
Abstract
Smooth muscle cells (SMCs) are important cell type for regenerative medicine. Previous studies showed that retinoic acid (RA) induces differentiation of SMCs from monolayer-cultured embryonic stem cells (ESCs) with high efficiency. However, the underlying mechanisms are still poorly defined. Here, we identified Wnt signaling as a primary regulator for RA-induced ESC differentiation. The activation of Wnt signaling inhibited the epithelial–mesenchymal transition during ESC differentiation, leading to inhibition of RA-induced SMC differentiation and promoting differentiation of ESCs toward primitive endoderm (PrE) lineage instead, while the inhibition of Wnt signaling promoted RA-induced SMC differentiation. Loss-of-function studies revealed that 7-like 2 (Tcf7l2) was the key transcription factor that Wnt operate through during RA-induced differentiation. Thus, this study revealed that the Tcf7l2-mediated Wnt signaling is a switch in determining the mesoderm/PrE fates in RA-induced ESC differentiation.
Introduction
Embryonic stem cells (ESCs) possess the potential of differentiation into all embryonic cell lineages [1], and even extraembryonic lineages such as primitive endoderm (PrE) [2] and trophoblast [3] under certain conditions. Therefore, ESCs are ideal models for studying early embryonic development and are important cell sources for regenerative medicine. Differentiation of ESCs is regulated by complex and coordinated molecular networks of extracellular cues and intracellular gene regulatory networks.
Retinoic acid (RA), the most physiologically active metabolite of vitamin A, is a potent signaling molecule and serves as a critical morphogen in multiple processes during embryonic development [4]. RA deficiency during embryogenesis causes spectra of organ defects [5]. In in vitro differentiation studies, RA has been found to play essential roles in the induction of ESC differentiation into various cell types of ectoderm and mesoderm origins, including neurons, chondrocytes, osteocytes, skeletal muscle, smooth muscle, and cardiomyocytes [6]. As RA is widely used in various in vitro ESC differentiation protocols presenting dosage- and context-dependent effects [7,8], the study of underlying molecular mechanism and crosstalk with other signaling pathways of RA during ESC differentiation is of great importance to understand cell lineage determination mechanism as well as to improve the efficiency in generating desired cell types.
Several signaling pathways, such as fibroblast growth factor, Hedgehog, and Wnt, have been shown to interact with RA signaling in vitro or in vivo [9]. The Wnt family is a group of cysteine-rich secreted glycoproteins whose signal transduction cascade is highly conserved in eukaryotes and can be divided into β-catenin-dependent canonical signaling and β-catenin-independent noncanonical signaling [10]. Wnt signaling is involved in multiple cellular processes, including proliferation, differentiation, polarization, and migration, as well as ESC self-renewal and differentiation. Several chemical inhibitors are widely used to modulate the activity of Wnt pathway. CHIR99021 can reduce the degradation of β-catenin by selectively inhibiting glycogen synthase kinase 3 (GSK-3), leading to activation of canonical Wnt signaling [11]. XAV939 [12] and PNU74654 [13] can antagonize Wnt signaling by promoting β-catenin degradation and inhibiting the interaction between β-catenin and Tcf7l2, respectively.
Extensive work has been conducted to investigate the crosstalk between RA and Wnt signaling in different contexts [14], but a consensus understanding has not been reached. Interaction between RA and Wnt signaling varies in different developmental and ESC differentiation schemes. For example, Wnt signaling is suppressed by RA during neural tube formation and neural differentiation, while activation of Wnt signaling blocks the neural inducing effect of RA [15]. In contrast, RA signaling increases Wnt activity in PrE differentiation [16].
We have previously established a highly efficient RA-induced ESC-smooth muscle cell (SMC) differentiation model [17]. By applying a high dose of RA (10 μM) to monolayer cultures of ESCs, we achieved up to 90% differentiation of SMCs. However, the underlying mechanisms, especially the potential crosstalk with other signaling pathways, are not well understood. Here, we identified Wnt as an important signaling pathway that interacted with RA signaling during SMC differentiation process and crucially affected the lineage identity of high-dose RA-induced ESCs. By blocking epithelial–mesenchymal transition (EMT), Wnt signaling activation attenuated the RA-induced differentiation to SMCs and promoted the differentiation toward PrE lineage. Importantly, Wnt inhibition further promoted the RA-induced SMC differentiation from ESCs. Moreover, we identified Tcf7l2 as the critical downstream effector of Wnt signaling in the context of RA-induced ESC differentiation. Our study thus provided insights into the importance of RA/Wnt crosstalk in the lineage differentiation of ESCs, which may facilitate the development of efficient methods for differentiation of ESCs into SMCs.
Experimental Procedures
Cell culture and differentiation
Undifferentiated D3 mESCs were cultured on mitomycin-inactivated mouse embryonic fibroblasts (MEFs) in DMEM (Gibco) supplemented with 15% fetal bovine serum (Gibco) and 1,000 U/mL LIF (Millipore) at 37°C in 5% CO2. ESCs were resuspended and plated onto gelatin-coated dishes at a density of 4 × 104/cm2 in DMEM with 15% serum and 10 μM atRA for 10 days, then DMEM with 10% serum for 6 days, to perform monolayer differentiation. The effects of PD0325901, LY294002, CHIR99021, XAV939, and PNU74654 (Tocris) added from day 0–10 on the RA-induced differentiation were examined.
Western blot analysis
Cells were harvested and lysed on ice for 1 h in RIPA buffer (Beyotime) containing protease inhibitor cocktail (Roche). Protein concentrations were determined using BCA protein assay kit (Thermo Scientific). Equal amounts of each protein lysate were loaded onto each lane of a 12% or 6% SDS-polyacrylamide gel and separated by electrophoresis. The separated proteins were then transferred onto polyvinylidene fluoride (PVDF) membranes (Millipore). After blocked with the blocking solution (5% nonfat milk and 0.1% Tween 20 in TBS) for 1 h, membranes were incubated with primary antibodies diluted in blocking solution at 4°C overnight, then horseradish peroxidase (HRP)-conjugated secondary antibodies were added. Immunoreactive protein bands were visualized using Clarity™ ECL Western Blotting Substrate (Bio-Rad), and signal detection was performed on ChemiDoc XRS+ System (Bio-Rad). Densitometric analysis was carried out using Image Lab Software (Bio-Rad), and values were normalized to β-actin or Histone H3. The primary and secondary antibodies used are listed in Supplementary Table S2.
Quantitative RT-polymerase chain reaction
Total RNA was extracted with TRIzol reagent (Takara) according to the manufacturer's instructions, and cDNA was reverse transcribed with the ReverTra Ace qPCR RT Kit (Toyobo) from 1 μg RNA. cDNA samples were subjected to quantitative polymerase chain reaction (qPCR) amplification on a LightCycler 480 (Roche) using THUNDERBIRD SYBR qPCR Mix (Toyobo) according to the manufacturer's guidelines. All reactions were performed in triplicate and normalized to β-actin (Actb), which served as a reference gene in the same sample. Primer sequences used were acquired from PrimerBank (
Immunofluorescence analysis
Cells were plated on poly-L-lysine-treated slides and fixed with 4% paraformaldehyde in PBS for 30 min at room temperature (RT), and then permeabilized with 0.25% Triton-X 100 (Sigma) in PBS for 20 min. Samples were incubated in blocking solution (10% normal goat serum and 0.25% Triton-X 100 in PBS) for 1 h at RT and stained with primary antibodies diluted in blocking solution overnight at 4°C. After washing thrice with PBST (0.1% Triton-X 100 in PBS), samples were incubated with secondary antibodies in blocking solution for 1 h at RT. Stained samples were mounted using fluoroshield mounting medium with DAPI (Abcam). The images were acquired with an LSM710 confocal laser scanning microscope (Zeiss). The primary and secondary antibodies used are listed in Supplementary Table S2.
Flow cytometry analysis
Cells were dissociated into single-cell suspensions with 0.25% trypsin-EDTA (Gibco), then fixed and permeabilized using Foxp3/Transcription Factor Staining Buffer Set (eBioscience) according to the manufacturer's instructions. After blocking with 2% normal goat serum in PBS (blocking solution) for 15 min, cells were incubated with primary antibody diluted in blocking solution for 1 h, washed with PBS twice, and then incubated with fluorescein-conjugated secondary antibody in blocking solution for 30 min. Finally, the stained cells were resuspended in PBS and analyzed using BD Accuri C6 flow cytometer, and data were analyzed with BD Accuri C6 Software or FlowJo software. Cells stained with the corresponding isotype antibodies were used as negative control to set gates. Antibodies used are listed in Supplementary Table S2
RNA-Seq
Total RNA was extracted and converted to cDNA libraries using the TruSeq RNA Sample Prep Kit (Illumina). The libraries were sequenced to a depth of ∼25 M paired-end reads using an Illumina HiSeq 2000 system. See Supplemental Experimental Procedures for details. The accession number is GSE123632.
Gene targeting
The sgRNAs against early exons of the target genes were designed using the CRISPR Design Tool (
Statistical analyses
All experiments were performed at least in technical triplicate. All data are presented as mean ± SEM. Statistical significance was calculated using Student's t test for pairwise comparisons. Significant differences were defined as *P ≤ 0.05, **P ≤ 0.01.
Results
Activation of Wnt signaling inhibits RA-induced ESC–SMC differentiation
To identify the potential signal pathways that modulate RA-induced SMC differentiation, we chemically modulated several major signaling pathways, including ERK [PD 0325901 (PD)-inhibition], PI3K-Akt [LY 294002 (LY)-inhibition], and Wnt [CHIR 99021 (CHIR)-activation] during RA-induced ESC–SMC differentiation, and evaluated the differentiation outcome. While PD treatment increased the expression of SMC markers (Acta2, Tagln, Cnn1, and Myh11), both LY and CHIR treatments decreased the expression of some of these markers. Western blotting results showed that the levels of nonphospho (active) β-catenin were significantly increased after the addition of CHIR, indicating that CHIR had activated Wnt signaling pathway (Supplementary Fig. S1A). CHIR treatment resulted in strong morphological changes (Supplementary Fig. S1B) and consistent decrease in SMC markers at both mRNA and protein levels (Supplementary Fig. S1C, D); therefore, we focused on Wnt signaling in this study.
To further characterize the effect of Wnt signaling on RA-induced SMC differentiation, we conducted detailed analysis with different dosages of chemical activator (CHIR) and inhibitors [XAV939 (XAV) and PNU74654 (PNU)]. Real-time RT-PCR analysis showed that CHIR significantly downregulated the expression of SMC markers, including Acta2, Tagln, Cnn1, and Myh11, and transcription factor Myocd (Fig. 1A top). The protein levels of Acta2 and Myh11 were significantly decreased (Fig. 1B). On the contrary, the expressions of all these SMC markers were augmented by the addition of Wnt inhibitor XAV or PNU when compared with standard differentiation (Fig. 1A bottom and C). In addition, immunofluorescence and flow cytometry analyses showed that the addition of CHIR significantly lowered the ratio of Acta2- and Myh11-positive cells (Fig. 1D, E); the addition of XAV or PNU, however, increased the ratio above the RA alone induced differentiation (Fig. 1D, F). Taken together, these data suggested that Wnt signaling represses RA-induced ESC–SMC differentiation.
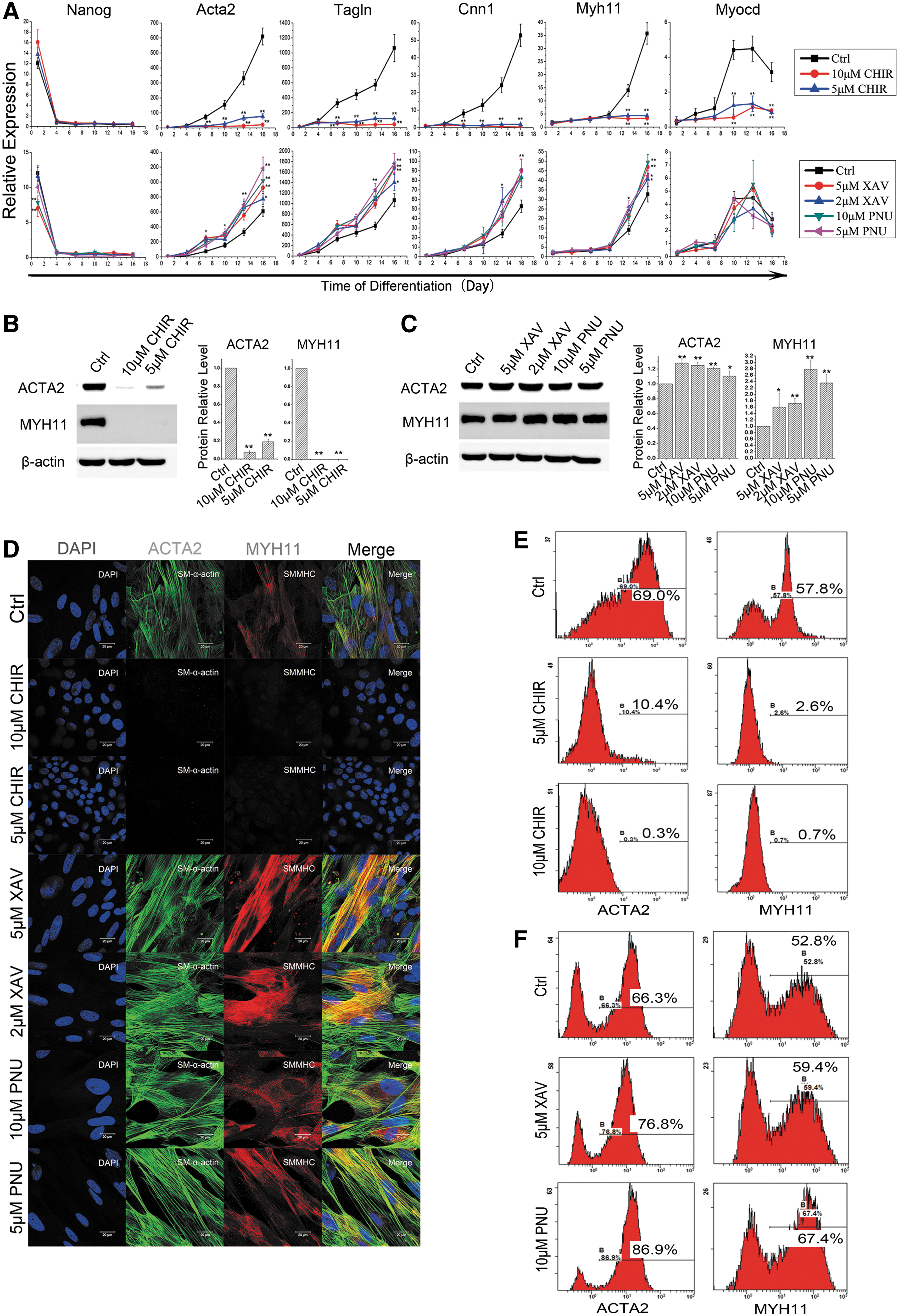
Activation of Wnt signaling inhibits SMC formation in RA-induced ESC differentiation.
Activation of Wnt signaling promotes RA-induced ESC differentiation toward PrE lineage
To uncover the mechanism that Wnt modulates RA-induced ESC differentiation, we conducted time-course RNA-seq analysis during the differentiation (Supplementary Fig. S2A). Interestingly, the RNA-seq and subsequent real-time RT-PCR data revealed a significant decrease in mesoderm (one of the major sources of vascular SMCs) markers such as Brachyury (T), Kdr, and Isl1, and increase in PrE markers such as Gata6, Pdgfra, and Dab2 in CHIR-treated samples (Fig. 2A), while other lineage markers showed no significant expression (Fig. 2B), implying a diverging to PrE fate by Wnt activation. Following this lead, we included ESC and a PrE-derived Xen cell line in RNA-seq analysis and performed hierarchical clustering analysis. As shown in Fig. 2C, the gene expression profiles of cells derived from RA induction alone clustered with adult SMCs, while cells derived from RA/CHIR induction gradually clustered to XEN cells with differentiation and were very close to XEN cells at day 16, suggesting that RA/CHIR-induced ESCs differentiated into PrE lineage. Moreover, expression profiles of cells derived from the RA induction at day 4 were clustered with cells from the RA/CHIR induction, indicating that the lineage identity of ESCs had not been diverted at day 4.

Activation of canonical Wnt signaling promotes RA-induced ESC differentiation toward PrE lineage.
We then assessed specific markers representative of key cell states: pluripotency, SMC, PrE, and PrE derivatives, which comprise parietal endoderm and visceral endoderm in the RNA-seq data (Fig. 2D). The pluripotency markers that were highly expressed in ESCs were rapidly downregulated with RA only induction, while some of these markers such as Esrrb, Zfp42, Oct4 (Pou5f1), and Klf4 continued to express at low level under RA/CHIR induction, consistent with the presence of these markers at early stages of PrE differentiation [20]. SMC markers and related transcription factors expressed at high levels specifically in RA-induced ESC differentiation and adult SMCs, while PrE markers had high expression in RA/CHIR-induced differentiation, especially at later period and in the positive control XEN, demonstrating that RA and RA/CHIR induced ESCs toward SMC and PrE lineage, respectively. Many of these results were further corroborated by immunofluorescence, western, and flow cytometry analysis (Fig. 2E–G).
Next, we performed gene set enrichment analysis (GSEA) of the Gene ontology (GO) and KEGG (Kyoto encyclopedia of genes and genomes) pathways. Upregulated genes in RA-induced differentiation were associated with focal adhesion, ECM–receptor interaction, vascular development, vascular smooth muscle contraction, and cytokine–cytokine receptor interaction, which was consistent with the differentiation of SMCs (Supplementary Fig. S2B) [21,22]. While RA/CHIR-induced differentiation was enriched for GO terms associated with lysosome, glycoprotein metabolism, glycosphingolipid metabolism, and cytokine–cytokine receptor interaction, consistent with the PrE identity (Supplementary Fig. S2C) [23,24].
As an inhibitor of GSK3, CHIR can have multiple effects, including the activation of canonical Wnt signaling by stabilizing β-catenin, as well as on insulin and Hedgehog pathways [25]. To further discern the regulatory mechanisms of CHIR on RA-induced ESC differentiation, we replaced CHIR with Wnt3a to activate canonical Wnt signaling at ligand level. RA/Wnt3a induction gave rise to the polygonal or cobble-shaped cells positive with PrE markers and showed gene expression patterns similar to those of RA/CHIR induction (Supplementary Fig. S3A–C), thus corroborating that the function of CHIR on RA-induced ESC differentiation was achieved through activation of Wnt signaling.
Noteworthily, addition of Wnt signaling activator (CHIR) or inhibitor (XAV or PNU) alone in the absence of RA demonstrated little influence on expression of either SMC or PrE phenotype, suggesting that lineage determination of ESC differentiation requires synergistic regulation of RA and Wnt signal pathways (Supplementary Fig. S3D).
Activation of Wnt signaling suppresses RA-induced ESC differentiation by diverging to PrE fate
To further corroborate the finding that Wnt activation suppressed RA-induced SMC differentiation of ESCs by diverging the differentiation to PrE lineage, we targeted disrupted Gata6, the earliest lineage specifier of PrE and previously shown to be essential for ESCs differentiation into PrE lineage [26] (Supplementary Fig. S4A–C), in ESCs by CRISPR technology and isolated homozygote clones (Supplementary Fig. S4F–J), and conducted differentiation experiments. When Gata6-null ESCs were set to differentiate in the presence of RA, they showed accelerated and more efficient differentiation into SMC cells than wild-type cells (Fig. 3A left), with stronger expression of SMC marker genes (Fig. 3B), so were the protein levels of Acta2 and Myh11, and the ratio of Acta2-positive cells (Fig. 3C top, D left and E). When CHIR was added to the RA-induced differentiation, the Gata6-null ESCs expressed none of PrE markers, and highly expressed SMC markers with the ratio of Acta2-positive cells reaching up 72% (Fig. 3B, C bottom, D right and E). In addition, inhibiting Wnt signaling by PNU during RA-induced differentiation into Gata6-null ESCs no longer promoted the expression of SMC markers (Supplementary Fig. S4K and Fig. 3E).

Activation of Wnt signaling suppresses RA-induced ESC differentiation by diverging to PrE fate.
These data demonstrated that RA can efficiently induce SMC differentiation of ESCs even under Wnt activation when the PrE differentiation route was blocked by Gata6 knockout, and the SMC differentiation promoting effects of Wnt inhibition were indeed mediated through suppressing PrE fate, therefore corroborating the idea that Wnt activation suppressed RA-induced SMC differentiation of ESCs by diverging the differentiation to PrE lineage, indicating that RA-induced ESC differentiation toward SMC or PrE lineage is an either–or process, and the determination between SMC versus PrE is fine-tuned by Wnt signaling.
Wnt signaling regulates ESC lineage differentiation by blocking EMT
EMT is an important process during ESC (epithelial) to SMC (mesenchymal) differentiation. Moreover, PrE lineage possesses epithelial characteristics [27]. Therefore, we explored the role of Wnt signaling in regulating EMT during RA-induced differentiation. To this end, we examined the expression of a series of EMT regulators during RA- or RA/CHIR-induced ESC differentiation. GSEA showed that GO terms associated with EMT and mesenchymal cells significantly enriched in RA-induced differentiation, while in the differentiation synergistically induced by RA/CHIR, the degrees of enrichment of these terms were significantly lower (Fig. 4A). These results demonstrated that RA-induced ESC–SMC differentiation required the EMT process, which can be blocked by CHIR, suggesting that the activation of Wnt signaling changes the lineage determination of RA-induced ESC by blocking EMT.
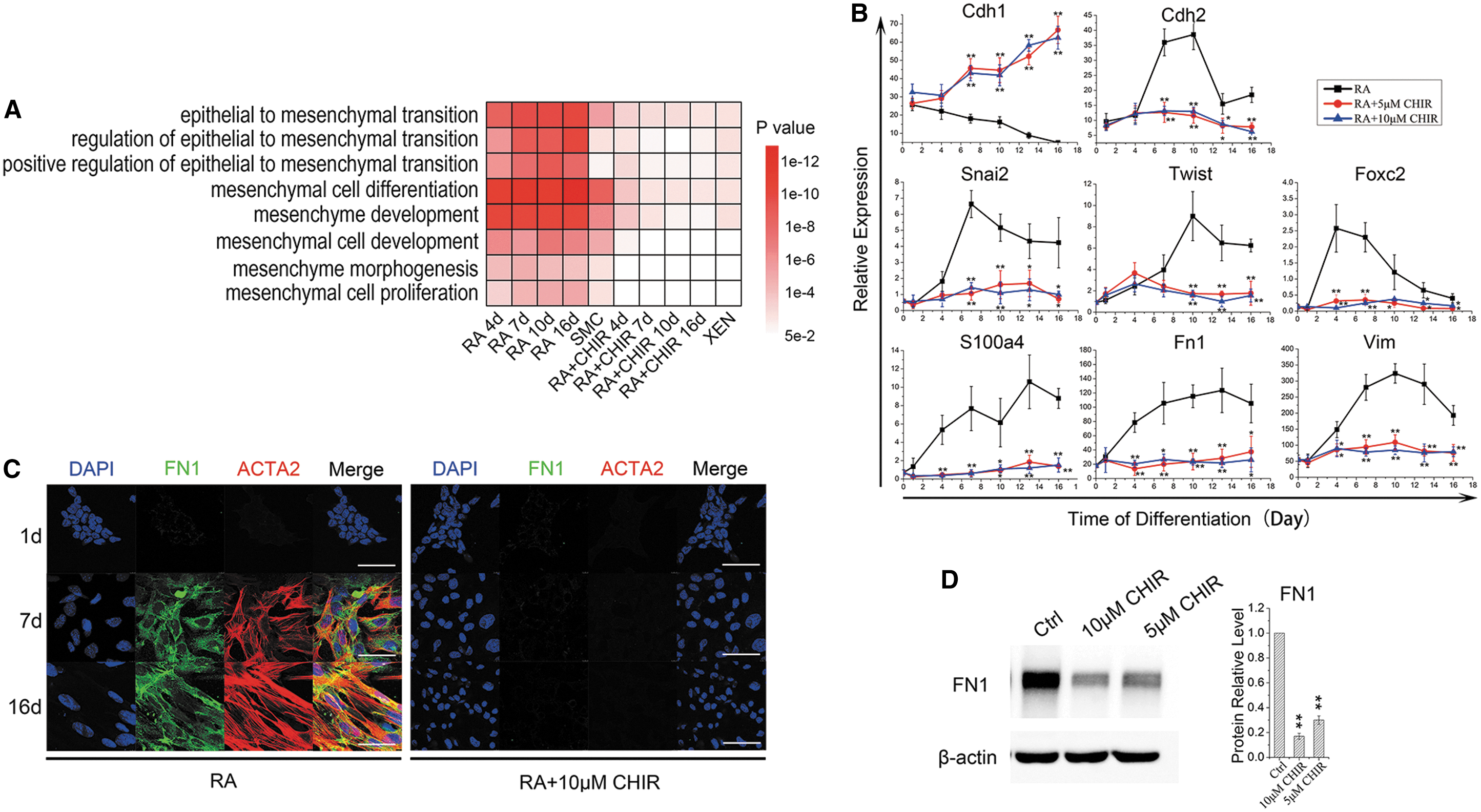
Wnt signaling regulates ESC lineage differentiation by blocking EMT.
We further detected the intermediate stages of RA-induced differentiation from ESCs into SMCs, and found that E-cadherin (Cdh1) was significantly downregulated, while the expression of N-cadherin (Cdh2) increased and peaked at day 10 (Fig. 4B). Time-course analysis demonstrated that transcription factors regulating EMT (Snail2, Twist, Foxc2) and EMT-related structural proteins (S100a4, Fn1, Vim) were significantly upregulated in ESC–SMC differentiation, while upon treatment with CHIR, these markers were barely expressed, corroborating that RA-induced ESC–SMC differentiation required EMT while CHIR blocked it (Fig. 4B–D). Again, replacing CHIR with Wnt3a recapitulated the inhibitory effects on EMT as CHIR did (Supplementary Fig. S3C).
Interestingly, Gata6 knockout ameliorated the EMT blocking effect by Wnt activation, shown by the recovery of the expression of EMT-related genes, including Cdh1, Fn1, and Snai2 (Fig. 3B), suggesting a potential role of Gata6 in the EMT suppressing function of Wnt in RA-induced ESC differentiation.
Wnt signaling regulates SMC/PrE lineage decision by Tcf7l2
Canonical Wnt signaling is transduced by a group of Lef/Tcf family transcription factors in a context-dependent manner. To further understand the molecule regulation of Wnt on RA-induced ESC differentiation, we set out to identify the particular Lef/Tcf that functions in this context. To this end, we first profiled the expression of all the Lef/Tcf factors, including Lef1, Tcf1 (official symbol Tcf7), Tcf3 (Tcf7l1), and Tcf4 (Tcf7l2) [28] throughout the process of differentiation. Among these, only Tcf7l2 was significantly upregulated or downregulated in response to the activation or inhibition of Wnt signaling, respectively (Supplementary Fig. S4D). In addition, we detected the expression and distribution of β-catenin and TCF7L2 during differentiation by immunocytochemistry, and found that β-catenin and TCF7L2 expression was upregulated and colocated in the nucleus under RA+CHIR treatment (Fig. 5A). Interestingly, as PrE markers, Snai1, Sox17, Stra6, and Vegfa are also recognized targets of Wnt signaling pathway [29], qRT-PCR results showed that the expression of these genes was significantly upregulated in the differentiation process of ESC-PrE induced by RA+CHIR (Supplementary Fig. S4E). These results suggested that Tcf7l2 might play a key role in relaying Wnt signal in RA-induced ESC differentiation.
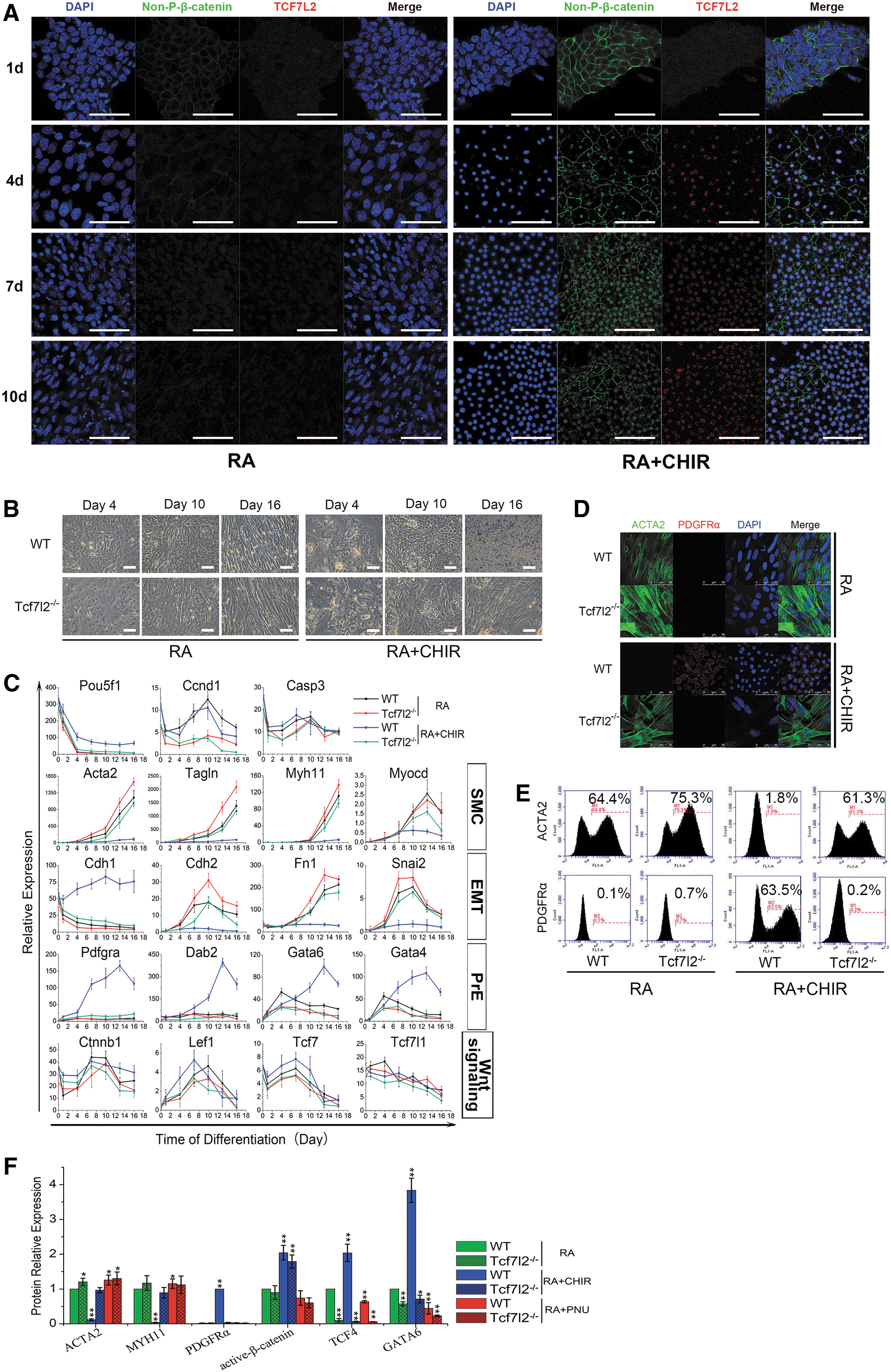
Wnt signaling regulates SMC/PrE lineage decision by Tcf712.
To demonstrate it, we knocked out Tcf7l2 using CRISPR-Cas9 method (Supplementary Fig. S4F–J). In RA-induced ESC–SMC differentiation, Tcf7l2-null ESCs expressed higher levels of SMC markers and EMT-related genes compared with wild-type line, and same was the ratio of Acta2-positive cells (Fig. 5B left, C, D top, E left and F). When Wnt signaling was activated with CHIR, instead of upregulating PrE markers, Tcf7l2-null ESCs activated SMC and EMT markers like wild-type ESCs under RA only induction, and the proportion of Acta2-positive SMC cells were also recovered (Fig. 5B right, C, D bottom, E right and F). These results suggested that deletion of Tcf7l2 abolished Wnt signaling regulation of ESC differentiation toward PrE lineage and promoted differentiation toward SMC. In addition, PNU treatment no longer promoted SMC differentiation into Tcf7l2-null ESCs (Supplementary Fig. S4K and Fig. 5F), corroborating that Wnt activation acted through Tcf7l2 to activate PrE differentiation and therefore suppressing SMC differentiation. The above results showed that the role of SMC/PrE lineage determination by Wnt signaling in RA-induced ESC differentiation is achieved through Tcf7l2 pathway.
Discussion
The lineage identity of a particular cell type is defined by balanced acts of different signaling pathways and transcription factors. Enormous evidence from different differentiation models in the past decade has revealed that various signaling pathways and molecules orchestrate the process of SMC differentiation from pluripotent stem cells [30]. Wnt signaling participates in the regulation of multiple cellular processes, while its role in regulating ESC self-renewal and differentiation is complicated and somewhat controversial. Several studies have revealed that Wnt signaling plays a role in maintaining pluripotent state [31], whereas more contradictory results show that its activation can lead to ESC differentiation [32,33]. However, little is known about the functional roles of Wnt signaling in SMC differentiation from ESCs. Here using a simplified and efficient in vitro differentiation system, we identified Wnt signaling as a switch that controls the lineage balance between SMC and PrE lineage during RA-induced differentiation, advancing our knowledge of the molecular mechanism regulating SMC differentiation.
Diversity of embryonic origins is a well-recognized hallmark of SMCs, and may influence the development and the site-specific localization of vascular disease [34]. Despite great progress in SMC differentiation in vitro models so far, most systems are focusing more on the efficiency of differentiation, and in most cases it is unclear which embryonic lineage the resulting cells represent for [35]. In this study, lineage identification through qRT-PCR and RNA-seq analyses showed that the SMCs obtained through RA-induced ESC differentiation contained a significant population originated from mesoderm, especially the lateral plate mesoderm, which ultimately differentiates into aortic root and coronary artery in vivo [34]. The observation that the process of RA-induced ESC–SMC differentiation going through EMT and an intermediate stage of mesenchyme found in this study further indicates the mesoderm origin of the derived SMCs.
Based on the observations in this study that activation of Wnt signaling diverged RA-induced ESC differentiation to PrE fate, we have shown that both blocking PrE differentiation by Gata6 knockout and inhibiting Wnt signaling by chemical inhibitor or Tcf7l2 knockout would greatly enhance the efficiency of SMC differentiation, while on the contrary, activation of Wnt signaling successfully induced ESCs into PrE lineage when high concentration of RA was presented. Interestingly, while previous studies showed that Gata6 could also participate in the transcription regulation of SMC markers [36], but Gata6 is not essential for RA-induced ESC differentiation toward SMC.
In summary, these findings demonstrated that canonical Wnt signaling was a key decision signaling for the differentiation of ESCs into mesoderm lineage and then into SMC or direct differentiation into PrE lineage under RA induction. Furthermore, we found that Tcf712 was the key mediating transcription factor for this regulatory effect of Wnt signaling. This may offer new insights into the pathways regulating the determination of different lineages within the embryo. Besides, this study provided new efficient means to generate functional SMC and PrE lineage in vitro, and methodological references for the future therapeutic applications in regenerative medicine.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was funded in part by Natural Science Foundation of China (31071201) to M.Z.; Initiative Design Project of Agricultural Research of Hangzhou (20162012A07) to H.H.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Table S1
Supplementary Table S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.