
Abstract
Marsupials have long attracted scientific interest because of their unique biological features and their position in mammalian evolution. Mesenchymal stem cells (MSCs) are of considerable research interest in translational medicine due to their immunomodulatory, anti-inflammatory, and regenerative properties. MSCs have been harvested from various tissues in numerous eutherian species; however, there are no descriptions of MSCs derived from a marsupial. In this study, we have generated Tasmanian devil (Sarcophilus harrisii) MSCs from devil induced pluripotent stem cells (iPSCs), thus providing an unlimited source of devil MSCs and circumventing the need to harvest tissues from live animals. Devil iPSCs were differentiated into MSCs (iMSCs) through both embryoid body formation assays (EB-iMSCs) and through inhibition of the transforming growth factor beta/activin signaling pathway (SB-iMSCs). Both EB-iMSCs and SB-iMSCs are highly proliferative and express the MSC-specific surface proteins CD73, CD90, and CD105, in addition to the pluripotency transcription factors OCT4/POU5F1, SOX2, and NANOG. Expression of the marsupial pluripotency factor POU5F3, a paralogue of OCT4/POU5F1, is significantly reduced in association with the transition from pluripotency to multipotency. Devil iMSCs readily differentiate along the adipogenic, osteogenic, and chondrogenic pathways in vitro, confirming their trilineage differentiation potential. Importantly, in vitro teratoma assays confirmed their multipotency, rather than pluripotency, since the iMSCs only formed derivatives of the mesodermal germ layer. Devil iMSCs show a tropism toward medium conditioned by devil facial tumor cells and express a range of immunomodulatory and anti-inflammatory factors. Therefore, devil iMSCs will be a valuable tool for further studies on marsupial biology and may facilitate the development of an MSC-based treatment strategy against Devil Facial Tumor Disease.
Introduction
Mesenchymal stem cells (MSCs), also known as mesenchymal stromal cells, represent a heterogeneous population of plastic-adherent, fibroblast-like cells that are capable of self-renewal and multipotent differentiation into adipocytes, osteoblasts, and chondroblasts [1]. MSCs continue to attract research interest in the development of cell-based therapies, particularly for degenerative diseases such as osteoarthritis and degenerative disc disease [2,3]. However, MSCs have also been shown to secrete a wide variety of immunomodulatory, anti-inflammatory, and angiogenic factors, and so their potential as a novel therapeutic for a range of immune-mediated and inflammatory disorders is also attracting interest [4,5]. A further important consideration is that MSCs appear to be immune evasive, which permits the use of allogeneic MSCs for therapy, circumventing the need to harvest MSCs directly from the individual being treated [6].
Tissue-derived MSCs have been isolated and characterized from numerous eutherian species [7]. However, when harvested from older donors, their limited proliferative ability, rapid decline of differentiation potential, and the variability in quality across different donors hinder their therapeutic or research utility. Moreover, MSC-based therapies require large numbers of highly proliferative cells, which are relatively homogenous in culture and that will readily differentiate, effectively limiting the donor source to young individuals. Therefore, induced pluripotent stem cell (iPSC)-derived MSCs present a viable alternative to harvested MSCs that have been shown to closely resemble conventional tissue-derived MSCs in terms of both phenotype and function [8,9]. Thus far, iPSC-derived MSCs have been generated in several eutherian models, including mouse, human, and dog [8,10 –12]; however, there are no descriptions of MSCs derived from a marsupial.
Previously, we have generated the first reported marsupial iPSCs from adult Tasmanian devil (Sarcophilus harrisii) dermal fibroblasts [13]. In this study, we report the generation of MSCs from the Tasmanian devil iPSCs (iMSCs), through both embryoid body (EB) formation assays and through inhibition of the transforming growth factor beta (TGF-β)/activin signaling pathway. These iMSCs exhibit MSC-like morphology, are highly proliferative, and express specific MSC surface proteins in addition to endogenously expressed pluripotency factors. Moreover, devil iMSCs readily differentiate along the adipogenic, osteogenic, and chondrogenic pathways in vitro and express a range of immunomodulatory and anti-inflammatory factors. These iMSCs are the first example of an iPSC-derived functional cell type from a marsupial and serve to further validate the devil iPSCs in addition to providing a unique resource for further in vitro studies. It is our hope that they also hold potential for the development of an effective MSC-based treatment strategy against Devil Facial Tumor Disease (DFTD), which is threatening Tasmanian devils with extinction.
Materials and Methods
The use of animal tissues was reviewed and permitted by the Animal Ethics Committee at The University of Queensland, Australia (Ethics approval number: SVS/550/12).
Culture of devil iPSCs
Two of the fully characterized clones of devil iPSCs (Clones A2 and A3) (13) were grown on irradiated mouse embryonic fibroblasts with iPSC medium prepared from KnockOut Dulbecco's modified Eagle's medium (DMEM) (Gibco, Invitrogen, Australia) with the addition of 15% (v/v) defined fetal calf serum (HyClone, GE Healthcare Life Sciences, Australia), 0.1 mM nonessential amino acid solution (NEAA) (Gibco), 1 mM
Mesengenic induction of devil iPSCs through inhibition of the TGF-β/activin signaling pathway (SB-iMSCs)
Devil iPSCs (Clones A2 and A3) were enzymatically passaged with TrypLE (Gibco) until small clumps of colonies were formed and iPSC bulk cultures were seeded into iPSC basal medium consisting of KnockOut DMEM supplemented with 15% (v/v) defined fetal bovine serum (FBS), 0.1 mM NEAA, 1 mM
Mesengenic induction of devil iPSCs through the EB differentiation method (EB-iMSCs)
A previously optimized protocol for human embryonic stem cells (ESCs) and iPSCs was followed to generate EBs from devil iPSCs [14]. Devil iPSCs (Clones A2 and A3) were enzymatically passaged with TrypLE and cultured in six-well ultra-low attachment plates (Costar) in iPSC medium. The culture medium was replaced every other day for 10–12 days. EBs were transferred to T25 tissue culture flasks with MSC medium (as in the previous section). EBs adhered to the flask, and differentiated cells grew outward from the center; ∼5–6 days later, the undifferentiated center of each EB was aspirated out manually, and the remaining cells were left in culture until they became confluent. A single cell suspension was generated using TrypLE, and the cells were seeded directly into T75 cell culture flasks with MSC medium and subsequently expanded.
Growth kinetics and doubling time
Both EB- and SB-iMSCs at passage 3, derived from Clones A2 and A3, were seeded into T75 tissue culture flasks at a cell density of 1 × 104 cells per cm2. Cells were passaged with TrypLE when they reached 80% confluency and counted with trypan blue using a cell counter (Bio-Rad Laboratories, Australia). The same seeding density was used for successive subcultures, and cell counting was performed for ∼35 days. Cumulative cell number and cumulative population doubling were determined as described previously [15].
RNA isolation, cDNA synthesis, and analysis of gene expression
Total RNA was extracted from devil iPSCs and iMSCs using an RNA Purification Kit (Macherey-Nagel, Thermo Fisher Scientific, Australia) as per the manufacturer's protocol. RNA was reverse transcribed with the iScript cDNA Synthesis Kit (Bio-Rad Laboratories). Quantitative polymerase chain reaction (PCR) analysis was carried out on a CFX96 real-time PCR detection system (Bio-Rad Laboratories) using SsoFast EvaGreen Supermix (Bio-Rad Laboratories). Expression data were normalized to the expression levels of devil β-ACTIN. The relative expression ratios of selected genes were calculated by the Delta Ct method. Devil iPSCs were used as the control. Validated primers and their product sizes are listed in Tables 1 and 2.
Tasmanian Devil Primer Sets Used to Confirm the Expression of Mesenchymal Stem Cell-Specific Surface Markers and Endogenously Expressed Pluripotency Factors
Tasmanian Devil Primer Sets Used to Confirm the Constitutive Expression of Immunomodulatory and Anti-Inflammatory Factors
Fluorescence immunocytochemistry
Devil iMSCs were seeded onto sterile coverslips and placed into 12-well plates (Costar) at 1 × 104 cells per cm2 and cultured until 75%–80% confluent. Cells were washed with phosphate buffered saline (PBS) (Invitrogen) three times before fixing with 2% paraformaldehyde (PFA) for 10 min at ambient temperature. Next, cells were permeabilized with 0.1% Triton X-100 in PBS for 20 min and blocked with 5% goat serum (Jackson ImmunoResearch) in PBS. Cells were incubated at 4°C overnight with the primary antibodies, which were diluted in 3% goat serum in PBS at the following dilutions: anti-OCT4 (1:50, MAB4401; Merck Millipore), anti-SOX2 (1:50, ab97959; Abcam, Australia), anti-NANOG (1:150, ab80892; Abcam), anti-CD73 (1:50, sc-25603; Santa Cruz Biotechnology), anti-CD90 (1:50, SM420P; Acris Antibodies, Germany), and anti-CD105 (1:50, ab156756; Abcam). In control groups, primary antibodies were excluded and cells were incubated only with 3% goat serum. Next day, cells were washed again three times with PBS and incubated with secondary antibodies, diluted in PBS to 1:1,000, at room temperature for 1 h. The following secondary antibodies were used: Goat Anti-Rabbit IgG H&L (Alexa Fluor® 568) (ab175471; Abcam), Goat Anti-Mouse IgG H&L (Alexa Fluor® 488) (ab150113; Abcam), Goat Anti-Rat IgG H&L (Alexa Fluor 488) (ab150157; Abcam), and Goat Anti-Mouse IgM μ chain (Alexa Fluor 488) (ab150121; Abcam). Coverslips were washed with PBS and mounted onto glass slides with ProLong mounting medium containing 4′,6-diamidino-2-phenylindole (DAPI) (Invitrogen). Photomicrographs were captured on an Olympus 1X51 inverted fluorescence microscope (Olympus, Australia).
Flow cytometry
Devil iMSCs were dissociated with TrypLE, and a total of 2 × 106 cells was used for the analysis of each marker. Cells were washed with PBS three times and fixed with 4% PFA for 20 min at room temperature. Cells were again washed with PBS three times and blocked for 30 min at ambient temperature with 5% goat serum added to 1% bovine serum albumin in PBS (PBA). Cells were then incubated overnight with primary antibodies at 4°C. Primary antibodies were diluted in 5% goat serum/PBA as follows: anti-CD73 (sc-25603; 1:50) and anti-CD90 (SM420P; 1:50). Negative controls were incubated only with PBA. Next day, cells were washed three times with PBA and incubated with secondary antibodies (as stated above) for 1 h at room temperature. Finally, cells were analyzed with a CytoFLEX Flow Cytometer (Beckman Coulter, Australia). Sixty thousand events were recorded for each sample.
In vitro mesodermal differentiation assays
The mesodermal differentiation potential was assessed using StemPro Adipogenesis, Osteogenesis, and Chondrogenesis Differentiation Kits (Life Technologies, Australia) as per the manufacturer's protocols. Osteogenic and adipogenic differentiation were assessed by plating cells into six-well tissue culture plates at a density of 1 × 104 cells/cm2. Cells were cultured with osteogenic or adipogenic induction medium, which was replaced every 2–3 days for 14 days. For the chondrogenic differentiation assay, 1 × 106 cells of devil iMSCs were collected by centrifugation (200 g for 3 min) in a 15 mL conical tube. The supernatant was discarded and substituted with chondrogenic induction medium. The medium was replaced every 3 days for 21 days. Chondrogenic and osteogenic differentiation were observed by staining with Alcian blue (Sigma-Aldrich, Australia) and Alizarin red S (Sigma-Aldrich), respectively. Adipogenic differentiation was confirmed with HCS LipidTOX Red staining (Life Technologies).
In vitro teratoma assays
Teratoma formation utilizing immune deficient mice is the “gold-standard” assay to define true pluripotent stem cells (PSCs), which form disorganized tumors containing tissue derivatives of the three embryonic germ layers [16]. However, as an alternative to increasingly controversial live animal testing, a modified protocol of the methylcellulose gel-supported three-dimensional in vitro teratoma assay was used as previously described [13,17]. Devil iMSCs were dissociated with TrypLE, and ∼3 × 105 cells from each group were cultured between two layers of 10% (w/v) low molecular weight methylcellulose (Sigma-Aldrich). iMSCs were cultured with DMEM containing 20% (v/v) FBS, 0.1 mM NEAA, 6 mM
Culture of Devil Facial Tumor 1 cells and preparation of conditioned medium
Devil Facial Tumor 1 (DFT1) cells were generously provided by Prof. Gregory Woods (The University of Tasmania, Tasmania, Australia) and cultured in complete Roswell Park Memorial Institute (RPMI) medium (Gibco) with the addition of 10% FBS, 1 mM
For the preparation of conditioned medium, DFT1 cells were cultured in T75 tissue culture flasks until they reached 75%–80% confluency before being washed with PBS thrice and refed with serum-free DFT1 culture medium (as above, but without FBS). Culture supernatant was collected at 24 and 48 h postincubation and pooled together. The collected conditioned medium was stored at −80°C before being used for the experiments.
Scratch assay
Each well of a six-well tissue culture plate was seeded with devil iMSCs (Clones EB-A2 and SB-A2) (Passage 5) to a final concentration of 1 × 105 cells/well, in MSC medium (as above), and maintained at 37°C and 5% CO2 for 24 h to allow cell adhesion and the formation of a confluent monolayer. A single scratch was made in the confluent monolayers with sterile 200 μL pipette tips. Each well was washed twice with serum-free MSC culture medium to discard dislodged cells. “Wound” closure was observed by collecting digitized images at 24, 48, and 72 h after the scratch was performed.
In vitro migration assay
In vitro migration assays were performed using the Cultrex 24-Well Cell Migration Assay system (R&D Systems, Australia). Devil iMSCs were cultured in T75 tissue culture flasks until they reached 80% confluency. In the final 24 h before the assay, cells were starved in a serum-free MSC medium (MSC medium without FBS). Following trypsinization, devil iMSCs (EB-iMSCs and SB-iMSCs) were seeded in the upper chamber (1 × 105 cells/well), while conditioned medium from DFT1 cultures was added to the lower chamber, of each well. Two wells were used for each cell line. Cells were then incubated at 37°C, 5% CO2 for 24 h. Serum-free medium was used as the negative control in place of DFT1-conditioned medium. The following day, medium from the top chambers was carefully aspirated, and each insert was washed with 100 μL of 1 × wash buffer according to the manufacturer's protocol. Medium in the bottom chambers was similarly aspirated, and each bottom chamber was washed with 500 μL of 1 × wash buffer. Next, 500 μL of cell dissociation solution/Calcein-AM was added to the bottom chambers, and the cell migration device was reassembled. After 1 h, the top chambers were removed and the plate was read at 485 nm excitation and 520 nm emission on a TECAN M200 plate reader (Tecan Group Ltd., Männedorf, Switzerland). Using a standard curve, relative fluorescence units were converted to cell number, and the percent invasion was determined as detailed in the manufacturer's protocol.
Statistical analysis
All values are reported as the mean ± standard error of the mean and are representative of at least three independent experiments except where specifically stated otherwise. Differences between individual groups were analyzed by analysis of variance. Means were compared by Student's t-test using GraphPad Prism 7 software. Significance is indicated as: not significant (ns) P > 0.05, *P ≤ 0.05, **P ≤ 0.01, ***P ≤ 0.001, ****P ≤ 0.0001.
Results
Inhibition of TGF-β/activin type I receptor promotes differentiation of devil iPSCs toward a mesenchymal-like morphology
Incubation of devil iPSCs with TGF-β/activin type I receptor inhibitor (SB431542) for 14 days induced their differentiation toward an MSC-like morphology. This epithelial to mesenchymal transition (EMT) was evident by the gradual formation of a monolayer of cells occupying the culture vessel, particularly around the periphery of a colony (Fig. 1). Following passage into MSC medium the cells rapidly adhered to the plastic surface. After subsequent passages (first or second passage), cells displayed a fusiform to stellate appearance and were uniformly distributed as single cells throughout the culture vessel, resembling adult MSCs described from other mammalian species. In contrast, in the absence of TGF-β/activin type I receptor inhibitor, undifferentiated devil iPSCs remained as dome-shaped, multilayered colonies of tightly packed cells (data not shown).
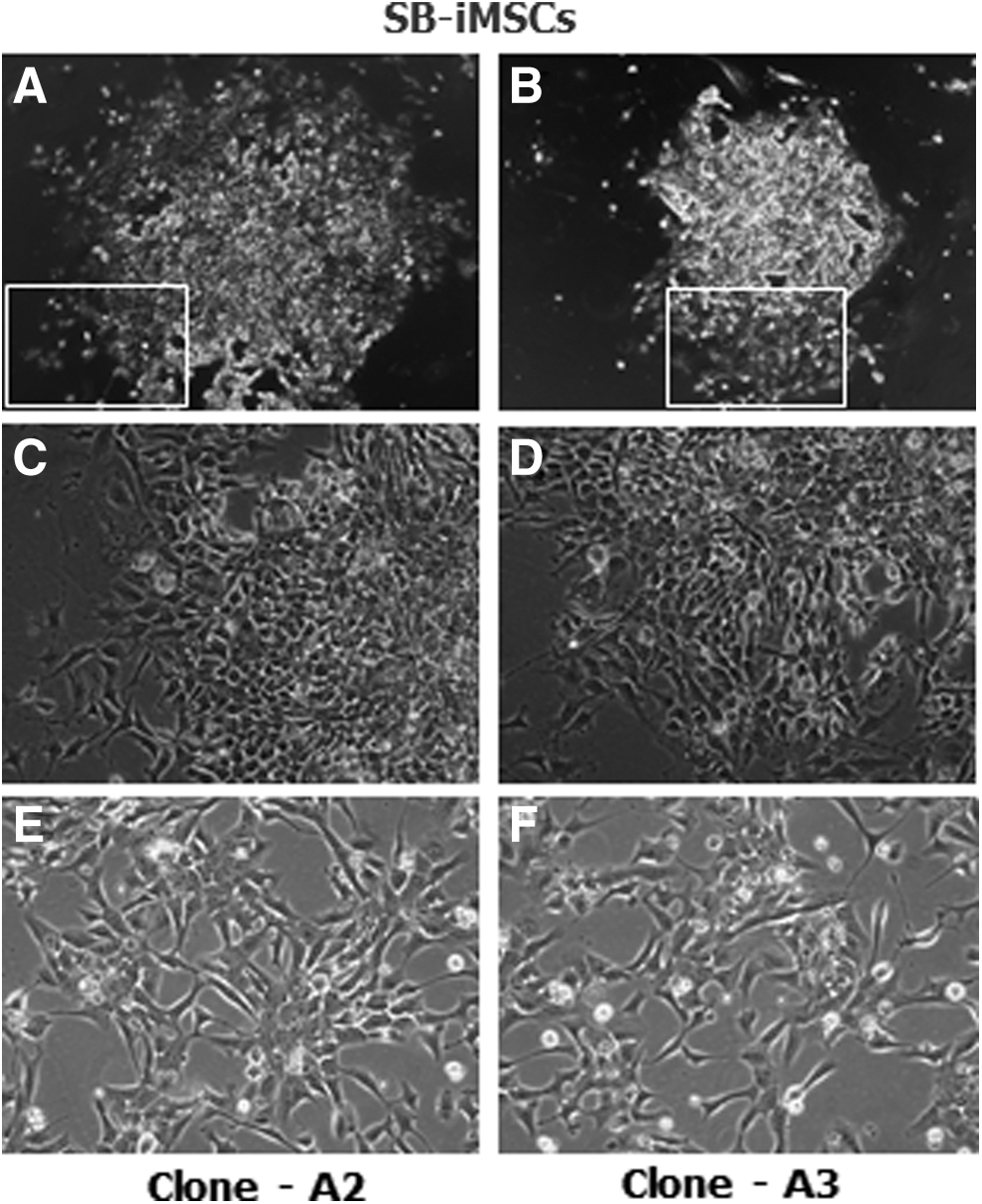
Morphology of devil iMSCs derived by the TGF-β/activin type I receptor inhibitor method.
Derivation of MSC-like cells through an EB differentiation assay
EBs were first generated for 10–12 days in iPSC medium as spheres and transferred to uncoated tissue culture flasks containing standard MSC medium. After transfer to tissue culture flasks, EBs attached to the surface of the culture vessel and, subsequently, cells with a differentiated morphology grew outward from the center of the EBs. By passage two or three these cells displayed an MSC-like morphology that was comparable to that observed in the SB-iMSCs (Fig. 2).
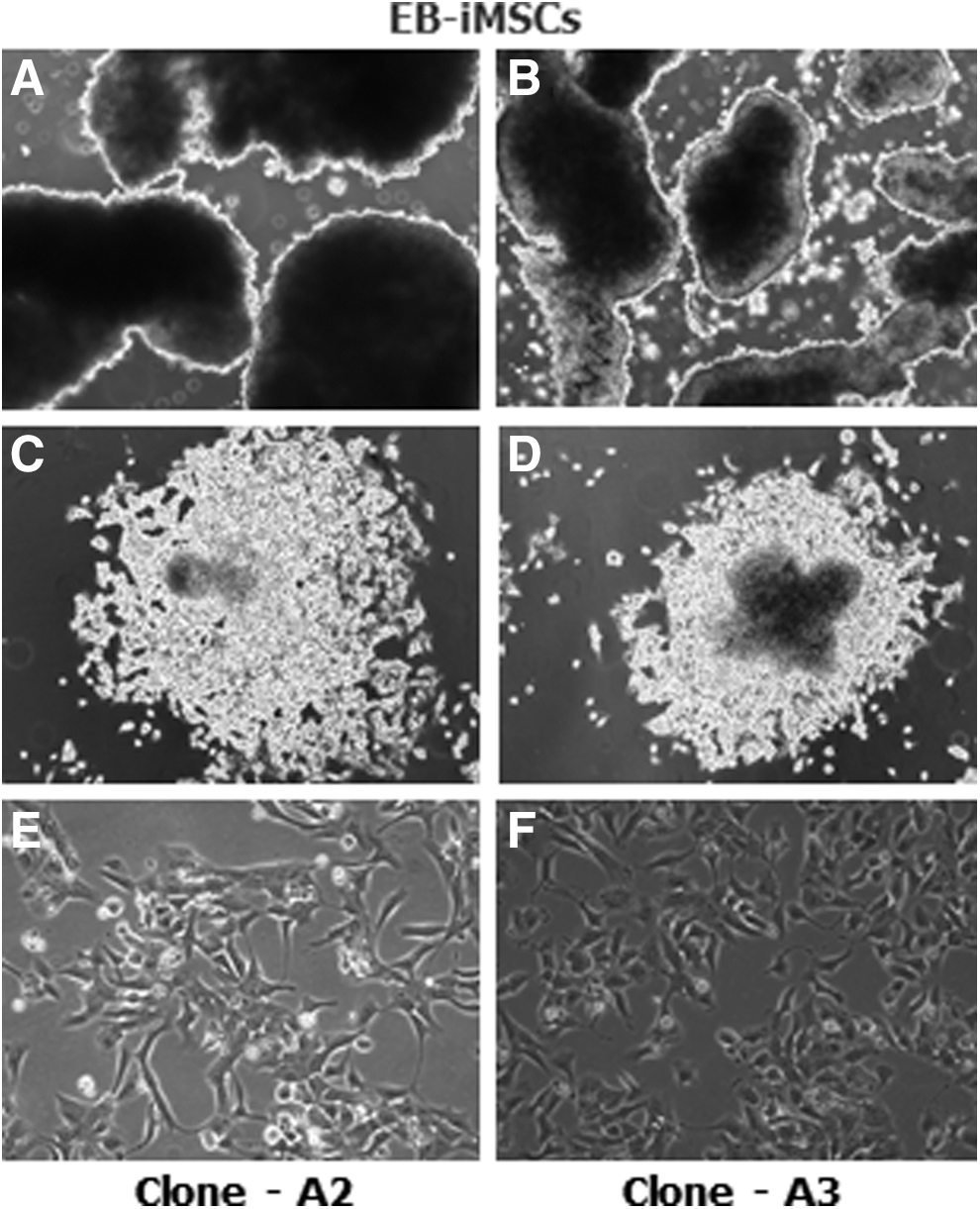
Morphology of devil iMSCs derived by the EB differentiation method.
Devil iMSCs express MSC-specific surface markers
As proposed by the International Society for Cellular Therapy (ISCT), MSCs should express a specific cell-surface antigen expression profile [18]. Therefore, we performed fluorescence immunocytochemistry to demonstrate that devil iMSCs possess an immunophenotype that is consistent with mammalian MSCs. As expected, devil iMSCs derived from both inhibitory and EB differentiation methods (SB-iMSCs and EB-iMSCs, respectively) robustly expressed characteristic MSC surface proteins CD73, CD90, and CD105 (Fig. 3A, B). Negative controls for nonspecific immunostaining are shown in Supplementary Fig. S1.
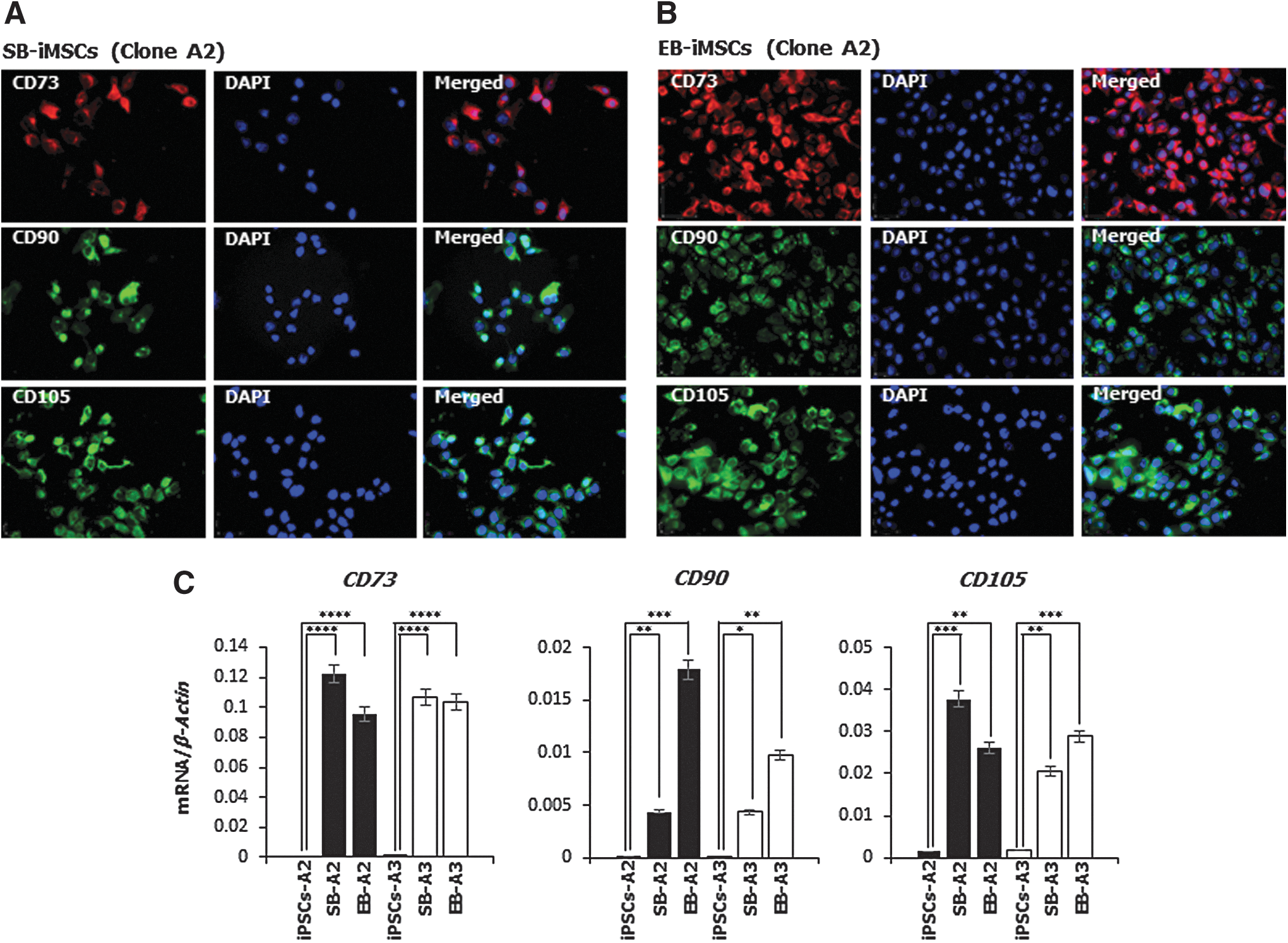
Devil iMSCs express MSC-specific surface markers.
The relative mRNA expression levels of these MSC-specific surface markers were further analyzed using quantitative reverse transcription-polymerase chain reaction (RT-PCR) analysis. Clones of both SB- and EB-derived iMSCs had significantly upregulated expression of CD73, CD90, and CD105 compared with devil iPSCs that did not express CD73 or CD90 and had extremely low levels of CD105 expression (Fig. 3C).
Based on flow cytometric analysis, between 82% and 97% of iMSCs (Fig. 4) express the cell surface proteins CD73 and CD90. Despite repeated attempts, with various adjustments to the protocol, we were unable to generate meaningful data using the antibody to CD105.
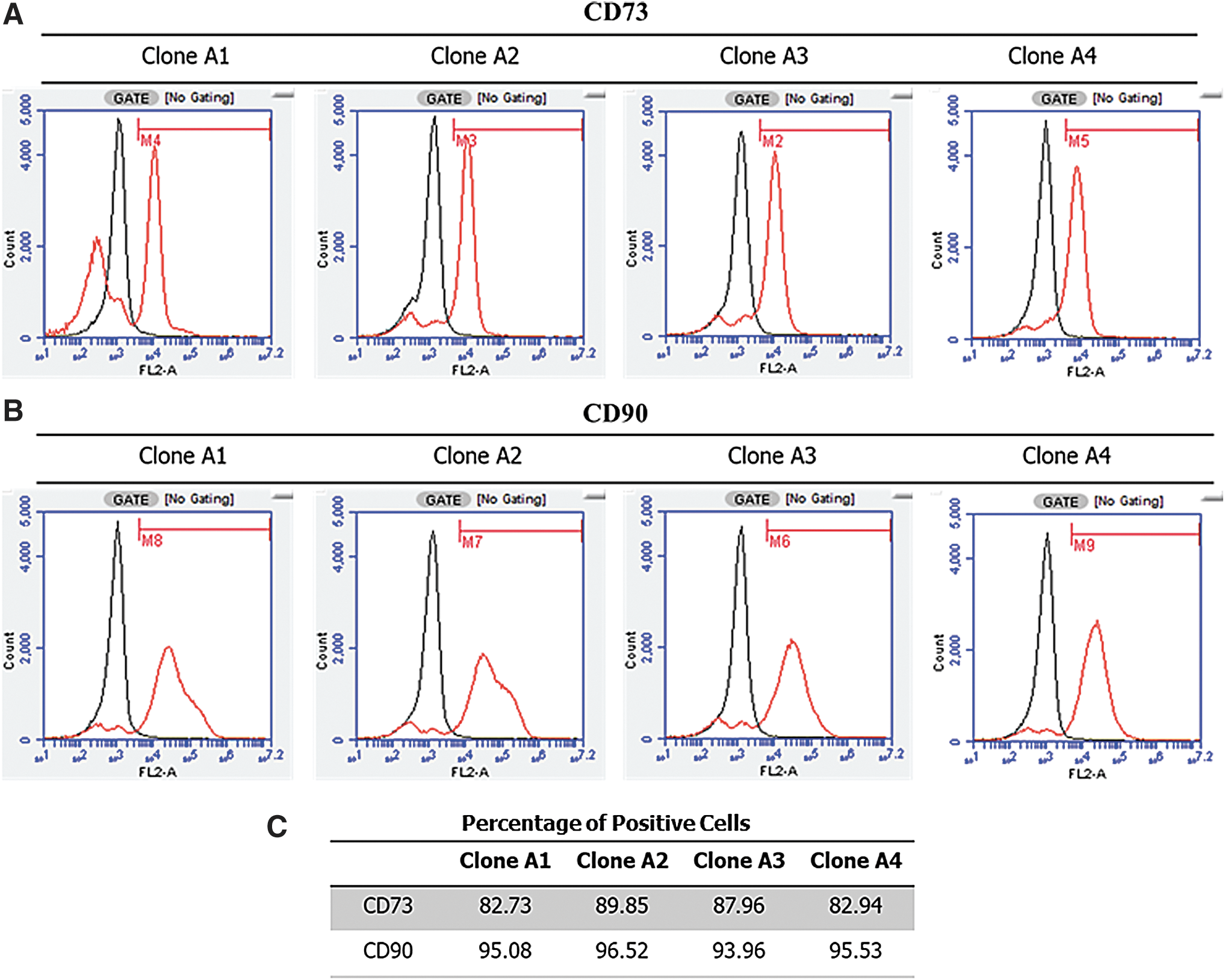
The majority of iMSCs express CD73 and CD90.
Devil iMSCs express devil endogenous pluripotency factors
We further characterized devil iMSCs with respect to their expression of pluripotency markers. Interestingly, devil iMSCs derived from both inhibitory and EB differentiation methods continue to express devil endogenous pluripotency factors OCT4, SOX2, and NANOG at both the mRNA and protein levels (Fig. 5). Expression of SOX2 and NANOG was reduced in both EB- and SB-MSCs compared to the iPSCs, with the exception of SOX2 expression in EB-iMSCs derived from iPSC clone A2 which did not change. This is in contrast to the expression of OCT4, which was not significantly different between iPSCs and iMSCs. The most significant change in expression between iPSCs and iMSCs was seen with the marsupial pluripotency factor POU5F3, a paralogue of OCT4, which was robustly expressed in both clones of iPSCs but diminished to low levels in all iMSCs (Fig. 5C). Negative controls for nonspecific immunostaining are shown in Supplementary Fig. S2.
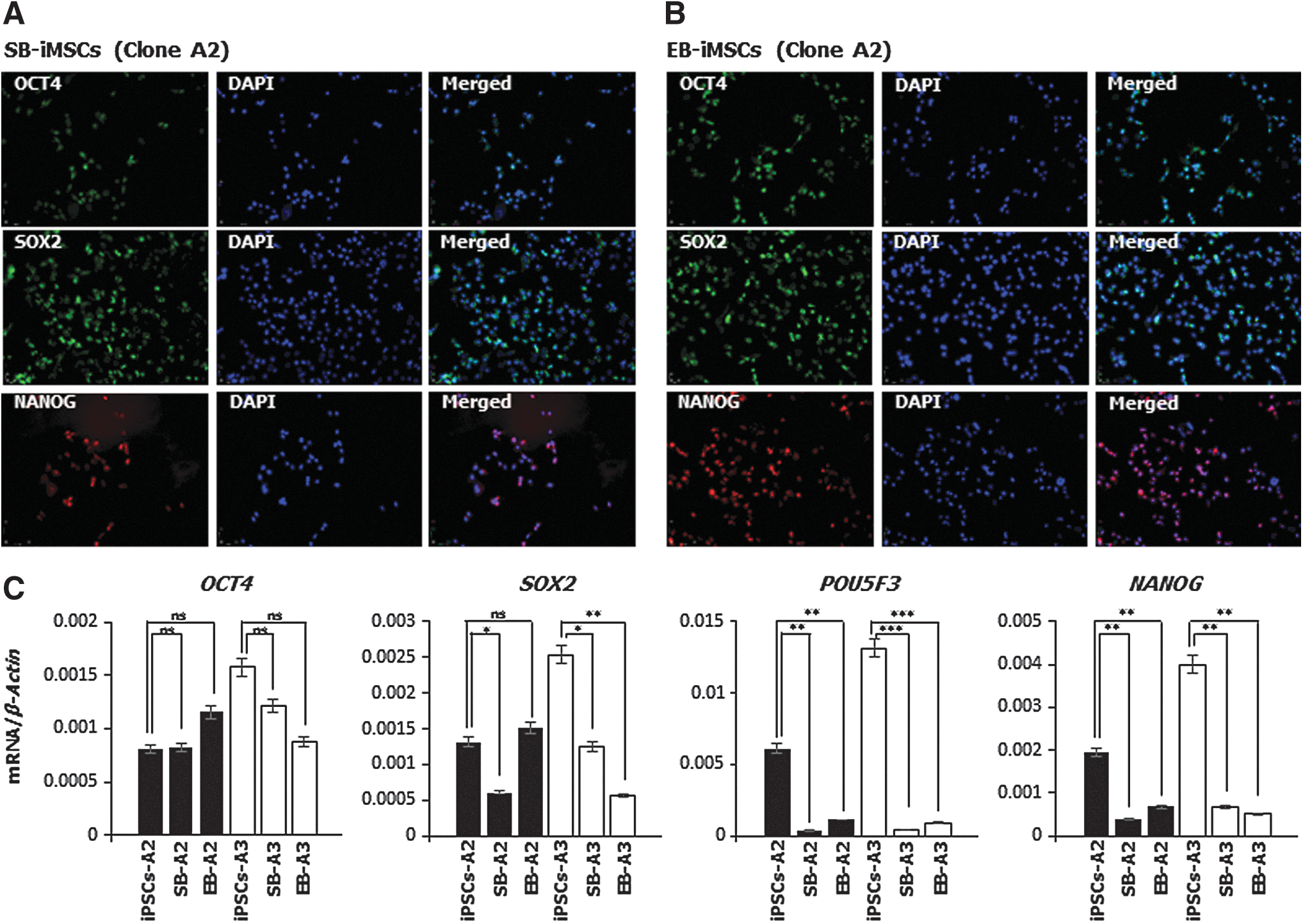
Devil iMSCs express pluripotency factors.
Because the devil iPSCs were generated using a lentiviral system with the transduction of human transcription factors, there is a possibility for reactivation of these transgenes during the EMT. We have previously shown that in devil iPSCs, all six human transcription factor transgenes were transcriptionally silenced at passage 20 (13). Likewise, devil iMSCs (which were generated from devil iPSCs at passage 22) showed no expression of the human transgenes (Supplementary Fig. S3), further confirming that the OCT4, NANOG, and SOX2 proteins detected in the iMSCs are the result of transcription from the devil genes.
Devil iMSCs undergo osteogenesis, chondrogenesis, and adipogenesis
The multipotency of MSCs is typically demonstrated by their differentiation potential into osteogenic, chondrogenic, and adipogenic lineages [19]. Therefore, devil iMSCs were next subjected to standard osteogenic, chondrogenic, and adipogenic differentiation protocols. Devil iMSCs (EB-iMSCs and SB-iMSCs; Clone A2) which were cultured in osteogenic differentiation medium demonstrated mineral deposits that stained strongly with Alizarin red S, characteristic for osteogenic differentiation (Fig. 6A). Devil iMSCs also underwent robust chondrogenic differentiation as demonstrated by Alcian blue staining (Fig. 6B) and adipogenesis as evidenced by the presence of intracellular lipid droplets (Fig. 6C). Negative controls are shown in Supplementary Fig. S4.
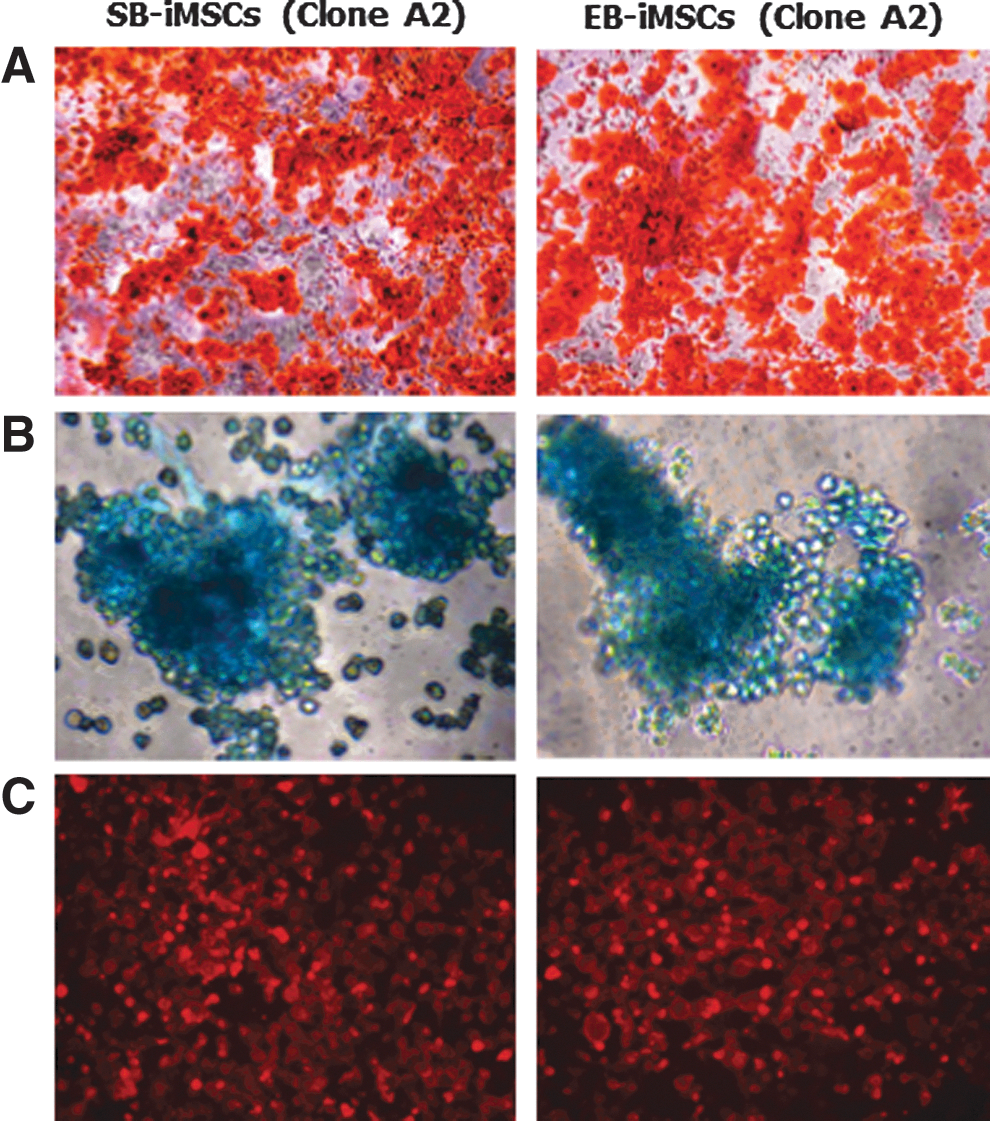
In vitro mesodermal differentiation potential. Devil iMSCs were exposed to standard adipogenic, osteogenic, and chondrogenic differentiation protocols.
The growth kinetics of devil iMSCs
The proliferative potential of cells is vital with regard to their application in cellular therapies. Therefore, we next evaluated the growth kinetics of devil iMSCs. After 30 days in culture, SB-iMSCs derived from Clones A2 and A3 reached a cumulative cell number of 2.7 × 1011 and 3.56 × 1012, respectively. Interestingly, during the same period EB-iMSCs derived from Clones A2 and A3 reached a cumulative cell number of 1.68 × 1010 and 8.6 × 1010, respectively (Fig. 7A, B). Over this period, cumulative population doublings of SB-iMSCs were greater than those of EB-iMSCs derived from both clones in that SB-iMSCs derived from Clones A2 and A3 achieved a cumulative population doubling of 16.8 and 20.5, respectively, whereas EB-iMSCs derived from Clones A2 and A3 reached a cumulative population doubling of 12.8 and 15.2, respectively (Fig. 7C, D). The live cell counts were between 86% and 94% over this period of evaluation and were consistent across all analyzed cell lines. Significantly, we have maintained these iMSCs for more than 25 passages without any loss of proliferative ability or cell viability during culture (data not shown).
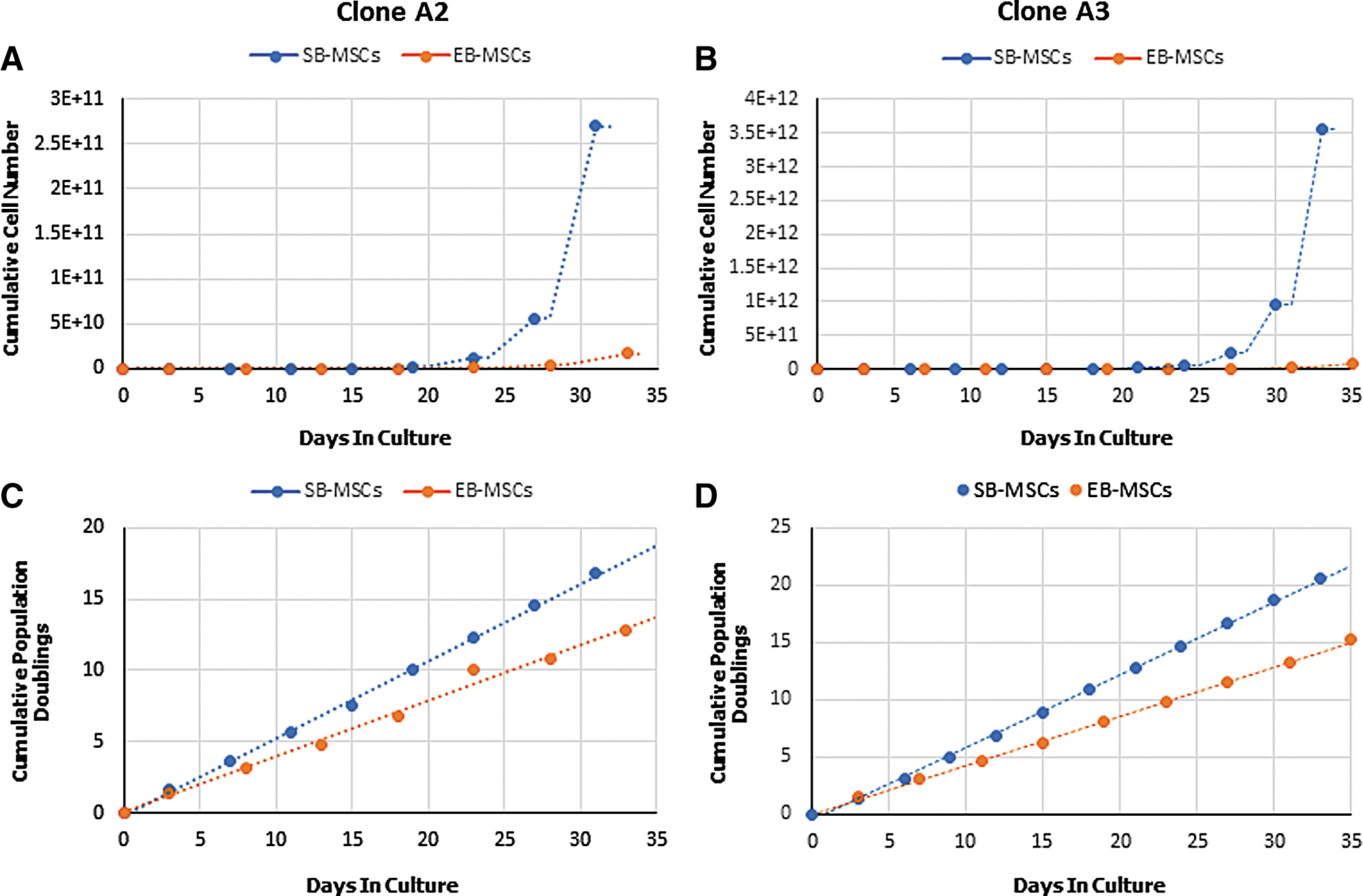
Growth kinetics of devil iMSCs.
Devil iMSCs are multipotent, but not pluripotent
Teratomas from devil iMSCs derived from both inhibitory and EB differentiation methods contained tissue derivatives only from the mesodermal germ layer. Teratomas from EB-iMSCs and SB-iMSCs clearly displayed adipose tissue and bone spicules (Fig. 8A, B) indicating that devil iMSCs are multipotent, but not pluripotent, cells that can differentiate toward the mesodermal lineage without the addition of exogenous differentiation factors.
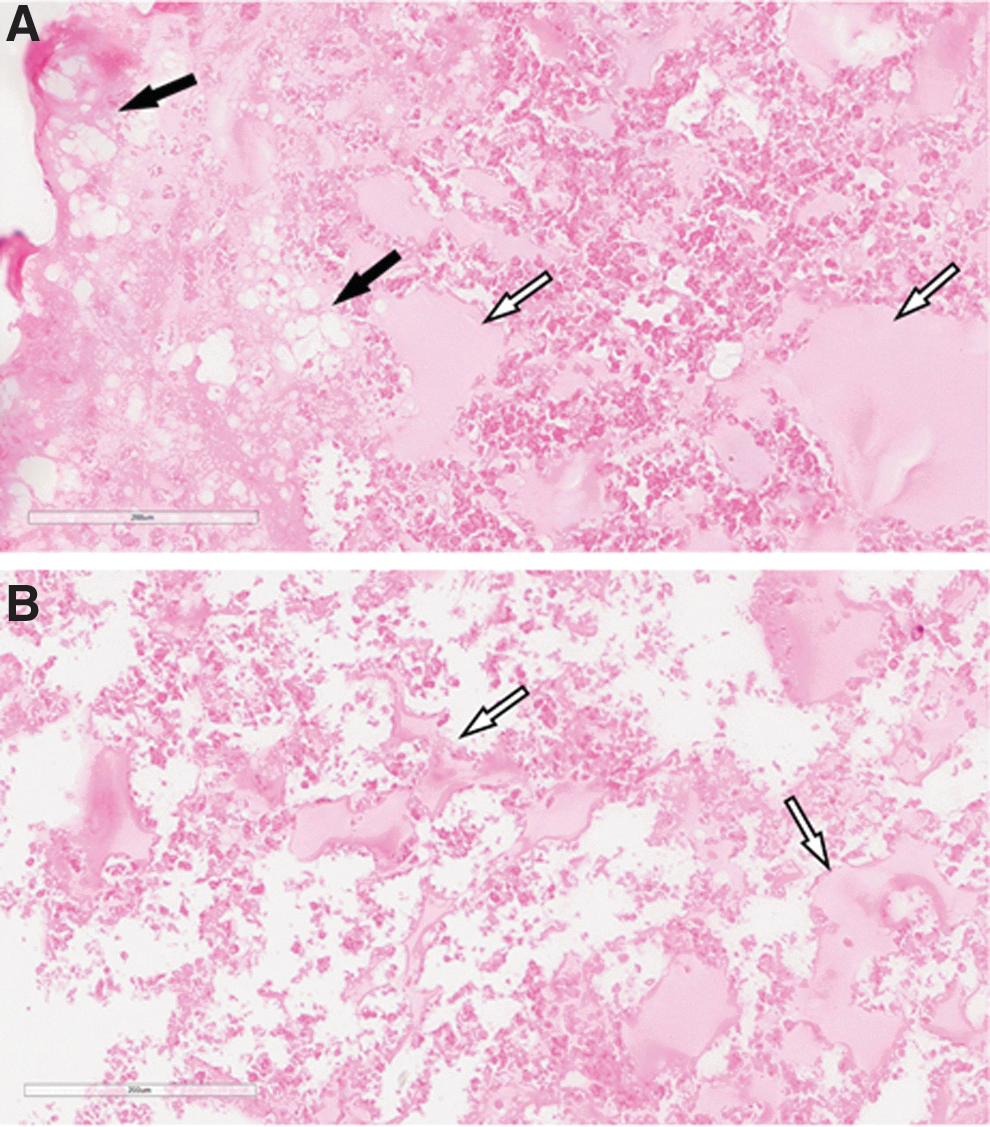
iMSCs are multipotent but not pluripotent. Both EB- and SB-iMSCs formed adipose tissue and bone in in vitro teratoma assays. The data shown are from EB-iMSC Clone A3.
Devil iMSCs display tropism toward medium conditioned by devil facial tumor cells
Human and mouse MSCs have been reported to have migratory properties and show a tropism toward tumors and sites of inflammation. To confirm a migratory potential in devil iMSCs, a traditional scratch/wound healing assay was performed and within 72 h iMSCs (SB-A2 and EB-A2) had filled the deficits created by the scratches (Fig. 9A).
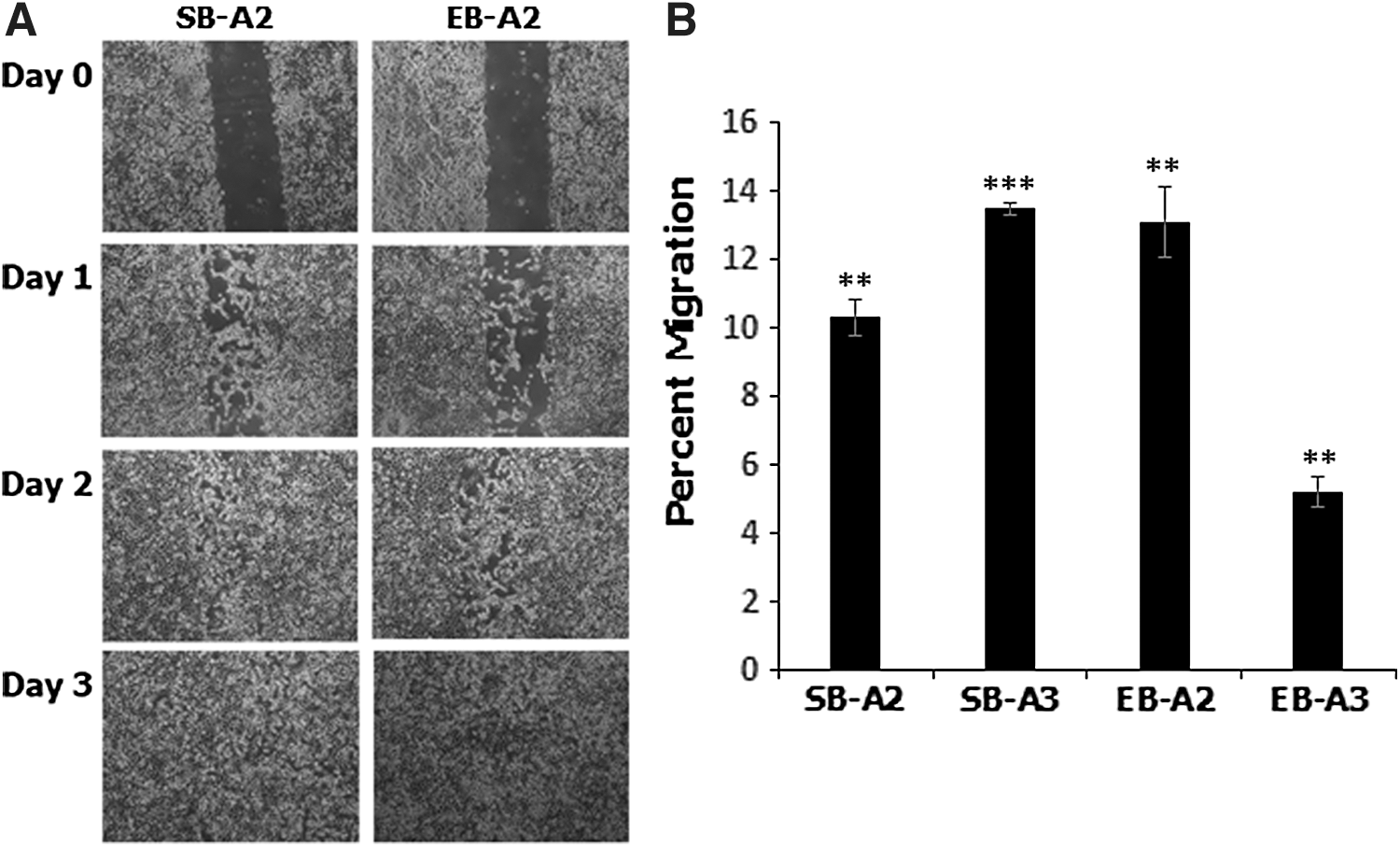
Devil iMSCs are migratory and display a tropism to medium conditioned by devil facial tumor cells.
A transwell migration assay (Boyden chamber assay) was used to determine if iMSCs possess a tropism toward devil tumor cells or, more specifically, toward medium conditioned by them. We observed a highly significant increase of 10.31% ± 0.55% (P ≤ 0.01) and 13.47% ± 0.18% (P ≤ 0.001) in migration of SB-A2 and SB-A3 iMSCs, respectively, toward DFT1-conditioned medium compared to control medium (Fig. 9B). EB-A2 and EB-A3 iMSCs displayed a 13.07% ± 1.03% (P ≤ 0.01) and 5.21% ± 0.45% (P ≤ 0.01) increase in migration, respectively, compared to control cultures (Fig. 9B).
Devil iMSCs express immunomodulatory and anti-inflammatory factors
MSCs across a range of eutherian species secrete factors that can modulate the immune system in response to inflammation [20]. Therefore, we analyzed the constitutive expression of key immunomodulatory and anti-inflammatory factors by devil iMSCs at the transcriptional level. Both EB- and SB-iMSCs express the immunomodulatory factors interleukin 6 (IL-6), transforming growth factor-β (TGF-β), prostaglandin E2 (PGE2), vascular endothelial growth factor (VEGF), and inducible nitric oxide synthase (iNOS) (Fig. 9A). A study of the expression of anti-inflammatory factors revealed differences between EB- and SB-derived iMSCs in the expression of interleukin 8 (IL-8) and hepatocyte growth factor (HGF), both of which are expressed at significantly higher levels in SB-iMSCs than in EB-iMSCs. Both EB- and SB-iMSCs express the anti-inflammatory factors galectin-9 (GAL-9), chemokine ligand 5 (CCL5), and stromal cell-derived factor 1 (SDF1) (Fig. 10).
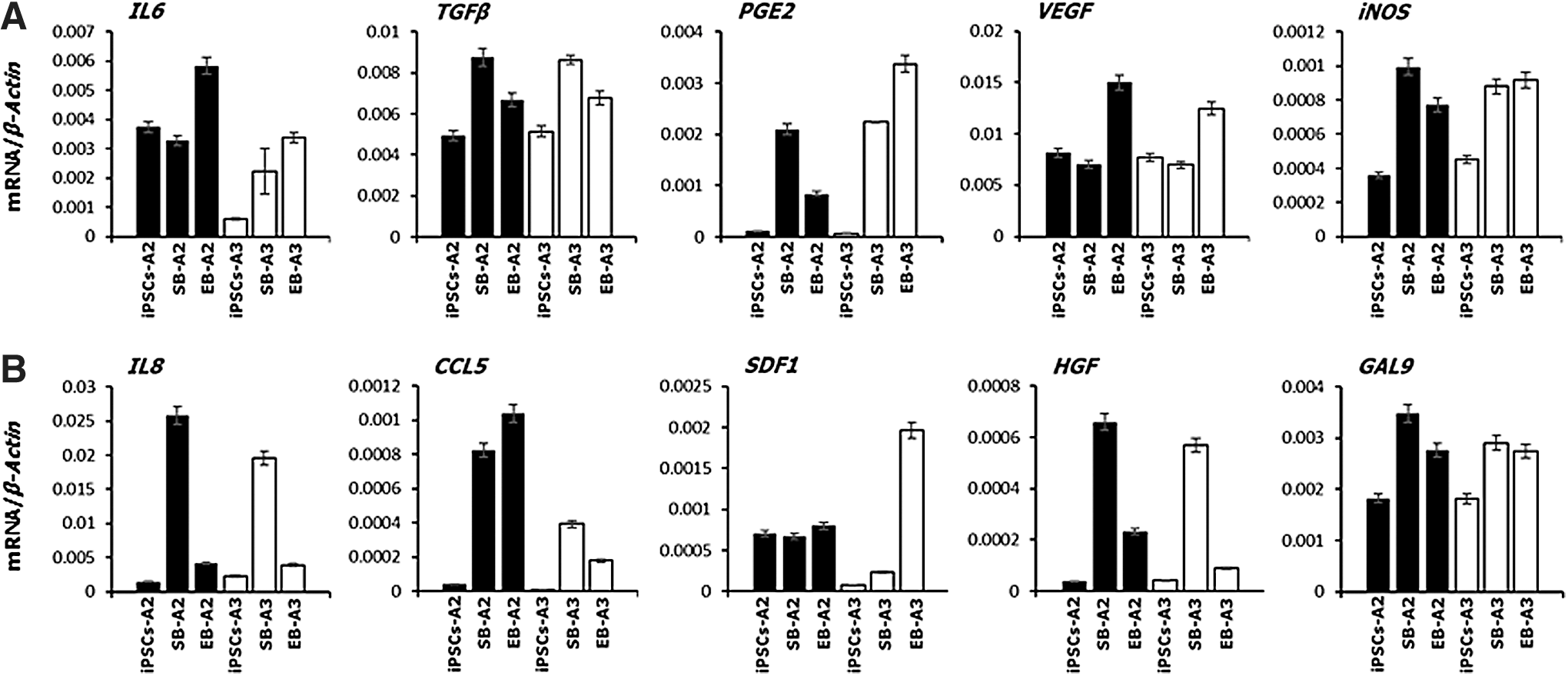
Devil iMSCs express immunomodulatory and anti-inflammatory factors. Quantitative RT-PCR analysis for
Discussion
MSCs, also known as mesenchymal stromal cells, are an important focus in translational research because of their ability to differentiate into a variety of mesoderm-derived tissues and their anti-inflammatory and immunomodulatory properties. MSCs are typically sourced from bone marrow and fat of adults [21,22], but their clinical potential is hampered due to the scarcity of MSCs in tissues, their heterogeneity, and the invasive nature of harvesting procedures [23]. Therefore, various protocols for generating MSCs from PSCs have been developed to circumvent the need to harvest them directly from live animals [14]. To date, MSCs have been isolated from numerous eutherian mammalian species but not from a marsupial.
The conventional technique for the differentiation of human PSCs (ESCs and iPSCs) into diverse cell types, such as MSC-like cells, involves the formation of EBs and subsequent culturing in appropriate differentiating medium [24,25]. However, Chen et al. described a novel protocol to differentiate human PSCs into MSCs by inhibition of TGF-β/activin signaling and subsequently culturing the cells in appropriate conditions supporting the EMT [14]. We have previously generated canine iPSC-derived MSCs [12] using this protocol and in this study we have demonstrated that it is equally as applicable to marsupial iPSCs as it is to those of eutherians. The devil MSCs (iMSCs) that we have generated using both the EB formation and TGF-β/activin inhibition approaches share traits that are characteristic of MSCs described from other mammalian species, particularly with regard to their morphology, plastic adherence, immunophenotype, and differentiation potential.
Mesenchymal marker proteins are widely used to identify and characterize multipotent stem cells that are capable of differentiation into cells of the mesenchymal lineage. MSCs from a range of eutherian species have been shown to express specific cell surface proteins, including CD73, CD90, and CD105 [26]. Similarly, devil iMSCs express MSC-specific surface markers CD73, CD90, and CD105 both at mRNA and protein levels. Furthermore, flow cytometry confirmed that the majority of these cells were positive for CD73 and CD90. In addition, the expression levels, as well as the high percentages of positive cells, for these markers were consistent between and within the SB-derived and EB-derived cell lines.
Many studies have demonstrated that MSCs from different eutherian species express combinations of the key pluripotency factors OCT4, SOX2, and NANOG, depending on the tissue from which the cells were isolated and the age of the donor. For example, MSCs derived from the bone marrow of young rats express Oct4, Sox2, and Nanog, whereas MSCs from the bone marrow of older rats express only Oct4 [27]. Equine [28] and feline [29] bone marrow-derived MSCs and canine MSCs isolated from adipose tissue [30], bone marrow [12,30], and amniotic fluid [31] similarly express OCT4, NANOG, and SOX2, while bone marrow-derived MSCs and those generated from canine iPSCs also express REX1 [12]. Similarly, MSCs from human amniotic membranes express OCT4, NANOG, SOX2, and REX1 [32], while those isolated from fetal lung [33] and fetal membrane and yolk sac [34] express only OCT4 and/or NANOG. Thus, the expression of OCT4, NANOG, and SOX2 by the devil iMSCs is consistent with observations in MSCs from eutherian species and suggests conserved roles for these genes in MSC self-renewal and differentiation in both eutherians and marsupials.
While expression of SOX2 and NANOG was diminished in both EB- and SB-generated MSCs compared to the iPSCs (with the exception of SOX2 expression in EB-iMSCs derived from iPSC clone A2 which did not change), expression of OCT4 remained similar across the iPSCs and iMSCs. This is of interest when contrasted with the change in expression levels of the marsupial pluripotency factor and paralogue of OCT4/POU5F1, POU5F3 (previously referred to in the literature as POU2). During early marsupial development, both OCT4 and POU5F3 are expressed by cells of the epiblast, the marsupial equivalent of the eutherian inner cell mass [35,36]. However, expression of OCT4 in the epiblast appears to be cytoplasmic, while POU5F3 transcripts are restricted to the nucleus [35]. Furthermore, while OCT4 expression is also present within the hypoblast and trophectoderm, POU5F3 expression is tightly restricted to the epiblast [35]. Our devil iPSCs express both OCT4 and POU5F3 as expected, but levels of POU5F3 are several fold higher than those of OCT4 [this study and 13], which is also seen in blastocysts from the tammar wallaby [36]. POU5F3 expression levels are the most significantly decreased in the transition from pluripotent devil iPSCs to multipotent iMSCs. Taken together, these data suggest that POU5F3 may play a more critical role in maintaining pluripotency in marsupials than OCT4. Low levels of POU5F3 expression in devil iMSCs may reflect the observation by Frankenberg et al. [37] that a range of adult tissues in the tammar wallaby, including skin, kidney, skeletal muscle, liver, and lung, express POU5F3; the authors speculate that POU5F3 may be involved in maintaining multipotency of the adult stem cell population within these tissues, and our detection of low levels of POU5F3 expression in iMSCs adds strength to this argument.
A comparison of the growth kinetics of devil EB-iMSCs and SB-iMSCs over 30 days identified a superior proliferative potential, and hence higher cumulative cell numbers, of SB-iMSCs compared to EB-iMSCs. Similar observations were reported by Chen et al., in human MSCs whereby those derived by the SB inhibitory differentiation method displayed faster growth kinetics and, hence, achieved a markedly higher cumulative cell number than the MSCs derived by the EB differentiation method during the early expansion phases [14]. Regardless of the generation method, the observed high proliferative potential of devil iMSCs in culture provides an opportunity to generate clinically efficacious numbers of cells within a short expansion period.
The beneficial effects of MSC-based therapies have been associated with their potent immunomodulatory and anti-inflammatory effects, affecting both the innate and adaptive immune responses [4,38]. These properties are the result of direct cell–cell contact-dependent mechanisms, secretion of a diverse range of cytokines and chemokines, or by a combination of both mechanisms [39]. Devil iMSCs constitutively express a variety of immunomodulatory and anti-inflammatory factors at the transcriptional level. These include immunomodulatory factors such as IL-6, TGF-β, PGE2, VEGF, and iNOS, in addition to anti-inflammatory factors such as IL-8, CCL5, SDF1, HGF, and GAL-9. Due to the unavailability of recombinant devil pro-inflammatory cytokines (for example, TNFα, IFNγ, and IL-1β) and the toxic effects of pro-inflammatory cytokines from other species that were trialed, the induced secretion of immunomodulatory and anti-inflammatory factors by devil iMSCs could not be evaluated.
One of the major challenges of cancer therapy lies in the inefficient delivery of therapeutic agents directly to the tumor sites, particularly in the case of relapsed or metastatic tumors which are often smaller and directly inaccessible [40]. Once systemically administered, MSCs can home to sites of neoplasia, metastases, and inflammation, where they arrest within the vasculature of the tissue before migrating across the endothelium and infiltrating into the tissue [41]. Given this tropism, MSCs have attracted significant scientific interest as potential delivery vehicles for the targeted transport of antitumor agents. Compared to other delivery platforms, their tumor tropic properties enable MSCs to accumulate specifically at the tumor site for better distribution of therapeutic agents, and their ability to modulate the immune response helps to increase MSC survival after transplantation [42]. Thus, MSCs have been genetically engineered to express various antiproliferative, pro-apoptotic, and antiangiogenic reagents and shown to preferentially engraft and exert their effects on various tumor models [43 –46]. Encouraged by the tropism of devil iMSCs that we observed toward medium conditioned by DFT1 cells, and our ability to expand these cells to clinically relevant cell numbers, we are currently exploring the potential of MSC-based treatment strategies against DFTD.
Conclusion
In conclusion, we report the generation of marsupial MSCs from Tasmanian devil iPSCs. These iMSCs express the pluripotency factors OCT4, SOX2, and NANOG, but show significantly reduced expression of the marsupial pluripotency factor POU5F1, suggesting a more critical role for POU5F1 over OCT4 in maintaining marsupial pluripotency. The persistent low level of POU5F1 expression in devil iMSCs suggests that it is involved in supporting multipotency. Importantly, from a clinical perspective, while devil iMSCs express pluripotency markers, they are capable only of differentiating into mesodermal derivatives and do not form tumors in in vitro differentiation assays. In addition, iMSCs express a variety of immunomodulatory and anti-inflammatory factors and demonstrate a tropism toward medium conditioned by devil facial tumor cells, supporting further investigation into their potential use as an MSC-based treatment strategy against DFTD.
Footnotes
Acknowledgment
The authors are extremely grateful to Prof. Gregory Woods from The University of Tasmania, Australia, who very generously provided the DFT1 cells.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was funded by a Morris Animal Foundation grant to Deanne J. Whitworth (Grant Number: D14ZO-080) and an Australian Government Research Training Program Scholarship awarded to Prasanna Weeratunga. The funding bodies had no involvement in the study design, data collection and analysis, preparation of the article, or decision to publish.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.