
Abstract
Bronchopulmonary dysplasia (BPD), the main complication of extreme prematurity, has lifelong consequences for lung health. Mesenchymal stromal cells (MSCs) prevent lung injury in experimental BPD in newborn rodents when given in the immediate neonatal period. Whether MSC therapy can restore normal lung growth after established lung injury in adulthood is clinically relevant, but currently unknown. Experimental BPD was achieved by exposing newborn rats to 95% O2 from postnatal days 4–14. Human umbilical cord-derived MSCs were intratracheally administered to rats (1 × 106cells/kg body weight) as a single dose at 3 or 6 months of age followed by assessment at 5 or 8 months of age, respectively. Lung alveolar structure and vessel density were histologically analyzed. O2-exposed rats exhibited persistent lung injury characterized by arrested alveolar growth with airspace enlargement and a lower vessel density at both 5 and 8 months of age compared with controls. Single-dose MSC treatment at 3 months partially attenuated O2-induced alveolar injury and restored vessel density at 5 months. Treatment with a single dose at 6 months did not attenuate alveolar injury or vessel density at 8 months. However, treatment with multiple MSC doses at 6, 6.5, 7, and 7.5 months significantly attenuated alveolar injury and improved vessel density at 8 months of age. Treatment of the adult BPD lung with MSCs has the potential to improve lung injury if administered in multiple doses or at an early stage of adulthood.
Introduction
Bronchopulmonary dysplasia (BPD), the chronic lung disease of prematurity that predominately affects preterm infants born less than 28 weeks of gestation [1], remains the most common complication of extreme prematurity [2]. Advances in perinatal care have modified the pattern of lung injury and enable the survival of infants born as early as 23–24 weeks of gestation. Interference with normal lung growth at these early stages results in dramatic impairment of alveolar and lung vascular growth. As a consequence, preterm survivors with BPD experience frequent hospital readmissions [3] and long-term respiratory complications that reach beyond childhood, including asthma, respiratory infections, low exercise capacity, early-onset emphysema, and pulmonary vascular disease [4 –9]. Clearly, novel therapies are needed to prevent neonatal lung injury and/or restore already established lung damage.
Exciting discoveries in stem cell biology over recent years have offered new insight into the pathogenesis of BPD and opened new therapeutic avenues. Within a decade, mesenchymal stromal cell (MSC) therapy has advanced from proof of concept [10,11] and pre-clinical experimental studies [12] to early phase clinical trials in preterm infants at risk for BPD [13 –15]. All these studies focused mainly on preventing and/or reducing lung injury in its early stages; however, it remains unknown whether MSCs can offer therapeutic benefits to the adult lung that was injured in neonatal life. This approach is relevant to ex-preterm adults who developed BPD as neonates and experience poor pulmonary outcome as adults [4], and it may also have implications for the treatment of adult lung diseases including emphysema and chronic obstructive pulmonary disease (COPD).
We have previously shown beneficial effects of MSCs administered within the first 3 weeks of life in an experimental animal model of BPD [11,16,17]. Here, using the same animal model of BPD, we investigate the effect of administering MSCs to the adult BPD lung to identify the window of opportunity for treating BPD with MSCs later in life.
Methods
All experimental procedures were approved by the University of Alberta Animal Care and Use Committee and the Human Research Ethics Board (Edmonton, Alberta, Canada).
Additional methods can be found in Supplementary Data.
Isolation and culture of human umbilical cord-derived-MSCs
Human umbilical cord-derived MSCs were isolated from the Wharton's Jelly of the UC of healthy, term pregnancies after parental consent, using enzymatic digestion as previously described [18]. Passage 2 MSCs displayed typical spindle-shaped morphology and fulfilled the minimal criteria of MSCs, including cell surface markers and tri-lineage differentiation potential in adipogenic, osteogenic, and chondrogenic assays. After characterization, MSCs were expanded and frozen in pentastarch-based cryomedium (6% HES 200/0.5, 5% DMSO) until use.
Animal model
Experimental BPD was achieved by exposing neonatal Sprague-Dawley rats (Charles River, QC, Canada) to hyperoxia (O2, 95% O2) from postnatal days 4–14 in sealed Plexiglas chambers (BioSpherix, Redfield, NY), as previously described [11,19]. After the hyperoxia exposure period, rats were then housed in room air (RA, 21% O2) until adulthood. Controls were only exposed to RA. Rats were grown to two different ages: postnatal months 3 and 6 before commencement of the experimental procedures (Fig. 1).
Experimental timeline. Newborn rats were exposed to hyperoxia (medium gray area) or RA (light gray area) from P4–14d, and they were then raised in RA until adulthood. MSCs were administered intra-tracheally (indicated by dotted line and syringe). Three different groups were assessed: group 1, MSC treatment at P3mo and assessment at P5mo; group 2, MSC treatment at P6mo and assessment at P8mo; group 3, MSC treatments at P6mo, P6.5mo, P7mo, and P7.5mo and assessment at P8mo. MSC, mesenchymal stromal cell; Pmo, postnatal month; RA, room air.
In vivo cell administration
Within each age group, rats were allocated into four groups: (1) RA control, (2) O2 control, (3) RA+MSCs, and (4) O2+MSCs. MSCs were administered at various time points via an intratracheal (i.t.) injection (1 × 106 cells per kg body weight in sterile phosphate-buffered saline; 100 μL). Rats were administered a single dose of MSCs at P3mo or P6mo followed by assessment at P5mo or P8mo, respectively (Fig. 1). In a separate experiment, we investigated the effects of a multiple-dose treatment regime consisting of i.t injections of MSCs at P6mo, P6.5mo, P7mo, and P7.5mo followed by assessment at P8mo (Fig. 1).
Lung morphometry and immunohistochemistry
The left lung was pressure-fixed in situ via the trachea with a zinc formalin solution at a constant pressure of 20 cm H2O, as previously described [20]. After tracheal ligation, the lung was removed and immersed in fixative overnight. Lungs were then processed, paraffin-embedded, sectioned at 5-μm thickness, and stained with Masson's Trichrome. Alveolar structures were quantified on a motorized microscope stage by measuring the mean linear intercept (MLI), as previously described [20], as well as the tissue and airspace fractions. Sections were also stained for von Willebrand Factor (vWF, 1:200, A0082; Dako Glostrup, Denmark) for the assessment of vessel density, as previously described [21]. All histological measurements were performed on coded slides to blind the observers to the treatment groups.
Statistical analysis
Values are expressed as the mean ± standard error of the mean. Intergroup comparisons were made by using an analysis of variance, with exposure (RA vs. O2) and treatment (no MSCs vs. MSCs) as factors. A value of P < 0.05 was considered statistically significant.
Results
Single-dose MSC treatment at P3mo, followed by assessment at P5mo
MSC treatment partially attenuates O2-induced lung injury
Exposure to O2 during the neonatal period results in persistent impairment in lung structure at P5mo, characterized by enlarged and simplified airspaces, compared with RA-exposed rats (Fig. 2). Treatment with a single dose of MSCs at P3mo partially attenuates lung injury in O2-exposed rats. Although both O2-exposed groups exhibited a significantly (P < 0.05) larger MLI compared with the RA-exposed groups, in O2-exposed rats treated with MSCs the MLI was decreased by ∼14% (P < 0.05) compared with in untreated O2-exposed rats (Fig. 2). Lung tissue fraction did not differ significantly between all groups (Fig. 2).
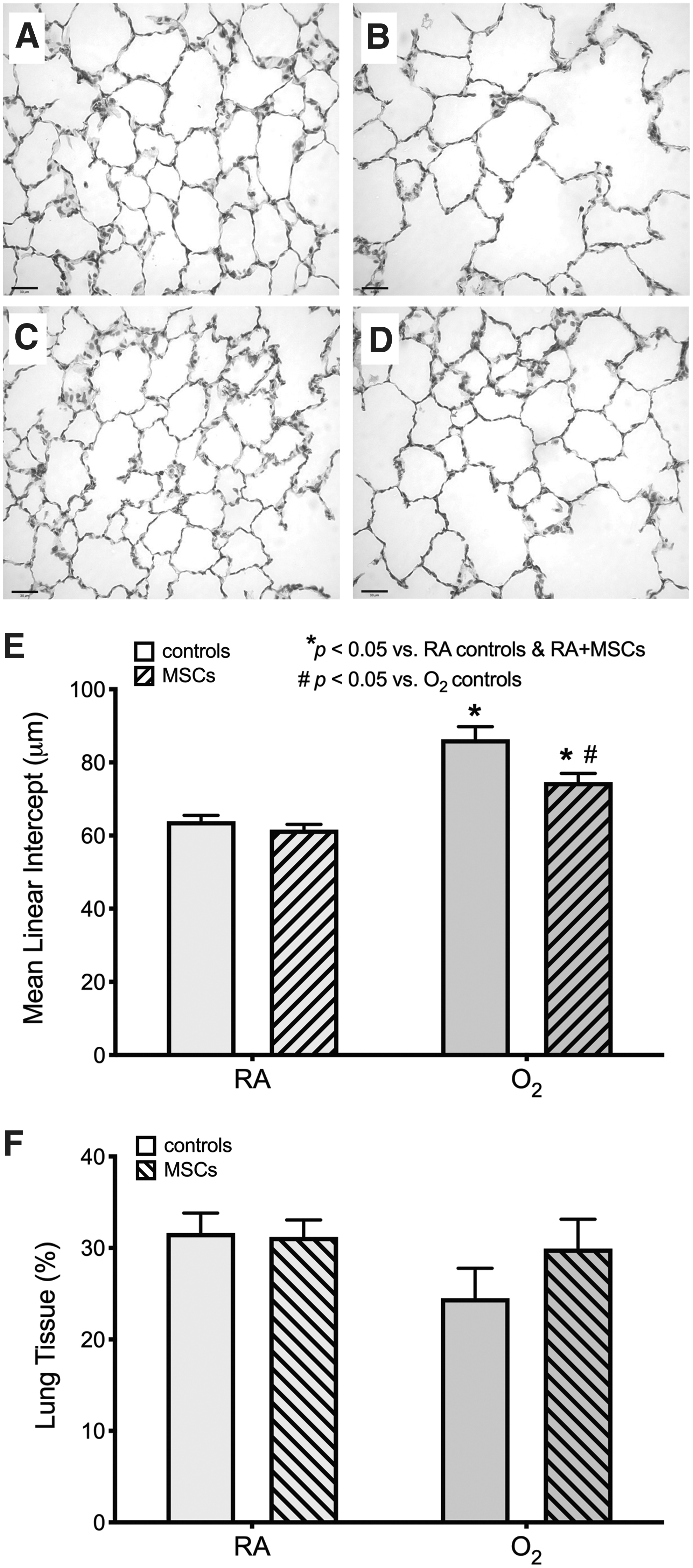
Treatment with a single dose of MSCs at P3mo partially improves O2-induced lung injury at P5mo. Representative photomicrographs of the lung structure at P5mo in a control RA-
Vessel density is improved in O2-exposed rats after MSC treatment
At P5mo, vessel density in O2-exposed rats was approximately half of that compared with RA-exposed rats (P < 0.05; Fig. 3). Treatment with MSCs at P3mo improved the vessel density in O2-exposed rats at P5mo to a similar level of the RA-exposed rats (both MSC-treated rats and controls); there was no significant difference in vessel density between RA-exposed rats and MSC-treated O2-exposed rats.
Treatment with a single dose of MSCs at P3mo restores vessel density in O2-exposed rats at P5mo. Representative photomicrographs of the vessel (arrows) density at P5mo in a control RA-
Blood oxygen saturation levels were measured before MSC treatment at P3mo, and then again at P5mo. Oxygen saturation levels were not affected by O2 exposure or MSC treatment (Supplementary Fig. S1). Assessment of lung function parameters at P5mo showed no significant difference between all groups, indicating that this particular rodent model of experimental BPD exhibits structural but not functional hallmarks of lung injury (Supplementary Fig. S1). There was no significant difference in MCP-1 concentrations between all groups (Supplementary Fig. S2). O2-exposed rats treated with MSCs had a significantly lower concentration of the inflammatory cytokine IL-1b compared with room air-exposed rats (P < 0.05) (Supplementary Fig. S2). O2-exposed rats treated with MSCs also had a significantly lower concentration of RGN compared with all other groups (P < 0.05) (Supplementary Fig. S2).
Single-dose MSC treatment at P6mo, followed by assessment at P8mo
MSC treatment does not attenuate O2-induced lung injury
Impairment in lung structure also persists at P8mo in O2-exposed rats (Fig. 4). Assessment of lung structure at P8mo, after a single dose of MSCs at P6mo, did not show any significant improvements in O2-exposed rats. Both untreated and MSC-treated O2-exposed rats exhibited a significantly (P < 0.05) greater MLI than both RA-exposed groups; MLI was approximately one-third larger (Fig. 4). In addition to an increased MLI, both O2-exposed groups had a significantly lower lung tissue fraction compared with the RA-exposed groups: ∼15% less parenchymal tissue (Fig. 4).
Treatment with a single dose of MSCs at P6mo does not improve O2-induced lung injury at P8mo. Representative photomicrographs of the lung structure at P8mo in a control RA-
MSC treatment does not improve O2-induced reduction in vessel density
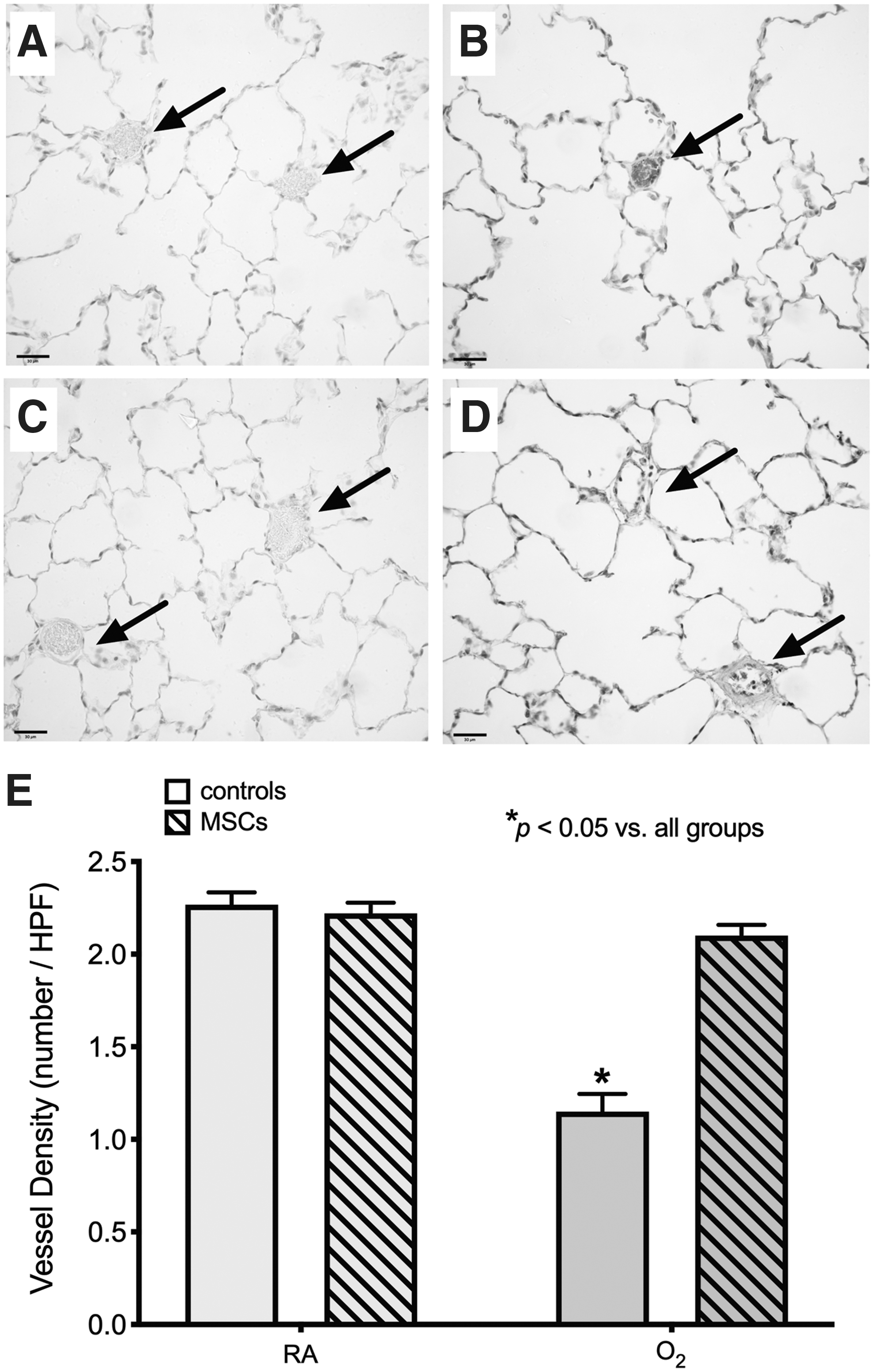
At P8mo, O2-exposed rats still exhibited a significant reduction in vessel density, by approximately one-third, compared with RA-exposed rats (P < 0.05; Fig. 5), demonstrating the persistent long-term effects of neonatal O2 exposure on angiogenesis. However, MSC treatment at P6mo did not provide any beneficial effects on vessel density in O2-exposed rats at P8mo; both O2-exposed groups displayed a significantly lower vessel density compared with RA-exposed rats (both MSC-treated rats and controls).
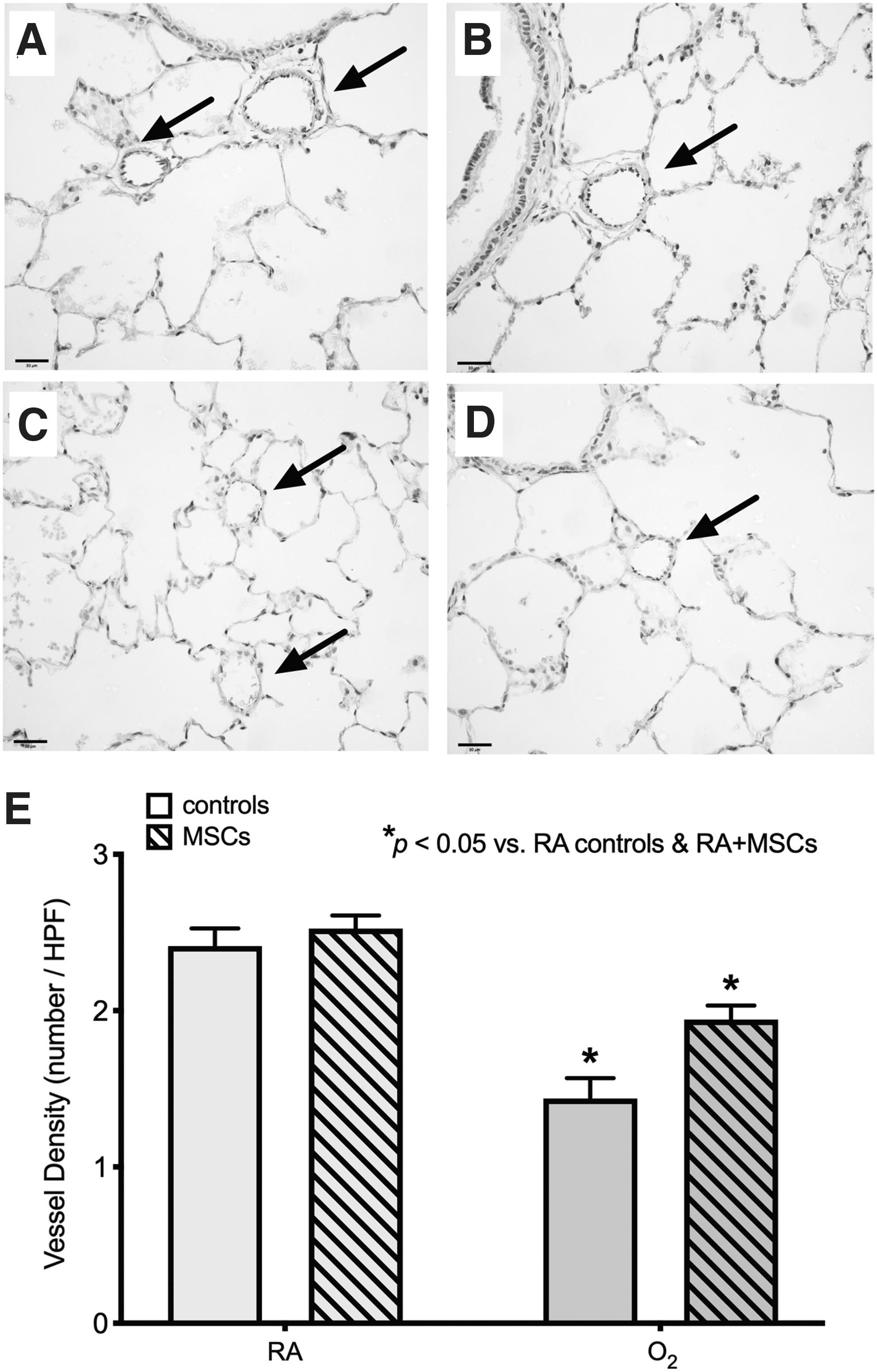
Treatment with a single dose of MSCs at P6mo does not restore vessel density in O2-exposed rats at P8mo. Representative photomicrographs of the vessel (arrows) density at P8mo in a control RA-
Blood oxygen saturation levels were measured before MSC treatment at P6mo, and then again at P8mo. Oxygen saturation levels were not affected by O2 exposure or MSC treatment (Supplementary Fig. S3). Lung function testing at P8mo did not show any significant difference in any of the lung function parameters between groups (Supplementary Fig. S3). There was no significant difference in MCP-1, RGN, VEGF, or FGF-basic concentrations between all groups (Supplementary Fig. S4).
Multiple-dose MSC treatment from P6–7.5mo, followed by assessment at P8mo
Multiple doses of MSCs attenuate O2-induced lung injury
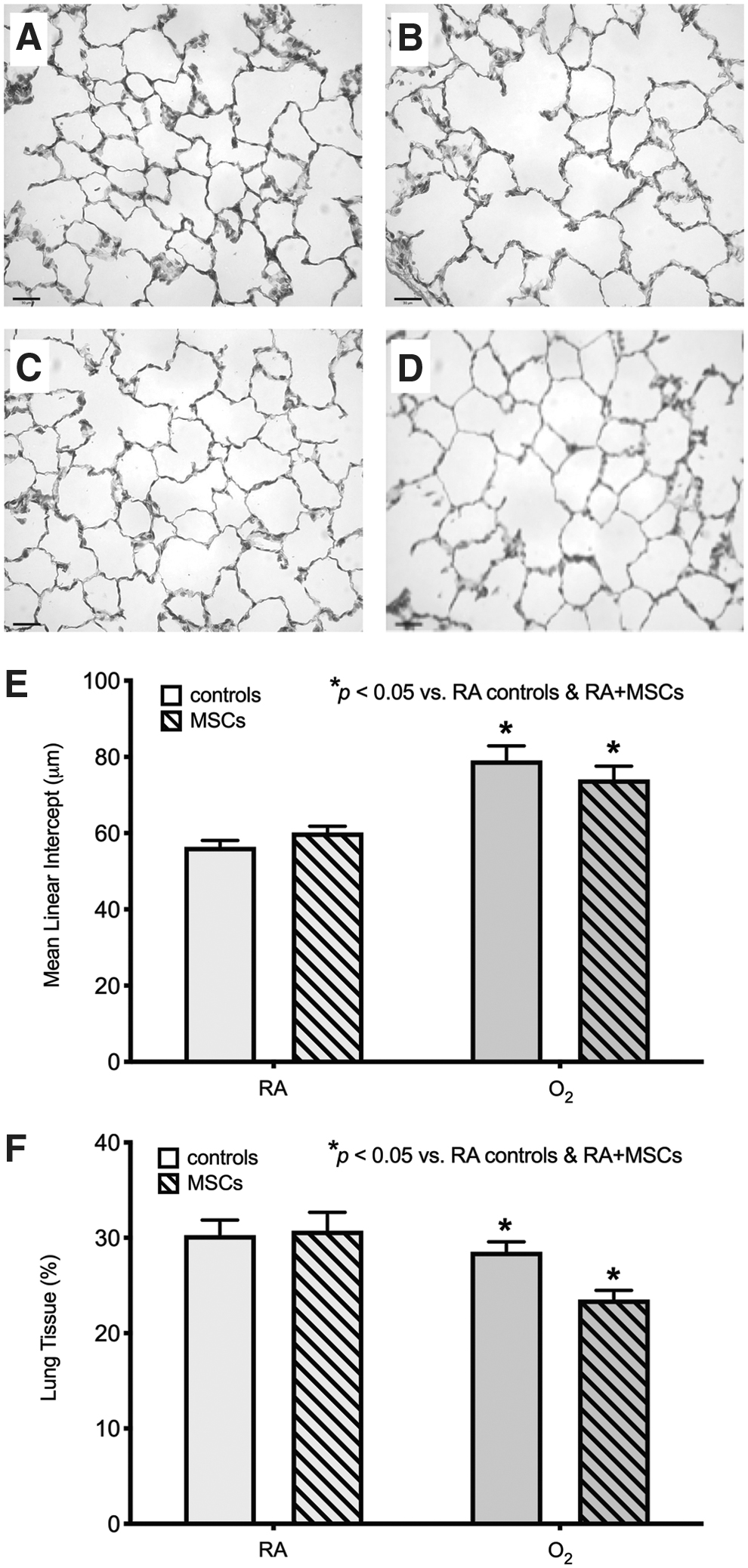
O2-exposed rats that were treated with multiple doses of MSCs from P6–7.5mo showed the greatest improvement in lung structure. There was no significant difference in MLI between MSC-treated O2-exposed rats and the RA-exposed groups. As expected, untreated O2-exposed rats had a significantly larger MLI, by ∼20%, indicating persistent lung injury after neonatal exposure to hyperoxic gas (Fig. 6). Even though untreated O2-exposed rats had a significant increase in MLI, their lung tissue fraction did not differ significantly from the other groups (Fig. 6).
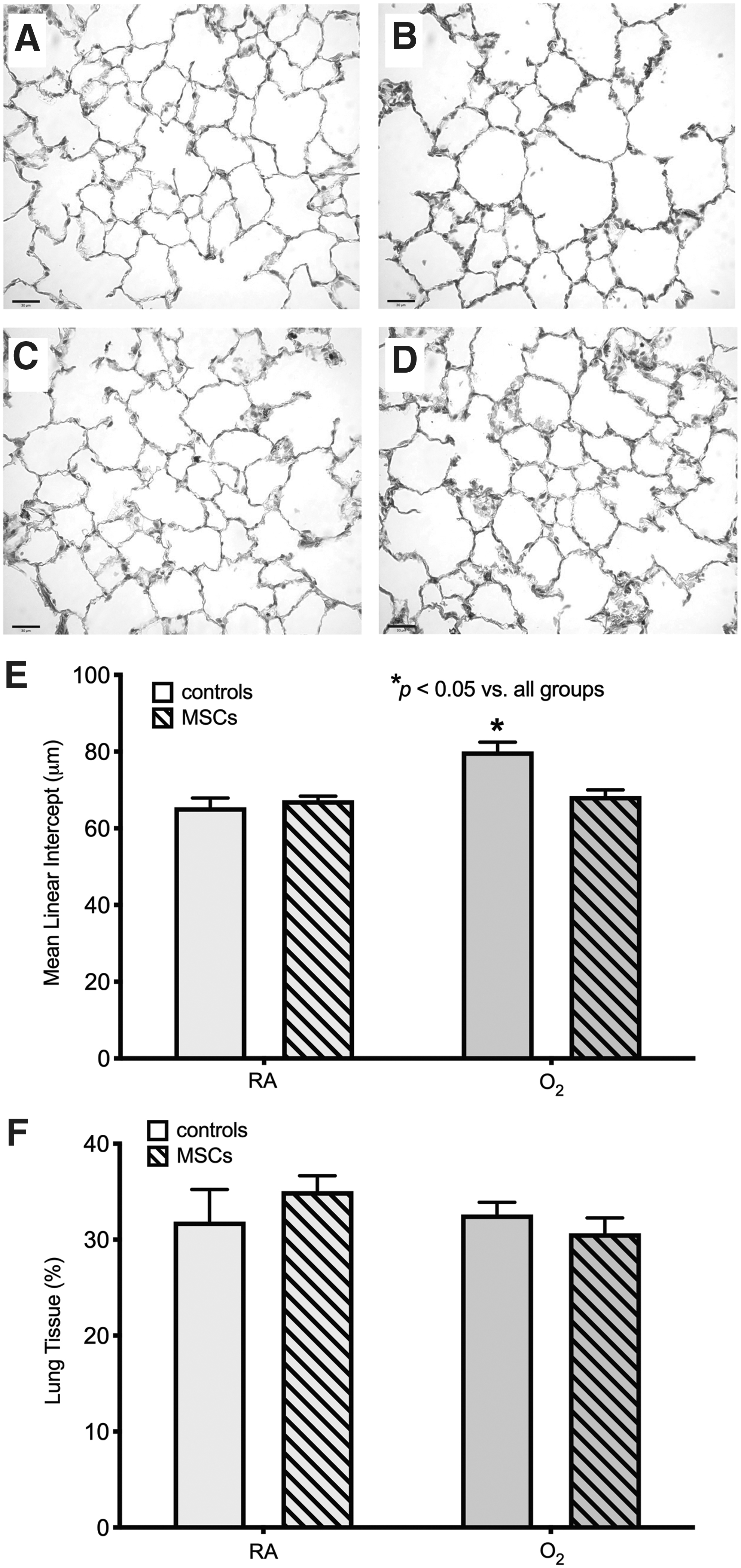
Treatment with multiple doses of MSCs from P6–7.5mo improves O2-induced lung injury at P8mo. Representative photomicrographs of the lung structure at P8mo in a control RA-
Multiple doses of MSCs improved vessel density in O2-exposed rats
As seen in the previous groups, O2 exposure during neonatal life resulted in a persistent reduction in vessel density into adulthood at P8mo. O2-exposed rats displayed almost half the vessel density of that of RA-exposed rats (P < 0.05; Fig. 7). However unlike the single dose of MSCs at P6mo, multiple doses of MSCs from P6–7.5mo significantly improved vessel density in O2-exposed rats, by ∼64%, and they had a significantly higher vessel density compared with untreated O2-exposed rats.
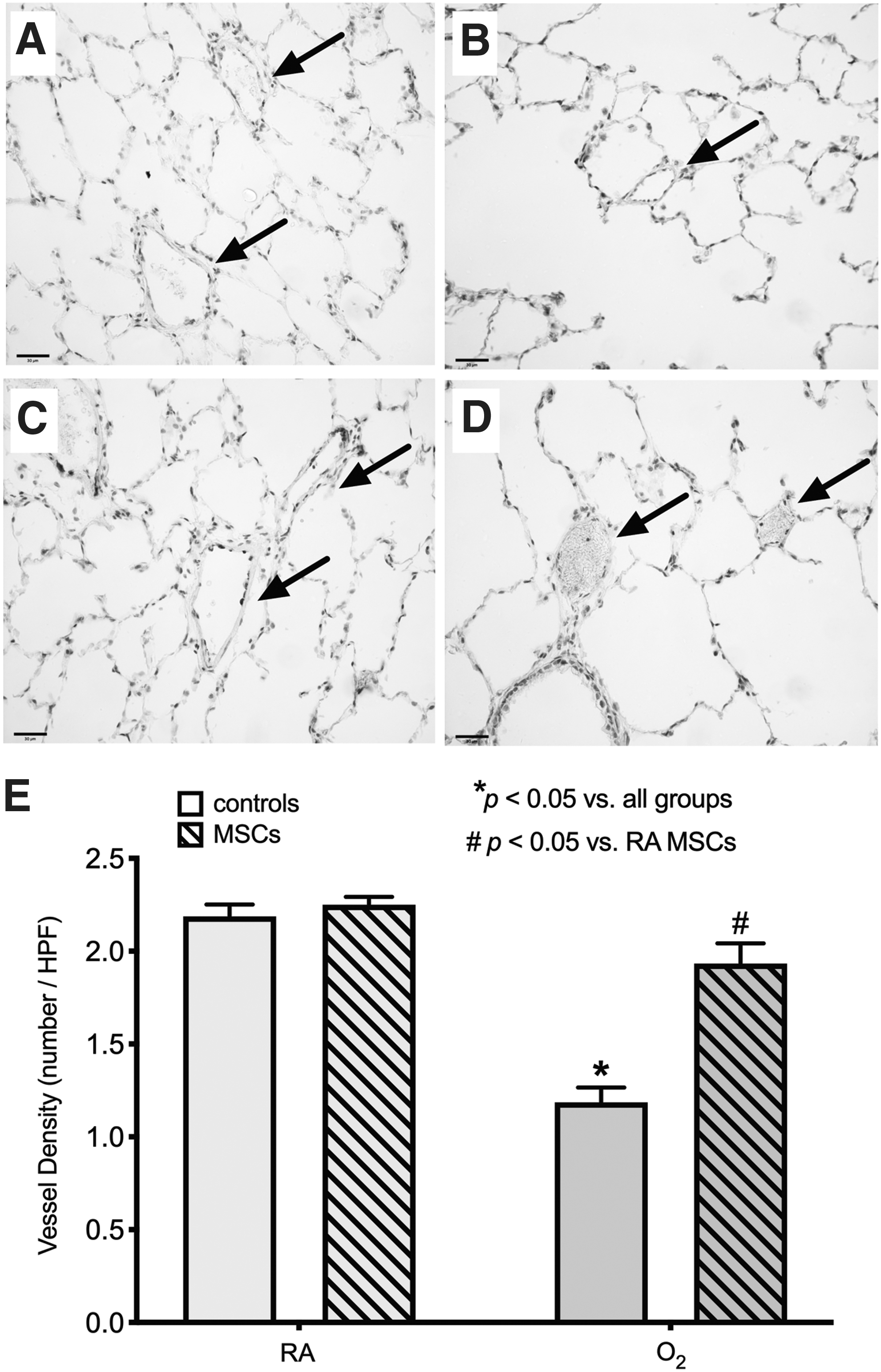
Treatment with multiple doses of MSCs from P6–7.5mo restores vessel density in O2-exposed rats at P8mo. Representative photomicrographs of the vessel (arrows) density at P8mo in a control RA-
Blood oxygen saturation levels were measured before the first MSC treatment at P6mo, and then again at P8mo. Oxygen saturation levels were not affected by O2 exposure or MSC treatment (Supplementary Fig. S5). Assessment of lung function parameters at P8mo showed no significant difference between all groups (Supplementary Fig. S5). There was no significant difference in MCP-1, RGN, VEGF, or FGF-basic concentrations between all groups (Supplementary Fig. S6).
Discussion
We demonstrate for the first time that treatment in adulthood, after the development of BPD, with MSCs has the potential to improve persistent lung injury if administered in multiple doses or at an early stage of adulthood. Specifically, we show that a single dose of MSCs administered at P3mo has the potential to partially attenuate lung injury and restore vessel density, whereas a single dose administered later in adulthood at P6mo does not provide therapeutic benefit. Conversely, repeated dosing from P6mo showed a more favorable outcome and attenuated both impaired alveolar structure and vessel density.
We harnessed the potential of the Wharton's Jelly due to its clinically relevant, easily accessible, ethically viable, and readily available source of MSCs [22,23]. MSCs are one of the most extensively examined cell types in experimental models of neonatal lung injury, and their therapeutic potential has already been translated into clinical trials [14,22]. Current preclinical experiments and clinical trials have focused on the administration of MSCs during the period of lung injury, which has proven to elicit favorable outcomes in terms of attenuation of lung injury and long-term efficacy. However, it is unknown whether MSC therapy can rescue the lung in adulthood from established lung injury if the ideal therapeutic window (ie, treatment coinciding or immediately after injury) has passed.
Here, we investigated the effects of administering MSCs at various time points in adulthood with the aim of providing insight into treating the adult BPD lung and at determining how wide the therapeutic window for neonatal lung disease extends. Our findings indicate that the adult lung can, to a certain extent, be rescued and that multiple doses of MSCs are more responsive to improving the lung architecture as compared with a single dose. These outcomes have significant clinical benefits, as the treatment of BPD with MSCs may now be extended past the neonatal period. Indeed, a prevention approach would be more favorable in the clinic as it could reduce or eliminate the impact that BPD has on preterm infants. However, since clinical trials have only recently commenced, the use of MSCs to treat BPD has not yet become standard practice in the Neonatal Intensive Care Unit (NICU). Thus, many preterm infants affected by BPD are being discharged from the NICU and are progressing into childhood and adulthood with an increased risk of lifelong pulmonary complications [4 –8]. Our findings may enable ex-preterm children and adults to still be provided with the opportunity to gain therapeutic benefit from MSCs. However, the need for several injections at 6 months of age in the rat model indicates that rejuvenating the adult lung presents a bigger challenge.
Our study focused on several ages in the rat, which can be correlated to age ranges in humans. Although it is difficult to compare a specific age in the rat with a specific age in the human, because many factors that determine development and maturity must be taken into consideration, in our study we considered 3 months in the rat to be approximately adolescence (late-teens) in humans, 5 months in the rat to be approximately early 20s in humans, 6 months in the rat to be approximately mid-20s in humans, and 8 months in the rat to be approximately late 20s to early 30s in humans.
Our assessment of rats at the different time points throughout their life demonstrates the persistent effect of being exposed to oxygen in the neonatal period. In our study, it is likely that the MSCs exerted their beneficial actions through paracrine-mediated mechanisms [24]. In particular, the beneficial effects demonstrated by improved vessel density indicate the possibility of MSCs to produce pro-angiogenic factors and possibly other lung growth promoting factors, which has been previously demonstrated [25]. Since lung angiogenesis and alveolar development are closely aligned [26], it is likely that a paracrine-mediated effect of MSCs on angiogenesis would also positively affect alveolar growth.
Repair of adult lung injury in experimental models of COPD and emphysema have been demonstrated by many studies utilizing MSCs (reviewed in [27,28]); in those studies, MSCs exerted their therapeutic benefit on structural repair and functional restoration in animal models of COPD/emphysema that were induced by various insults, including cigarette smoke, papain, elastase, and bleomycin. The proposed mechanism of action in adult lung injury is via paracrine and immunomodulatory effects that reduce lung inflammation and increase the expression of growth factors, rather than cell engraftment. It is likely that the approach to repairing the lung in ex-preterm adults will closely align to that of COPD/emphysema, especially since they already have an increased risk of early-onset emphysema and are often exposed to secondary respiratory challenges such as smoking and infection. Although inflammatory cell characterization was beyond the scope of this study, further studies investigating the lung inflammation and immunomodulatory effects of MSCs in the ex-preterm lung would provide a deeper understanding of MSC's mechanism of action.
Limitations
Our use of an adult rat model of BPD is a strength of this study, as it allows the investigation of MSC therapy at a later stage in life after the establishment of lung injury; however, several limitations should be considered. Although it is speculated that the MSCs exerted their beneficial actions through paracrine-mediated mechanisms, this study focused on descriptive histological outcomes and did not investigate any mechanistic approaches of the exhibited beneficial outcomes. In addition, even though it is possible for MSCs to use their paracrine-mediated effects to repair the lung, it remains unknown whether various aspects of cell regeneration, such as vascular, epithelial, and fibroblastic components, are, in fact, “normal” in the repaired lung. Further in-depth investigation into the mechanism of action of MSCs in adult BPD lung injury is warranted, as well as characterization of the repaired lung tissue components. Nevertheless, our findings are still relevant and very important to the field of preclinical studies for clinical applications to a growing population of adult patients born preterm with or without complications of BPD [4,9].
Conclusions
MSCs derived from the Wharton's Jelly of the umbilical cord show promise as a cell-based therapy for established lung injury, especially when given in multiple doses, and may open a new avenue to treating ex-preterm patients later in life.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by a grant from the Canadian Institutes for Health Research (CIHR; B.T.) and the University of Alberta “New Breath” Grant (B.T.). M.O. was the recipient of a Molly Towell Fellowship in Perinatal Research. B.T. was supported by a Canada Research Chair and Alberta Innovate Health Solutions (AIHS) clinical investigator award and by the Canadian Foundation for Innovation (CFI).
Supplementary Material
Supplementary Data
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Figure S5
Supplementary Figure S6
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.