
Abstract
Mesenchymal stromal cells (MSCs) promote wound healing by expediting the inflammatory phase. Local injection of MSCs into injured vocal folds (VFs) is effective in animal models, suggesting suitability for clinical translation. Despite their therapeutic potential, MSCs do not persist within the VF. This study evaluates whether hyaluronan (HA) hydrogels offer a safe delivery vehicle for local injection of MSCs into VFs, and increase longevity of the cells within the injured tissue. MSCs ± HA hydrogel were exposed to interleukin (IL)1β, IL8, and chemokine (C-C motif) ligand 4, and evaluated for mRNA expression of matrix remodeling genes and secretion of immunomodulatory/prohealing factors. Chemotaxis/invasion in response to inflammation was evaluated. A lapin model of VF injury evaluated in vivo effects of MSCs ± HA hydrogel on enhancing VF healing. Histological evaluation of inflammation, type I collagen expression, HA hydrogel resorption, and MSC persistence was evaluated at 3 and 25 days after injury. MSCs within HA hydrogel were responsive to their extracellular environment, upregulating immunomodulatory factors when exposed to inflammation. Despite delayed migration out of the gel in vitro, the MSCs did not persist longer within the injured tissue in vivo. MSCs ± HA hydrogel exerted equivalent dampening of inflammation in vivo. The gel was resorbed within 25 days and no edema was evident. HA hydrogels can be safely used in the delivery of MSCs to injured VFs, minimizing leakage of administered cells. MSCs within the HA hydrogel did not persist longer than those in suspension, but did exert comparable therapeutic effects.
Introduction
Vocal fold (VF) scarring is a major cause of severe voice problems, characterized by increased levels of disorganized collagen and decreased elastin within the lamina propria (LP) [1 –3]. This results in a strained or aphonic voice, where the vibratory capacity of the VF is limited or absent [1]. At present no single treatment modality has been proven to be effective for VF scarring [4].
Hyaluronan (HA), is a glycosaminoglycan found throughout the VFs and essential to its biomechanical properties and oscillations during phonation. In addition to its role in conveying the functionality of the VF, HA is involved in the acute phase of wound healing, with anti-inflammatory properties in its high-molecular-weight form [5]. Its abundance in fetal wounds, which heal without scar formation, suggests a further role in tissue regeneration [6]. HA has been utilized extensively in in vitro, preclinical, and clinical applications. In hydrogel form, HA has been used as a drug delivery vehicle and in the controlled release of prohealing factors, such as interleukin (IL)-10, transforming growth factor beta 1 (TGFβ1), basic fibroblast growth factor, and most recently secreted factors from the mesenchymal stromal cell (MSC) secretome [7 –10]. Within the wound healing field, HA represents a nonimmunogenic, biocompatible material for the delivery of cells, including MSCs, and as a component in dressings to promote endogenous healing responses [11 –14].
HA's safety profile has been demonstrated in multiple clinical applications, including in the treatment of osteoarthritis, ethmoidectomy surgery, and glottal insufficiency, where for over 15 years, HA has been utilized for temporary augmentation in VF paresis, atrophy, and scarring [15 –20]. Endogenous tissue HA levels are reduced with injury to the VF, and alongside its biocompatibility and resorption, 1–12 months after delivery (dependent on the application), HA offers great advantages for clinical application in VF repair [16,21,22].
MSCs have been utilized extensively within the clinic for their immunomodulatory and prohealing properties [23 –26]. MSCs are plastic in their mechanism of action, exerting differential effects through cell/cell contact, and their secretome/extracellular vesicles, dependent on the extracellular milieu that they are exposed to [27]. We have previously demonstrated the ability of MSCs to promote wound healing within the injured VF [28 –30]. Our work has demonstrated that through modulating the phenotype of invading peripheral monocytes and tissue-resident macrophages, MSCs can promote an anti-inflammatory reaction, resolving the acute wound healing phase and promoting tissue repair and regeneration [28]. Interestingly, MSCs exert these long-term effects despite low-level persistence within the VF into which they are administered [28 –30].
The aim of this study was to evaluate the safety of HA hydrogels as a delivery vehicle for the administration of MSCs into injured VFs. Furthermore, we aimed to establish whether HA hydrogel could increase longevity of the MSCs within the VF tissue, potentially enhancing their prohealing effects.
Materials and Methods
Isolation and culture of human MSCs
Human MSCs were isolated from the iliac crest of healthy donors as previously described (n = 4) [24]. Ethical approval was obtained from the Local Ethics Committee, Stockholm, and all donors provided written consent. Expansion and characterization of MSCs were performed according to guidelines of the European Blood and Marrow Transplantation Group and approved by the Swedish National Board of Health and Welfare. Flow cytometry confirmed a MSC surface profile as per the International Society for Cellular Therapy guidelines [positive for CD73, CD90, CD105, and human leucocyte antigen (HLA)-I and negative for CD14, CD34, CD45, and HLA-II] [31].
MSCs were maintained in DMEM supplemented with 10% fetal calf serum (FCS), 100 IU/mL penicillin, 0.1 mg/mL streptomycin, and 0.25 μg/mL Fungizone
Incorporation of MSCs into a HA hydrogel
Auxigel™ (Termira AB, Stockholm, Sweden) was prepared by mixing 0.9% (w/v) HA in phosphate-buffered saline (PBS; part A) with 0.1% (w/v) polyvinyl alcohol derivative in PBS (to induce crosslinking, part B) at a 3:1 ratio. The HA hydrogel was mixed with an MSC cell suspension in culture media (in vitro studies) or PBS (in vivo studies) before utilization. Cell concentration was experiment dependent.
Assessment of MSC cell number in HA hydrogel
The effect of HA gel incorporation on MSC viability and proliferation was assessed using the CyQUANT® Cell Proliferation Assay (Thermo Fisher). MSC cell suspensions (8.125E+05 cells/mL; n = 4; P3) were combined with the HA gel components as described. The gel/MSC mix was syringed through a 27G needle, to exert equivalent shear stress to that utilized in delivering the cells in vivo, into a 96-well microplate (100 μL/well). The gels were solidified before adding DMEM supplemented with 20% FCS and 100 IU/mL penicillin, 0.1 mg/mL streptomycin and 0.25 μg/mL Fungizone. Cells were harvested immediately after gel solidification on day 0, day 1, and day 3. Supernatant was harvested and the gels snap frozen on dry ice and stored at −80°C. Plates were thawed at room temperature before adding CyQUANT GR dye diluted in the provided lysis buffer as per the manufacturer's instructions. The plate was incubated for 5 min at room temperature before reading on a fluorescence microplate reader with filters for 480 nm excitation and 520 nm emission (VarioSkan® Flash; Thermo Fisher). Data are expressed as percentage change in cell number relative to the day 0 control.
Evaluating MSC migration and invasion in HA hydrogel
The capacity of MSCs to migrate out of the HA gel and respond to proinflammatory cytokines was assessed using the Cultrex® 96-well Collagen I Cell Invasion Assay (R&D Systems, Abingdon, United Kingdom). MSCs (n = 4; P4) were serum starved 24 h before commencing the experiment by incubating in DMEM supplemented with 0.5% FCS and 100 IU/mL penicillin, 0.1 mg/mL streptomycin, and 0.25 μg/mL Fungizone (herein termed low-serum media). Detached MSCs were incorporated within the HA gel, as previously described, at a concentration of 1E+06 cells/mL in low-serum media. Cell suspension in HA or media (50 μL/well, equivalent to 5E+04 cells) was added to the top of the Cultrex chambers (±coating with type I collagen). Gels were allowed to solidify before the proinflammatory factors IL1β, IL8, and chemokine (C-C motif) ligand 4 (CCL4; PeproTech Nordic, Stockholm, Sweden) were diluted to a working concentration of 10 ng/mL in low-serum media and added to the bottom of the plate. The plate was incubated for 24 h at 37°C/5% CO2. Media, gel, and nonmigrating/invading cells in the top chamber were discarded, before the membrane was washed with provided buffer and incubated with calcein solution diluted in dissociation solution as per the manufacturer's protocol. Cell number was quantified using a fluorescence microplate reader with filters for 480 nm excitation and 520 nm emission (Infinite F200®PRO; Tecan Group Ltd., Männedorf, Switzerland).
Transcriptional regulation of matrix remodeling genes by MSCs incorporated in HA hydrogel
MSCs were assessed for the expression of hyaluronidase (HYAL), hyaluronan synthase (HAS), and matrix metalloproteinases (MMPs) by quantitative polymerase chain reaction (QPCR). MSCs (n = 4; P6; 1E+05 cells/well) were combined with the HA gel as previously described. MSCs seeded in serum-free culture media supplemented with 100 IU/mL penicillin, 0.1 mg/mL streptomycin, and 0.25 μg/mL Fungizone served as controls. IL1β, IL8, or CCL4 were added to the cells/gels at a working concentration of 10 ng/mL. Plates were incubated for 24 h at 37°C/5%CO2. Conditioned media were stored at −80°C for analysis. The gels and adherent cells were placed into RNAlater™ solution (Thermo Fisher) and stored at −20°C before assay.
RNA was isolated using the RNeasy® Mini Kit (QIAGEN AB, Sollentuna, Sweden), eluted in 30 μL of RNase-free water and quantified using a NanoDrop 2000C (Thermo Fisher). cDNA was generated using the High-Capacity cDNA Reverse Transcriptase Kit (Thermo Fisher) and QPCR performed using Fast SYBR® Green Master Mix (Thermo Fisher) on a CFX384 C1000 Touch Real-time system (Bio-Rad Laboratories AB, Solna, Sweden). Primer sequences for each of the investigated genes are outlined in Table 1. Data are expressed as relative expression normalized to the housekeeping gene, β-actin [β-actin cycle threshold (Ct)—gene of interest Ct].
Primer Sequences Used for Quantitative Polymerase Chain Reaction
Effects of HA hydrogel incorporation on the secretion of soluble factors modulating wound healing by MSCs
Conditioned media, harvested from the above experiments, were assessed for levels of IL8, hepatocyte growth factor (HGF), IL6, vascular endothelial growth factor (VEGF), and TGFβ1 by Enzyme-Linked Immunosorbent Assay (ELISA). All assays were performed according to the manufacturer's protocol (R&D Systems). Absorbance was read at 450 nm using a VarioSkan Flash (Thermo Fisher).
Evaluating the in vivo effects on VF wound healing by MSCs incorporated in HA hydrogel
New Zealand white rabbits (3–4 kg; n = 18) were utilized for these experiments. Animal studies were conducted in accordance with the American principles of Laboratory Animal Care and Swedish regulations on animal care ethics conforming to Directive 2010/63/EU.
Induction of VF injury
Rabbits (n = 14) were premedicated, anesthetized, and VF injuries induced as previously described [30]. All the animals survived the procedure. Four animals were not operated on and served as normal VF controls, together with two normal VFs from a data bank.
Treatment with MSCs
MSCs (P5) were resuspended in PBS or incorporated with the HA, as previously described, to a final concentration of 1 × 106/mL. Immediately after VF injury, 1 × 105 MSCs ± HA gel (0.1 mL/VF) were injected into the damaged area of LP and thyroarytenoid muscle using a Medtronic Xomed laryngeal injector with a 27G cannula under video monitoring (n = 5 VFs MSC/PBS mix; n = 9 VFs MSC/HA gel mix). Injured VFs injected with PBS (0.1 mL/VF; n = 3) or HA hydrogel only (0.1 mL/VF; n = 3) served as injured controls.
All animals were monitored for wellbeing and no side effects were noted during the course of the experiment. Animals were sacrificed after 2 days (QPCR; n = 3; see below), 3 days (n = 4 injected with MSC/HA hydrogel), and after 25 days (all remaining animals) with an overdose of intravenous pentobarbital sodium (100 mg/kg body weight). The larynx was dissected from each animal and divided down the posterior midline.
Establishing acute inflammatory responses to VF injury by quantitative PCR
The RiboPure™ RNA Purification Kit (Thermo Fisher) was used to extract total RNA from VF tissue ± injury (n = 2 injured and n = 1 normal/uninjured). cDNA was prepared using the RT2 First-Strand Kit (QIAGEN), and RNA was quantified using a NanoDrop 2000C (Thermo Fisher). The RT2 Profiler PCR array for rabbit inflammatory cytokines and receptors (PANZ011Z; QIAGEN), containing 84 genes, was performed on a CFX384 C1000 Touch Real-time system (Bio-Rad) as per the manufacturer's instructions.
Histological analysis of injured VFs after treatment with MSCs
VFs were fixed in formaldehyde (4% v/v) for 24 h and transferred to 70% ethanol (v/v) before embedding in paraffin wax. Five-micrometer-thick sections were cut in the horizontal plane, deparaffinized in xylene, and rehydrated in decreasing concentrations of ethanol, before staining as detailed below.
Histological analysis of fibrosis and inflammation
Sections, taken from multiple tissue layers, were stained with Mayer's Hematoxylin Plus (Histolab Products AB, Stockholm, Sweden) and 0.2% (w/v) Eosin (Histolab). Slides were dehydrated in increasing concentrations of ethanol, washed in xylene, and mounted using Pertex® mounting medium (Histolab). All sections were digitalized using a 3D HISTECH Midi Scanner System and images viewed using the Pannoramic Viewer Software 1.15 (3DHISTECH Ltd., Budapest, Hungary).
Degree of inflammation and level of fibrosis from VFs, at 3 and 25 days after injury ± MSC treatment, were rated blindly by an experienced pathologist using a previously described scale [30]. Degree of inflammation was based on the presence and position of inflammatory immune cells. A 3-grade scale was used: 1—no/minimal inflammation; 2—moderate inflammation within the LP and/or superficial thyroarytenoid muscle; 3—severe inflammation within the LP and/or superficial and deeper muscle. For fibrosis, four categories were used depending on the grade of scarring [30]. Grade A—no/minimal fibrosis; Grade B—focal or noncompact fibrosis in the LP or superficial vocal muscle; Grade C—compact fibrosis in the LP and superficial muscle; Grade D—compact fibrosis in the LP and superficial and deep vocal muscle.
Immunohistochemistry for type I collagen
Immunohistochemical staining was performed as previously described [30]. Briefly, rehydrated sections were blocked in PBS containing 3% (w/v) bovine serum albumin (BSA; Sigma-Aldrich Sweden AB, Stockholm, Sweden). Slides were incubated in mouse anti-cow type I collagen monoclonal primary antibody (#ab6308; Abcam, Cambridge, United Kingdom) diluted 1:200 in 1% (w/v) BSA overnight at 4°C, before incubation with 5 μg/mL Cy™3 AffiniPure goat anti-mouse IgG secondary antibody (#115-165-146; Jackson ImmunoResearch Laboratories, Inc., PA), diluted in 1% (w/v) BSA for 1 h at room temperature. Sections were washed with PBS before mounting with VECTASHIELD® containing a 4′,6-diamidino-2-phenylindole (DAPI) counterstain (Vector Laboratories, Inc., Burlingame, CA). Slides were visualized using an Olympus IX81 fluorescence microscope coupled to an XC10 camera (Olympus Soft Imaging Solutions GmbH, Muenster, Germany).
Relative type I collagen levels were quantified from the digitized images after a color filtering and normalization process using custom software (Hans Larsson, Karolinska Institutet) as previously described [30]. Relative staining for each VF layer was calculated by normalizing to the whole VF.
Alcian blue staining
Rehydrated sections were incubated with Alcian Blue stain and counterstained with Nuclear Fast Red solution as per the manufacturer's instructions (#SS012; BioGenex, CA). Dehydrated sections were mounted using Pertex mounting medium. Staining was rated blindly by an experienced pathologist using a 3-grade scale: 0—no/little staining; 1—focal increase in staining within the LP and superficial vocal muscle; 2—staining of the LP, superficial, and deep muscle.
Fluorescence in situ hybridization
Fluorescence in situ hybridization was used to verify the persistence of MSCs within the VFs at 3 and 25 days, as previously described [30]. Positive and negative cells were quantified in five fields of view for each VF.
Statistics
In vitro data were assessed for normality using the Shapiro–Wilk test and equal variances using the F-test. Statistical analysis was performed using Student's t-test (parametric) or Mann–Whitney U test (nonparametric). Statistical analyses of the histological and immunohistological data were conducted using nonparametric methods (χ2, χ2 analyses, Wilcoxon and Mann–Whitney tests; StatView® 5.0; SAS Institute, Inc., NC). Statistical significance was assumed at P < 0.05.
Results
MSCs demonstrate delayed migration when incorporated within a HA hydrogel
The migratory ability of MSCs in HA hydrogel or liquid cell suspension was evaluated in response to key proinflammatory factors demonstrated to be upregulated after injury to the VF (Supplementary Table S1). MSCs incorporated within the HA hydrogel demonstrated a reduction in cell migration compared with controls in liquid suspension (Fig. 1b; P < 0.01). The addition of IL1β, IL8, or CCL4 suppressed the migration of MSCs in suspension (Fig. 1b; P < 0.001 IL1β; P < 0.05 IL8 and CCL4). No further reduction in migration was seen in response to proinflammatory cytokines by MSCs incorporated within HA (Fig. 1b).
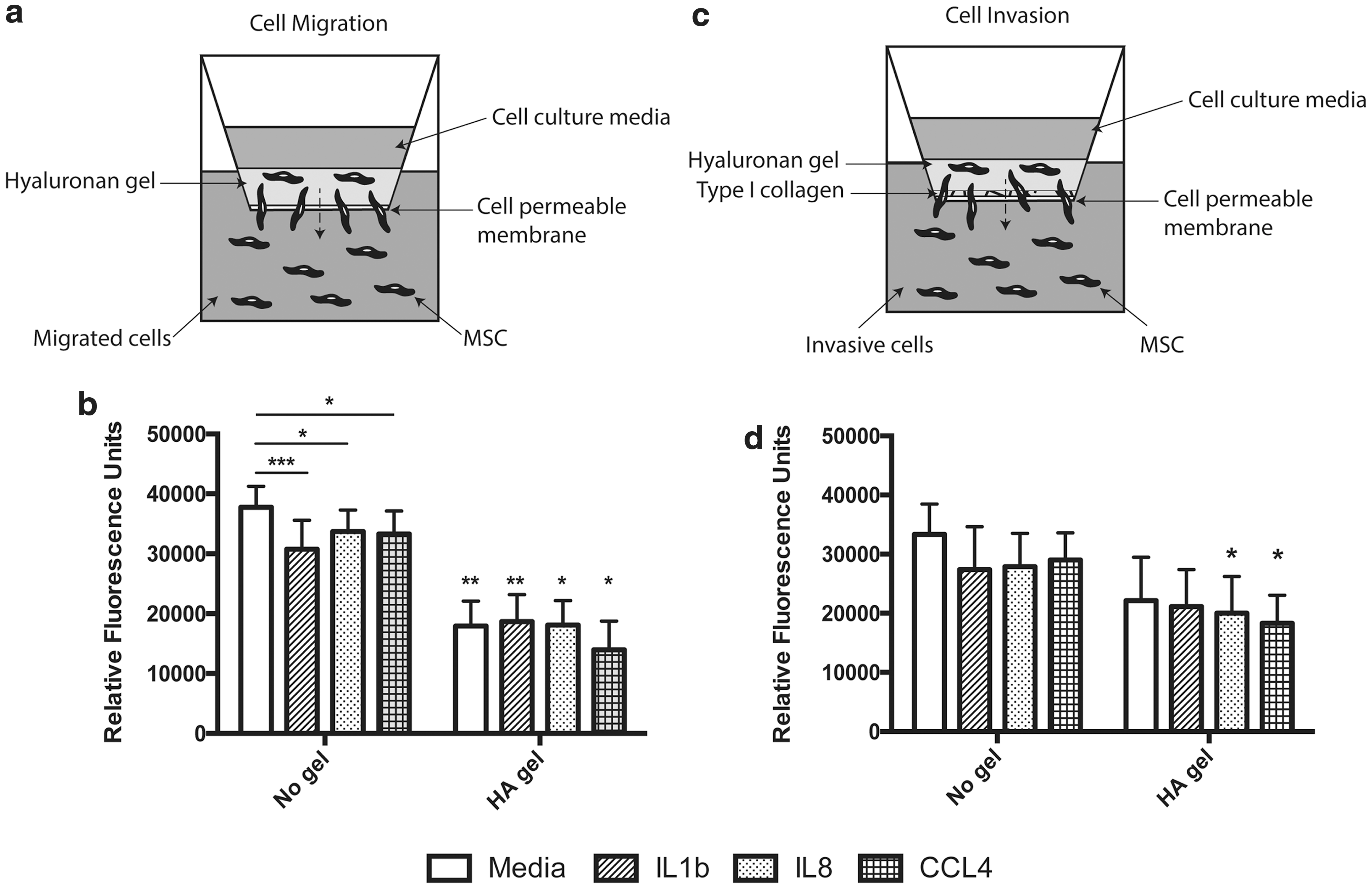
HA hydrogels delay directed migration of MSCs in vitro. MSCs ± HA hydrogel was seeded onto cell permeable membranes ± type I collagen coating to evaluate
The ability of MSCs ± HA to invade through a collagen matrix was further evaluated. MSCs within HA hydrogel invaded through the matrix at a comparable frequency to liquid suspension controls (Fig. 1d). Under these conditions only IL8 and CCL4 suppressed the invasion of the HA-incorporated MSCs (Fig. 1d; P < 0.05) compared with controls. No reduction in invasion was seen with exposure to proinflammatory cytokines with MSCs in suspension (Fig. 1d).
MSCs modulate their expression of matrix remodeling genes in response to HA hydrogel incorporation and inflammation
MSCs expressed all three isoforms of the HAS gene (Fig. 2a–c). Levels of HAS1 remained unaltered by gel incorporation or exposure to proinflammatory stimuli (Fig. 2a). HAS2 expression was downregulated with IL1β exposure in MSCs incorporated in HA gel compared with controls (Fig. 2b; P < 0.05). HAS3 was upregulated in MSCs exposed to IL1β in control cultures (Fig. 2c; P < 0.01).
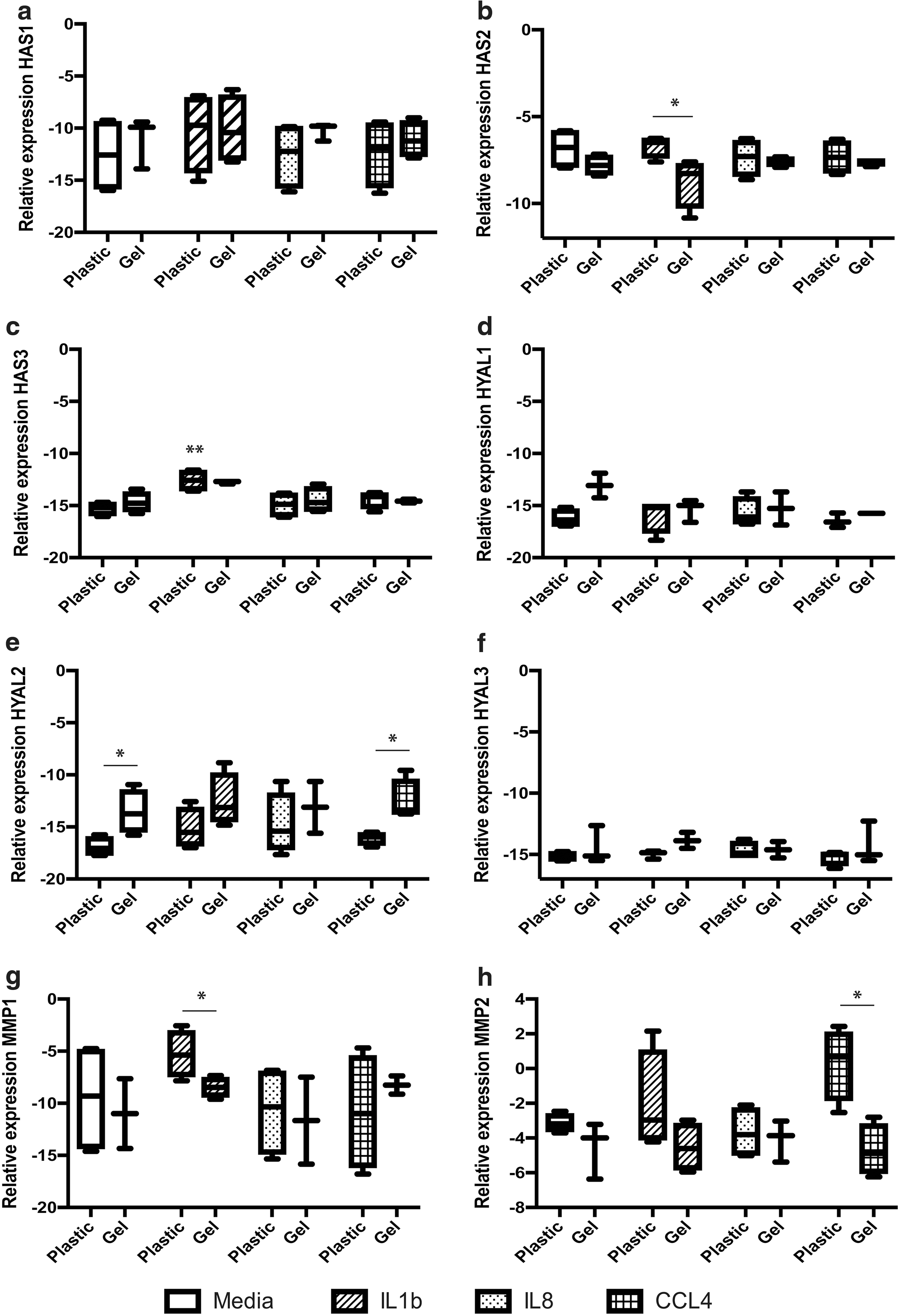
HA hydrogel and exposure to proinflammatory factors differentially regulates MSC expression of matrix remodeling genes. MSCs ± HA hydrogel were incubated with the proinflammatory factors, IL1β, IL8, and CCL4, for 24 h and assessed for their mRNA expression of
HYAL1–3 were expressed by MSCs, although HYAL1 and 2 were most highly expressed (Fig. 2d–f). HYAL4 was weakly expressed, and HYALP and PH-20 were not detected within the MSCs (data not shown). HYAL2 was upregulated with HA gel incorporation of the MSCs (Fig. 2e; P < 0.05). This effect was also seen in the CCL4-treated cultures (Fig. 2e; P < 0.05). No significant changes in expression were seen in HYAL1 or 3 between experimental groups (Fig. 2d, f).
MMP expression was differentially regulated by exposure to proinflammatory stimuli (Fig. 2g, h). Expression of MMP1 was decreased by IL1β stimulation in HA hydrogel cultures compared with IL1β-treated controls (Fig. 2g; P < 0.05). In contrast, MMP2 was downregulated in HA gel cultures exposed to CCL4 compared with their respective controls (Fig. 2h; P < 0.05). MMP8 and 9 were not detected after HA gel incorporation, with similar effects evident on MMP13 transcript (data not shown).
MSCs within HA hydrogels upregulate secretion of immunomodulatory factors in response to inflammatory stimuli
MSCs ± HA gel upregulated their secretion of IL6 in response to IL1β compared with their respective controls (Fig. 3a; P < 0.05). IL8 secretion was only upregulated in plastic cultured MSCs, compared with controls, in response to IL1β (Fig. 3b; P < 0.05). No significant changes in the antiscar-promoting growth factor, HGF, or VEGF, were seen between plastic and HA gel-contained cells, in control, or treated samples (Fig. 3c, e). In contrast, an upregulation of the immunomodulatory and scar-promoting factor, TGFβ1, was seen with MSCs in HA gel exposed to IL1β compared with MSCs on plastic (Fig. 3d; P < 0.05).
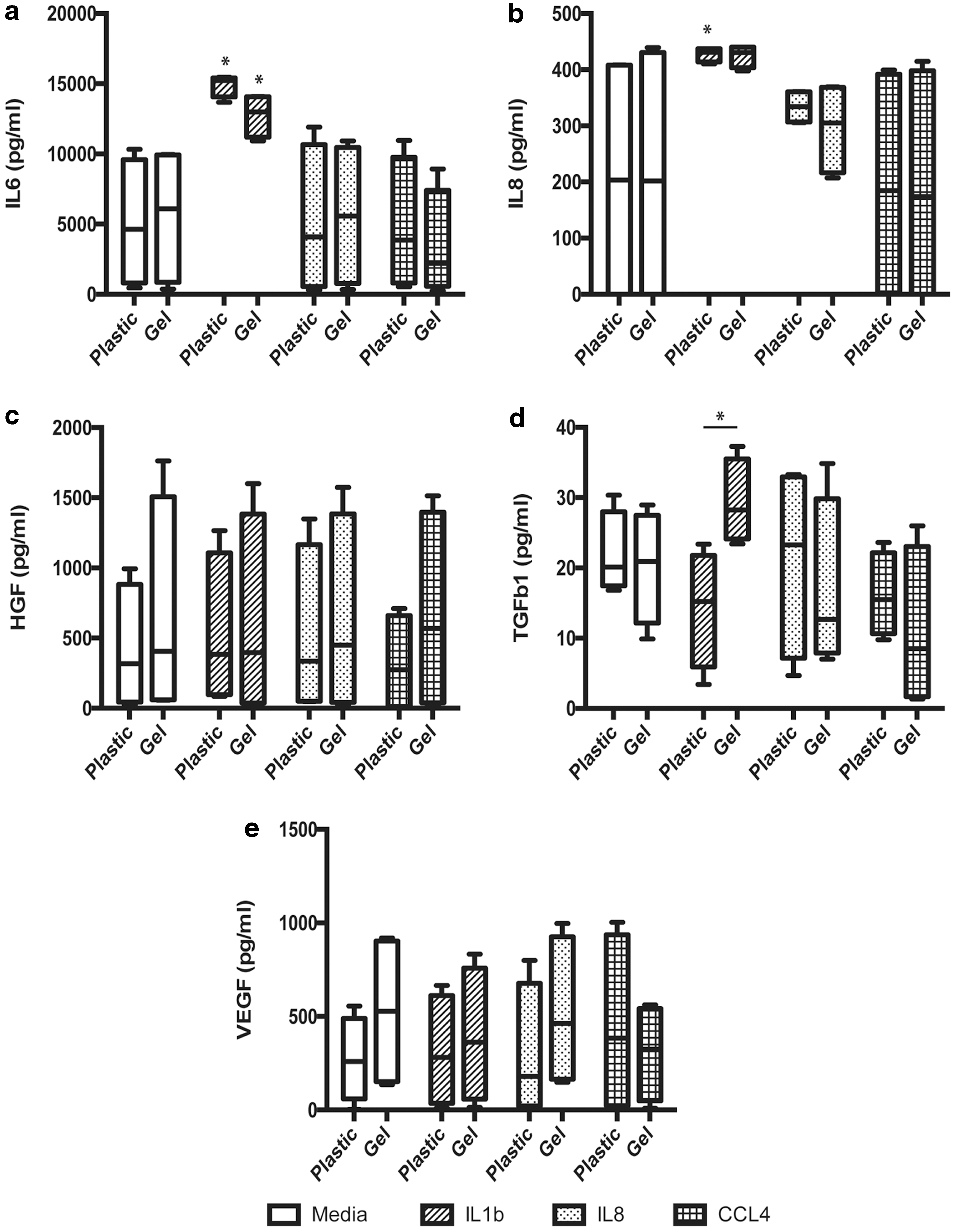
HA hydrogel incorporation modulates MSC responsiveness to inflammation. MSCs ± HA hydrogel were incubated with the proinflammatory factors, chemokines IL1β, IL8, and CCL4, for 24 h and assessed for secreted levels of the immunomodulatory prohealing factors
MSCs suppress inflammation within the injured VF
To evaluate whether HA hydrogel-incorporated MSCs exhibit a delayed inflammatory resolution effect, or if the gel itself induced inflammatory responses during the resorption process, we histologically evaluated levels of inflammation within the animals after treatment. At day 25, no significant differences in the level of inflammation between MSC-treated VFs and normal unoperated controls were observed (both MSCs in suspension and HA hydrogel; Fig. 4a, d, e and Table 2). Injured VFs ± HA hydrogel, but no MSCs, had a significantly higher rating for inflammation compared with unoperated controls (P < 0.05; Fig. 4a–c and Table 2). Injury induced fibrosis within the VF compared with uninjured controls (Table 2). Delivery of MSCs did not reverse fibrosis, with no difference in score between injured VFs and any of the treatment groups (Table 2).

MSC treatment diminishes inflammation in injured VFs. Hematoxylin and Eosin staining of rabbit VFs at day 25, depicting no difference in the level of inflammation between animals treated with MSCs in
The Effects of Mesenchymal Stromal Cell Treatment ± Hyaluronan Hydrogel on the Vocal Fold
The effect of MSC treatment ± HA hydrogel was assessed on vocal fold fibrosis, inflammation, Alcian Blue staining levels, LP thickness and relative type I collagen content.
(A) No/minimal fibrosis, (B) focal or noncompact fibrosis in the LP or superficial vocal muscle, (C) compact fibrosis in the LP and superficial muscle, (D) compact fibrosis in the LP and superficial and deep vocal muscle.
(0) No/limited, (1) moderate inflammation within the LP and/or superficial thyroarytenoid muscle, (2) severe inflammation within the LP and/or superficial and deeper muscle.
(0) No/minimal staining, (1) focal increase in staining within the LP and superficial vocal muscle, (2) staining of the LP, superficial and deep muscle.
P < 0.05 compared with uninjured (d) and injury only (e) controls.
HA, hyaluronan; LP, lamina propria; MSC, mesenchymal stromal cell; SD, standard deviation of the mean.
MSCs reduce type I collagen levels
As expected, the injured VFs had a significantly higher relative level of type I collagen compared with uninjured controls (P < 0.05; Table 2). VFs treated with MSCs in suspension, but not HA hydrogel, demonstrated a reduction in type I collagen content compared with injured controls, with levels comparable to that of the uninjured control group (P < 0.05; Table 2).
HA hydrogel degrades in vivo
Alcian Blue staining demonstrated no difference in the levels of mucopolysaccharides (including HA) between treatment groups and control groups at day 25 (Fig. 5 and Table 2).

HA hydrogels are degraded in vivo. Alcian Blue staining was performed on VF sections at day 25 to detect mucopolysaccharides (blue color), including HA. No difference between
HA hydrogel incorporation does not support persistence of MSCs in vivo
In vitro quantification of cell number after HA hydrogel incorporation and syringing confirmed no significant differences compared with controls (day 0) at 1 or 3 days (Fig. 6a). Despite this, only low numbers of MSCs were detected in vivo within VFs injected with HA hydrogel-delivered cells at day 3 (2.5% ± 3.4%; Fig. 6b). No MSCs were detected after 25 days in either suspension or HA hydrogel-delivered cells (Fig. 6c, d).
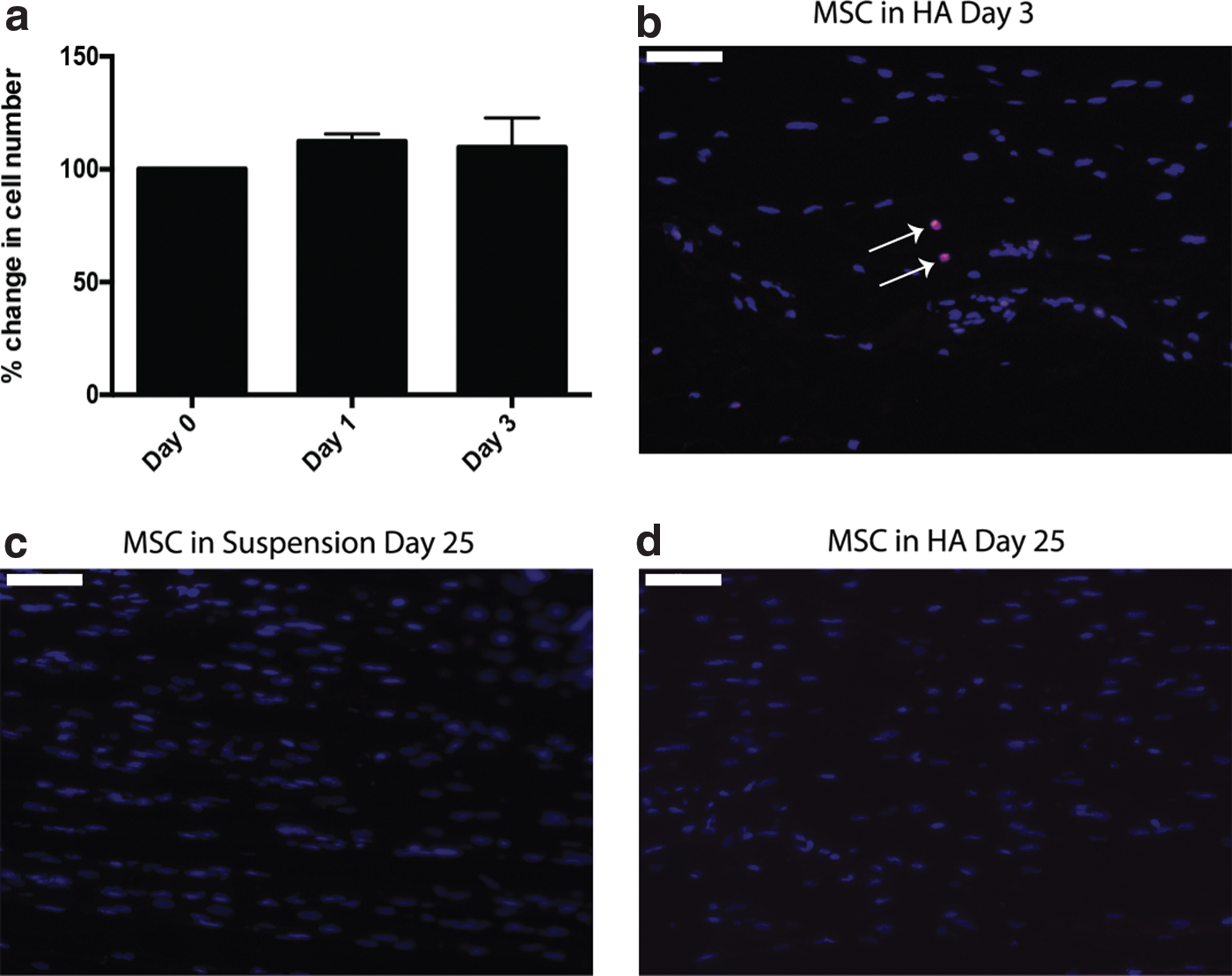
HA hydrogels do not enhance MSC persistence in vivo.
Discussion
The potential for MSCs to modulate the immune response during wound healing has been exemplified in many contexts and systems [32]. We, as well as others, have previously demonstrated that local injection of MSCs into VF wounds, as seen after excision of fibrotic scar tissue within the clinical setting, aids in tissue regeneration and restoration of VF function [2,29,30,33 –37]. These in vivo findings have demonstrated the potential for MSC therapy in the context of clinical VF scar resection. Limitations to this approach exist though, with large, critical-sized defects requiring supporting matrix to bridge the injury site. Furthermore, the issue of MSC leakage from the tissue with delivery in a liquid suspension requires addressing. The principle aim of this study was to evaluate whether delivery of MSCs within a HA hydrogel offers a safe alternative to current liquid delivery vehicles, holding the MSCs at the site of injury, potentially increasing their longevity, minimizing leakage, and providing a scaffold akin to the extracellular matrix (ECM) of the VF for large defects. To ensure the therapeutic effect of MSCs within the hydrogel is not negatively impacted, this study also addresses the effect of HA hydrogel incorporation on MSC viability and functionality to resolve inflammation and promote VF healing.
Previous studies have explored the effects of combining MSCs with a scaffold or HA hydrogel to enhance implantation of the cells into the VF and potentially improve healing with normal VF architecture and VF ECM, reduced type I collagen, and improved tissue viscosity [33,38,39]. Our in vivo experimentation supports studies within the field demonstrating that HA hydrogel delivery of MSCs into VF defects offers a safe alternative to currently employed cell suspension systems [33 –37]. No animals exhibited serious adverse side effects, such as edema within the VF, in response to the gel. Furthermore, no inflammatory reaction was triggered by the gel.
In vitro analysis was performed to investigate whether delivery of the MSCs within a HA hydrogel would impede cell migration and/or limit cellular responsiveness to the extracellular milieu. Our data demonstrated that the MSCs remained viable within the gel and exhibited a delayed, although not halted, migration compared with MSCs in liquid suspension. The presence of chemotactic collagen types within the LP may also encourage the cells to leave the HA hydrogel upon administration [40]. In vitro invasion assays confirmed that the presence of an underlying type I collagen matrix supported extrication from the gel and invasion through the collagen matrix in a manner comparable to the liquid suspension cells.
We have previously reported that MSCs are retained within the VF tissue for short periods of time, exerting their long-term immunomodulatory and prohealing effects through indirect mechanisms, including induction of the innate immune system and the promotion of anti-inflammatory macrophages [28]. These initial analyses suggested that MSCs would not be retained within the HA hydrogel upon in vivo delivery, where a rich collagen matrix would be present; thereby exposing the cells to immune cells resulting in their elimination. These findings correlated with our in vivo data, with low-level detection of the MSCs within the VF tissue demonstrated, irrespective of the delivery vehicle used.
Within this study we used PCR array of the whole VF tissue to evaluate the change in environmental milieu in response to the resection injury that we are inducing with this model. Our analysis confirmed that the principal proinflammatory signals seen within the tissue directly after injury were cytokines, such as IL1β and chemokines, including the neutrophil recruitment and activation factors IL8 and CCL4. We, therefore, wished to evaluate the impact of the HA hydrogel on the responsiveness of the MSCs to these factors. IL6 was not chosen for evaluation, despite its high degree of upregulation, due to the fact that MSCs themselves produce high levels of this cytokine, impacting on our ability to delineate mechanistic effects in vitro.
IL1β, a key cytokine present during the acute injury phase of the wound healing process, was demonstrated to be the principal inflammatory regulator of HA signaling pathways within MSCs. HAS2 was downregulated in response to IL1β in HA-encapsulated MSCs, whereas HAS3 was upregulated in suspension cells. HAS2 has been linked to early injury responses, promoting long-chain, anti-inflammatory HA production within the VF. Such downregulation by the gel-encapsulated MSCs may indicate an intrinsic recognition of HA presence within the cell's immediate extracellular environment [41]. The upregulation of HAS3 within the suspension cells, and a similar trend, although not statistically significant, within the HA-incorporated cells, follows the expression pattern seen in nonscarring oral mucosal fibroblasts [42]. This is of significant interest, as the stark contrast in HAS3 expression response at the mRNA level upon detection of IL1β in oral fibroblasts, compared with scar-forming fibroblasts, has been hypothesized to represent the differential scar formation evident between these two tissues [42,43]. Interestingly, the gel itself induced an upregulation of HYAL2, one of the principal HYALs within somatic tissues, confirming that the HA hydrogel and MSCs are communicating, most likely through the HA-CD44 pathway [44,45].
The proinflammatory cytokines evaluated within this study exerted differential effects on the mRNA expression of MMPs within the MSCs. A consistent trend of downregulated expression of the collagenases, MMP8 and 13, was seen with HA hydrogel presence, with undetected transcript in all or the majority of donors evaluated. This may be a direct result of the HA crosslinking procedure during gel formation, with one previous study suggesting that different crosslinking methods can be utilized to exert collagenase and HYAL inhibition in the treatment of osteoarthritis [46]. However, this finding is also supported by the wider literature with evidence suggesting low-level mRNA detection of MMP8 post-adipose MSC administration in a model of VF scarring [36]. Interestingly, IL1β exposure significantly decreased MMP1 expression in HA hydrogel-incorporated MSCs. This licensing, or shift in the MSC phenotype to upregulate anti-inflammatory responses, was also evident when evaluating their secretome. Elevated levels of IL6 and TGFβ1 (only MSCs in the hydrogel) were seen upon IL1β stimulation, with TGFβ1 known to decrease MMP1 expression in dermal fibroblasts [47]. IL6 is the principal cytokine linked to skewing of monocytes to anti-inflammatory macrophages by MSCs [48]. Interestingly, increased IL6 and associated enhanced stromal–monocyte interactions facilitated by IL1β, may play a role in the skewing of monocytes that we have previously reported with MSC injection to the injured VF [28].
Although TGFβ1 has been linked to scar formation within the VF, elevated levels during the acute injury response have also been linked to immunomodulation, with TGFβ1 exerting multifaceted roles in innate (monocyte and natural killer cell) immune suppression, and also in promoting the induction of regulatory T cells [49]. Our in vivo experiments support this, with no increased fibrosis with HA-incorporated MSCs compared with MSCs alone. Injury itself induced fibrosis and elevated levels of type I collagen, although no significant difference in relative type I collagen content was detected in any treatment group compared with uninjured controls. A recent detailed analysis of VF tissue composition and architecture, post-MSC administration, using atomic force microscopy alongside immunohistochemical and histological analyses, demonstrated that MSCs can restore the natural tissue architecture and ratio of type I and III collagens [37]. Alcian Blue staining confirmed that HA hydrogels can be used safely for the delivery of MSCs, inducing no edema or inflammatory responses and supporting knowledge that the gel can be resorbed within 1 month [6,16,21]. While, as previously reported, MSCs administered in HA hydrogel do not exhibit an enhanced healing response, leakage noted when injecting MSCs in liquid suspension into the VF was greatly reduced with HA hydrogel delivery [33,50].
Inflammation was consistently reduced by the presence of the MSCs, with the HA hydrogel exerting no inflammatory resolving effects alone. These data support those reported by King et al., who demonstrated no macrophage inflammatory response using MSCs in combination with HA hydrogels in a porcine model of VF injury [50]. These findings confirm that MSCs exert anti-inflammatory, prohealing effects within the injured VF and while this function is not impeded by the HA hydrogel, gel alone is not sufficient to enhance tissue repair. These findings correlate with previous preclinical and clinical reports, where pure HA injections have been noted as safe, although with variable efficacy in the treatment of scarred VFs [21,22,33,50].
This study demonstrates that HA hydrogels can be safely employed for the delivery of MSCs into injured VFs. Use of the HA hydrogel minimizes leakage of cell suspension during injection and provides a scaffold supporting the healing of critical-sized defects. Furthermore, the gel was resorbed within 1 month of delivery with no evidence of increased edema within the tissue. MSCs incorporated within the gel did not persist longer within the VF, but exerted comparable therapeutic effects and demonstrated responsiveness to the extracellular environment both in vitro and in vivo. We conclude that HA hydrogels offer a safe and effective method for the delivery of MSCs into injured VFs.
Footnotes
Acknowledgments
The authors acknowledge Tim Bowden at the Department of Chemistry, Ångström Laboratory, Uppsala University for discussions over the study and help with the preparation of hyaluronan. This work was supported by the Swedish Medical Research Council (K2011-X-20742-04-6), Stockholm County Council (20110152), Stiftelsen för Strategisk Forskning (IKE-2014-00354), Laryngfonden (50/14), and Karolinska Institutet.
Author Disclosure Statement
No competing financial interests exist.
Supplementary Material
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.