
Abstract
Pretransplant normothermic machine perfusion (NMP) of donor kidneys offers the unique opportunity to perform active interventions to an isolated renal graft before transplantation. There is increasing evidence that mesenchymal stromal cells (MSCs) could have a paracrine/endocrine regenerative effect on ischemia-reperfusion injury. The purpose of this study was to determine which cytokines are secreted by MSCs during NMP of a porcine kidney. Viable porcine kidneys and autologous whole blood were obtained from a slaughterhouse. Warm ischemia time was standardized at 20 min and subsequent hypothermic machine perfusion was performed during 2–3 h. Thereafter, kidneys were machine perfused at 37°C during 7 h. After 1 h of NMP, 0, 107 cultured human adipose tissue-derived MSCs, or 107 cultured bone marrow-derived MSCs were added (n = 5 per group). In a fourth experimental group, 7-h NMP was performed with 107 adipose tissue-derived MSCs, without a kidney in the circuit. Kidneys perfused with MSCs showed lower lactate dehydrogenase and neutrophil gelatinase-associated lipocalin levels in comparison with the control group. Also, elevated levels of human hepatocyte growth factor, interleukin (IL)-6, and IL-8 were found in the perfusate of the groups perfused with MSCs compared to the control groups. This study suggests that MSCs, in contact with an injured kidney during NMP, could lead to lower levels of injury markers and induce the release of immunomodulatory cytokines.
Introduction
To date, kidney transplantation is regarded as the best treatment for patients with end-stage renal failure. In an attempt to reduce the waiting time by increasing the deceased donor organ pool, the use of donor organs from donation after circulatory death donors and extended criteria donors plays an important role [1]. However, many of these organs are older and of suboptimal quality, rendering kidneys more susceptible to ischemia-reperfusion injury (IRI) when compared to the organs derived from living donors [2].
Preserving the function of ischemically damaged allografts is of crucial importance for successful transplantation. The basic principle of organ preservation is to prevent ischemic damage by replacing the circulating blood with a preservation solution, followed by retrieving and cooling the organ. Static cold storage is the most widely used organ preservation method for deceased donor kidney grafts. However, prolonged cold ischemia time is associated with a higher risk of delayed graft function and poorer allograft survival [3]. Ex vivo cold (1°C–8°C) machine perfusion is now considered to be the method of choice for preserving higher risk kidney allografts obtained from deceased donors [4].
Pretransplant normothermic (37°C) machine perfusion (NMP) allows for metabolic activity of renal cells. Therefore, it may enable better quality assessment of donor kidneys during preservation, by providing information on renal hemodynamics and allowing for sample collection of perfusate, urine, and tissue to quantify graft injury and function [5,6]. In addition, it provides a platform to safely expand the preservation time and potentially improve the condition of kidney grafts, for example, by administering ex vivo pharmacological agents or cellular therapy. Several autotransplantation studies in pigs have shown positive effects of NMP on graft function, leading to an increasing number of clinical studies investigating NMP [7 –9].
Due to the unique combination of immunomodulatory properties and abilities of self-renewal, mesenchymal stromal cells (MSCs) and MSC-derived products have attracted substantial interest as promising cellular therapeutic agents to improve clinical outcome of kidney transplantation [10,11]. MSCs are a population of multipotent stromal cells that can be isolated from different sources, including adipose tissue (A) and bone marrow (BM). MSCs have been shown to exert a wide range of anti-inflammatory and immunomodulatory effects on cells of the innate and adaptive immune system both by cell-to-cell contact and by secreting biologically active substances, such as chemokines, cytokines, and growth factors, which have antiapoptotic, antifibrotic, anti-inflammatory, and mitogenic features. MSCs are assumed to be beneficial to reduce transplantation-related IRI [12]. MSCs from different sources present differences in the expression of surface markers and also in cytokine, chemokine, and growth factor production [13 –15]. Although morphologically identical, proliferation rate and differentiation capacity are not completely similar [16]. There is also evidence that the therapeutic potential of BM-MSCs and A-MSCs could be different [17], and to date, it remains unclear which source of MSCs is most suitable to target IRI.
The administration of MSCs at the time of implantation or shortly after in vivo reperfusion of a donor kidney may reduce the severity of injury and might accelerate regenerative and reparative processes. As there is little evidence that MSCs can home to damaged tissue, anatomically targeting a specific (transplanted) organ by means of systemic intravenous cell therapy administration to the transplant recipient is difficult to accomplish. Two clinical studies, in which MSCs were administered intravenously after renal transplantation, focused on the immunomodulatory aspects of MSCs and reported signs of systemic immunosuppression, but did not take an in-depth look into reparative and regenerative potential of these cells on the kidney [18,19]. MSCs can be delivered directly to the donor kidney relatively easily, without exposing the entire patient/animal to MSCs, through ex vivo NMP by infusing the cells into the afferent tubing toward the renal vasculature. The first study to report infusion of MSCs into the renal artery during sub-NMP of human kidneys was performed by Brasile et al. In their study, MSCs of undefined source and origin were administered to human discarded kidneys leading to a reduced inflammatory state and an increased renal metabolism [20].
In our previous work, we showed that A-MSCs or BM-MSCs that are infused during NMP are predominantly retained in glomerular capillaries and a proportion of those cells will remain viable and detectable after 7 h of ex vivo perfusion [21]. The purpose of this study was to determine if ex vivo infused A-MSCs and BM-MSCs are viable and functional during NMP and which cytokines are secreted into the perfusate as a result of the addition of MSCs during NMP of porcine kidneys.
Materials and Methods
Kidney and blood retrieval
Viable porcine kidneys (sow; type: Topigs 20) and autologous blood were obtained from a slaughterhouse in Groningen (Kroon Vlees, Groningen, The Netherlands). After circulatory arrest, kidneys were subjected to 20 min of warm ischemia before flush with 180 mL of cold saline (NaCl 0.9%; Baxter, Utrecht, The Netherlands). Approximately 2.5 L of blood was collected in a beaker containing 25.000 IU heparin (LEO® pharma; Ballerup, Denmark). Excess fatty tissue was removed from the kidney. Subsequently, the kidney was connected to a Kidney Assist Transport device (Organ Assist, Groningen, The Netherlands) for oxygenated hypothermic machine perfusion (HMP) at 4°C with University of Wisconsin solution for machine perfusion (Belzer UW-MP solution; Bridge to Life Ltd., Columbia) at a mean pressure of 25 mmHg for 2–3 h. The whole autologous blood was leukocyte depleted using a BioR leukocyte filter (BioR 02 plus BS PF; Fresenius Kabi, Zeist, The Netherlands) and centrifuged for 20 min at 3,000 g. Next, the supernatant plasma was removed and the erythrocytes were washed with phosphate-buffered saline, after which a pure red blood cells (RBC) concentrate was obtained.
Experimental groups
Four experimental groups were defined (n = 5 per group). Kidneys were allocated to each group before the start of the study and experiments were planned in such a way that those belonging to the same group were not subsequently clustered in time. In three groups, the kidneys were perfused at 37°C for 7 h. After 1 h of NMP, 0, 107 A-MSCs, or 107 BM-MSCs were added to the perfusion circuit. To determine the isolated effect of machine perfusion on MSCs and the production of cytokines, an additional experimental group was created in which 7-h perfusions at 37°C were performed without a kidney (also n = 5). In this group, 107 A-MSCs were added to the perfusion circuit after 1 h of NMP and left to circulate during the remaining 6 h. For the group without a kidney, we chose to test A-MSCs as it was easier to generate a high number of A-MSCs because large amounts of adipose tissue were available.
NMP setup
After 2–3 h of HMP, kidneys were disconnected from the cold perfusion device. An 8 Fr cannula (Vygon, Valkenswaard, The Netherlands) was introduced into the ureter and a SealRing cannula (Organ Recovery Systems, Itasca, IL) was placed around the aortic patch of the kidney. Next, kidneys were connected to a perfusion setup consisting of a LifePort® organ chamber with cradle (also Organ Recovery Systems), a centrifugal pump with magnetic pump head (Deltastream DP2; Medos Medizintechnik AG, Stolberg, Germany), and an oxygenator with an integrated heat exchanger (HILITE 1000 Infant Hollow Fibre Oxygenator; Medos Medizintechnik AG). This system had been primed with the perfusion solution, which was oxygenated with 95% oxygen/5% carbon dioxide (carbogen) at 0.5 L/min. The temperature of the perfusion solution could be adjusted using a temperature-controlled water bath. All kidneys were perfused in a pressure-controlled, sinusoid pulsatile manner at a fixed pressure of 110/70 mmHg during all experiments. Kidneys were connected to the NMP setup when the circulating perfusate had reached a temperature of 30°C. Kidneys were then gradually rewarmed to 37°C in the first 60 min of NMP. After these 60 min, 0, 107 A-MSCs, or 107 BM-MSCs were added to the circuit. In the A-MSC group without a kidney, an artificial resistance was connected to the arterial tubing to imitate the average vascular resistance of a kidney.
Perfusion solution
The perfusion solution consisted of 288 mL of pure erythrocytes (final hematocrit between 0.37 and 0.40 L/L), 100 mL of 20 g/L human albumin (Albuman 20 g/L; Sanquin Plasma Products BV, Amsterdam, The Netherlands), 300 mL sodium chloride 0.9% (Fresenius Kabi Nederland BV, Zeist, The Netherlands), 8 mL sodium bicarbonate 8.4% (B. Braun Melsungen AG, Melsungen, Germany), 4.8 mL calcium gluconate 10% (also B. Braun), 9.6 mL glucose 5% (Baxter BV, Utrecht, The Netherlands), 8 IU insulin (Novo Nordisk A/S, Bagsværd, Denmark), 0.016 g mannitol (Sigma Aldrich, Zwijndrecht, The Netherlands), 1,000 μM creatinine (also Sigma Aldrich) ,and 8 mL of amoxicillin/clavulanate 1,200 mg/20 mL sterile water (Sandoz BV, Almere, The Netherlands).
Before the start of NMP, a bolus of 1 mL of 2.5 mg/mL verapamil (also Sandoz BV) was added to the perfusate through the arterial line. During perfusion, 1.6 mL of 1,200 mg/20 mL Augmentin was added every hour. Also, hourly Sterofundin (also B. Braun) was added to the perfusate to account for lost volume as a result of sampling. During perfusion, glucose 5% solution was added if perfusate glucose levels became lower than 4.5 mM and calcium was adjusted if the perfusate ionic concentration was lower than 1.1 mM.
Isolation and culturing of MSCs
The choice to use human MSCs during this experiment was based on the idea that we would test the actual future clinical product. Due to the absence of an intact immune system, xeno-effects are unlikely to occur in our ex vivo perfusion setup, with a perfusate that does not contain any leukocyte or plasma. To obtain A-MSCs, these cells were isolated from human perirenal fat, a clinical waste product derived from weekly living donor nephrectomies in the University Medical Center Groningen. First, 2 mL of Hank's Balanced Salt Solution supplemented with 1% p/s (100 U/mL penicillin and 100 mg/mL streptomycin) was added to the adipose tissue. Next, the tissue was cut into small pieces and enzymatically digested using 0.5 mg/mL collagenase type IV (Life Technologies) dissolved in RPMI medium (RPMI 1640 medium+GlutaMAX™; Life Technologies) supplemented with 1% p/s during 30 min at 37°C. After centrifugation, the cell pellet was resuspended in minimal essential medium eagle-alpha supplemented with 10% fetal bovine serum, 1%
Clinical-grade BM-MSCs were obtained from a human volunteer donor. Informed consent was obtained according to local ethical review board-approved procedures. Using aseptic techniques according to hospital protocols, bone marrow was aspirated under full anesthesia. After filtration, the bone marrow aspirate was diluted in a 2% albumin/NaCl solution (Sanquin Plasma Products BV). Next, bone marrow mononuclear cells (BM-MNC) were obtained using Ficoll density separation. BM-MNCs were plated in T175 flasks and expanded in Dulbecco's modified Eagle's medium supplemented with 10% fetal bovine serum of a selected batch (GE Healthcare) and antibiotics. At confluency, the cells were harvested using TripleSelect (Invitrogen) and expanded.
For each experiment, an MSC suspension from freshly trypsinized MSCs (passage 2–4) of three pooled donors was prepared ∼30 min before infusion. The cells were dissolved in 10 mL Sterofundin in a syringe. After 60 min of NMP, MSCs were added in a time span of 10 s through the arterial sample port, which was located a few centimeters from the renal artery.
Characterization of MSCs
To confirm that the cells cultured were indeed MSCs, we performed flow cytometric analysis on the cells that were used during the experiments on a BD FACS Canto II (BD Biosciences Nederland BV, Vianen, The Netherlands). MSCs were stained with FITC Mouse Anti-Human CD31, PE Mouse Anti-Human CD73, PerCP-Cy5.5 Mouse Anti-Human CD90, and APC Mouse Anti-Human CD105 (all BD Biosciences Nederland BV). As negative control, unstained MSCs were used.
Perfusate and urine analysis
Hourly, arterial perfusate and urine samples were taken, and arterial blood gas samples were analyzed. Perfusion parameters such as temperature, renal blood flow, renal resistance, and urine production were documented every 30 min. Standard clinical assays were used to measure the concentration of sodium, potassium, creatinine, urea, calcium, magnesium, phosphate, chloride, lactate, albumin, glucose, lactate dehydrogenase (LDH), and aspartate aminotransferase (ASAT). These values were used to calculate fractional sodium excretion (FENa+) and creatinine clearance. Thiobarbituric acid-reactive substances' (TBARS) levels in the perfusate were determined by measurement of the concentration of malondialdehyde (MDA) using a quantitative assay (Sigma). The concentration of N-acetyl-β-
Multiplex analysis
Concentrations of interleukin-6 (IL-6), interleukin-8 (IL-8), interleukin-10 (IL-10), interleukin-17 (IL-17), vascular endothelial growth factor A (VEGF-A), vascular endothelial growth factor c (VEGF-C), platelet-derived growth factor AA (PDGF-AA), monocyte chemoattractant protein 1, chemokine (C-C motif) ligand 2 (CCL2), chemokine (C-C motif) ligand 4 (CCL4), chemokine (C-X-C motif) ligand 10 (CXCL10), angiopoietin-1, endothelin-1, interferon gamma (IFN-γ), tumor necrosis factor alpha (TNF-α), fibroblast growth factor (FGF), hepatocyte growth factor (HGF), kidney injury molecule-1 (KIM-1), and neutrophil gelatinase-associated lipocalin (NGAL) were measured using a human cytokine/chemokine magnetic bead panel multiplex assay according to the manufacturer's protocol (Merck Millipore, Billerica, MA and R&D Systems, Minneapolis, MN). Briefly, a mixture containing all antibody-coated magnetic beads was prepared and incubated overnight at 4°C. Next, a biotinylated detector antibody and a streptavidin-conjugated fluorescent protein were added. Fluorescence was measured on a Luminex® 100/200 cytometer (Luminex, Austin, TX) using Xponent software.
Histology
At t = 0 and t = 240 min of NMP, needle biopsies were taken with a BioPince™ device (Argon Medical Devices Netherlands BV, Maastricht, The Netherlands) and fixed in formalin and RNAlater (Sigma). At t = 420 multiple surgical tissue samples were taken from the upper, lateral, and lower cortex. These biopsies were also fixed in formalin, RNAlater (also Sigma), or liquid nitrogen. The formalin-fixed, paraffin-embedded biopsies were stained with Periodic Acid-Schiff. These biopsies were examined and a damage scoring system was developed, based on previously published scoring systems [22]. As differences in histology between upper, lateral, and lower pole were very minimal, per experiment, three cortical biopsies per kidney of only the upper pole at t = 0, t = 240, and t = 420 were scored on a scale of 0–3 on glomerular dilatation, tubular dilatation, and tubular necrosis (0 = none; 1 = mild; 2 = moderate; and 3 = severe) by two independent and blinded observers. In case of disagreement about the score, findings were discussed until a consensus was reached.
Statistics
For all cytokines and other continuous longitudinally measured variables, the area-under-the-curve (AUC) of their release in time was calculated. A one-way ANOVA was used to compare AUC values between groups if the data had homogeneity of variances and were normally distributed, tested by means of a Bartlett and d'Agostino-Pearson test. If data were not normally distributed and/or did not have homogeneity of variances, the Kruskal–Wallis test was used. For post hoc comparisons, in case of an ANOVA, an unpaired t-test was utilized. A Mann–Whitney test was used for post hoc comparisons if a Kruskal–Wallis test had been used. Two-sided P values of 0.05 or lower were assumed to indicate statistical significance.
Results
Characterization of MSCs
MSCs were plastic adherent and showed a typical spindle-shaped morphology. A-MSCs and BM-MSCs were analyzed using flow cytometric analysis and showed expression of markers CD105, CD73, and CD90 and no expression of marker CD31.
Perfusion parameters
Arterial flow pattern was similar in all groups with a kidney in the circuit. The group without a kidney was corrected by calculating the average weight of all kidneys from the other groups. Figure 1 shows the average perfusate flow rate of every group in mL/min/100 g. The groups with a kidney in the perfusion circuit showed a small flow peak at t = 60, with a mean value of 165 ± 103 mL/min in the control group, 175 ± 52 mL/min in the BM-MSC group, and 182 ± 42 mL/min in the A-MSC group at this time point. During the remaining NMP time, the flow decreased slowly. A one-way ANOVA did not show a significant difference between the groups.
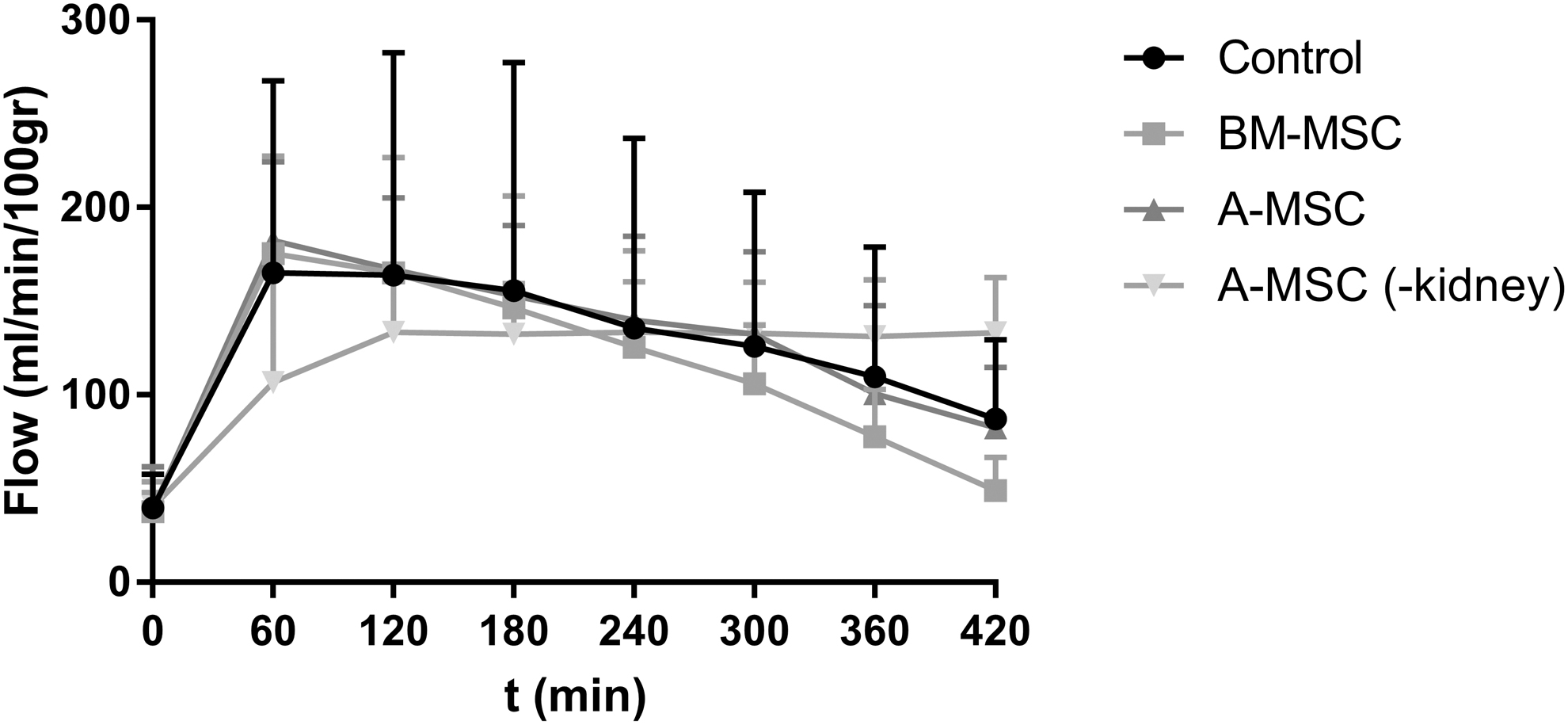
Arterial flow (mL/min/100 g) of the four experimental groups during NMP (mean ± SD). NMP, normothermic machine perfusion; SD, standard deviation.
Urine and perfusate analysis
Functional markers
All kidneys produced urine during the 7 h of NMP and there were no significant differences in cumulative diuresis between the experimental groups. Mean cumulative diuresis at t = 420 was 165 ± 120 mL in the control group, 135 ± 92 mL in the BM-MSC group, and 169 ± 117 mL in the A-MSC group.
Figure 2A shows the creatinine clearance in mL/min in every group. Although the A-MSC group showed higher levels of creatinine clearance with mean levels of 6.33 ± 1.63 mL/min, this did not reach statistical significance in comparison with the control group (2.68 ± 0.37 mL/min) and BM-MSC group (2.48 ± 1.01 mL/min). Comparisons in AUC did not show any significant difference either (Fig. 2B).
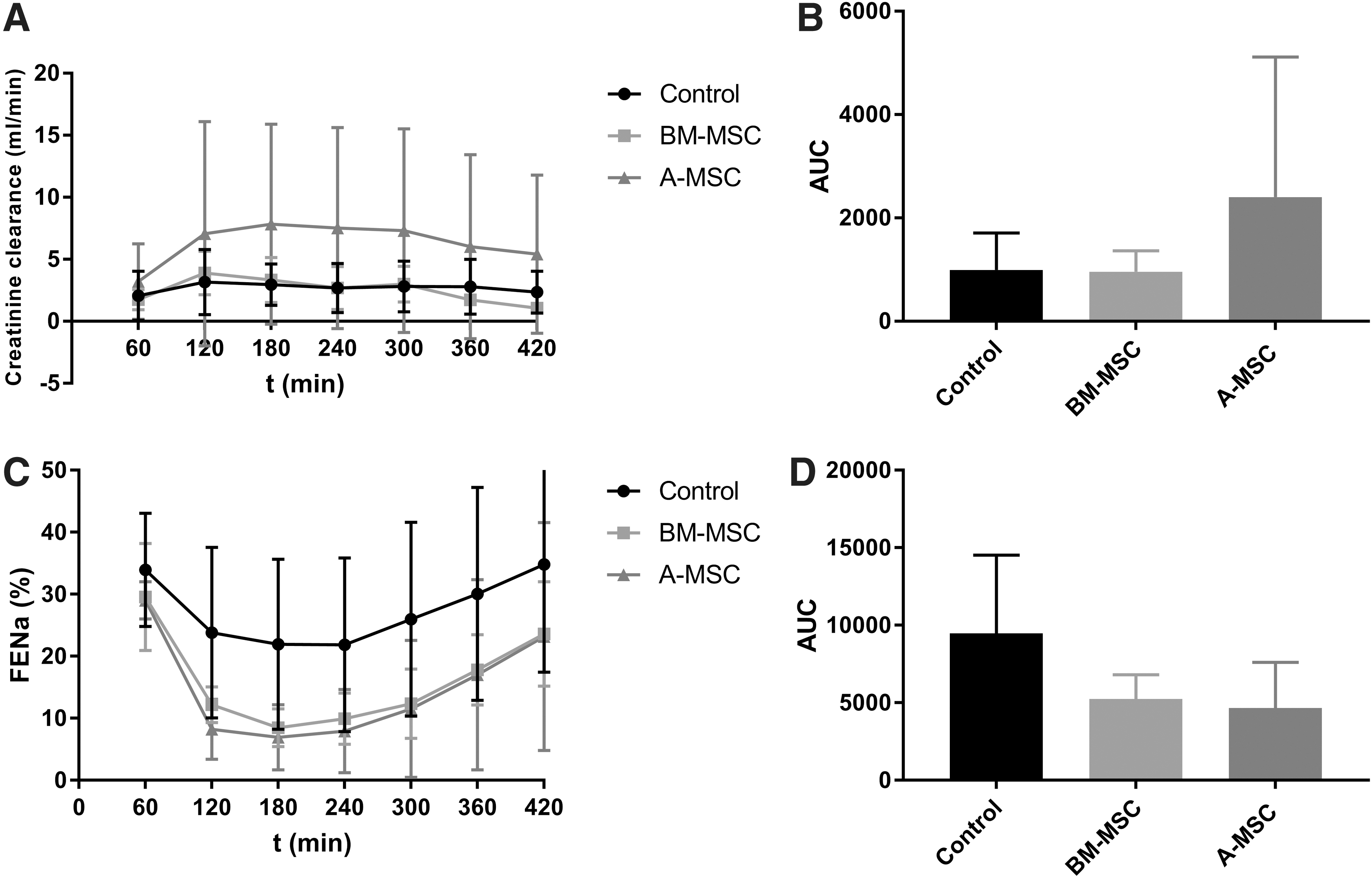
Functional markers of the three experiments with a kidney in the perfusion circuit during NMP.
FENa+ levels of each group are presented in Fig. 2C. Every group, with the steepest decline in the MSC groups, showed a decrease in the first 180–240 min, after which the FENa+ increased again. Mean values during perfusion were 27.5% ± 5.5% in the control group, 16.3% ± 7.8% in the BM-MSC group, and 14.8% ± 8.6% in the A-MSC group. A one-way ANOVA comparing AUCs per group did not show significant differences between the experimental groups (Fig. 2D).
Damage markers
TBARS, quantified by the concentration of MDA, were measured in all four groups as a marker for lipid peroxidation. In all groups, levels at the start of perfusion were ∼2 μM and increased to values between 8 and 12 μM after 420 min of NMP (Fig. 3A). There were no significant differences in AUC between the groups (Fig. 3B).
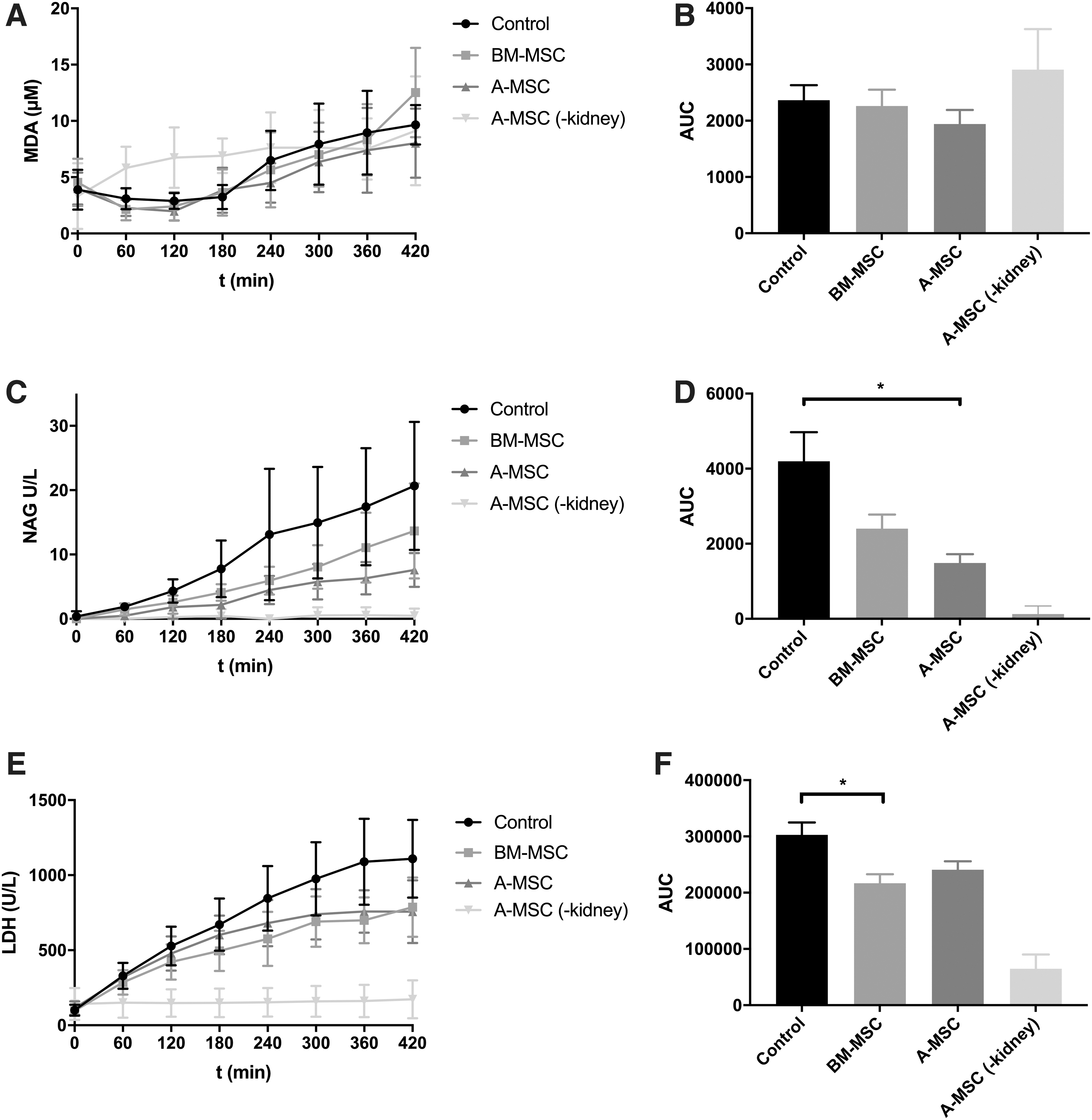
Damage markers measured in the perfusate of the four experimental groups during NMP.
To quantify tubular damage, the concentration of NAG in the perfusate and urine was measured. Figure 3C shows the concentration of NAG in the perfusate in U/L. Values of the group without a kidney were considered baseline values. The control group (without MSCs) had a significantly higher perfusate NAG concentration in comparison with the A-MSC group (P < 0.05) (Fig. 3D). There were no other significant differences between the groups. The groups had similar urinary secretion of NAG, which increased after 300 min in the control and A-MSC group, while it remained stable in the BM-MSC group. A one-way ANOVA on the AUC showed no significant difference between the groups (Supplementary Fig. S1).
The concentration of ASAT in the control group increased more than in the other groups, and in comparison with the A-MSC group, this almost reached statistical significance (P = 0.067) (Supplementary Fig. S1). The mean values of ASAT during NMP was 311 ± 229 U/L in the control group, 182 ± 85 U/L in the BM-MSC group, and 153 ± 80 U/L in the A-MSC group. The LDH values showed a similar pattern as the ASAT values (Fig. 3E). However, the LDH values in BM-MSC group were significantly lower than those in the control group (P < 0.05), and the difference between the control group and the A-MSC group was at the threshold of statistical significance (P = 0.051) (Fig. 3F). ASAT and LDH levels of the group without a kidney were considered baseline values.
Multiplex analysis
Using a magnetic bead panel multiplex assay, the concentrations of different proteins in the perfusate were measured. Renal injury markers KIM-1, endothelin-1, and NGAL were only measured in the groups with a kidney in the perfusion circuit. KIM-1 levels remained under the detection threshold. NGAL concentration was significantly higher in the control group in comparison with the BM-MSC group (P < 0.05) and the A-MSC group (P < 0.05) (Fig. 4A, B). Endothelin-1 levels in the BM-MSC group were significantly higher than in the A-MSC group (P < 0.05). There were no other statistically significant differences between groups (Fig. 4C, D).
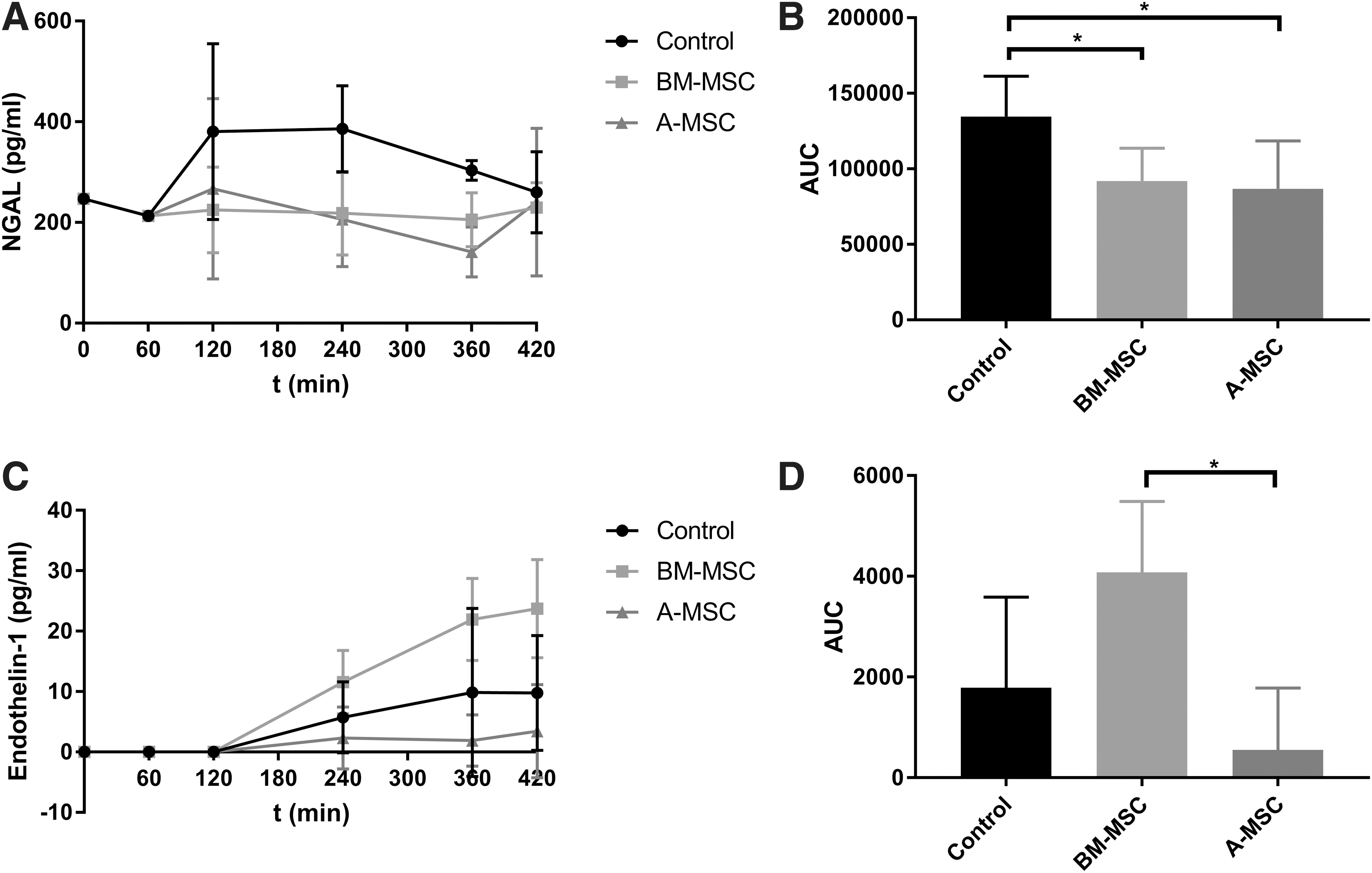
Luminex damage markers in the perfusate of the three experimental groups with a kidney in the circuit during NMP.
Growth factors
There were no significant differences in angiopoietin-1, VEGF-A, and VEGF-C levels between groups. FGF and PDGF showed no significant difference between the groups, except for the comparison of the groups with and without a kidney in the perfusion circuit (Supplementary Fig. S2).
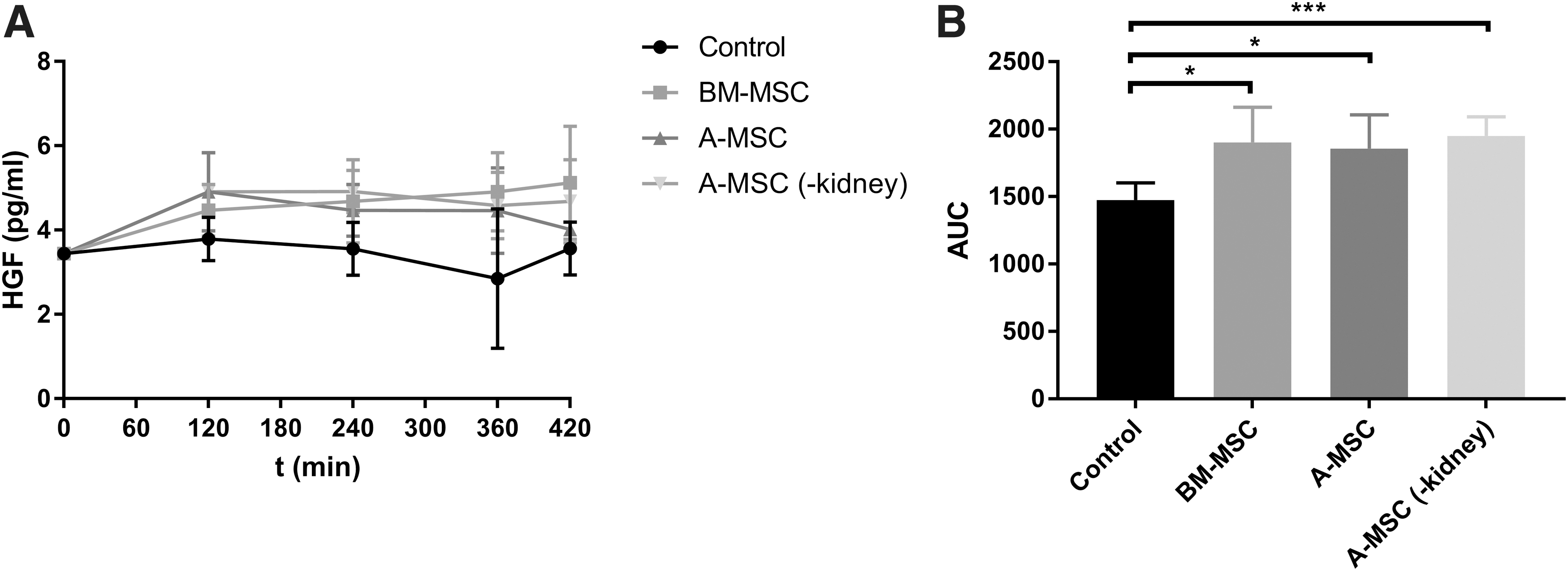
Figure 5A shows the average HGF concentrations in the different groups. The concentration in the control group was lower when compared to the BM-MSC group (P < 0.05), the A-MSC group (P < 0.05), and the A-MSC without a kidney (P < 0.01) (Fig. 5B).
Cytokines and chemokines
CXCL10, IL-17, and IFN-γ levels were undetectable. There were also no significant differences between groups concerning levels of CCL4, CCL2, IL-10, and TNF-α. These graphs can be found in the Supplementary Fig. S3.
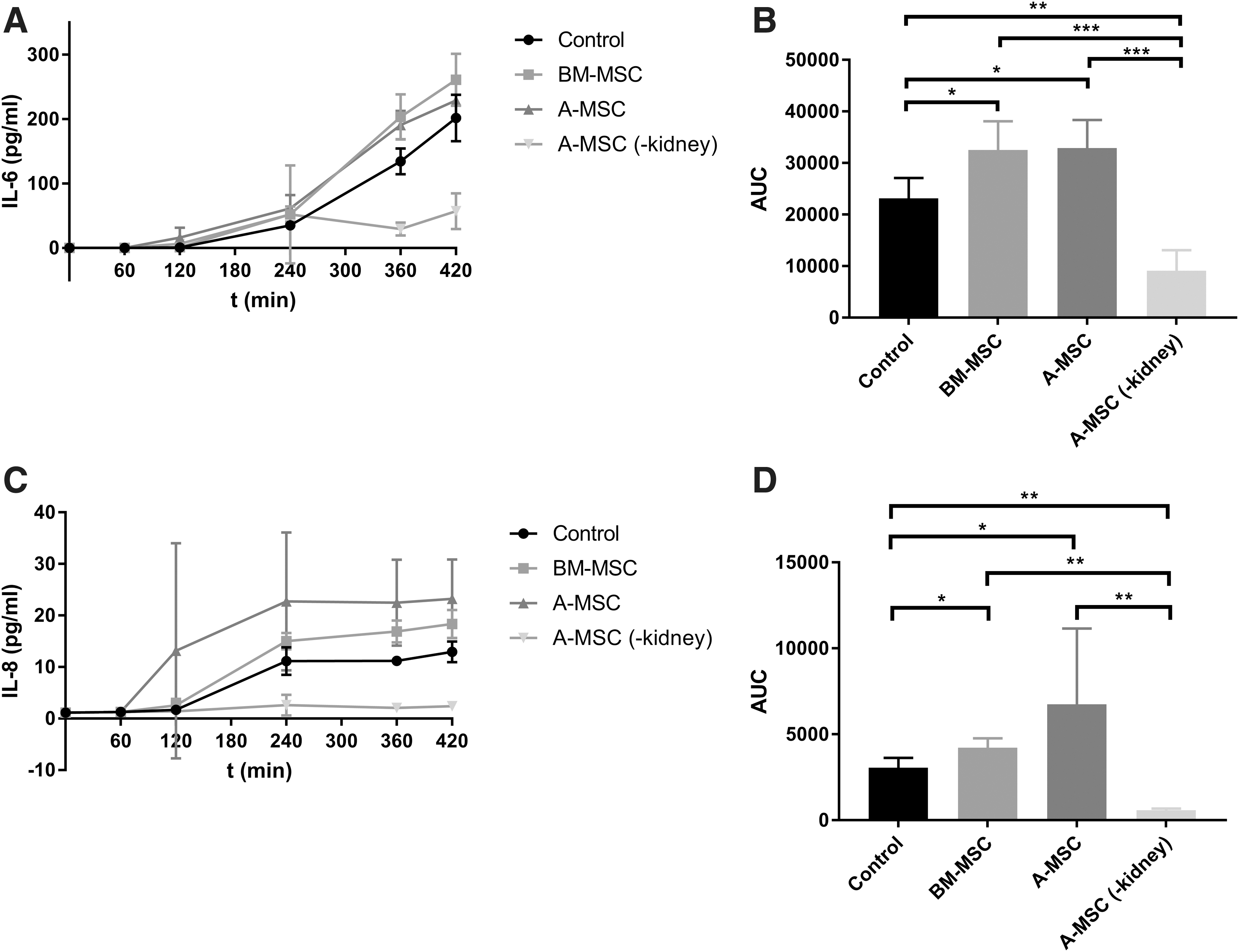
IL-6 levels increased during NMP and were elevated in the kidneys treated with MSCs (Fig. 6A). Lowest levels of IL-6 were measured in the group with A-MSCs without a kidney. Of the three experimental groups with a kidney in the perfusion circuit, the concentration in the control group was significantly lower than the BM-MSC group (P < 0.05) as well as the A-MSC group (P < 0.05). Analysis of the AUC also revealed that IL-6 levels were lower in the A-MSC group without a kidney in comparison with the BM-MSC group and the A-MSC group (both P < 0.001) and the control group (P < 0.01) (Fig. 6B).
Cytokines in the perfusate of the four experimental groups during NMP.
The average concentrations of IL-8 in the perfusate of the different groups can be found in Fig. 6C. IL-8 concentration was very low in the A-MSC group without a kidney. The highest levels of IL-8 were found in the A-MSC group with a kidney, although there was a large degree of variation. The concentration of IL-8 was significantly lower in the control group in comparison with the BM-MSC group (P < 0.05) and the A-MSC group (P < 0.05). The IL-8 levels were also significantly lower in the A-MSC group without a kidney in comparison with the control and BM-MSC group (both P < 0.01) and the A-MSC group (P < 0.01).
Histology
Of each experiment, cortical biopsies of the upper pole were scored on a scale of 0-3 for glomerular dilatation, tubular dilatation, and tubular necrosis. There were no significant differences in scores between groups after 0, 240, and 420 min of NMP (Supplementary Fig. S4).
Discussion
The aim of this study was to determine whether infused MSCs have an effect on renal function and to analyze which cytokines are secreted into the perfusate as a result of the addition of MSCs during renal NMP. Based on previous work by other groups, we hypothesized that MSCs remain viable and functional during NMP and that they may lay a base for regenerative processes by inducing release of growth factors and cytokines.
In this study, we did not find any significant difference between groups in functional markers, such as creatinine clearance and FENa+ during NMP, nor in the histological evaluation of kidneys. When evaluating kidney function over time, the A-MSC group seemed to perform better than the other groups, but this was not statistically significant. Levels of damage markers such as MDA were expected to be lower in the MSC groups as MSCs are known to be able to suppress oxidative stress [23]. This was, however, not the case in our data, with relatively high MDA levels in the group with MSCs without a kidney in comparison with the other groups. Nevertheless, NAG, a marker of tubular damage, was significantly lower in the A-MSC group in comparison with the control group. LDH and NGAL levels were significantly lower in the MSC groups in comparison with the control group. These results suggest that kidneys in the MSC groups appeared to have suffered less injury.
As regard to our analysis of growth factors, only HGF levels were significantly different between groups. In the kidneys of the A-MSC and BM-MSC group, levels were significantly higher than in the control group. As the A-MSC group without a kidney also showed a significantly higher concentration, it is most likely that secretion of this growth factor was a result of the presence of MSCs during NMP, and not necessarily the interaction between MSCs and injured renal tissue. The exact implication of this finding remains unclear. Previous research has shown that HGF can have mitogenic or trophic effects on many cell types [24]. Furthermore, there is evidence that high HGF levels are associated with better late graft function [25].
During the 7 h of NMP, cytokine levels of IL-8 and IL-6 were significantly higher in the kidneys treated with MSCs. IL-8 is an inflammatory cytokine with proangiogenic properties, which can in turn also induce VEGF production [26]. Thus, it could facilitate revascularization of injured tissues. IL-6 is mostly regarded as a more general proinflammatory cytokine, but also possesses anti-inflammatory properties. It has been found that IL-6 suppresses the secretion of proinflammatory cytokines such as IL-1 and granulocyte macrophage colony-stimulating factor (GM-CSF), and that it induces the synthesis of glucocorticoids [27]. IL-6 may play an important regenerative role: in mice, IL-6 stimulates regenerative proliferation in the epithelium of the gut, and when IL-6 is absent, an aggravation of inflammation occurs [28]. It is unclear whether IL-6 can stimulate the regeneration of kidney tissue. An important observation is that in the A-MSC group without a kidney, there was no secretion of IL-6 and IL-8. Hence, our data suggest that the secretion of these ILs will only take place in the presence of an injured kidney. It remains unclear if the MSCs induce the production of these cytokines by the kidney or that the injured kidney induces their secretion by MSCs and if the secretion of these cytokines is beneficial to the injured kidney. As both porcine and human gene primers gave signals in the human control samples, quantitative polymerase chain reaction (qPCR) could not be used to discriminate between the source of cytokines. Potentially, the only way to discriminate between the source of cytokines in future experiments is the use of heavy labeling during cell culture and subsequent mass spectrometry analysis.
In this study, we did not find an association between the addition of MSCs and very early renal function during NMP. To date, only a few preclinical studies have been performed using MSCs in combination with renal transplantation. A study in which MSCs/MSC-derived extracellular vesicles were administered during ex vivo HMP of isolated rat kidneys showed that, overall these kidneys suffered less ischemic damage [29]. The levels of damage markers such as LDH were lower in the kidneys that received this therapy, as was also the case in our study. The results of other preclinical studies using MSCs in kidney transplantation [30 –32] are difficult to compare to our study as in those other studies, MSCs were injected into the renal artery directly after reperfusion of the transplanted kidney or several days after transplantation and not in an ex vivo setting. The results of several studies show that the injection of MSCs leads to less fibrosis and improved function of glomeruli and tubules after 3–6 months [31,33]. For a fully reliable comparison to these aforementioned studies, a future study should be performed in which ex vivo MSC-treated porcine kidneys are actually transplanted to allow for a much longer follow-up in vivo.
In our study, we found no convincing difference in effect of A-MSCs and BM-MSCs during NMP. Until now, most research has been performed using only one source of MSCs. Very few analyses have been published that directly compare MSCs from different sources. The scarce data that have been published comparing BM-MSCs and A-MSCs show that the immunosuppressive potential of BM-MSCs may be inferior to that of A-MSCs [34,35]. As such effects have previously only been studied in vitro, further research is necessary to evaluate if differences in MSC modes of action can also be confirmed in vivo. In addition, future mechanistic studies could focus on the regenerative properties of different types of MSCs, rather than merely on immunomodulatory properties.
The choice to use human MSCs was based on the idea we would be testing a potential future clinical product. Also, there is evidence that porcine and human MSCs behave differently, with a higher reduction of viability of porcine MSCs, while in suspension, in comparison with human MSCs. Next to this, previous research has shown that the secretory profile of human MSCs is not influenced by the perfusion solution [36]. As it is unlikely that porcine MSCs will ever be administered to humans, we feel that the results of this study are more translatable to a setting in which human MSCs will be delivered to a human kidney, than investigating the effect of porcine MSCs in porcine kidneys would be.
Our study had several limitations. First, we did not focus on homing and survival of MSCs, and can therefore not be sure that the rather subtle short-term effect of MSC treatment that we observed is due to death of these cells as a result of being in continuous suspension during NMP. However, from previously performed similar research, we can conclude that when MSCs are administered to an isolated kidney during NMP, at least a small proportion remains viable, and that their effects do not solely rely on their viability [37,38]. Although MSCs are not immune privileged they are, to a certain extent, considered to be immune evasive [39,40]. Several studies have shown that MSCs are well tolerated and function across species [41 –43]. Nonetheless, as we used human MSCs in a porcine kidney, we cannot fully rule out the possibility of a xeno-effect as a result of human MSCs interacting with passenger leukocytes in the porcine kidney. Therefore, an important limitation of our study is that we did not measure the efflux of passenger leukocytes into the perfusion solution. This efflux is known to contain large populations of T cells, which have the ability to drive inflammation and secrete proinflammatory cytokines leading to higher levels of IL-6, IL-8, and IFN-γ [44]. However, the reported levels of these cytokines in the study by Stone et al. are a 1,000-fold higher than in this study. Possibly, this could be explained by the fact that there had already been a substantial efflux during HMP, which resulted in fewer passenger leukocytes being present during subsequent NMP. To date, the efflux during HMP remains undetermined, as does the fate of this immune efflux [44,45].
Summarizing, this study was able to demonstrate that the addition of human MSCs to an ischemically damaged porcine kidney during NMP will lead to a reduced expression of injury markers and an increased secretion of HGF, IL-6, and IL-8. Transplant studies are now needed to further determine whether these findings are associated with a beneficial effect on the injured kidney regarding short- and longer-term graft function, as well as affect graft survival.
Footnotes
Acknowledgment
We would like to thank Petra Ottens for her help with NAG, TBARS, and Luminex analyses.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was financially supported by research grants from the Dutch Kidney Foundation (grant no. 14OKG01) and the Netherlands Organization for Health Research and Development (ZonMw) (grant no. 90715606). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the article.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.