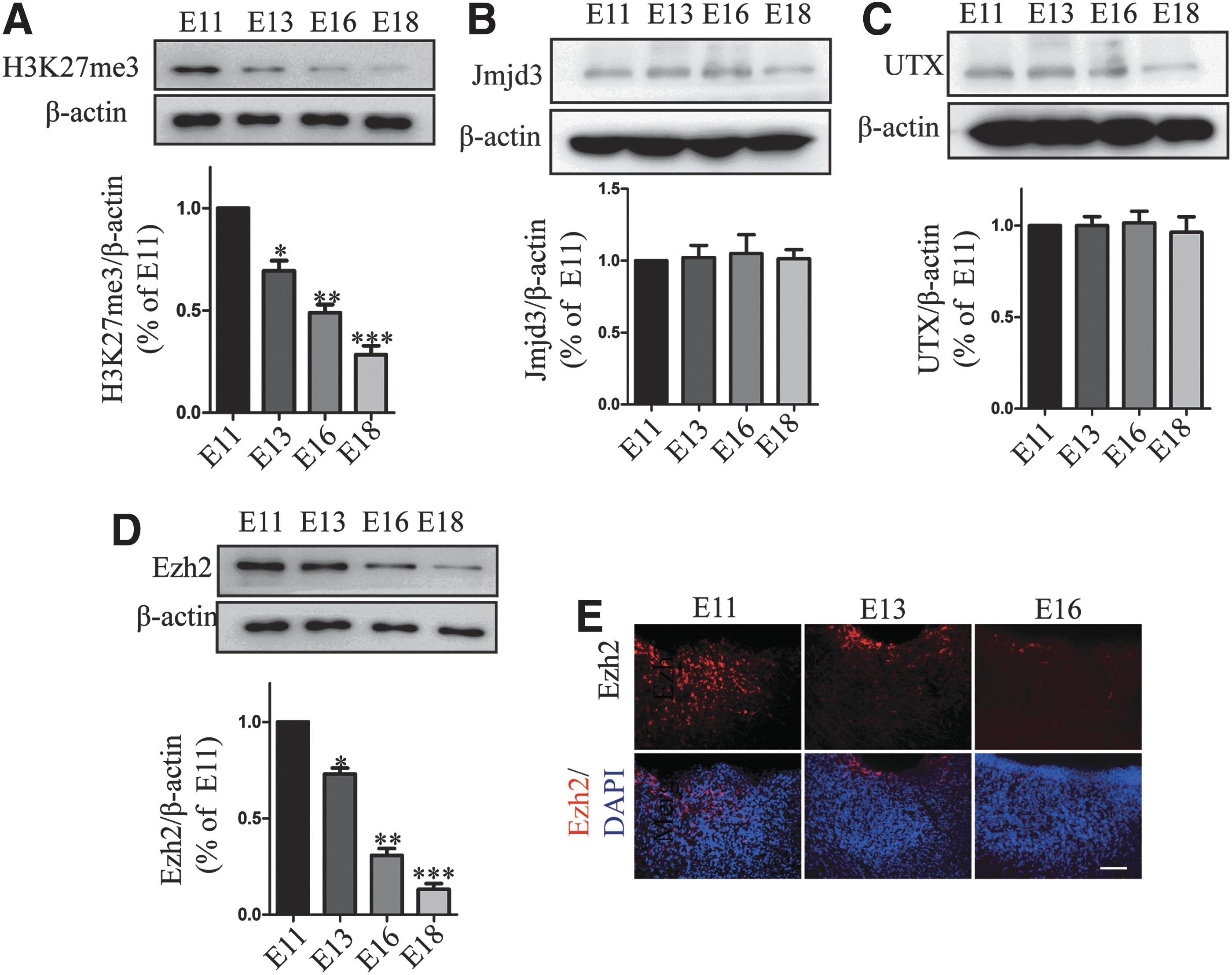
H3K27me3 and Ezh2 decrease in developing midbrain.
Loss of Ezh2 contributes to differentiation of DA neurons in vivo
To assess the effect of Ezh2 decline in vivo, we treated Ezh2 RNAi with AAVs in rat E11 embryos. We detected the effect of Ezh2 RNAi 7 days later. The protein expression level of Ezh2 and H3K27me3 was clearly lower than that of the Ezh2 RNAi group with control group in the midbrain (Fig. 7A). To explore the effect of Ezh2 depletion on differentiation of DA neurons, we counted the number of TH+ neurons (Fig. 7B) and found a significant increase in Ezh2 RNAi embryos than in controls. In addition, Nurr1 expression as well as DA precursor marker was increased with Ezh2 sh-RNA treatment (Fig. 7C). Importantly, we detected the number of TH+ neurons 6 weeks later after AAV injection treatment and found a significant increase in Ezh2 RNAi midbrain than in controls (Fig. 7D). To further confirm the effect of Ezh2 on the development of DA neurons, we overexpressed Ezh2 with AAVs in rat E11 embryos. The expression of Ezh2 and H3K27me3 was detected 7 days later (Fig. 8A). Interestingly, differentiation of DA neurons was not affected by Ezh2 overexpression (Fig. 8B). Since Ezh2 has a critical role in cell self-renewal [33,34], we detected the proliferative ability of cells with Ki67. The increase in number of Ki67+ cells after Ezh2 overexpression was observed (Fig. 8C). These pieces of evidence indicated that the decline of Ezh2 contributed to differentiation of DA neurons. Ezh2 overexpression did not affect differentiation of DA neurons but Ezh2 itself was required for the proliferation of cells in the developing midbrain.
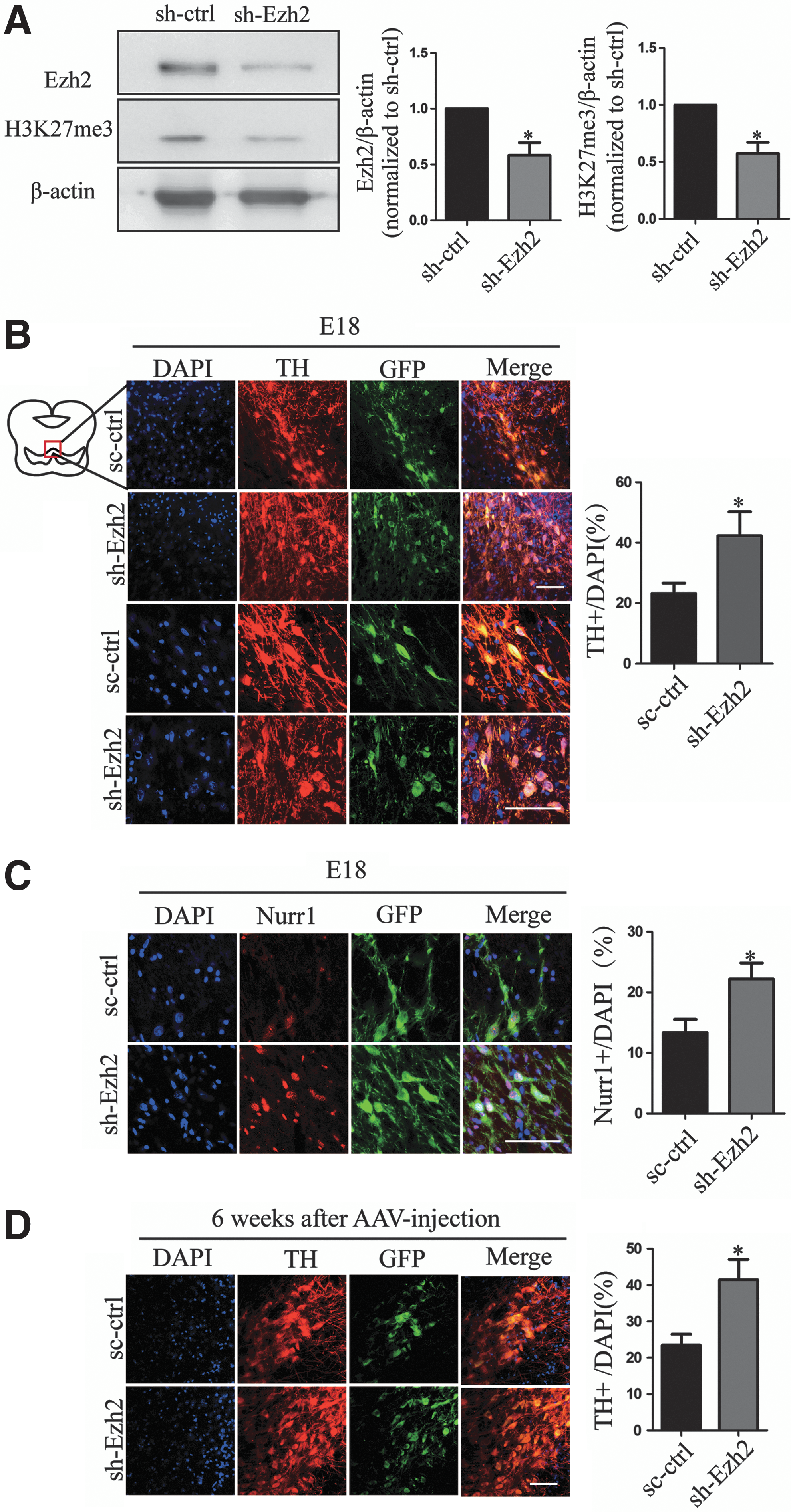
Deletion of Ezh2 promotes differentiation of DA neurons in midbrain.

Overexpression of Ezh2 does not affect differentiation of DA neurons but promotes the proliferation ability of cells in midbrain.
Discussion
The development of DA neurons requires a precise transcriptional cascade at a specific time point [3,35]. In our study, we proved that the decline of Ezh2 reduced H3K27me3 during midbrain development. It seems H3K27me3 dynamically changes in the midbrain and upregulated H3K27me3 is detrimental to the development and maintenance of DA neurons. Ezh2 is present in an overexpressed form in multitumors and it proves to be a promising target for diseases [36]. It has been proved that Ezh2 regulates the self-renewal of stem cells by modifying cyclin-dependent kinase (CDK) [37] and expression of tumor suppressors [34,38]. In nonsmall cell lung cancer, overexpressed Ezh2 can repress p57 expression by trimethylating H3K27 on CDKN1C [39]. As a CDK inhibitor, p57 blocks the G1/S phase in the cell cycle and represses the proliferation progress [40,41].
Using Ezh2-selective inhibitor and VM-NSCs approaches is an appropriate way to find out the correlation between Ezh2 inhibitor [27] and inhibits its enzyme activity by binding Su(var)3-9, Enhancer-of-zeste, Trithorax (SET) domain competitively [42]. As a novel small molecular, EPZ005687 effectively reduced H3K27me3 in VM-NSC cultures and promoted the differentiation of VM-NSCs into DA neurons. To determine how Ezh2 affects the development of DA neurons, transcription factors can be a cue. Previous studies have proved that high level of H3K27me3 represses Nurr1 expression even in adult rat midbrain [14]. Nurr1 can bind to sequence-specific promoter elements as a monomer, homodimer, or heterodimer with the retinoid X receptor, and regulates the expression of TH and l-aromatic amino acid decarboxylase in immature and mature neurons [43,44]. Our research showed that the capacity of differentiation of VM-NSCs into DA neurons was significantly repressed in the absence of Nurr1, even under EPZ005687 treatment. As we know, Nurr1 alone is not sufficient to induce maturation of DA neurons, and our research did not rule out other transcription factors, such as FoxA2, Pitx3, and Wnt/β-catenin signals [45 –47].
Although we reported here that the requirement for Ezh2 decline in regulating the development of DA neurons is related to its role in the proliferation of stem cells and the expression of gene, it is clear that the role of H3K27me3 and Ezh2 in the development of DA neurons is rather more complex. Previous studies showed that the loss of Ezh2 promoted a midbrain-to-forebrain identity switch by direct gene derepression. These pieces of evidence showed that though the decline of Ezh2 has an important role in the development of DA neurons, Ezh2 is still required for the development of midbrain.
Conclusion
To conclude, we found that the decline of Ezh2 reduced H3K27me3 in the developing midbrain. The deletion of Ezh2 promoted differentiation of DA neurons and Ezh2 overexpression enhanced cell self-renewal in midbrain development. Inhibition of Ezh2 inhibited the proliferation of VM-NSCs and facilitated the expression of DA neurons developmental regulators, and upregulated Nurr1 played a central role in Ezh2 inhibition-mediated differentiation of DA neurons.
Footnotes
Acknowledgments
This study was supported by grants from “Key New Drug Creation and Manufacturing Program” of the National Science and Technology Major Project (grant no. 2014ZX09102-001-05) and the Scientific Innovation Project of the Chinese Academy of Sciences (grant no. XDA01040304).
Author Disclosure Statement
No competing financial interests exist.