
Abstract
Human perivascular progenitor cells, including pericytes, are well-described multipotent mesenchymal cells giving rise to mesenchymal stem cells in culture. Despite the unique location of pericytes, specific antigens to distinguish human pericytes from other cell types are few. Here, we employed a human tissue microarray (Human Protein Atlas) to identify proteins that are strongly and specifically expressed in a pericytic location within human adipose tissue. Next, these results were cross-referenced with RNA sequencing data from human adipose tissue pericytes, as defined as a fluorescence activated cell sorting (FACS) purified CD146+CD34−CD31−CD45− cell population. Results showed that from 105,532 core biopsies of soft tissue, 229 proteins showed strong and specific perivascular immunoreactivity, the majority of which (155) were present in the tunica intima. Next, cross-referencing with the transcriptome of FACS-derived CD146+ pericytes yielded 25 consistently expressed genes/proteins, including 18 novel antigens. A majority of these transcripts showed maintained expression after culture propagation (56% of genes). Interestingly, many novel antigens within pericytes are regulators of osteogenic differentiation. In sum, our study demonstrates the existence of novel pericyte markers, some of which are conserved in culture that may be useful for future efforts to typify, isolate, and characterize human pericytes.
Introduction
Pericytes are mesenchymal progenitor cells that express mesenchymal stem cell (MSC) surface markers in situ and in culture, and they display multipotentiality toward osteogenic, adipogenic, and chondrogenic cell lineages. Pericytes are ubiquitously present around arterioles, capillaries, and venules in human organs [1 –3]. Current data support the notion that pericytes have organ-specific functions in tissue repair while maintaining progenitor cell properties [4]. Traditional pericyte markers include NG2 [1], CD146 [5], platelet-derived growth factor receptor-beta (PDGFRβ) [5], and α-smooth muscle actin (αSMA) [6], with absence of endothelial cell (EC) specific antigens such as CD31, CD34, or von Willebrand factor (vWF). However, there exists a paucity of specific markers for the identification of human pericytes.
Here, we used the Human Protein Atlas (HPA) to help define such markers by identifying proteins expressed in a pericytic location within human adipose tissue. The HPA is a large tissue microarray (TMA) dataset used to provide protein expression information at the tissue, cellular, and, occasionally, subcellular level [7]. The HPA was initiated in 2003 and a first version of the public database
To identify novel antigen expression within pericytes, candidate surface markers of pericytes were identified via HPASubC analysis. Therefore, cell protein surface markers found from TMA were cross-referenced with RNA-Seq data from freshly sorted or cultured pericytes to validate protein expression. Pericytes from different tissue origins might express differential surface markers. Thus, these proteins were further investigated by TMA for expression in tissues other than adipose tissue. Our study demonstrates the existence of novel pericyte markers, some of which are conserved in culture that may be useful for future efforts to typify or isolate human pericytes.
Materials and Methods
HPASubC software
The HPASubC software has been validated, published [9,10], and can be downloaded from
General HPASubC usage
A list of all proteins evaluated at HPA is first accessed by using normal_tissue.csv.zip, available at
HPASubC usage to identify novel markers of pericytes
For identification of novel pericyte markers, all images with weak, moderate, or strong staining of blood vessels and moderate nonspecific staining of other tissues were excluded. Conversely, samples with strong, nonspecific staining were included. Images were scored on a scale of 0–3 based on the following parameters: 3: Specific positive staining of blood vessels; 2: Positive staining of tissues other than blood vessels AND blood vessels have stronger staining than other kinds of tissues; 1: Positive staining of more than two kinds of tissues on the slide OR has at least one other tissue stained more intensely than blood vessels; 0: Poor images; very little staining of blood vessels. However, it is challenging to distinguish whether pericytes or ECs are stained from the IHC images on the TMA. In this study, proteins that were repeat scored 3 were cross-referenced with fresh and cultured pericyte RNA-seq datasets.
Isolation of human pericytes from adipose tissue
Human adipose lipoaspirate specimens used for fluorescence activated cell sorting (FACS), culture, and RNA-seq analysis were obtained from six healthy female donors (age range: 35–63 years; body mass index range: 21.1–32.0) undergoing elective liposuction from lateral axillary areas at local plastic surgery clinics under IRB approval. Tissue dissociation and pericyte isolation were performed as previously described [11]. The cells were collected by centrifugation and incubated in Aldefluor assay buffer containing fluorescently labeled, mouse antihuman monoclonal antibodies (PE- or APC-Cy7-conjugated CD45, and PE- or PerCP-Cy5.5-conjugated CD146; BD BioSciences, San Jose, CA) for 20 min on ice to allow immunophenotypic staining for FACS. Cells collected by FACS were either lysed immediately for RNA extraction or seeded for culture expansion. To adjust for gene expression changes brought about in culture, after three passages, cultured pericytes were stained with CD31, CD45, and CD146 fluorescent antibodies and sorted by FACS a second time. Pericytes that had retained CD146 expression in culture were isolated for RNA-seq analysis.
RNA extraction and sequencing
Subpopulations of freshly isolated pericytes and their cultured counterparts were sorted into a cell lysis buffer solution, and RNA was extracted by using the Qiagen RNeasy Micro kit (Qiagen). Total RNA was quantified and measured for RNA integrity numbers of 7.7 or greater (2100 Agilent Bioanalyzer). For each subpopulation, complementary DNA (cDNA) was then prepared by using the NuGen's Ovation RNA-Seq System V2 kit from ∼100 ng of RNA pooled from two or three donors and used as input for Ovation's Ultralow DR Multiplex System 1–8 (Nugen, San Carlos, CA). High-throughput sequencing was conducted on an Illumina Hiseq 2000 platform to generate 100-bp paired-end reads at a depth of coverage of ∼79 million reads. Cluster formation achieved ∼86%, whereas the quality of reads exceeded 95%. Fragment sequence identification and alignment were performed by using TopHat and Bowtie, respectively. Samples were normalized by RPKM (reads per kilobase of exon per million mapped reads), which takes into account both library size and gene length in within-sample comparisons. Cross-library, individual gene comparisons were made possible by using a similar number of reads between libraries (ie, depths of coverage).
Results
Identification of membrane proteins specific to and consistently expressed on blood vessels
To identify new perivascular antigens, we analyzed proteins from HPA adult human normal soft tissue images. A total of 105,532 soft tissue core images with 19,613 proteins were obtained from the HPA to be analyzed (Fig. 1). In step 1, we parsed out the ENSG IDs, and we obtained 59,529 images (56.4% of total soft tissue images), which contained 7,272 proteins (37.08% of total proteins) with perivascular immunoreactivity. In step 2, images containing proteins with a high specificity for perivascular expression (qualitative score of 3) were selected for further analysis (Fig. 2A). This yielded 1,952 images (1.85% of total soft tissue images) with 1,467 different proteins (7.48% of total proteins) that were highly specific for blood vessels.

Schematic of the selection process of HPA images. All HPA records are obtained and used to identify Ensemble gene IDs of immunostained proteins. HPA “Soft tissue” images are parsed out to capture and download TMA images. Images were selected and scored by specificity and consistency to identify novel markers for pericyte cell sorting. In step 1, images were divided into those with or without clear perivascular staining. In step 2, those remaining images were divided into those in which immunostaining was either specific or nonspecific to the perivasculature. In step 3, those remaining images were divided based on whether or not perivascular immunostaining was consistent across images. In the end, 328 proteins were found to be both specific and consistent within the perivascular space within the normal adult human “soft tissue” TMA dataset. HPA, Human Protein Atlas; TMA, tissue microarray.
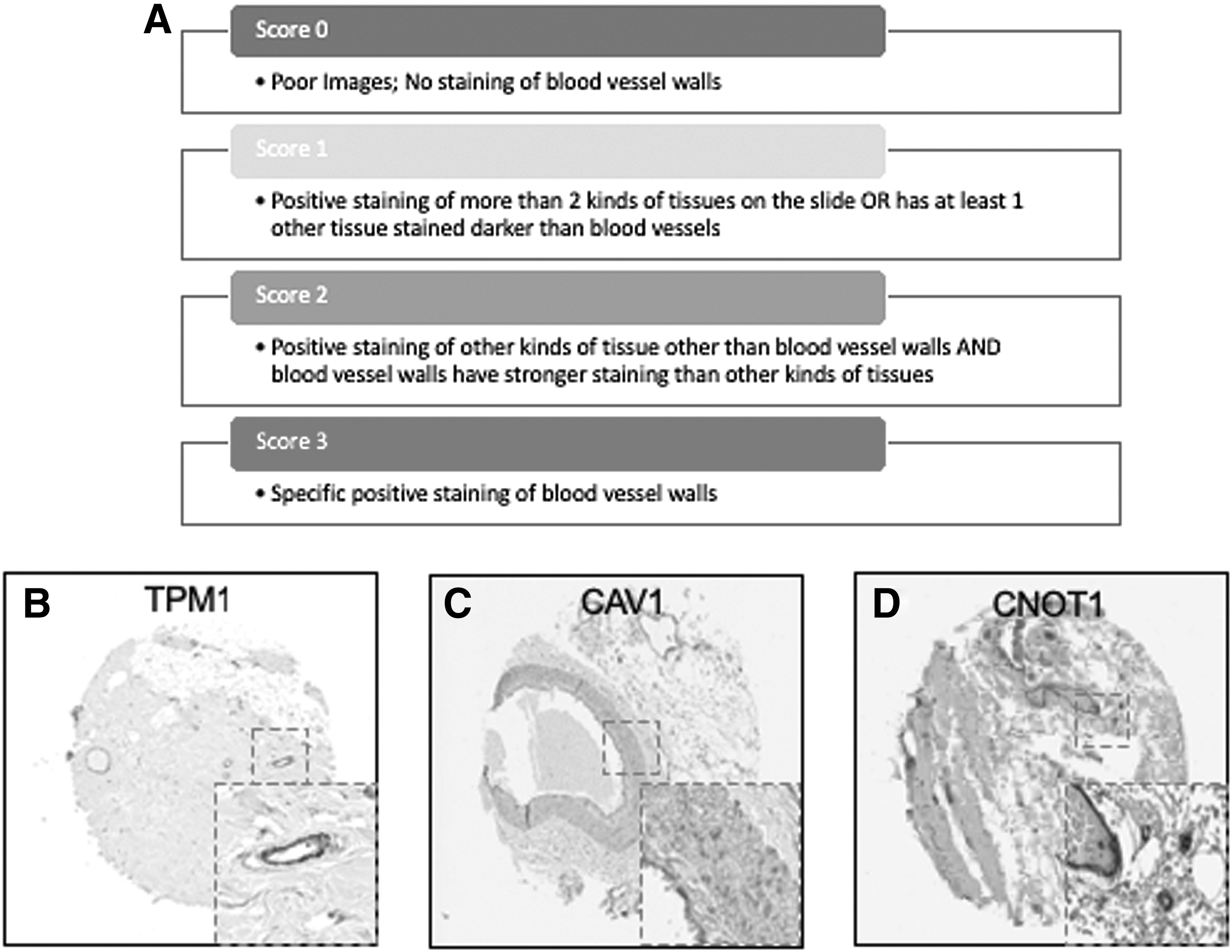
Scoring system of TMA imaging selection, and examples of specificity staining Score 3. Images with blood vessels were scored on a scale of 0–3 based on the following parameters: 3: specific positive staining of blood vessel walls; 2: positive staining of other kinds of tissue other than blood vessels AND blood vessel walls have stronger staining than other kinds of tissues; 1: positive staining of more than two kinds of tissues on the slide OR has at least one other tissue stained darker than blood vessel walls; 0: poor images; very little staining of blood vessel walls
In the TMA database, each protein of interest was probed by two or more antibodies. In some situations, one antibody might be more or less specific than the others. Thus, these images were further investigated to determine whether staining with different antibodies produced staining of the 1,467 proteins in the same perivascular location. Proteins with only 1 image with a score of 3 were excluded in Step 3, leaving 841 images (0.07%) of 328 proteins (1.67%) that were consistently found in the perivasculature (examples shown in Fig. 2B–D).
Pericytes reside within the tunica intima in a subendothelial location. Next, these 841 images of 328 proteins were analyzed to categorize their location within either the (1) tunica intima, (2) media, or (3) adventitia. Of the 328 remaining proteins, 155 are specific to the intima (eg, ENG), one (CDCP1) is specific to the adventitia, and 92 are specific to the media (eg, VWA9). Four were shared between the intima and adventitia, two (RDH16 and TNR) were shared between the adventitia and the media, 69 were shared between the intima and the media (eg, TPM1), and one (CAAP1) was shared among all three locations (Fig. 3A and Supplementary Table S1). Ninety-five media-specific proteins were excluded from further analysis, as medial SMCs are not considered to represent a multipotent cell type. Twenty-seven proteins were additionally excluded due to inconsistencies related to blood vessel staining pattern between samples (eg, DES). This left 229 proteins (Supplementary Table S2), which were consistently and specifically expressed within the tunica intima.
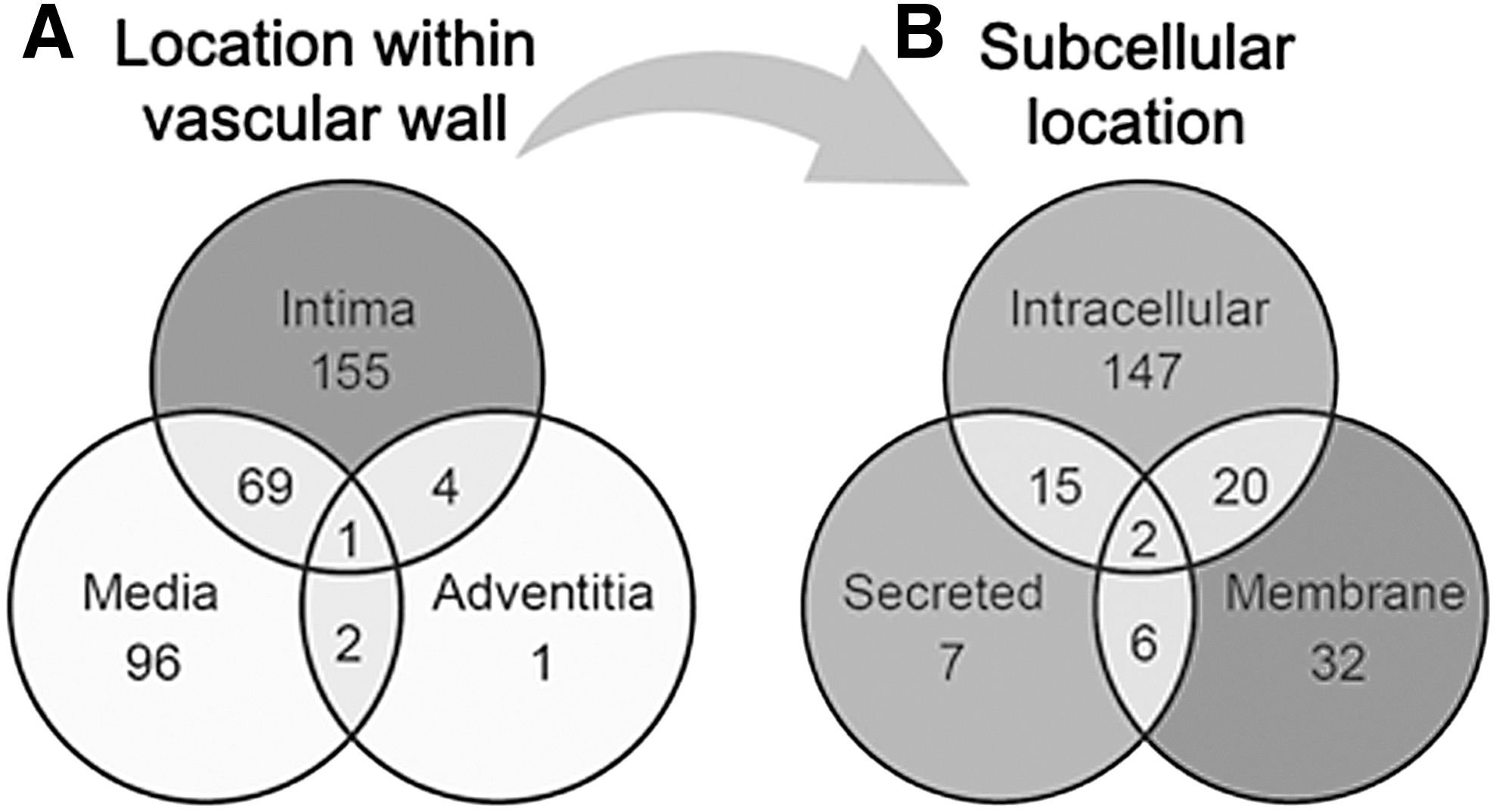
Histologic and subcellular location of perivascular antigens. (Left) Venn diagram to demonstrate the location of n = 328 perivascular antigens within different layers of the vessel wall, including the tunica intima, media, and adventitia. The majority of antigens were found within the tunica intima only (n = 155)
Among these 229 proteins (Supplementary Table S2), well-known pericyte markers ENG, Myl9 [12] have been reported to be associated with pericytes. Moreover, ABI3 [13], BECN1 [14], and ADPIOQ [15] have been reported to be highly expressed within pericytes. Endothelial markers, such as vWF [16], were included in this list, as it is difficult to distinguish from the TMA images whether ECs or perictytes are stained. In this study, these 229 proteins were further categorized by their subcellular location due to the nature of instability of secreted proteins and the inability to locate intracellular proteins.
The 229 proteins were then categorized by location within the cell to identify membrane proteins to serve as potential pericyte markers for FACS. One hundred and forty-seven proteins were intracellular only (eg, MYL3), 32 were restricted to the membrane (eg, GFRA3), and 7 were secreted proteins (such as COL15A1). Twenty proteins were both intracellular and membrane proteins (such as VWDE), 6 were both membrane and secreted proteins, 15 were both intracellular and secreted proteins, and 2 were present in all three locations (eg, C12orf76) (Fig. 3B and Supplementary Table S2). In sum, 60 out of the 229 proteins found to be specific to the tunica intima were also membrane-bound proteins, serving as possible novel surface marker proteins for pericyte isolation (shown in Table 1).
Sixty Membrane Proteins Specifically Expressed in the Tunica Intima of Vessel Wall in Normal Adult Human Adipose Tissue
Confirmation of protein expression within an RNA-Seq data base of human CD146+ pericytes
To confirm whether these 60 membrane proteins expressed within the tunica intima were significantly expressed in purified human adult adipose-derived pericytes, a cross-comparison was next performed by using RNA Sequencing data from freshly sorted and cultured human adipose-derived pericytes [17] (Fig. 4). Adipose-derived pericytes were isolated by incubation in Aldefluor assay buffer containing fluorescently labeled, mouse antihuman monoclonal antibodies to CD146, as well as negative selection for CD31 and CD45.
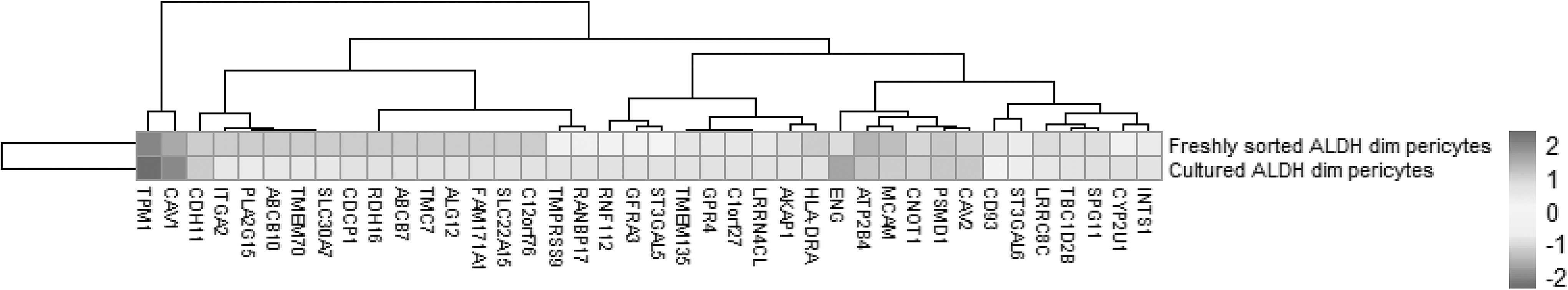
Heatmap of gene expression in adipose tissue-derived fresh sorted and cultured pericytes. CD146+Aldhdim human pericytes were derived from n = 6 lipoaspirate samples from healthy female donors. Total RNA-Seq was performed on purified human pericytes, either immediately after FACS (freshly sorted) or after culture propagation (cultured). Samples were normalized by RPKM. The culture time point (y-axis) and level of gene expression (x-axis) is color-coded from dark gray to light gray. CD146 (MCAM) expression is included as a positive control. FACS, fluorescence activated cell sorting; RPKM, reads per kilobase of exon per million mapped reads.
RNA-Seq data showed that 38 out of the 60 TMA-identified proteins were expressed in adipose-derived human pericytes (Tables 2 and 3). As a positive control, the pericyte marker CD146 (MCAM) used to isolate pericytes was also examined, which showed consistent expression among purified cells, both freshly after sorting and after culture expansion. Eight out of 38 have only weak expression in pericytes (RPKM = 1 freshly sorted and after culture). Five out of 38 had low expression (RPKM = 1) in freshly sorted pericytes, but increased expression after culture. Fourteen out of the 25 membrane proteins were consistently expressed both on freshly sorted pericytes and after culture. Several of these transcripts increased in expression after culture, such as TPM, CNOT1, CYP2U1, and INTS1 (Table 3). Many transcripts were detectable at lower levels after culture, including CAV1, ENG, ATP2B4, CAV2, PSMD1, SPG11, TBC1D2B, LRRC8C, ST3GAL6, and CD93. The remaining 11 genes were expressed on the membranes of freshly sorted pericytes, but they were lost after culture (HLA-DRA, AKAP1, TMEM135, GPR4, C1orf27, LRRN4CL, GFRA3, ST3GAL5, RNF112, TMPRSS9, and RANBP17) (Table 3).
Perivascular RNA Transcripts Expressed Among Both Freshly Sorted and Culture Expanded CD146+ Human Pericytes
Of the 25 genes/proteins expressed present in human pericytes by both TMA and RNA-Seq, 14 genes were expressed among both freshly sorted and cultured pericytes. Data were normalized by per million mapped reads (RPKM) dataset. Corresponding heatmap is shown in Fig. 4.
RPKM, reads per kilobase of exon per million mapped reads; TMA, tissue microarray.
Perivascular RNA Transcripts Expressed Among Freshly Sorted but Not Culture Expanded CD146+ Human Pericytes
Of the 25 genes/proteins expressed present in human pericytes by both TMA and RNA-Seq, 11 genes were expressed among freshly sorted pericytes, but they were lost after culture propagation. Data were normalized by per Million mapped reads (RPKM) dataset. Corresponding heatmap is shown in Fig. 4.
Confirmation of novel pericyte antigen expression in other tissues
Emerging concepts suggest that pericytes may have tissue-specific properties [18 –20], including tissue-specific antigen expression [21]. For this reason, we next examined immunolocalization of these 25 novel “pericyte” antigens in all other tissues available within the HPA (including 11 additional tissue depots from visceral and mesenchymal locations). Each image was again scored based on perivascular specificity (see again Fig. 2). A wide range of perivascular specificity was observed (Table 4), and most antigens showed significant variation across tissues in terms of relative specificity for blood vessels. Several markers were highly specific for a pericytic location across tissues, including, for example, CD93 (Fig. 5A–C, mean specificity score: 2.9) and ST3GAL5 (mean score: 2.58).
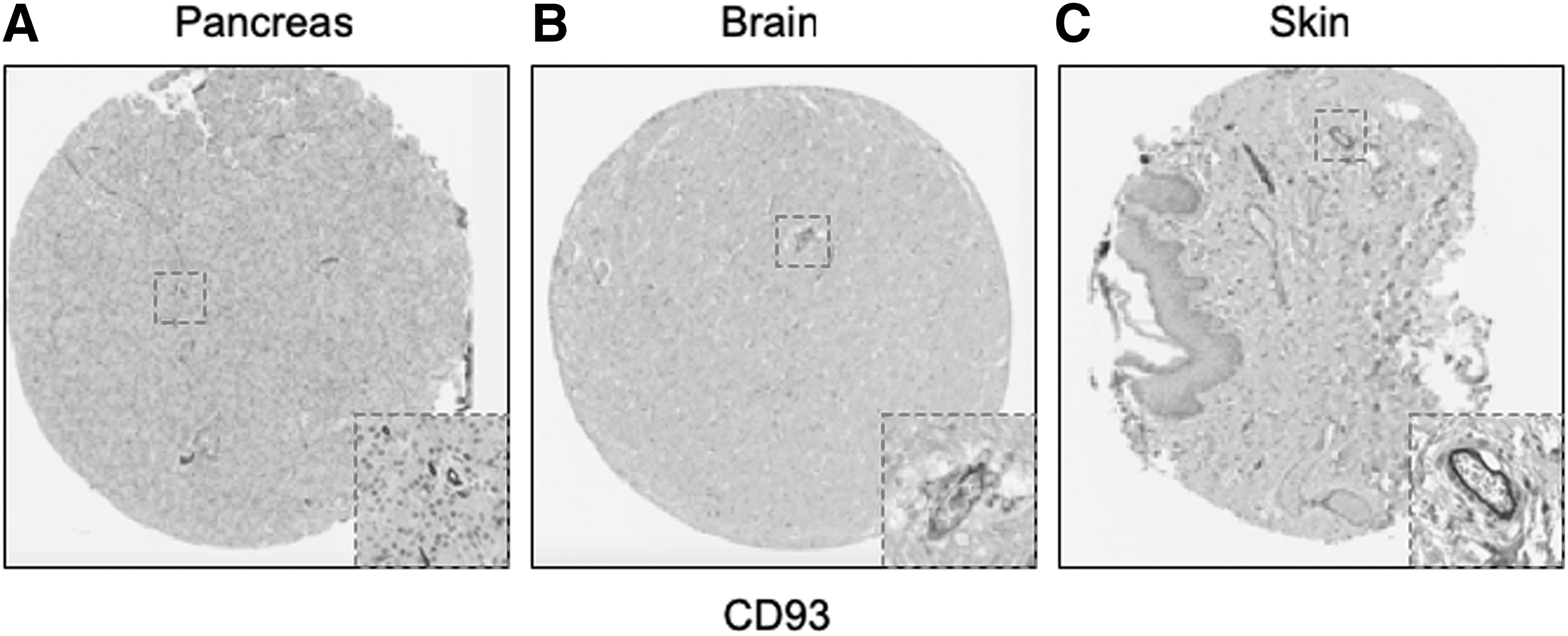
Representative examples of conserved pericytic CD93 immunoreactivity across tissues. Of all identified pericyte antigens, CD93 showed the highest specificity for a pericytic location across human tissues. Examples shown within the HPA TMA, including
Twenty-Five Pericyte Antigens and Their Expression Across Diverse Human Tissues Within the Human Protein Atlas
Twenty-five novel “pericyte” antigens in all other tissues available within the HPA (including 11 additional tissue depots from visceral and mesenchymal locations). Each image was again scored based on perivascular specificity (see again Fig. 2).
HPA, Human Protein Atlas.
Discussion
In summary, using the HPA TMA, 60 novel membrane proteins were found to be consistently and specifically expressed in the tunica intima. Twenty-five of these proteins were confirmed by RNA-Seq data to be expressed in purified human adipose-derived pericytes. Of these, several pericyte antigens were conserved across organ systems. These 25 proteins serve as potential pericyte cell surface markers for future efforts in the FACS isolation of human adipose tissue pericytes. Interestingly, a large minority (7) of these 25 antigens expressed at the gene and protein level in human pericytes have known roles in regulating osteogenic differentiation. Specifically, GPR4, CAV1, TMEM135, ENG, SLC30A7, CDCP1, and ITGA2 have all been reported to have roles in MSC differentiation and osteogenesis [22 –33]. Among the seven membrane proteins that have been reported to have functions in MSC differentiation and osteogenesis, ITGA2 and TMEM135 have positive regulatory effects on osteogenic differentiation [21,25,27,34]; whereas ENG, GPR4, CAV1, SLC30A7, and CDCP1 have negative regulatory effects [23,26,28,29,32,33].
As mentioned, and of these new pericyte antigens, a large minority have been previously reported to have functions in the regulation of osteogenic differentiation of cells. For example, both Integrin alpha-2 ITGA2 and TMEM-135 have been reported to positively regulate MSC osteogenic differentiation. The membrane glycoprotein ITGA2 regulates pericyte osteogenic differentiation and osteoblast maturation [25,27,32]. TMEM135 has also been shown to have effects on osteogenesis in bone marrow stromal cells [34]. Conversely, several pericyte antigens negatively regulate osteogenic differentiation. Endoglin (ENG, CD105) is a co-receptor for transforming growth factor beta (TGF-β) and canonical “MSC” marker, which has been previously observed to be expressed on human pericytes across tissues [3,35]. Most studies suggest that the absence of CD105 on mouse and human MSC results in increased osteogenic differentiation [33,36]. G protein-coupled receptor 4 (GPR4) is expressed in multiple organs, including the vasculature [23], and has been observed to inhibit the osteogenesis of bone marrow stem cells [24]. Caveolin-1 (CAV1) is a scaffolding protein of cholesterol-rich caveolae lipid rafts in the plasma membrane [37]. The caveolin-1 null mouse has increased bone mass and formation, and its bone marrow-derived MSCs have enhanced osteogenic potential [32]. SLC30A7(ZnT7) inhibits osteogenesis through Wnt/β-catenin signaling. CUB domain-containing protein 1 (CDCP1) is almost exclusively expressed in a subset of CD34(+) stem/progenitor cells in the bone marrow and is downregulated during MSC differentiation down osteogenic, adipogenic, and chondrogenic cell lineages [38]. Given the high number of negative regulators of ossification endogenously present within human pericytes, it is interesting to speculate whether these have functional relevance during pathophysiologic calcification and ossification of vessel walls.
The remaining 18 proteins have no reported function in osteogenesis on MSCs and can be divided into 5 categories based on current knowledge of their location or function: (1) highly expressed on ECs; (2) predominately expressed in neurons and the brain; (3) function in immune cells; (4) functions in stem cell differentiation, cancer progression, or cell metabolism; and (5) unknown.
Three proteins found in our study are markers that are shared with the endothelium or neurons. Chromosome 1 open reading frame 27 (C1orf27), SLC22A15 (FLIPT1), and CD93 are shared with the endothelium. C1orf27 exhibits cytoplasmic expression in epidermal tissue and plays a role in cell fate specification and development [39]. SLC22A15 (FLIPT1) is expressed mainly in ECs [26] and mediates the uptake of pharmaceutical drugs to different tissues [40]. CD93 is a highly glycosylated transmembrane protein expressed in ECs and stem cells [41], and it has been previously studied in EC adhesion and migration and tumor angiogenesis.
In our study, 13 proteins have been reported to function in cell metabolism (CYP2U1, TBC1D2B, AKAP1, PSMD1, TMEM70, TMPRSS956) [42], tumor progression (CAV-2, TPM1), and stem cell differentiation (ST3GAL5, ST3GAL6 ATP2B4, CNOT1, RANBP17) [22,29,42 –50]. The functions of both C12orf76 and GFRA3 are currently unknown. However, the artemin-GFRA3 signaling system has a role in regulating early embryonic development and apoptosis [51]. The diverse roles of pericyte cell surface marker proteins demonstrate the importance of pericytes to several different systems in the body.
Recently, pericytes have emerged as tissue-specific progenitor cells, and traits of pericytes are different depending on their tissue of origin [52]. For example, previous studies have shown that in certain regions of the brain, pericytes are neural crest derivatives [53], whereas pericytes in other organs are mesoderm derivatives [54]. Further, neural versus non-neural pericytes have been suggested to have differences in abilities of differentiation. For example, recent studies identified that human cardiac pericytes express mesenchymal stem/stromal cell markers, including CD44, CD73, CD90, and CD105. These pericytes have the capacity to differentiate into osteo-, chondro-, and adipogenesis, but there is no potential for skeletal myogenesis in vitro [55]. On the other hand, in the central nervous system, pericytes play an essential role in the maturation and maintenance of the blood–brain barrier [56,57]. Our study demonstrates the tissue-intrinsic differences in human pericyte antigen expression. Comparing results from soft tissue and brain tissue in this study, some proteins such as CAV1, TPM1, and ENG (Fig. 4) showed specific staining in both brain and soft tissue. Other protein markers, such as CYP2U1, RNF112, and TMPRSS9, only showed specific staining in soft tissue but not in the brain.
Notably, the transcriptomic analysis of this study demonstrates large changes in pericyte gene expression before and after culture expansion. This is remarkable, since much of what we know about pericytes is based on in vitro culture experiments [58,59]. Gene expression profile changes after culture may be caused by a variety of reasons, including cell culture medium, stiffness and composition of the cell culture substrate, confluency, and cell cycle stage. Further investigation of these surface markers and their functions may aid in profiling the different conditions of stromal cells.
Several limitations to the study just mentioned exist. Some well-studied pericyte markers were not available for review within the currently available TMA, such as RGS5 and TBX18. Along these lines, the quality of each individual IHC antibody is another limitation. In some situations, when more than two antibodies were sometimes used to stain the same protein of interest, clear discrepancies were identified. However, this issue was mitigated by excluding any inconsistent or nonspecific staining. Unfortunately, some common pericyte antigens also were noted to have nonspecific staining within the TMA, including, for example, CD29 and NG2. To further mitigate any spurious results from TMA analysis, our cross-referencing with the transcriptome of FACS-derived pericytes identified a large number of novel antigens expressed on the cell membrane which could be used in the future to isolate or further partition human pericytes.
In sum, combined use of a human TMA dataset with the transcriptome of adipose tissue-derived human pericytes yielded 25 cell surface antigens, including 18 novel antigens expressed within human pericytes. Our study demonstrates the existence of novel pericyte markers, some of which are conserved in culture and that may be useful for future efforts to typify or isolate human pericytes.
Footnotes
Acknowledgments
A.W.J. was funded by NIH/NIAMS (R01 AR070773, K08 AR068316), NIH/NIDCR (R21 DE027922), USAMRAA (W81XWH-18-1-0121, W81XWH-18-1-0336, W81XWH-18-1-0613), the American Cancer Society (RSG-18-027-01-CSM), the Orthopedic Research and Education Foundation with funding provided by the Musculoskeletal Transplant Foundation, the Maryland Stem Cell Research Foundation, and the Musculoskeletal Transplant Foundation. The content is solely the responsibility of the authors and does not necessarily represent the official views of the National Institute of Health, Department of Defense, or US Army. The authors thank Dr. Marc Halushka for his assistance in HPASubC analysis.
Author Disclosure Statement
None of the authors state that they have conflicts of interest with this article.
Supplementary Material
Supplementary Table S1
Supplementary Table S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.