
Abstract
Chronic persistent inflammation is thought to impede axon regeneration and cause demyelinating disease also with neuropathic pain, leading to more severe dysfunction after peripheral nerve injury. Increasing evidence indicates that neural stem cells (NSCs) have immunomodulatory effects, and previous studies have shown that many of the beneficial effects attributed to stem cell therapy may exert their therapeutic effects through paracrine mechanisms. In this research, the repairing effect of NSC-conditioned medium (NSC-CM) on sciatic nerve injury and its mechanism of repair were further explored. The present research showed that NSC-CM promoted histopathological and functional recovery after crush injury in rats, and what counts is that NSC-CM inhibited the inflammation of sciatic nerve in the late stage of injury. NSC-CM significantly downregulated the infiltration of proinflammatory factors [tumor necrosis factor alpha (TNF-α), interleukin 6 (IL-6), and IL-1β] as well as decreased the CD68 inflammatory macrophages infiltrating in the sciatic nerve. In addition, to study the effect of NSC-CM on the inflammatory state of macrophages in vitro, lipopolysaccharide (LPS) was used to induce the proinflammation of macrophages. The results showed that NSC-CM decreased the expression of macrophage proinflammatory-related proteins (IL-6, IL-1β, TNF-α, inducible nitric oxide synthase) induced by LPS. The activation of Sirt-1 signaling in macrophages effectively countered the proinflammation induced by LPS in the presence of NSC-CM. Using Sirt-1-specific inhibitor EX527 partially weakened the anti-inflammatory effect of NSC-CM. Altogether, this study demonstrated for the first time that NSC-CM promotes functional recovery after sciatic nerve crush injury in vivo and also inhibits the inflammation in activated macrophages by activating Sirt-1 signaling pathway in vitro.
Introduction
Peripheral nerve injury (PNI) is mainly related to trauma, tumors, and iatrogenic lesions, which causes neurological deficits and dysfunction in the body. Unlike the central nervous system, the axons of the peripheral nervous system can regenerate after injury. However, this regeneration is usually incomplete, which always leads to neuropathy [1]. Wallerian degeneration after PNI, including the occurrence of axonal necrosis, disappearance of myelin breakdown, and proliferation of nerve sheath and cell phagocytosis, has been regarded as the key event of peripheral nerve regeneration [2]. Successful Wallerian degeneration depends on multiple cellular events, which, in addition to axon regeneration, includes dedifferentiation of Schwann cells and activation of immune cells. Macrophages play an important role in the type of immune cells that function at the site of PNI [3]. The application of macrophages includes the removal of myelin fragments and the regulation of the activity of Schwann cells. Moreover, they promote axon regeneration by releasing a large number of axon regeneration-related factors. Importantly, macrophages also contribute to inflammatory events after injury [2]. Inflammation plays different roles in different periods after PNI. In the early stages, proinflammatory factors such as IL-β and tumor necrosis factor alpha (TNF-α) mainly secreted by dedifferentiated Schwann cells after losing contact with axons recruit macrophages to infiltrate the site of injury [4]. Meanwhile, the persistent inflammatory response causes demyelinating disease and neuropathic pain [5 –7]. Therefore, inhibiting the late-stage inflammation after sciatic nerve injury is an important event to inhibit demyelinating disease and promote nerve regeneration.
With a strong self-renewal and multidirectional differentiation potential, neural stem cells (NSCs) can differentiate into neurons, astrocytes, and oligodendrocytes. NSCs play an active role in a variety of neurological diseases, such as ischemic stroke [8], Alzheimer's disease [9], and spinal cord repair [10,11]. NSCs have also been shown to promote repair after PNI [12 –14]. Characteristics of NSCs, such as differentiating into Schwann-like cells and neurons, the expression of neurotrophic factors involved in neuronal protection and angiogenesis, and their roles in repair and regeneration of damaged peripheral nerves, including myelination and immunoregulation, enable NSCs to play the role of repairing peripheral nerve damage [14].
NSCs have been shown to exert great influence in anti-inflammation through macrophages. Specifically, NSCs transplants decrease the number of inflammatory mononuclear phagocytes, and promote the healing of the injured central nervous system (CNS). Coculture with macrophages also showed that NSCs reduced the expression of macrophage proinflammatory factors [15]. The conditioned medium of stem cells shows promising potential in tissue repair. Previous studies have also shown that stem cell-conditioned medium promoted the recovery of central nervous system diseases such as stroke and spinal cord injury [16 –18].
Given the established immune modulatory properties of NSCs and the therapeutic effects of stem cell-conditioned medium, we hypothesized that NSCs may exert their therapeutic effects in chronic neuroinflammation by modulating macrophage inflammation toward the reduction of peripheral nervous system (PNS) damage.
In this study, we investigated whether the NSC-conditioned medium (NSC-CM) could facilitate recovery within PNI. Furthermore, we explored its effects and possible mechanisms on macrophages.
Materials and Methods
Ethics, consent, and permissions
This study was approved by the Ethical Review Board of Jiangsu University.
Animals
Male Sprague-Dawley (SD) rats (120–140 g) were purchased from the Laboratory Animal Center of Jiangsu University (Zhenjiang, China). Rats were housed as 3 per cage in a controlled environment with 12 h light/dark cycle, and provided with water and food. All animal experiments were approved by the Institutional Animal Care and Use Committee of Jiangsu University (Permit Number: SCXK 2018-0012).
Cells culture
All cells were cultured at 37°C in a humidified atmosphere of 5% CO2. As previously described [19,20], NSCs were obtained from human embryonic stem cells (hESCs, SHhES2, passage 49), in short, hESCs were cultured with neural induction medium: Dulbecco's modified Eagle's medium (DMEM)/F12, 1 × B-27 (Gibco), 20 ng/mL basic fibroblast growth factor (PeproTech), 20 ng/mL epidermal growth factor (PeproTech), 1% nonessential amino acid (Thermofisher), and 1% penicillin–streptomycin. After 18–21 days, the cultured cells were resuspended in neural induction medium named P0 NSCs. The human monocyte THP-1 cell line (THP-1), from Professor Zhou (Jiangsu University), was cultured in RMPI1640 supplemented with 10% fetal bovine serum (FBS; Gibco). To obtain resident peritoneal macrophages (PMs), healthy SD rats were sacrificed with CO2 asphyxiation, and the peritoneal cavity lavages with 10 mL cold high-glucose DMEM (Gibco) were collected. As previously described [21], briefly, the peritoneal lavage was washed with phosphate-buffered saline (PBS) at 1,500 rpm for 10 min. The cells were resuspended in DMEM with 10% FBS and 1% pen/strep incubated for 1 h at 37°C in 5% CO2. Suspended cells were washed out with the DMEM. For macrophages activation, 100 ng/mL of LPS (from E. coli O111:B4, Sigma-Aldrich, Germany) were added for 6 h. Afterward, NSC medium (NSC-M) or NSC-CM was added with macrophages for 12 h and 24 h. To inhibit Sirt-1, 1 μM of EX527 (Selleck) was preadded for 6 h.
NSC-CM collection
NSC-M was refreshed when the cell confluence is ∼60%. After 48 h, conditioned medium was harvested and centrifuged at 1,000 g for 3 min to pellet cells; then, the collected supernatant was filtered with the 0.22 μm membrane (Millipore, Germany). NSC-CM was collected from P10-P20 NSCs and stored at −80°C. NSC-M was treated same and used for control. For vivo experiments, the NSC-CM was concentrated 5-fold and 10-fold by the lyophilizer (Labconco FreeZone, England) to create 5 × NSC-CM and 10 × NSC-CM, respectively, and then reconstituted with NSC-M. For vitro experiments, the NSC-CM and NSC-M were used to treat cells.
Sciatic nerve crush injury model
As described previously [5], the surgeon was blinded to the group identity. In brief, rats were anesthetized by intraperitoneal injection of pentobarbital (50 mg/kg). The gluteal muscle was bluntly separated to expose the right sciatic nerve. The exposed nerve proximal from sciatic notch was subjected to tight crush for 30 s using vascular clip. The vascular clip was marked with black ink to cause a crush injury of 3 mm. The sham-operated group rats went through the same surgical procedures, but without sciatic nerve crush injury. The NSC-M or PBS was used as the control, and different concentrations (NSC-CM, 5 × NSC-CM, 10 × NSC-CM) of NSC-CM were used to treat sciatic nerve crush injury. The equal volume of treatments were injected into the rats through the caudal veins (1 mL/rat) on the first day after injury.
Sciatic functional index
The assay was carried out as described previously [22]. The footprint analysis of rats was performed at 3, 7, 10, and 14 days after surgery to assess the recovery of hind limb function. Sciatic functional index (SFI) was calculated according to the following formula:
E and N represent experimental and normal, respectively; the entire plantar length (PL) indicates the distance from the heel to the third toe; the toe spread (TS) indicates the distance from the first to the fifth toe, and the intermediary toe spread (IT) indicates the distance from the second to the fourth toe. SFI reflects the recovery of sciatic nerve. A score of −100 represents the total loss of function, whereas oscillating ∼0 indicates an efficient nerve.
Hematoxylin–eosin staining
The injured nerve and gastrocnemius muscles of the rats were harvested, fixed in 4% paraformaldehyde for 24 h, and embedded in paraffin. Longitudinal sections (12 μm in thickness) and cross-sections (4 μm in thickness) of the harvested nerves and gastrocnemius muscles were prepared, and then subjected to hematoxylin–eosin (HE) staining.
Immunohistochemistry
The immunohistochemistry (IHC) of the sciatic nerve was conducted as described previously [23]. In brief, the sciatic nerve at 3 days and 14 days after injury was taken for sectioning. The anti-interleukin 6 (IL-6) antibody (1:300; Bioss, China), anti-IL-1β antibody (1:300; Servicebio, China), anti-TNF-α antibody (1:300; Servicebio), and anti-CD68 antibody (1:500; Servicebio) were used. The mean densities of IL-6, IL-1β, and TNF-α also with the positive number of CD68 were determined by using Image-Pro Plus software (Media Cybernetics, Bethesda, MD).
Transmission electron microscope
The midpoints of injured nerve were resected at 14 days after surgery for transmission electron microscope (TEM) (HITACHI-HT7700, Japan) analysis. As previously described [23], the ultrathin sections were observed under the transmission electron microscopy. And the number of myelin lamellae per field and the thickness of myelin sheaths were calculated by Image-Pro Plus software (Media Cybernetics).
Real-time quantitative-polymerase chain reaction
Total RNA from all samples in vitro was extracted using Trizol reagent (Sigma) following the manufacturer's protocol, and then reverse transcribed into cDNA by HiScript II QRT SuperMix (Vazyme, China). The replication of PCR products was monitored by SYBR™ green dye and analyzed with StepOnePlus™ (ABI). The method of 2−ΔΔCt was adopted to obtain the relative fold change of the target gene normalized to β-actin expression compared with control samples. The genes and the related specific primers used are shown in Table 1.
Genes and Sequence of Primer Pairs Used for Real-Time Quantitative-Polymerase Chain Reaction
IL-6, interleukin 6; iNOS, inducible nitric oxide synthase; TNF-α, tumor necrosis factor alpha.
Western blot
In brief, PMs were lysed in the lysis buffer (with protease/phosphatase inhibitors) to extract the total proteins of cells. The protein concentration of cell lysates was quantified by the BCA Protein Assay kit (Cwbio, China). The western blot analysis was performed as described previously [19]. The following primary antibodies were used for western blot: anti-HIF-1α (1:500; Wanleibio, China), anti-Sirt-1 (1:800; Wanleibio), anti-IL-1β (1:400; Wanleibio), anti-TNF-α (1:1,000; Bioworld, China), antinuclear factor kappa B (NF-κB) (1:1,000; Cell Signaling Technology), anti-p-NF-κB (1:1,000; Cell Signaling Technology), anti-inducible nitric oxide synthase (iNOS) (1:500; Santa Cruz Biotechnology), and anti-β-actin (1:500; Santa Cruz Biotechnology). Protein bands were detected using ECL (Millipore) reagents and acquired using FUSION Solo (France). The density of each band was normalized to housekeeping band β-actin and quantified using Image J software (NIH, Bethesda, MD).
Enzyme-linked immunosorbent assay
Cell medium was collected and centrifuged at 300 g for 10 min at 4°C, and supernatants were stored at −80°C for subsequent analysis. The expression of IL-6 and IL-1β was measured using ELISA kits (Multi Sciences, China) following the manufacturer's instructions.
Statistical analysis
All values were given as mean ± standard deviation (SD). One-way ANOVA was used to compare the differences among groups. And LSD-t test is used for further pairwise comparison. P < 0.05 was considered to be statistically significant.
Results
NSC-CM improves the functional recovery of sciatic nerve
To determine the optimal concentration of NSC-CM for sciatic nerve injury, different concentrations of conditioned medium were selected for sciatic nerve injury. The SFIs at 3 and 7 days were analyzed after sciatic crush injury, and the gastrocnemius muscle was weighed at 7 days after the injury. As shown in Fig. 1A, B, compared with NSC-M, NSC-CM and 5 × NSC-CM are not statistically significant; however, the SFI of 10 × NSC-CM has significantly improved compared with NSC-M at 7 days after the injury. Therefore, the dose that could alleviate the injury to the most extent was 10 × NSC-CM.
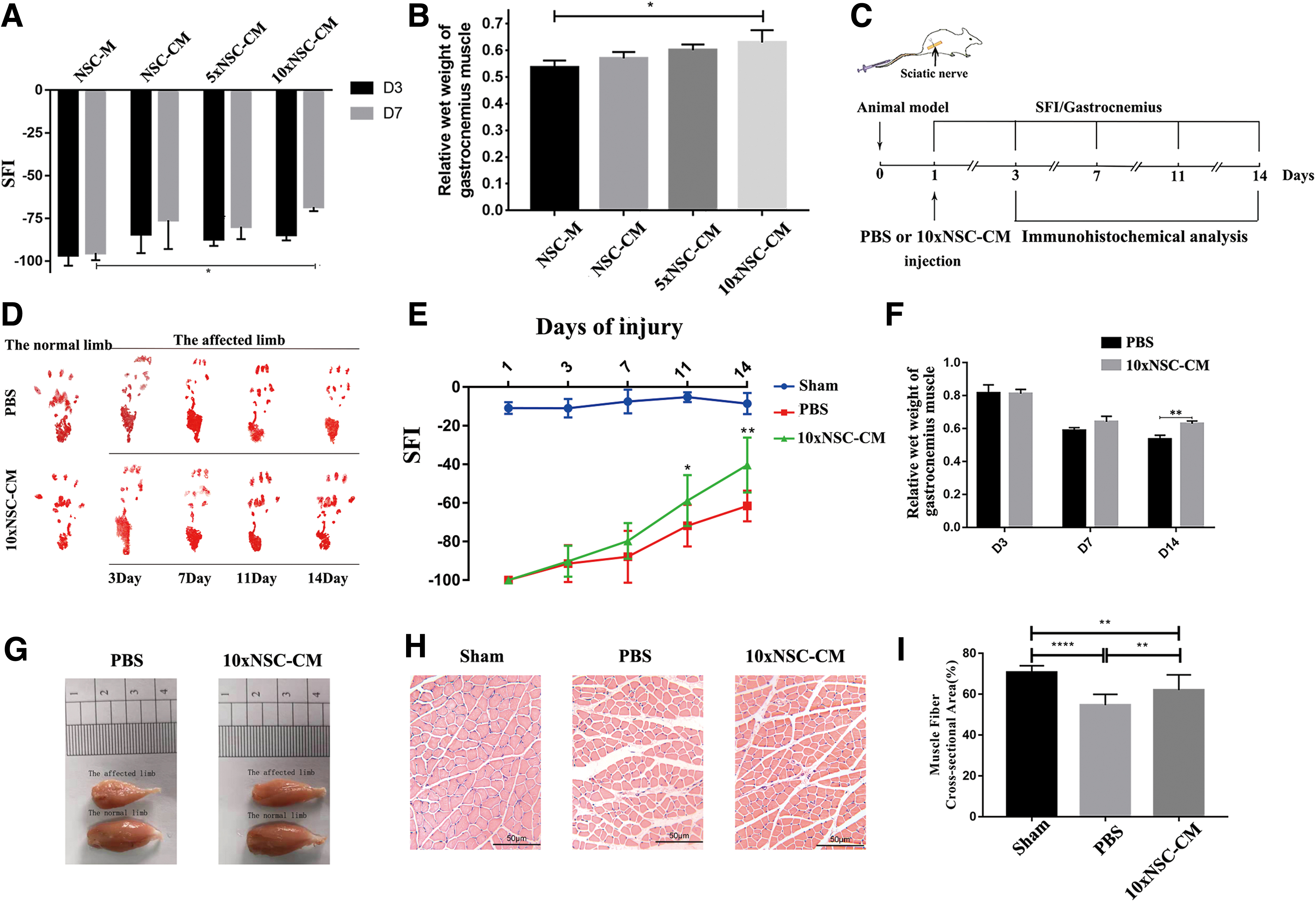
NSC-CM improves the functional recovery of sciatic nerve.
For further research, our study included three groups: the sham-operated group (Sham), the control group (PNI + PBS), and the NSC-conditioned medium treatment group (PNI +10 × NSC-CM). Figure 1C shows a schematic of the experimental process after 10 × NSC-CM or PBS treatment. The results of walking footprint analysis are shown in Fig. 1D to assess the motor function recovery of rats. The footprints of the 10 × NSC-CM group stretched at 11 days and approached the footprints of the normal limb at 14 days. After the sciatic nerve crush injury, SFI (Fig. 1E) values dropped to −100 in PBS group and 10 × NSC-CM group after 1 day, indicating a complete loss of sciatic nerve function. The SFI of 10 × NSC-CM group drastically increased relative to those of the control group at 11 and 14 days after surgery. Long-term denervation leads to gastrocnemius atrophy. But compared with the control group, the degree of mass loss of the gastrocnemius muscle in the 10 × NSC-CM group was weaker. As shown in Fig. 1F and G, on the 14th day after injury, the wet weight of the gastrocnemius muscle in the 10 × NSC-CM group was higher than that in the PBS group. The wet weight of the gastrocnemius muscle in the PBS group on the 14th day after injury was 0.53 relative to the normal side, while the wet weight of the gastrocnemius muscle in the 10 × NSC-CM group was 0.63 on the 14th day after injury.
As shown in Fig. 1H, HE staining showed that the normal gastrocnemius muscle fibers were neatly arranged and uniform in size, and the myocardial space was small and uniform. Compared with the PBS group, the gastrocnemius muscle of the 10 × NSC-CM group was closer to the normal muscle fiber morphology, and the atrophy of the muscle fiber and the thickening of the myenteric space were significantly improved. Moreover, the cross-sectional area of muscle fibers in the 10 × NSC-CM group recovered compared with the PBS group, which was ∼62% of the normal cross-sectional area, was closer to normal (Fig. 1I). These results indicated that NSC-CM could promote neurological recovery in rats after sciatic crush injury and neuromuscular reinnervation to slow the contraction of gastrocnemius muscle compared with PBS.
NSC-CM improves axonal regeneration and remyelination
HE staining was used to evaluate the regenerated nerves of the longitudinal sections at 14 days after crush injury. As shown in Fig. 2A, the 10 × NSC-CM group had more regenerated nerve fibers than the PBS group, and the arrangement was more regular at the site of crush injury. Consistently, TEM revealed that the numbers and the thicknesses of myelinated fibers in 10 × NSC-CM-treated rats were higher than those in PBS-treated rats (Fig. 2B–F). These results indicated that NSC-CM were favorable for axon outgrowth and remyelination after the damage of sciatic nerve.
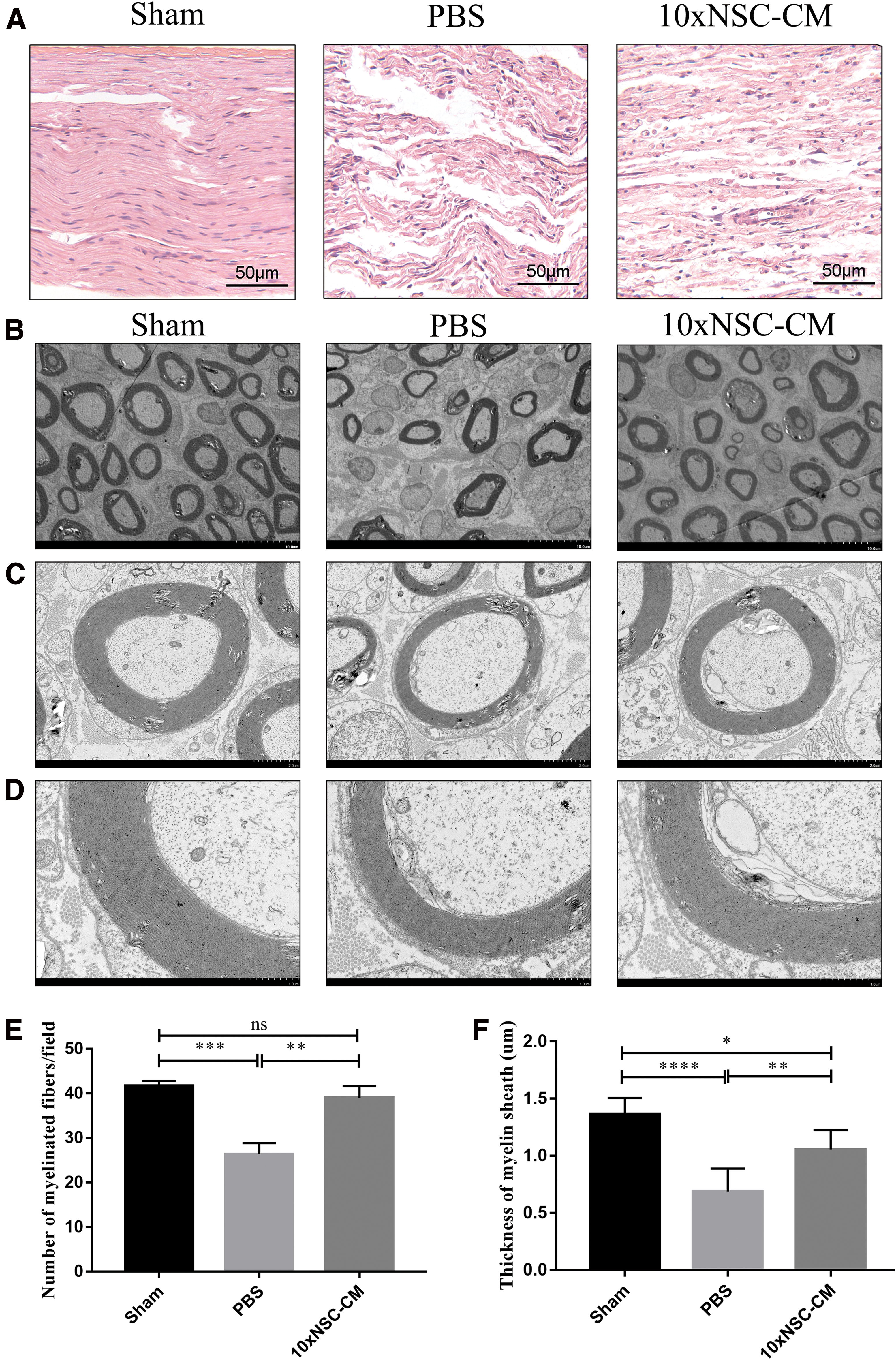
NSC-CM improves axonal regeneration and remyelination.
NSC-CM reduces inflammation after injury
The distal nerve stump was subjected to histochemical staining at 3 and 14 days after surgery. As shown in Fig. 3A–D, the distal sciatic tissue treated with PBS and 10 × NSC-CM showed increased expression of proinflammatory cytokines (IL-6, IL-1β, TNF-α) compared with the sham group, but there was no statistical difference between the two treatment groups at 3 days after injury. At 14 days after injury, the expression of proinflammatory cytokines in the 10 × NSC-CM group was significantly reduced compared with the PBS group.
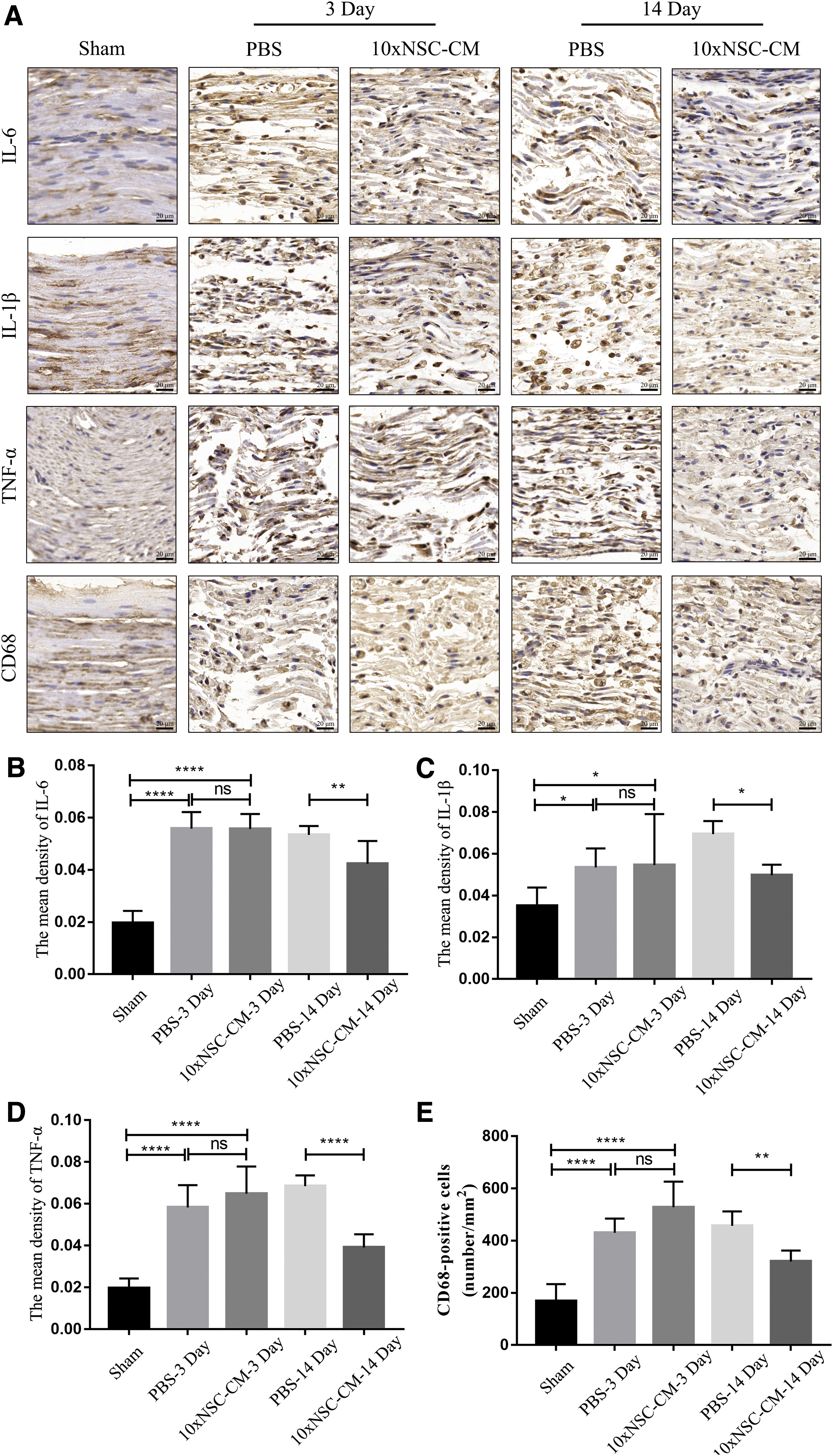
NSC-CM reduces inflammation after injury.
Moreover, 3 days after the injury, CD68-positive macrophages began to recruit in the damaged sciatic nerve, but there was no significant difference between PBS and 10 × NSC-CM treatments. However, consistent with the infiltration of proinflammatory cytokines, the recruitment of CD68-positive macrophages was significantly reduced in 10 × NSC-CM compared with PBS at 14 days after injury (Fig. 3A, E). These results indicated that NSC-CM reduced tissue inflammation and macrophages recruitment in the late stages of sciatic nerve crush injury.
NSC-CM significantly inhibits the expression of proinflammatory factors in macrophages
NSC-CM alleviated neuroinflammation after sciatic nerve injury and was associated with the recruitment of macrophages in vivo. Thus, we further explored whether NSC-CM regulates the inflammatory response of macrophage induced by LPS in vitro. The resident PMs and THP-1 were used as exploring objects. As shown in Fig. 4, pretreatment with LPS significantly increased the mRNA levels of IL-6 (Fig. 4A), IL-1β (Fig. 4B), TNF-α (Fig. 4C), and iNOS (Fig. 4D) in macrophages, while NSC-CM treatment reversed the increase of LPS-induced inflammatory genes. Consistent with expression at the transcriptional level, as shown in Fig. 4E–H, the stimulation of NSC-CM significantly reduced the protein expression of iNOS, TNF-α, and IL-1β compared with LPS pretreatment.
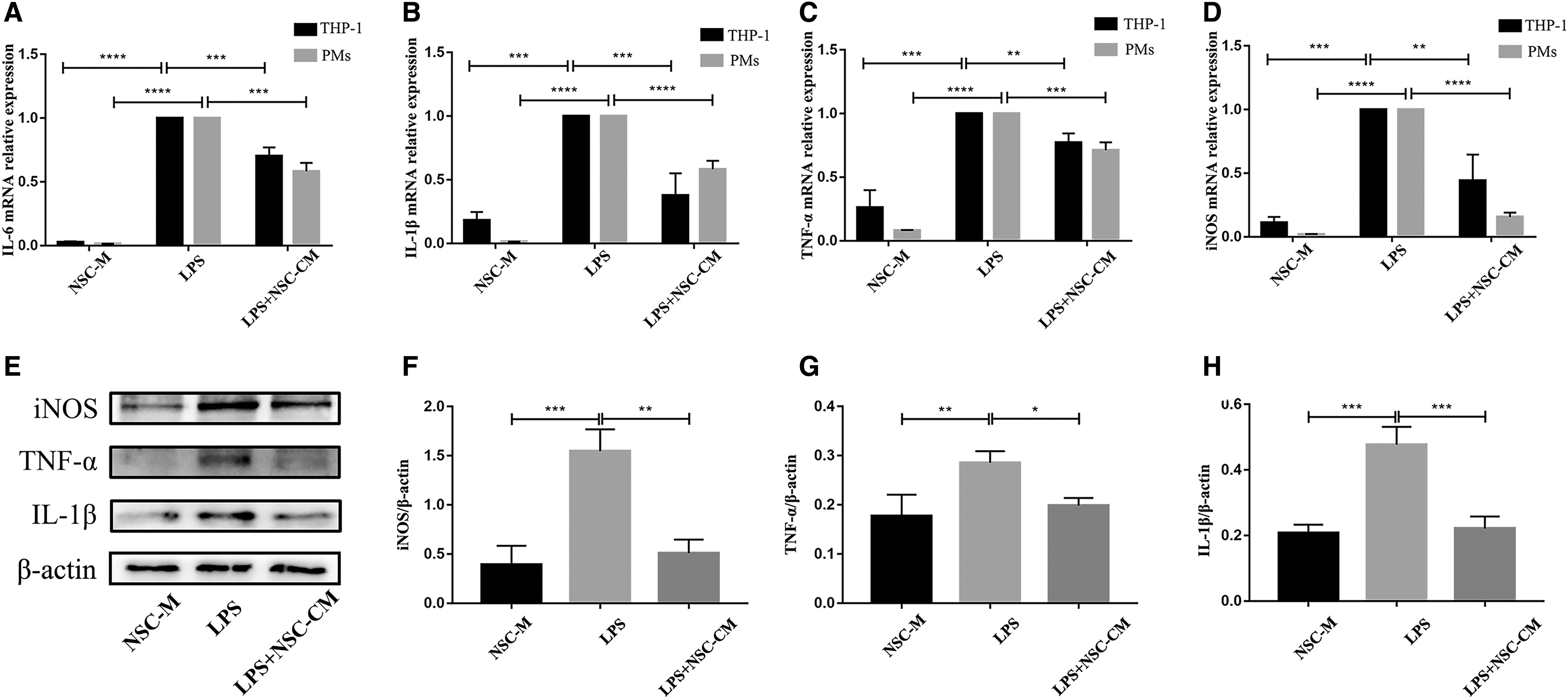
NSC-CM significantly inhibits the expression of proinflammatory factors in macrophages. THP-1 and PMs were incubated with or without LPS (100 ng/mL) for 6 h, and then with NSC-M or NSC-CM for 12 h after LPS treatments. The mRNA levels of
These results indicated that NSC-CM inhibited the proinflammatory response at the transcriptional and protein levels in macrophages.
NSC-CM activates Sirt-1 to inhibit HIF-1α and NF-κB phosphorylation
The protein levels of Sirt-1 and its downstream signaling molecules HIF-1α and NF-κB/p-NF-κB were detected. Figure 5A, B shows that NSC-CM significantly increased Sirt-1 expression in PMs, while treatment with LPS alone significantly inhibited the expression of Sirt-1. At the same time, NSC-CM inhibited the activation of downstream signaling molecules p-NF-κB and HIF-1α by activating Sirt-1 (Fig. 5A–D). These results indicated that NSC-CM inhibited HIF-1α and NF-κB phosphorylation by activating Sirt-1 in macrophages.

NSC-CM activates Sirt-1 to inhibit HIF-1α and NF-κB phosphorylation. PMs were incubated with or without 100 ng/mL LPS for 6 h, and then incubated with NSC-M or NSC-CM for 12 h.
The involvement of Sirt-1 in the inhibition of HIF-1α and NF-κB signaling is exerted by NSC-CM in macrophages
We verified whether the anti-inflammatory effect of NSC-CM was mediated through the Sirt-1 inactivation at the cellular level. The PMs were pretreated with EX527, a specific Sirt-1 inhibitor. As shown in Fig. 6A–D, the expression of Sirt-1 was suppressed by EX527, and the inhibitory effect of NSC-CM on the LPS-induced activation of NF-κB and HIF-1α was counteracted by EX527. To determine the relationship between the activation of Sirt-1 and the expression of proinflammatory factor in macrophages, we verified the expression of the downstream molecules already identified above. As shown in Fig. 6E–H, the inhibition of Sirt-1 greatly offset the inhibitory effect of NSC-CM on the expression of iNOS, IL-1β, and TNF-α. The data were confirmed by ELISA analysis of proinflammatory cytokines (IL-6 and IL-1β) secreted by macrophages (Fig. 6I, J).
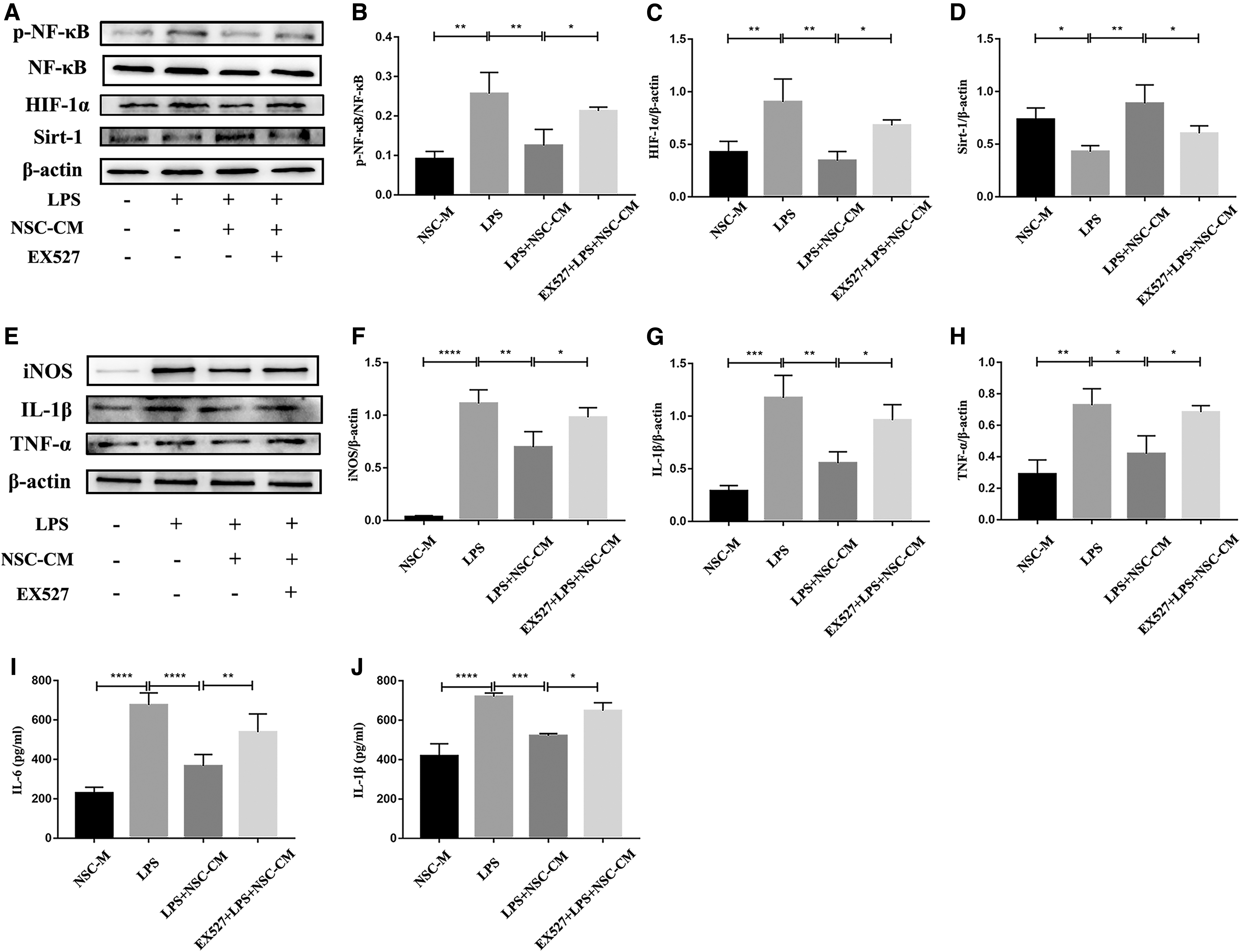
The involvement of Sirt-1 in the inhibition of HIF-1α and NF-κB signaling exerted by NSC-CM in macrophages. The PMs were pretreated with EX527 for 6 h, and incubated with or without 100 ng/mL LPS for 6 h, and then incubated with NSC-M or NSC-CM for 12 h and 24 h.
These results indicated that the inhibition of NF-κB and HIF-1α by Sirt-1 activation was involved in the suppression of NSC-CM on LPS-induced macrophage inflammatory response.
Discussion
Although the regeneration of the peripheral nervous system is more impressive than that of the central nervous system, the motor and sensory functions of regeneration often remain incomplete. And the persistent inflammatory response usually causes demyelinating disease and neuropathic pain. Therefore, there is an urgent need to develop alternative repair strategies and treatment methods [24].
The increasing evidence suggests the supportive role of stem cells in peripheral nerve regeneration after injury [25]. Many studies have demonstrated that NSCs differentiated from human embryonic-derived support cells have efficacy in rodent models of peripheral nerve regeneration [26,27].
However, stem cells transplantation still faces many limitations, such as teratoma formation, immune response, difficulty in harvesting cells, limited cell proliferation and differentiation. These major issues prevent stem cells from being routinely used in the clinic as a treatment option [28]. Derived products of stem cells such as conditioned medium and microvesicles have proven to be potentially therapeutic [29]. Previous research has also shown that stem cell-conditioned medium can repair nerve damage in rats [30]. And stem cell transplantation inhibited sciatic nerve inflammation through conditioned medium [31].
In this study, we used NSC-CM to treat crush injury of sciatic nerve in rats. We first discovered the potential therapeutic effects of conditioned medium from NSCs on PNI. We found that after NSC-CM treatment, the extent of toe extension of the affected side was closer to normal toes at 14 days after injury. And NSC-CM promoted the motor function recovery after sciatic nerve crush injury compared with PBS. Compared with the control group, NSC-CM slowed the atrophy of the gastrocnemius muscle, indicating that NSC-CM promoted neuromuscular reinnervation. Moreover, NSC-CM treatment improved axonal regeneration and remyelination.
Appropriate microenvironment is important for remyelination and axonal regrowth, and persistent inflammation hinders myelin sheath and axon regeneration [7]. Therefore, we used IHC to observe the expression of inflammatory factors in the distal sciatic stump in the early (3 days) and late (14 days) stages of injury. The inflammatory factors including IL-6, IL-1β, TNF-α, and CD68 were used to indicate inflammatory infiltration in sciatic nerve [2,7]. Importantly, we found that NSC-CM reduced inflammation in the late stages of nerve injury in vivo. And with the reduction of inflammatory expression, macrophage infiltration was also reduced in the late stages of nerve injury.
After sciatic nerve injury, the early inflammatory factors are mainly secreted by Schwann cells (SCs), which are used to recruit monocytes and tissue macrophages in the blood circulation to the injury site [1,32]. Macrophages are recruited to the injury site 3 days after the injury and reach a peak 7 days after the injury, so the inflammation in the later stage is mainly involved by macrophages. The study showed that after 10 × NSC-CM treatment, the inflammation in the early stage of injury did not change significantly, but the inflammation infiltration in the late stage of injury was suppressed. These results suggest that 10 × NSC-CM may downregulate the expression of proinflammatory factors by macrophages after sciatic nerve injury. Therefore, it is necessary to further explore the effects of NSC-CM on macrophage inflammation in vitro.
So we investigated the effect of NSC-CM on macrophage inflammation in vitro. We used LPS to induce inflammation to mimic the macrophage proinflammatory state and then treated the macrophages with NSC-CM [3,33]. The results demonstrated that NSC-CM treatment attenuated the expression of IL-6, IL-1β, TNF-α, and iNOS in macrophages induced by LPS at the transcriptional and protein levels, respectively.
Strong evidence has indicated that sirtuins, especially Sirt-1 and Sirt-2, play an important role in neuroinflammation. Increased expression of Sirt-1 inhibited LPS-induced inflammation of microglia BV-2 cells and reduced the production of a range of cytokines, and it also protected neuronal PC12 cells from caspase-3 dependent apoptosis [34,35].
Sirt-1 affects gene expression through the action of deacetylase and is closely related to a variety of neurological diseases. As a deacetylase, Sirt-1 not only participates in the deacetylation of histones but also performs deacetylation modification with certain important transcription factors and cofactors, including transcription factors such as NF-κB and iNOS, thereby suppressing inflammatory expression [36]. The lack of Sirt-1 promoted the production of proinflammatory cytokines in macrophages. The deletion of Sirt-1 resulted in a high degree of acetylation of NF-κB and therefore reduced an excessively active NF-κB signaling. In addition, Sirt-1 deactivated HIF-1α acetylated, while HIF-1α is a key transcription factor that regulates the expression of proinflammatory genes, including IL-1β [37].
In this study, we found that NSC-CM inhibited LPS-induced macrophage inflammation at the transcriptional and protein levels, respectively, and found that the inhibitory effect of NSC-CM on macrophage inflammation was achieved by activating the macrophage Sirt-1 signaling pathway. At the same time, NSC-CM treatment changed the expression levels of NF-κB and HIF-1α, which are the downstream of Sirt-1 signaling pathway. To verify whether the expression of inflammatory factors was inhibited by NSC-CM works through Sirt-1, we used Sirt-1-specific inhibitor EX527 to pretreat PMs [38]. The results showed that the inhibitory effect of NSC-CM on the LPS-induced activation of NF-κB and HIF-1α was counteracted by EX 527, suggesting a role of Sirt-1 in the modulatory effect of NSC-CM on macrophage proinflammation.
In general, our experimental results showed that NSC-CM promotes functional recovery after sciatic nerve crush injury in vivo and inhibits macrophage proinflammatory by activating Sirt-1 signaling pathway in vitro. And above all, the application of NSC-CM may be a potential treatment strategy for neuroinflammatory diseases. Certainly, we will further explore the mechanism by which NSC-CM activates Sirt-1 expression in macrophages in later studies.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by the National Natural Science Foundation of China (grant no. 81571221), a project funded by the Priority Academic Program Development of Jiangsu Higher Education Institutions (PAPD) and the Qing Lan Project of Jiangsu Province.