
Abstract
Ischemia/reperfusion (I/R) injury remains a major problem in liver transplantation. I/R causes inflammatory cytokine release, apoptosis, and necrosis. Bone marrow-mesenchymal stem cells (BM-MSCs) can differentiate into hepatocytes in vivo, and differentiation further increases when hepatocytes are damaged. Exosomes are important mediators of cellular connections. Recently, exosomes of hepatocytes have been shown to play a pivotal role in inhibiting hepatocyte apoptosis and promoting hepatocyte regeneration. Therefore, we induced MSCs to differentiate into hepatocyte-like cells and extracted their exosomes; we then injected the exosomes into a mouse hepatic I/R model through the tail vein. Simultaneously, CoCl2 was used to mimic I/R in vitro. Our data indicated that in vivo, mesenchymal stem cell-derived hepatocyte-like cell exosomes (MSC-Heps-Exo) effectively relieve hepatic I/R damage, reduce hepatocyte apoptosis, and decrease liver enzyme levels. Consistent with the in vivo results, the in vitro experiments confirmed that exosomes effectively increased hepatocyte tolerance to ischemia and reduced hepatocyte apoptosis. We thus found that autophagy enhancement may be the mechanism by which exosomes protect the liver from I/R injury. These results indicate that exosomes play a protective role in hepatic I/R, and that the use of BM-MSCs for hepatocyte induction and exosome extraction may provide a new clinical treatment method through bioengineering.
Introduction
Hepatic ischemia/reperfusion (I/R) injury is a serious complication in surgical procedures, such as hepatectomy and orthotopic liver transplantation, and adversely affects patient and graft outcomes [1,2]. Impaired autophagy, mitochondrial dysfunction, and consequent damage to cellular homeostasis are the characteristics of hepatic I/R injury [3,4]. Although there in-depth studies have examined the mechanisms of hepatic I/R injury, no effective treatments are available for reducing or preventing liver I/R injury. Therefore, there is an urgent need to develop effective therapeutic strategies for I/R-induced liver injury.
There are many mechanisms for hepatic I/R injury, including oxidative stress, complement activation, inflammatory cell infiltration, inflammatory factor release, and apoptosis [4 –6]. Recent studies have shown that enhancement of autophagy can improve the activity and function of hepatocytes after I/R injury [7]. Autophagy has long been considered an adaptive response to cellular stress, where cellular stress is an adaptive means of preventing cell death. Under normal liver physiological conditions, basal levels of autophagy can degrade mitochondria and long-lived cytosolic and damaged proteins, to regulate hepatocyte lipid metabolism, immune responses, and cell death. When cells are subjected to metabolic stressors such as ischemia and hypoxia, autophagy is activated. Currently, limited details regarding the changes and effects of autophagy during liver I/R injury are available.
Exosomes are membrane-derived nanoscale vesicles (30–150 nm) that are released by many types of cells, including hepatocytes under normal and pathological conditions [8]. The effects of exosomes on cellular function have been linked to a variety of physiological processes, including intercellular communication, cancer metastasis, immunomodulatory activity, and the spread of infectious agents [9]. Recent studies have shown that hepatocyte-derived exosomes provide a synthetic mechanism for the formation of sphingosine-1-phosphate in target hepatocytes and accelerate cell proliferation and liver regeneration after I/R injury or partial hepatectomy [10]. Importantly, studies have shown that exosomes secreted by hepatocytes play an important role in the treatment of acute and chronic liver diseases.
However, difficulties in obtaining fresh isolated hepatocytes and cell cultures have hindered the application of this process [11]. Bone marrow mesenchymal stem cells (BM-MSCs) are easily obtained from the BM and can be differentiated into mesodermal cell lineages, such as osteoblasts, adipocytes, and chondrocytes [12,13]. However, it remains to be determined whether mesenchymal stem cell-derived hepatocyte-like cell exosomes (MSC-Heps-Exo) can reduce hepatic I/R injury and whether they are efficacious in activating autophagy. Therefore, this work aims to explore the potential therapeutic effects in an established mouse model of 70% hepatic I/R injury, and to characterize this potential effect in vitro using a liver cell hypoxia model with the chemical reagent, cobalt chloride.
Materials and Methods
Animals
C57BL/6 mice, male, weighing 22 ± 2 g, were purchased from Beijing HFK Bioscience Co., Ltd. (Beijing, China) and were maintained in the animal facility of the Institute of Organ Transplantation, Tongji Hospital, Tongji Medical College, Huazhong University of Science and Technology. All procedures involving animal use in this study were performed and monitored in accordance with the guidelines of the Chinese Council on Animal Care, and were approved by the Institutional Animal Care and Use Committee of the Tongji Medical College, Huazhong University of Science and Technology. Throughout the study, all efforts were made to minimize any suffering of the animals.
Isolation, culture, and hepatic differentiation of BM-MSCs
BM-MSCs were obtained from C57BL/6 mice and cultured as previously described [13]. Identification of primary MSCs was confirmed by morphology, phenotypic analysis, and ability to differentiate into adipocytes and osteoblasts [13]. MSCs were also capable of differentiating into chondrocytes. The protocol of chondrocyte differentiation and Alcian Blue staining was followed as per the Mouse Mesenchymal Stem Cell Chondrogenic Differentiation Medium Kit instructions (MUBMX-90042, Cyagen, Guangzhou, China). MSCs of passage 3 were treated with 10 ng/mL fibroblast growth factor-4 (FGF-4), 20 ng/mL hepatocyte growth factor (HGF), 10 ng/mL oncostatin M (OSM), 1 × insulin–transferrin–sodium selenite (ITS), and 20 μg/L dexamethasone (Dex) at specific times, as presented in Fig. 2A.
After differentiation, the hepatocyte-like cells were identified by morphological observation and immunofluorescence staining with specific antibodies against alpha fetoprotein (AFP) and albumin (ALB). Primary antibodies to AFP (diluted at 1:80) and ALB (diluted at 1:100) were added to the tissue sections and the sections were incubated overnight in a humid box at 4°C. After rinsing thrice with phosphate-buffered saline (PBS) (pH 7.4) (5 min each), the samples were incubated with fluorescence-tagged secondary antibodies (diluted at 1:100) in the dark in the humid box at 37°C for 1 h. Tissue sections were mounted in buffered glycerol, viewed under a fluorescence microscope, and photographed. Furthermore, hepatic nuclear factor-3β (HNF-3β) expression was analyzed by quantitative reverse transcription/polymerase chain reaction (qRT-PCR). Gene expression was analyzed using a StepOnePlus Real-Time PCR System and relative gene expression was calculated by the 2-ΔΔCT method using GAPDH as the reference gene. The primer sequences used for the experiment are as follows: HNF-3β forward: GGCAACATGTTCGAGAACGG, reverse: GGTCTTCTTGCCTCCGCTAC; GAPDH forward: AGGTCGGTGTGAACGGATTTG, reverse: TGTAGACCATGTAGTTGAGGTCA.
Exosome isolation and characterization
Exosomes were isolated from BM-MSC-derived hepatocyte-like cells using the Total Exosome Isolation Kit (Invitrogen, California). Total exosome isolation reagent (0.5 mL) was added to each 1 mL of filtered conditioned medium and was mixed well by inverting. The mixture was centrifuged at 12,000g for 70 min at 4°C after overnight incubation at 4°C. All the supernatant was then removed by aspiration and exosome pellets were resuspended in a convenient volume of PBS. Exosome size and concentration were determined using a Zetasizer Nano (Malvern Instruments, Malvern) and electron microscopy. Exosome markers, including CD9, CD63, and TSG101, were measured by western blot analysis.
Hepatic I/R injury model
Hepatic I/R injury was modeled using 70% hepatic I/R animals. The mice were anesthetized with 60 mg/kg of sodium pentobarbital by intraperitoneal injection. Then, the artery and portal vein were interrupted using a noninvasive clip for 60 min to prevent blood flow to the left and middle lobes of the liver. During the operation, the mice were maintained at 37°C with a warming pad and incubator. After 6 h of reperfusion, the mice were sacrificed for blood and liver tissue collection. Mice in the I/R-Exo group were injected with 100 μg MSC-Heps-Exo before and after the operation through a tail vein. In the I/R group, an equal volume of saline was injected at the same time point. The control group only underwent laparotomy without clamping the blood vessel.
Hepatocyte hypoxia model
The cell hypoxia model was established using the chemical reagent, cobalt chloride (CoCl2; Sigma, St. Louis, MI). The mouse hepatocyte cell line AML12, was used in this experiment. For cell propagation, the cell line was cultured in DMEM/F12 (HyClone, Logan City, UT) and supplemented with 10% fetal bovine serum (FBS; Gibco, Grand Island, NY), ITS liquid media supplement (Sigma), and 40 ng/mL Dex (Sigma). The cells were maintained in a humidified incubator at 37°C and 5% CO2. In the cell hypoxia model, AML12 cells were cultured in FBS-free DMEM/F12 with 200 μM CoCl2 for 24 h when the cell confluence reached 60%–70%.
Biochemical analysis
Serum alanine aminotransferase (ALT) and aspartate aminotransferase (AST) levels were measured using a standard clinical automatic analyzer in the clinical laboratory of Tongji Hospital, Tongji Medical College, Huazhong University of Science and Technology.
Pathological analysis
Fresh liver tissues were fixed in 4% paraformaldehyde for paraffin embedding. Then, the embedded tissues were sectioned at 5 μm and stained with Hematoxylin and Eosin (HE). Suzuki's histological grading was performed to evaluate any histopathological damage. The assessment was based on three aspects, including sinusoidal congestion, hepatocyte necrosis, and ballooning degeneration. Each criterion was scored on a scale of 0 to 4 depending on the degree of liver damage. In the absence of any congestion, vacuolization, or necrosis, the best score of 0 was assigned. Severe congestion accompanied with severe vacuolization and >60% necrosis was assigned a score up to 12. Histological changes were evaluated in randomly chosen fields at 200 × magnification.
TdT-mediated dUTP Nick-End Labeling (TUNEL) assay was used to detect apoptosis in the paraffin-embedded liver sections. The assay was performed according to the Cell Death Detection Kit (Roche, Basel, Switzerland) instructions, and the sections were observed under a microscope (NIKON ECLIPSE CI, Tokyo, Japan). The number of TUNEL-positive nuclei was counted in six randomly chosen images from nonoverlapping areas of each group. The data are presented as the percentage of TUNEL-positive cells.
Cell viability assay
Cell viability was detected using a CCK-8 Kit (Dojindo, Japan). Cells were plated in 96-well plates at a density of 5,000 cells/well and incubated for 24 h. Then, cells were subjected to hypoxia treatment as described above and 10 μL of CCK-8 solution was then added to each well. The cells were incubated for another 100 min at 37°C. Finally, absorbance values were determined at 450 nm using a microplate reader.
Flow cytometric analysis for cell apoptosis
Apoptotic cells were stained with the Annexin V-FITC and the Propidium Iodide (PI) Staining Kit (MultiSciences, Hangzhou, China) according to the manufacturer's instructions. AML12 cells were suspended and centrifuged at 300g for 5 min, followed by incubation with Annexin V-FITC and PI for 5 min in the dark. The apoptotic cells were quantified (%) by flow cytometry on a FACSCalibur system (BD Biosciences), and FlowJo software (Oregon) was used for data analysis.
Western blot analysis
Protein content was determined using the Bradford method for the standardization of bovine serum albumin (Sigma). Sodium dodecyl sulfate/polyacrylamide gel electrophoresis was used to isolate the protein samples and the samples were then transferred to PVDF membranes (Millipore). Tris-buffered saline with Tween containing 5% bovine serum albumin, was used to block the nonspecific binding sites for 1 h. The membranes were incubated overnight at 4°C with primary antibodies against β-actin (1:100,000 dilution), p62/SQSTM1 (1:1,000), beclin-1 (1:1,000), LC3 (1:1,000), and cleaved caspase-3 (1:1,000) (Cell Signaling Technology, Beverly, MA). The membrane was then incubated with the secondary antibody at room temperature for 1 h; the signal was then washed and detected. Protein expression levels were quantified using Image-Pro Plus software (Media Cybernetics).
Electron microscopy
Exosomes in PBS were immobilized in 1.5 M sodium dimethyl arsenate buffer (pH 7.4), adsorbed onto a copper mesh (Electron microscopy Sciences, Hatfield, PA), and stained with 2% uranyl acetate. Liver tissue was fixed with 2.5% glutaraldehyde, cut into ultrathin sections, and stained with uranium acetate and lead citrate. Samples were observed and images were taken using an H7700 transmission electron microscope (Hitachi, Tokyo, Japan) for data acquisition. The number of exosomes and autophagosomes in each sample was counted randomly at the same magnification.
Statistical analysis
All data are shown as the mean ± standard error of the mean (SEM). Comparisons between groups were performed through the two-tailed Student's t-test. A P < 0.05 was considered statistically significant.
Results
Isolation and identification of BM-MSCs
MSCs obtained from the BM of C57BL/6 mice exhibited a spindle-shaped morphology and their identity was confirmed by flow cytometry. The results showed that BM-MSCs have a typical MSC phenotype, positive for CD29, CD44, CD105, and Sca-1, and negative for CD11b, CD31, CD45, and CD117. Isotype controls are shown as shaded curves in Fig. 1A. In addition to the surface marker phenotype, MSCs also differentiated into adipocytes (Fig. 1B), osteoblasts (Fig. 1C), and chondrocytes (Fig. 1D) in vitro. These evidences suggested that typical MSCs were obtained from the BM of mice.

Identification of MSCs.
Hepatic differentiation and exosome isolation
Differentiation of BM-MSCs into hepatocyte-like cells in vitro was performed according to the protocol shown in Fig. 2A. Cytokines were added to the medium in the following order: FGF-4 was added on day 0, HGF on day 3, and HGF, OSM, ITS, and Dex on day 6. On day 21, hepatocyte-like epithelioid cells were observed (Fig. 2B). The expressions of liver-specific protein markers AFP and ALB were analyzed by immunofluorescence. In addition, the hepatocyte-characterized nuclear factor HNF-3β, was assayed by qRT-PCR. We found that the HNF-3β level in induced hepatocyte-like cells was lower than that in the AML12 hepatocyte cells, but was significantly higher than that in the MSCs (Fig. 2C, **P < 0.01). As shown in Fig. 2D, much higher AFP and ALB expression levels were observed in the hepatocyte-like cells induced from BM-MSCs than from normal BM-MSCs. These results showed that BM-MSCs differentiated into hepatocyte-like cells.

Acquisition and identification of hepatocyte exosomes induced by BM-MSCs.
Exosomes released into the culture medium were isolated for biophysical and biochemical analyses. The expectant cup shape was revealed by electron microscopy analysis of exosomes (Fig. 2E). Quantitative determination of exosome size and concentration through Zetasizer Nano showed that the average vesicle diameter was 144 nm and the concentration was 4.9 × 108 particles/mL (Fig. 2F). In addition, western blot was performed to validate the exosome markers. The results showed that CD9, CD63, and TSG101 were highly expressed in the exosomes of hepatocyte-like cells. All these analyses were in agreement with previously described features of hepatocyte exosomes [10].
MSC-Heps-Exo reduce hepatic I/R injury in vivo and in vitro
To investigate the effect of MSC-Heps-Exo in hepatic I/R injury, histological changes and serum liver enzyme levels were examined. Histologically, HE staining of liver tissues for the I/R group showed a large amount of hemorrhage and necrosis, and the structure of the liver lobe was severely damaged (Fig. 3A), whereas the liver damage was significantly lighter in the exosome-treated group (Fig. 3B, C). Similarly, a large apoptotic region (Fig. 3D) was found in the I/R control group following TUNEL staining, whereas apoptosis was greatly reduced in the exosome-treated group (Fig. 3E, F). In addition, the level of serum AST and ALT after 6 h was significantly lower in hepatic I/R mice treated with exosomes than in the I/R control group. The level of ALT and AST in the exosome-treated group was less than one-fifth of that in the I/R control group (Fig. 3G). Thus, a large number of hepatocytes survived during the liver I/R injury due to the MSC-Heps-Exo injection.
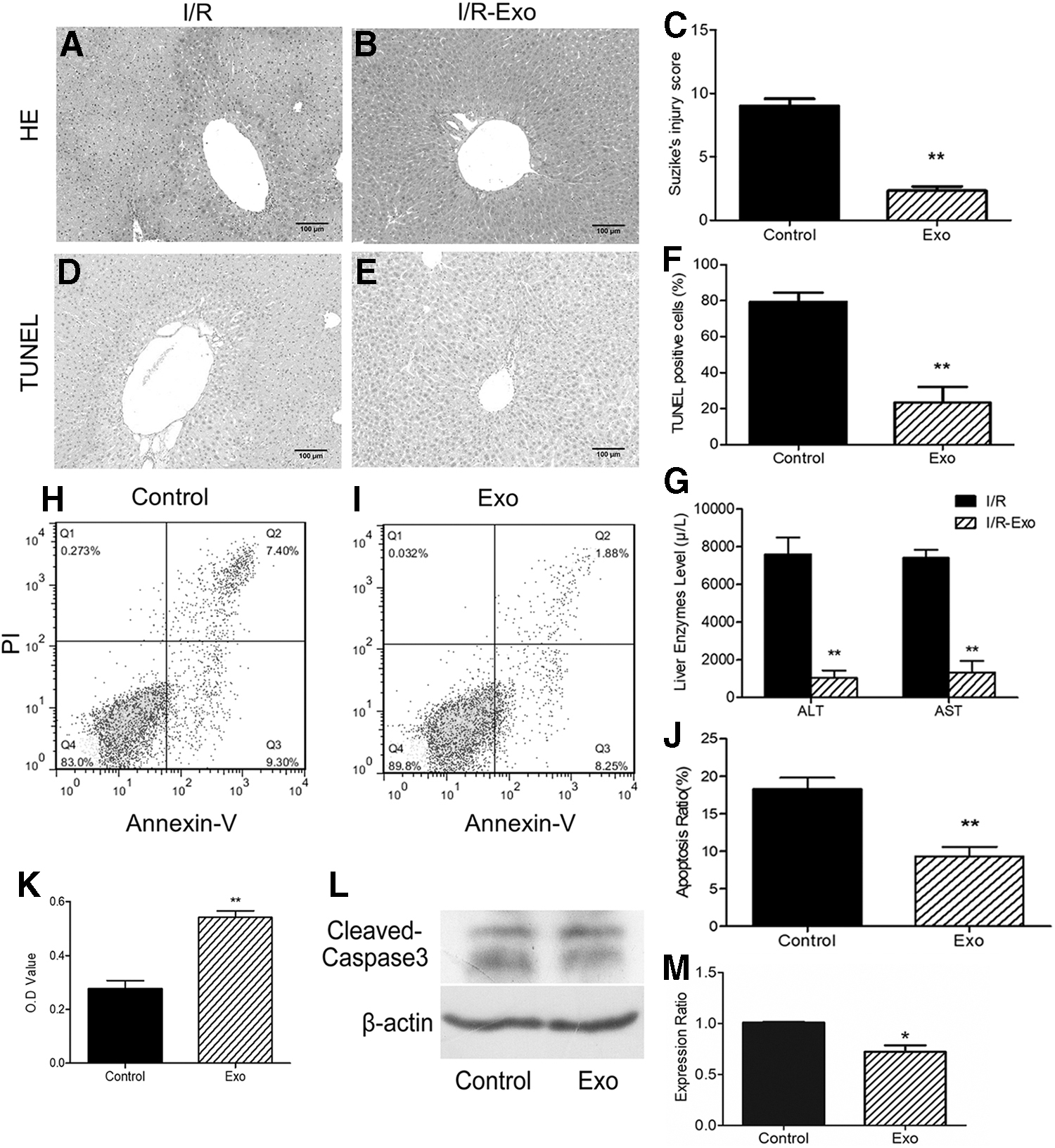
MSC-Heps-Exo reduces hepatic I/R injury in vitro and in vivo
In vitro models were subsequently designed to validate the protective effect of exosomes in vivo. Since CoCl2 can be used as a hypoxia simulator, we treated AML12 cells with CoCl2 to establish a hypoxic model in vitro. The CCK-8 cell viability assay showed that AML12 cell viability in the exosome-treated group was significantly higher (almost double) than that in the I/R control group (Fig. 3K). Using flow cytometric analysis, we also observed that apoptosis in the exosome-treated cells was lower than that in the I/R control group; especially, late-stage apoptotic cells (Annexin-V+/PI+) were greatly reduced after exosome treatment (Fig. 3H–J). In addition, the level of the apoptosis protein, cleaved caspase-3, was decreased in the exosome treatment group (Fig. 3L, M). These in vitro results also demonstrated that MSC-Heps-Exo treatment can improve the tolerance of hepatocytes to hypoxia.
MSC-Heps-Exo enhance liver autophagy during hepatic I/R injury
Autophagy has recently been found as an important protective measure that prevents I/R damage by removing damaged mitochondria, thereby inhibiting excessive ROS production [14]. Intolerance to I/R damage is poor in the aged liver due to autophagy damage, which promotes mitochondrial permeability transition and cell death [15]. We then attempted to investigate whether MSC-Heps-Exo affects autophagy during liver I/R injury. As shown in Fig. 4A and B, the levels of LC3-II, a standard indicator of autophagy activity, were increased in the MSC-Heps-Exo-treated I/R mice compared with those in I/R control mice. Additionally, p62/SQSTM1, which is degraded during the autophagic process, was decreased after MSC-Heps-Exo treatment. Moreover, changes in Beclin-1, a component of the PI3K complex that is required for autophagy, were similar to those in LC3-II (Fig. 4A, B). Autophagy is a highly dynamic and multistep process. TEM analysis showed that autophagy was increased in the liver of mice treated with exosomes compared with that in the I/R control mice (Fig. 4C–E). These results suggest that MSC-Heps-Exo treatment enhances autophagy during hepatic I/R injury.
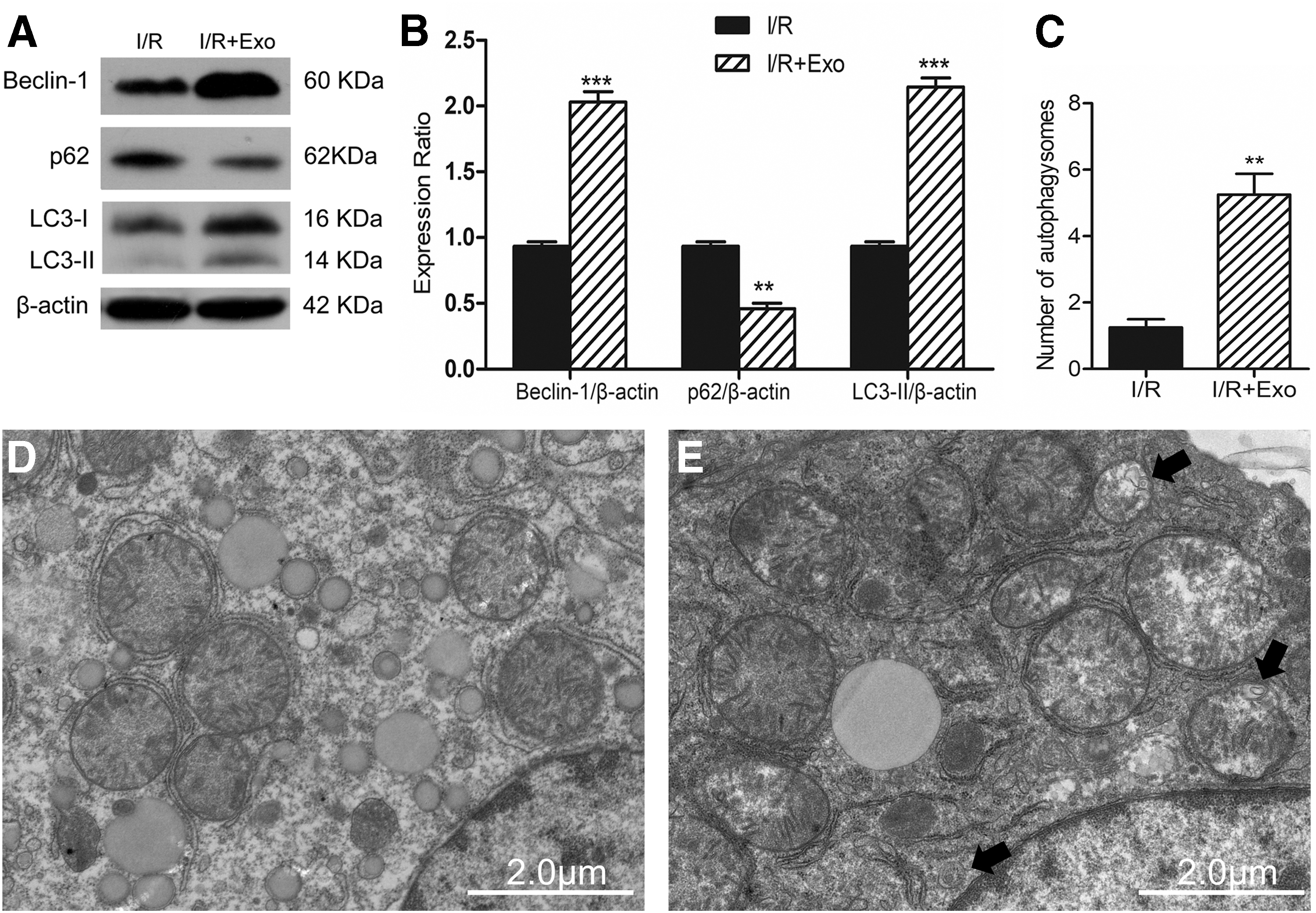
MSC-Heps-Exo pretreatment enhances liver autophagy in hepatic I/R injury.
Discussion
In our study, we found that the exosomes of hepatocyte-like cells induced from BM-MSCs can reduce liver I/R injury in vivo and inhibit damage to the hypoxic culture of hepatocytes in vitro. Moreover, exosomes of hepatocyte-like cells induced from MSCs increased autophagy in liver tissues in vivo and in hepatocytes in vitro.
Exosomes play a series of regulatory roles in cells. Recent studies have shown that exosomes exert protective effects on tissue I/R injury. Exosomes can reduce the extent of myocardial infarction and improve renal I/R injury by inhibiting inflammatory responses, reducing oxidative stress, inhibiting fibrosis, and promoting angiogenesis [16 –19]. Recent studies have also found that hepatocytes can secrete exosomes and play an important role in hepatocyte regeneration and repair [10]. The advantages of the clinical application of exosomes are enormous compared with those of cell therapy. By intravenous infusion of cells, most cells stick to alveolar capillaries and the host survival rate is low [20]. However, exosomes are not associated with problems such as cell embolism or stem cell differentiation. In addition, exosomes can be better protected than cell therapy by enriching some proteins and RNA [21].
However, there are still many limitations in the acquisition and in vitro culture of primary hepatocytes. BM-MSCs have multidirectional differentiation potential and can differentiate into a variety of cell types, including hepatocytes [12]. Importantly, mesenchymal liver cells are more readily available for cultivation in vitro. Previous studies have found that in the animal and human livers, the proportion of BM-MSCs that differentiate into hepatocytes varies with the degree of liver damage [22,23]. Differentiation of BM-MSCs into hepatocytes in vitro is a physiological process consistent with natural body processes and can thus be used as compensation for the damaged liver.
Therefore, we collected exosomes from the culture supernatant of MSC-derived hepatocyte-like cells and verified their effects on in vivo and in vitro models of hepatic I/R injury. Pathological results showed that I/R injury caused hepatocyte degeneration and necrosis, destruction of hepatic lobule structure, inflammatory cell infiltration, hepatic sinus congestion, swelling, and disordered arrangement. After exosome injection, the water-like degeneration area of hepatocytes in the I/R+Exo group, necrotic area, inflammatory cell infiltration, and sinusoidal swelling were all decreased. ALT and AST, the most common indicators of liver function, entered the blood in large amounts after liver cell damage, and continued to increase significantly. After exosome injection, ALT and AST expression was significantly lower in the I/R+Exo group than that in the I/R group, suggesting that exosomes can reduce hepatocyte necrosis and improve liver function. Additionally, in a CoCl2-induced hepatocyte hypoxia model, exosomes increased the hepatocyte viability after cell hypoxia injury.
Hepatic I/R injury involves a range of mechanisms, including metabolic acidosis, apoptosis, inflammatory responses, and oxidative stress [4 –6]. Our study showed that exosomes play an antiapoptotic role in the CoCl2-induced hepatocyte hypoxia model. Additionally, recent evidence suggests that autophagy can digest cytoplasmic material to produce the necessary metabolic substrates and energy to keep cells alive as a response to stress conditions such as I/R [24,25]. In our study, we also found that exosomes can enhance autophagy during hepatic I/R injury. At present, some researchers have proposed that enhanced autophagy may be a novel approach to improve hepatocyte viability and function following I/R injury, and that the protective effect of autophagy may be related to its antiapoptotic and anti-inflammatory activities [26].
Our research provides an innovative approach to the clinical treatment of hepatocyte injury, especially after liver transplantation, and expands the application of stem cells through exosomes. Many studies related to exosomes derived from stem cells have shown that they have similar functions as stem cells, such as regulating the body's immune function and promoting the regeneration of damaged tissues; these cells are thus considered to have great research potential in regenerative medicine. However, only a few studies have examined the exosome function of specific cell types differentiated from stem cells. Our results can thus be used as a bioengineering tool to explore the acquisition of hepatocytes along with other more robust applications. However, the specific components of exosomes that protect liver I/R injury and methods to obtain exosomes more efficiently still deserve further study.
Footnotes
Author Disclosure Statement
The authors declare no conflicts of interest.
Funding Information
This work was supported by funds from the National Natural Science Foundation of China (grant no. 81770652) and the Natural Science Foundation of Hubei Province (grant nos. 2017ACA096 and 2017CFB748). The authors gratefully acknowledge these funding sources.