
Abstract
The renin-angiotensin system (RAS) has been suggested to play an important role in cardiac remodeling after acute myocardial infarction (AMI). We have confirmed that bone marrow mesenchymal stem cell-derived exosomes (BMSC-EX) had similar types of repair like effects upon tissues as BMSC, but the mechanisms remain unknown. BMSC were cultured to the third generation and were induced to release exosomes. Rats were injected with exosomes (100 μg/mL) or stem cells (1 × 106/mL) through the tail vein immediately after AMI was built, compared to those treated with physiological saline. Thereafter, all groups were analyzed for cardiac function, infarction sizes, and the levels of expression of BNP, ACE, ACE2, AngII, Ang1-7, and other factors in the plasma. After H2O2 makes contact with H9C2 cardiomyocytes, cell proliferation activity and apoptotic rates were measured by using CCK8 kits, to facilitate investigation of the effect of exosomes on H9C2 cells. In vivo, the index of cardiac remodeling and cardiac function was improved in both groups of exosomes and stem cells after AMI. Furthermore, exosomes may have helped to regulate the balance of the RAS system, upregulate ACE2—Ang1-7—Mas, and downregulate the ACE—AngII—ATIR pathway. Therefore, its effects were such as to accelerate the conversion of Ang II to Ang 1-7, thereby improving cardiac remodeling and forming sustained myocardial protection. In vitro, exosomal intervention was found to have increased the levels of activity of H9C2 cardiomyocytes under H2O2 injury and improved adverse effects of AngII upon H9C2 cells. All procedures for this study were reviewed and approved by the Institutional Animal Care and Use Committee (IACUC) at Guangdong Medical University. BMSC-EX improved cardiac remodeling and cardiac function, and had effects upon RAS system-related factors in plasma. Similarly, BMSC-EX also helped to protect H9C2 cells under attack from H2O2 or AngII, and may thus play beneficial roles by facilitating regulation of the balance of the RAS system.
Introduction
Globally,
Recent research has indicated that RAS is composed of two regulating axes acting on opposite sides related to the angiotensin-converting enzyme, angiotensin II, angiotensin receptor subtype AT1 (ACE -AngII- ATIR), and the negative regulation of ACE2-Ang1-7-Mas. ACE2-Ang1-7-Mas is known to be an important role player in the dynamics of research field of cardiovascular and cerebrovascular diseases, and may impart antihypertensive, antifibrotic, and antiproliferative characteristics [4].
As a homolog of ACE, ACE2 can degrade AngII and generate Ang- (subtypes 1-7), and is the main driver of both AngII degradation and Ang- (1-7) formation in myocardium [5,6]. Related research also indicated that the levels of expression of Ang- (1-7) in the periphery of the infarct were significantly enhanced, suggesting that Ang- (1-7) was associated with myocardial remodeling, and that ACE2 was the key enzyme that drove the decomposition of AngII into Ang- (1-7) [7].
Gurzu [8] found that Ang- (1-7) could inhibit AngII-induced vasoconstriction, which indicated that Ang- (1-7) could play a role through binding with AT1 receptors. ACE2- Ang1-7- Mas activates the G protein coupling receptor Mas, thus blocking the MAPK signal transduction pathway, exerting an antifibrosis and anti-inflammatory effect [9 –11]. That antagonizes the effects of Ang-II, increasing the blood flow of the coronary artery and reducing left ventricular dysfunction after ischemia [12], all while meanwhile inhibiting and reversing myocardial remodeling and ultimately protecting against effects of heart failure [13]. XNT is the activator of ACE2, which is believed to reduce blood pressure, improve cardiac function, reverse myocardial hypertrophy, and limit peripheral vascular fibrosis [14 –16].
Exosomes are membrane vesicles with a diameter of 30–120 nm and are released into extracellular matrices postfusion of multivesicular bodies and cellular membranes. Exosomes were discovered by Johnstone and other researchers who were studying the dynamics of transformation processes from reticulocytes to mature red blood cells. Protein, miRNA, and other substances are wrapped inside the exosomes [17]. Exosomes are released from cells, released into blood and other body fluids, and finally can be engulfed by other cells, and this process is an important medium for intercellular communication [18]. It is believed that large numbers of bioactive substances, membrane-anchored receptors, adhesion molecules, and cell surface-specific antigens are expressed upon surfaces of exosomes. Thus, exosomes play an important role in intercellular information transmission by way of combining with antigen antibodies of damaged target cells and play a role similar to the roles played by bone marrow mesenchymal stem cells (BMSCs) [19,20].
H9c2 cells are subcloned cell lines derived from embryonic heart tissues of BD1X rats, which are widely used in in vitro experimental studies of myocardial diseases. Zhao [21] found that exocrine derived from human umbilical cord mesenchymal stem cells can be used to induce inhibition of H9c2 cell apoptosis by way of regulating the levels of expression of proteins belonging to the Bax and Bcl2 families. These studies suggested that exocrine can be used to treat myocardial infarction and to protect myocardial cells. However, at present, the methods of assessment of cardiac function have remained mainly focused upon echocardiography and pathological section, and there are no uniform standards for evaluation methods.
Thus, the aim of this study was to explore therapeutic effects and mechanisms of exosomes derived from BMSCs used for treatment of AMI and whether or not findings were associated with RAS.
Materials and Methods
Specific pathogen-free (SPF) male SD (Sprague–Dawley) rats were provided by the Animal Experimental Center of Guangdong Province (Guangzhou, China) for use in our experiments. All animals were housed in conditions with standardized humidity, temperature, a natural light cycle of 12-h dark/12-h light, and with free access to water and food provided ad-libitum. Laboratory environments and equipment were used at and provided by the Clinical Medical Research Center in Affiliated Hospitals of Guangdong Medical University. All procedures for this study were reviewed and approved by the Institutional Animal Care and Use Committee (IACUC) at Guangdong Medical University.
Isolation, morphological observation, and detection of exosomes
The third-generation MSCs were cultured whereupon when confluences reached 80%–90%, cells were then added to the α-MEM medium without fetal bovine serum. After 48 h of culture under the conditions of 37°C and 5% CO2, the conditioned culture supernatants were collected. The supernatant was centrifuged (2,000 g) at 4°C for 30 min to remove whole cells and debris. Then, supernatant was ultracentrifuged at 100,000 g for 1 h, 100,000 g for 2 h, and 170,000 g for 2 h at 4°C. Exosomes were resuspended with 200 μL of PBS, and were then stored at −80°C. The concentrations of exosomes were measured by using bicinchoninic acid protein assay kits (KeyGen, Biotech, Inc.) following manufacturer's protocols. The morphology of exosomes was observed using transmission electron microscopy (JEM-1400) according to guidelines outlined in our related previous studies. Flow tests for exosomes were completed following all manufacturer protocols.
Animal experiment design and evaluation
We used a 7% chloral hydrate 5 mL/kg intraperitoneal injection for ECG-based monitoring. In the middle of the neck, the trachea was cut and intubated with an animal ventilator for positive pressure ventilation. The tidal volume of the ventilator was 8–10 mL/kg, exposing the heart by thoracotomy. The left anterior descending coronary artery (LAD) was ligated between the pulmonary artery cone and the left atrial appendage. I, aVL lead ST segment arched elevation of more than 0.1 mv and lasts more than 30 s as a sign of successful ligation. The laser Doppler blood flow measuring instrument was used to determine measures of blood flow perfusions of the heart. SPF male SD rats (8 weeks old) were divided randomly into seven treatment groups (N = 15 each group), which included a: control group (Nor group), 3 days postmyocardial infarction group (MI3d group), 28 days postmyocardial infarction group (MI28d group), 7 days post-treatment of bone marrow mesenchymal stem cells (BMSC7d group), 28 days post-treatment of bone marrow mesenchymal stem cells (BMSC28d group), 7 days post-treatment of exosomes (EXOSOMES7d group), and 28 days post-treatment of exosomes (EXOSOMES28d group). In addition to the control group, the other groups of rats were underwent surgery and confirmed the successful construction of myocardial infarction model.
Rats in the two BMSC groups were injected with MSCs (1 × 106/mL) [22] and the two EXOSOMES groups were injected with exosomes (100 μg/mL) [23] through the tail vein immediately after the operation. Rats in the control group and the two MI groups were injected with equivalent amount of physiological saline.
Measurement of the area of ischemia and the pressure of the left ventricle
Rat hearts were exposed after tracheal intubation and implementations of respiratory support. Measures of blood flow of the heart were detected by laser Doppler blood flow analyses and were made when the rat heart was in a normal state of beating. The blood flow analyzer has the options of assessing measures of function during both normal and ischemic states respectively. Image-Pro-Plus image software was used to facilitate the determination of the intercept of myocardial infarction and assessed images for the different treatment that contained measures of surrounding areas of the same sizes. Images indicated that the yellow colored regions were indicative of ischemic afflicted areas, whereas red colored areas were normal, and blue and gray areas were areas and region that lacked any blood flow (Supplementary Fig. S2). The images and corresponding values of left ventricular pressure in rats were measured by common carotid artery intubation. The right cervical artery was separated and exposed, the common carotid artery was punctured, and finally, the polyethylene catheter was sent from the common carotid artery to the left ventricle. The waveform-based images and pressure data were obtained for further analyses (Supplementary Fig. S1).
Histology staining
The heart tissues of each group were collected, tissues were cleaned, and heart tissues surrounding myocardial infarctions were dissected out, fixed in 4% Formaldehyde Solution for 24 h, and paraffin-embedded slices with a thickness of 5 μm were derived. Cardiac tissue staining followed all manufacturer's protocols and standard methods. After HE staining and Masson trichromatic staining were completed, tissue sections were observed under light microscopy. The areas of myocardial infarction area and volumes of fractions of collagen fibers were measured by using the image analysis system under low magnification.
Determination of ACE, ACE2, AngII, Ang1-7, and BNP factors in plasma
For plasma isolation, we collected whole blood from animals into a tube containing a anticoagulant and protease inhibitor cocktail, followed by centrifugation at 2,000 g for 15 min. Procedures were carried out following all manufacturer's protocols and instructions provided for the ELISA kit (Uscn Life Science).
Detection of proliferation activity and apoptosis of H9C2 cells by cytology
H9C2 cardiomyocytes of 4–6 generations were cultured and injured by applications of different concentrations of hydrogen peroxide. Measures of activity and apoptotic rates of H9C2 cells were detected after treatments with hydrogen peroxide. Cells were pretreated with different concentrations of exosomes. Cell proliferation activity was measured following manufacturer's protocols provided in the CCK8 kit. Optimum time points and concentrations of exosome preconditioning cells for hydrogen peroxide injuries were selected. Samples for experiments were divided into a normal control treatment group (Ctr group), a hydrogen peroxide-injured cell treatment group (hydrogen peroxide group), and an exosome co-culture pretreatment cell group (hydrogen peroxide + exosomes group). After the cells were treated at optimum concentrations and through optimum time points, CCK8 assays were used to facilitate determination of measures of proliferation activity of cells in each group. Flow cytometry was used to facilitate determinations of apoptotic rate.
Effects of AngII on the Morphology and Proliferation Activity of H9C2 Cells
H9C2 cardiomyocytes of 4–6 generations were co-cultured with AngII and H9C2 cells at different concentrations. Cell morphological changes were observed at different time points, and cell proliferation activity was measured by CCK8. The experiment was divided into normal control group (Ctr group), AngII and cell co-culture group (AngII group), and AngII + exosome and cell co-culture group (AngII + EXOSOMES group). Select the best concentration and time point of angiotensin II acting on cells. Morphological changes were observed in each group. CCK8 assay was used to determine the proliferation activity of cells in each group.
Statistical analyses
Data were composed and analyzed as the mean ± standard error of the mean (SEM). Statistical analyses were performed using one-way analysis of variance (ANOVA) followed by Tukey's tests. Differences with a value of P < 0.05 were considered statistically significant.
Results
Exosomes improved cardiac perfusion, reduced ischemic area, and improved cardiac function postmyocardial infarction
Our results indicated that BMSC and exosomes had therapeutic effects upon measures of AMI in rat based models. The laser Doppler blood flow-measuring instrument was used to detect measures from the cardiac surface flow images (Fig. 1A–C) in each treatment group, and the areas of ischemia-afflicted regions were calculated. That the ischemic area of the exosome treatment group and BMSC group was significantly lower compared with the control group upon the 28th day post-AMI (P < 0.05) was in our view, a very interesting and perhaps unexpected finding (Fig. 1D). Carotid artery intubation was used to measure left ventricular pressure (Supplementary Fig. S3) in each treatment group. We found that Pmax values in MI treatment group were significantly lower than in the Nor treatment group (P < 0.001) for the 28-day treatment groups. The Pmax values of both the exosomes and BMSC treatment groups were lower than were found in the Nor treatment group (P < 0.01), which were higher than the values observed in the MI treatment group (P < 0.05) (Fig. 2A). The values for Pmin in the MI treatment group were significantly higher than the values in the Nor treatment group (P < 0.01). Pmin values in both the exosome and BMSC treatment groups were lower than were the respective values observed in the MI treatment group (P < 0.05), and there were no significant differences between the exosomes and Nor treatment groups (P = 0.28) (Fig. 2B). The results suggested that both BMSC and exosomes could induce increases in measures related to ventricular systolic pressure postmyocardial infarction, and could induce reductions in end diastolic pressure. Ultimately, these effects could enhance myocardial contractility and improve diastolic dysfunction postmyocardial infarction. Furthermore, the levels of expression of BNP in plasma in the MI 3d group were higher than the respective levels in the Nor treatment group by way of measurements made with ELISA kits and assays (P < 0.05). Likewise, the levels of expression of plasma BNPs in the MI 28d treatment group were significantly higher than were observed in the Nor treatment group (P < 0.01). The levels of expression of plasma BNP in MI 28d treatment group were higher than were observed for the MI 7d treatment group (P < 0.05), and the levels of expression of plasma BNP in the Exosomes 28d treatment group were lower than were observed in MI 28d treatment group (MI <0.05) (Fig. 4C). These observations suggested that exosomes may be able to reduce the possibility of heart failure when applied post-AMI.
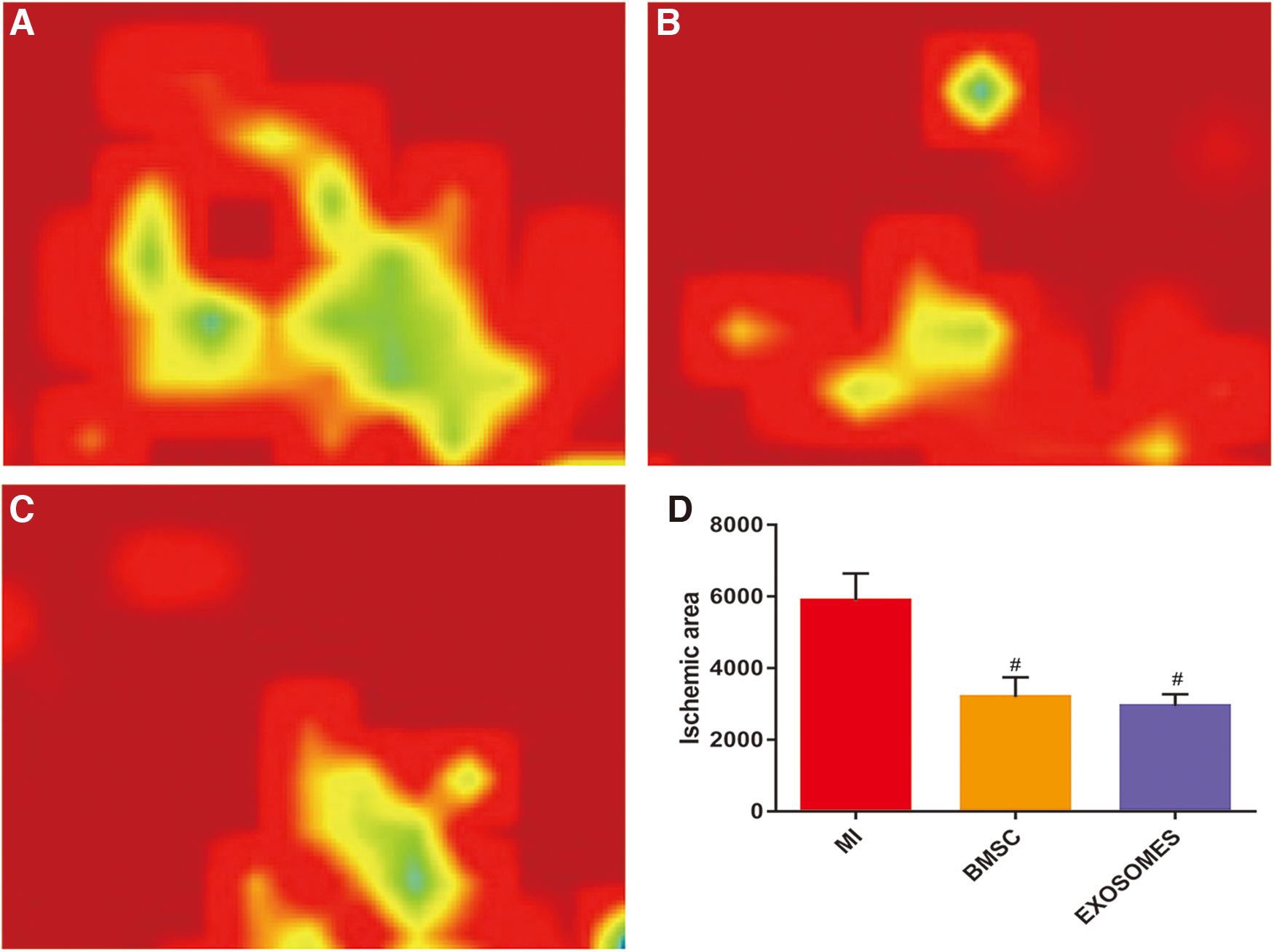
Exosomes improved cardiac perfusion and reduced ischemic area postmyocardial infarction. Regional blood flow image of myocardial infarction in the MI28d treatment group

Exosomes improved cardiac function postmyocardial infarction. Twenty-eighth day postmyocardial infarction, Left ventricular systolic pressure in rats for each treatment each group
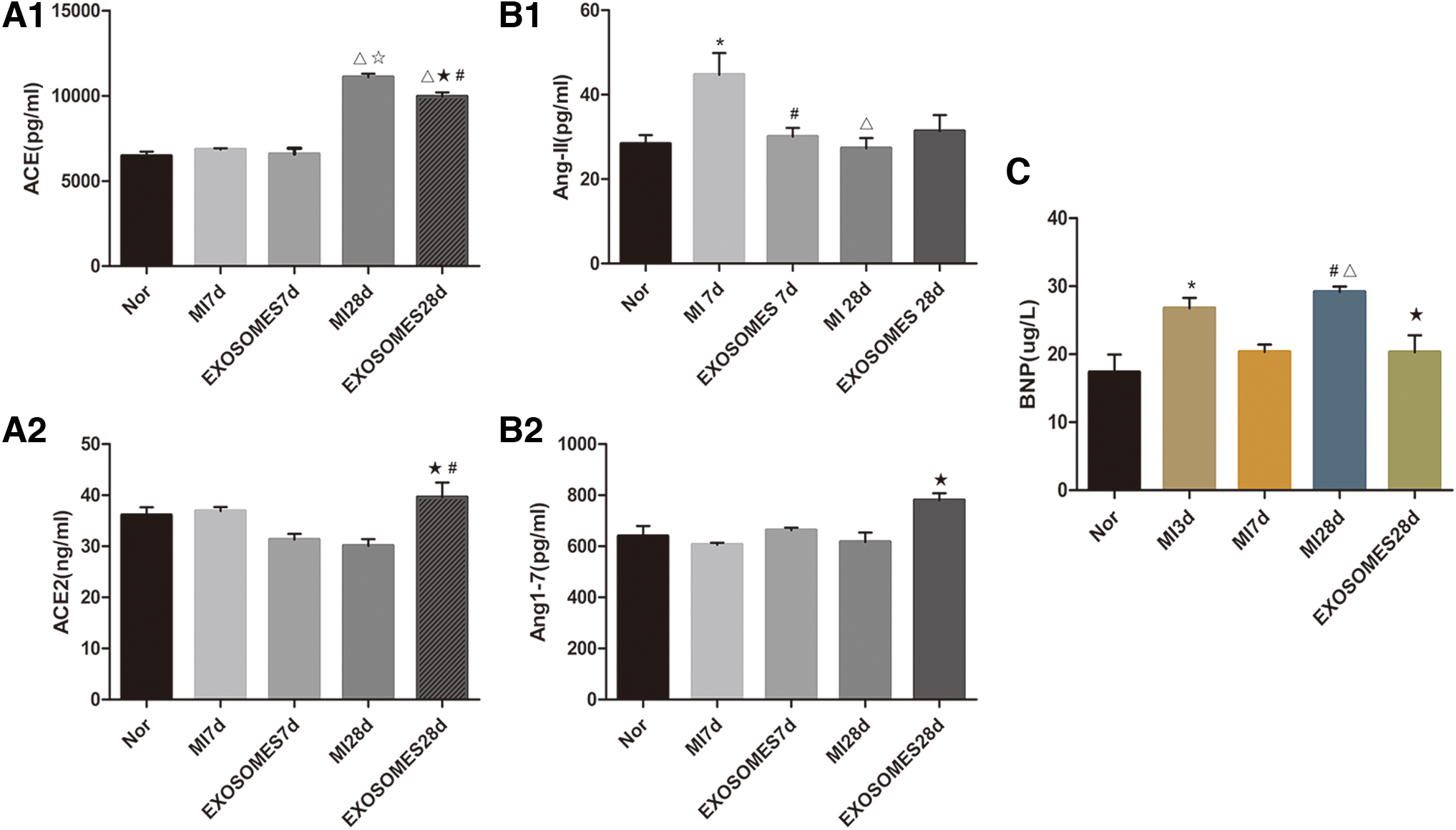
Exosome-based therapies may regulate the concentrations of RAS factor in plasma during treatment of acute myocardial infarction. The levels of expression of ACE and ACE2 in the plasma of rats in each treatment group
Exosomes reduced infarct size and decreased myocardial fiber volume fraction
Masson-trichrome staining was performed to facilitate the assessment of measures of the extent of fibrosis and collagen volume fraction, as well as the sizes of infarct areas (Infarct Size) in heart tissues (Fig. 3). The characteristic of Masson staining is that the part of myocardial fibrosis on pathological section is blue, and the area of the blue part can be calculated by image analysis software. Infarct size was defined as the sum of the epicardial and endocardial infarct circumference divided by the sum of the total LV epicardial and endocardial circumferences; the collagen content was determined by measuring the ratio of the blue area to the total area. Each parameter was quantitatively evaluated using the Image-Pro Plus software (Bethesda, MD) [24]. We discovered that heart tissues had extensive depositions of collagen in the MI 28d treatment group (P < 0.05), whereas in both the BMSC and exosome treatment groups, these depositions were effectively relieved (P < 0.05) (Fig. 3E) [19]. Myocardial infarcted areas in both the BMSC 28d treatment group and Exosomes 28d treatment group were smaller than we observed in the MI 28d treatment group (P < 0.05) (Fig. 3D).
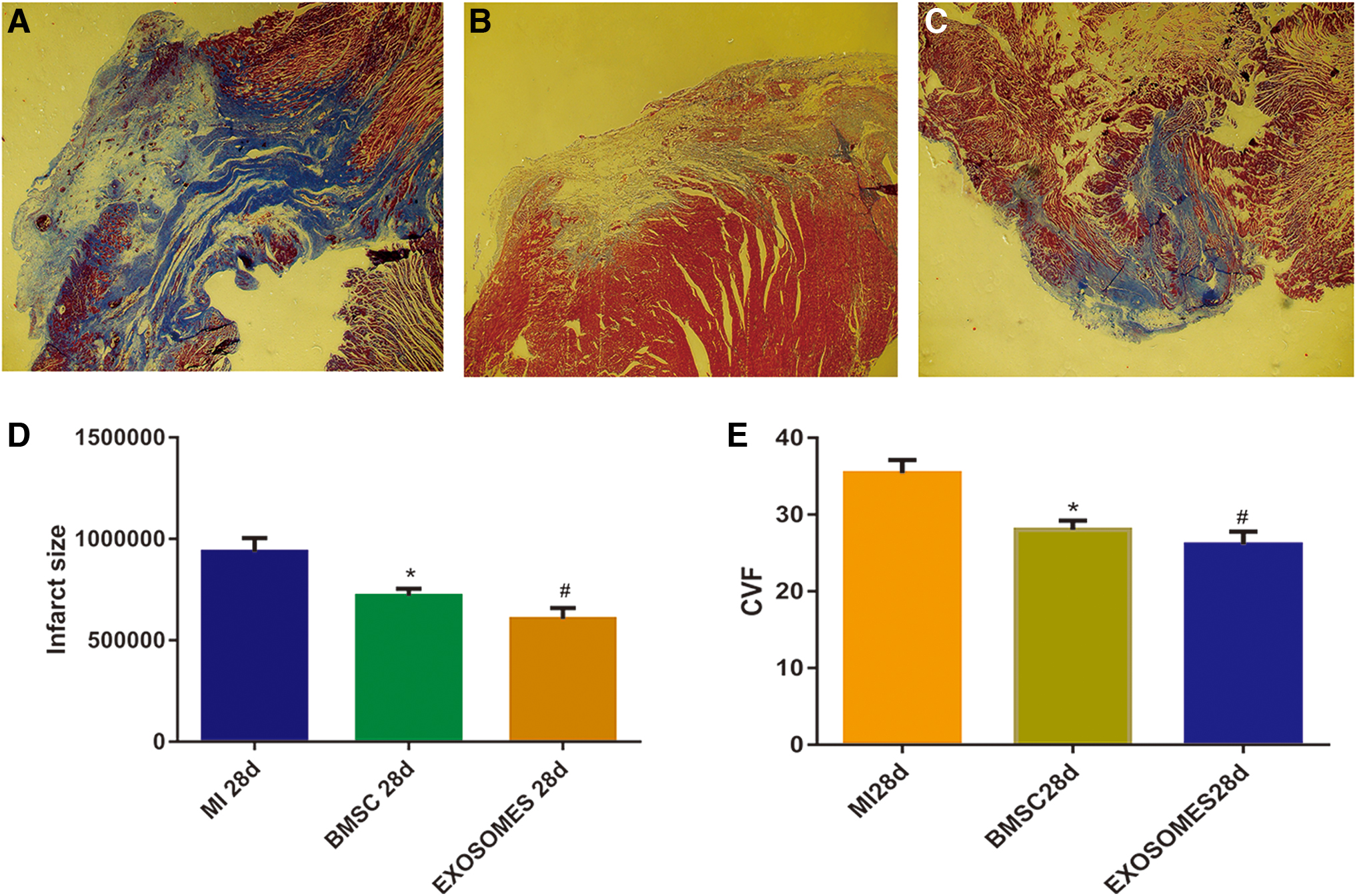
Exosomes reduced infarct size and decreased myocardial fiber volume fraction.MI28d treatment group myocardial infarction area (magnification, × 40)
Exosome-based therapies may regulate the concentrations of RAS factor in plasma during treatment of AMI
Measures of levels of expression of ACE and ACE2 in plasma of rats in each treatment group were detected by ELISA. There were no significant differences in plasma ACE expression between the MI 7d treatment group and Exosomes 7d treatment group. The levels of expression of plasma ACE in the MI 28d treatment group were significantly higher than we observed in the Nor treatment group (P < 0.001), and were significantly higher than we observed in the MI 7d treatment group (P < 0.001). The levels of expression of plasma ACE in the Exosomes 28d treatment group were significantly higher than in Nor treatment group (P < 0.001), and were also significantly higher than in the Exosomes 7d treatment group (P < 0.001), but were lower than in the MI 28d treatment group (P < 0.05). The levels of expression of plasma ACE2 in the Exosomes 28d treatment group were higher than we observed in the Exosomes 7d treatment group (P < 0.001), which was higher than we observed in the MI 28d treatment group (P < 0.05) (Fig. 4A). The levels of expression of AngII in plasma of MI 7d group were higher than we observed in the Nor treatment group (P < 0.05). The levels of expression of plasma AngII in the MI 28d treatment group were lower than we observed in the MI 7d treatment group (P < 0.05). The plasma AngII expression in Exosomes 7d group was lower than that in MI 7d group (P < 0.05), and there were no significant differences in plasma AngII expression among other groups (P = 0.48). The levels of expression of Ang- (1-7) in the plasma of the Exosomes 28d treatment group were higher than we observed in the MI 28d treatment group (P < 0.05). There were no significant differences in the plasma Ang- (1-7) expression between the other groups (P = 0.38) (Fig. 4B).
Exosomes alleviated damage from H2O2 to H9c2 cells
While 200 μg/L H2O2 was exposed to H9c2 cells for 2 h, the proliferative activity of cells was measured by CCK8. Results indicated that cellular activity of the H2O2 treatment group was 60% lower compared with the normal treatment group (P < 0.001) (Fig. 5A). Co-cultures of 0.5 μg/μL of exosomes and H9c2 cells for 5 h, whereupon after 2 h of additional exposure to 200 μg/L of H2O2 acting upon H9c2 cells facilitated determinations of measures of cell proliferation and activity by way of using CCK8. Results indicated that cellular activity of the exosome treatment group was higher compared with the H2O2 treatment (P < 0.01) (Fig. 5B). Flow cytometry assessments indicated that apoptosis rates of H9c2 cells in the control group were 4.2% ± 0.225% (Supplementary Fig. S4). Apoptosis rates of H2O2 treatment group were 10.72% ± 2.656%. Apoptosis rate of H9c2 cells in the exocrine group significantly decreased to a measure of 7.720% ± 0.965% (P < 0.05) (Fig. 5C).

Exosomes alleviated damage from H2O2 to H9c2 cells. Cell viability of H9c2 cells, which were treated with different concentrations of H2O2 for 2 h
Exosomes alleviated the adverse effects of Ang-II on H9c2 cells
Results from co-cultures with 0.3 μg/μL of AngII and H9c2 cells for 72 h indicated that the morphogenesis of H9c2 cells was significantly changed, and that the cellular arrangements were more disordered than were observed in the control group (Fig. 6A). Morphologies and arrangements of cells in the exosome treatment group were similar to those in the control group (Fig. 6B, C). Cellular activity observed in the AngII treatment group was lower than we observed in the control treatment group (P < 0.001), whereas cellular activity in the AngII+Exosomes treatment group was lower than we observed in the control treatment group (P < 0.01), which was higher than that we observed in the AngII group (P < 0.05) (Fig. 6D). These findings suggested that co-culturing of Ang-II and H9c2 cells can induce hypertrophy and apoptosis of H9c2 cells.

Exosomes alleviated the adverse effects of Ang-II on H9c2 cells. Control group cells (magnification, × 100)
Discussion
At present, endogenously applied plasma exosomes are believed to facilitate improvements in myocardial ischemia, protection of myocardial cells, and prevention of ischemia-/reperfusion-related injuries. Echocardiographies are often used to evaluate cardiac function after traditional myocardial infarction; however, in this approach it is difficult to avoid subjective factors in measurement processes. In the processes related to cardiac pathological sectioning, random errors are also unavoidable. Therefore, it has been difficult to achieve more objective and standardized assessments of functional and pathological changes postmyocardial infarction. Therefore, in this study, we used a more intuitive approach, which employed laser Doppler blood flow detection methods. The blood flow perfusion values and images of the heart were collected directly in each group. Ischemic regions were determined by specific color difference-based imaging. Areas of color, which differed, and imaging-based areas were calculated by way of using image analysis software, and areas of ischemic regions were evaluated more directly. In the measurement of ventricular pressure, we used the common carotid artery intubation manometry to obtain the left ventricular pressure image and corresponding values for pressure. The catheter passed through the common carotid artery to the left ventricle, and the pressure waveform and data were collected by the pressure sensor. It is considered that the highest value represents the systolic force of the rat heart, and the lowest value represents the diastolic force of the rat heart, thus reflecting the cardiac function [25]. Relevant animal studies have pointed out that PCSK9 is strongly expressed in the marginal area of myocardial infarction in rats, which is related to obvious cardiac systolic dysfunction [26]. Experimental evidence demonstrates that SGLT2 inhibitors exert cardioprotective effects in animal models of AMI through improved function during the ischemic episode, reduction of infarct size, and a subsequent attenuation of heart failure development [27]. Therefore, we can also reflect the cardiac function of rats by measuring the infarct size of rats.
In this study, we found that MSCs and MSC-exo could be used to facilitate reductions in levels and measures related to ischemic area and could induce decreases in measures of end-diastolic pressure, while also increasing systolic pressure of ventricular myocardium postmyocardial infarction in rats. Thus, our findings suggested that MSCs and MSC-exo could be used to improve cardiac function in cases of myocardial infarction. Smaller infarctal sizes and lower measures related to fibrosis also confirmed these conclusions to some extent. Stem cells and exosomes reduced the ischemic area postmyocardial infarction in our assessments using rats by way of analyses of cardiac surface perfusion values and images. Pathological sections and Masson trichrome-based staining findings indicated that stem cells and exosomes could significantly reduce size of infarction. Analyses of the volumes of fractions of cardiac tissue fibers implied that measures of the degrees of tissue fibrosis postmyocardial infarction could be improved. Left ventricular pressure measurements indicated that myocardial ischemia and hypoxia occurred in infarcted area postinduction of AMI, that myocardial cell necrosis occurred, and that consequently, the infarcted area was gradually replaced by fibrous tissues. We also found that measures of the degree of myocardial fibrosis were proportional to the severity of myocardial infarction. Therefore, inhibiting postinfarction myocardial fibrosis can be used a methodology to facilitate improvement of postinfarction cardiac function and myocardial remodeling, and can have a significant impact upon measures of long-term prognosis. Furthermore, we found that BNP increased within plasma upon the third day postmyocardial infarction in rats, and increased again on the 28th day postmyocardial infarction. The rises of BNP upon the third day may have been associated with heart failure postmyocardial infarction; however, this finding may also have been affected by way of increases of BNP in acute phases of myocardial infarction. The reasons underlying the second rounds of increases of BNP likely had to do with heart failure after myocardial infarction. This likely signified that exosomes could reduce the possibility of heart failure postmyocardial infarction. Accordingly, related research has demonstrated that RAS plays an important role in cardiac remodeling [5,6]. Our findings were supported based upon the levels of expression of RAS-related factors in plasma detected at different time intervals after exosomal treatments for myocardial infarction-afflicted rats. The results illustrated not only that upon the 28th day postmyocardial infarction, the levels of expression of plasma ACE in the control treatment group were significantly increased but also that the levels of expression in the exosome treatment group were lower than we observed in the control treatment group. The levels of expression of ACE2 in the control treatment group were not significantly different from those we observed in the normal treatment group, whereas the levels of expression of ACE2 in the exosome treatment group had significantly increased. These results are consistent with the goals of our research wherein we sought to target means to improve ventricular remodeling after activation of the ACE2 pathway. As the two promoters of RAS, the complex relationships and dynamics between the concentrations and activities of the promoter are still being explored; however, our findings suggested that differences in levels of expression and concentrations may imply there is a correlation between them. Thus, exosomes may be associated with RAS in considerations of treatments of myocardial infarction. Furthermore, we found that in the control treatment group, the levels of expression of Ang II increased upon the seventh day postmyocardial infarction, and that the levels of expression of Ang II in the exosomes treatment group were lower than in the control treatment group. However, there were no significant differences in the levels of expression of Ang 1-7 upon the seventh day; however, upon the 28th day, the levels of expression of Ang 1-7 in the exosome treatment group were higher than that in each of the other treatment groups. The levels of expression of Ang II in the control and exosome treatment groups were higher than in the other treatment groups upon the seventh day postmyocardial infarction. Thus, our findings suggested that exosomes may accelerate the transformation of Ang II to Ang 1-7 by way of activating the ACE2 pathway, such that the levels of expression of Ang 1-7 increase, as was found upon the 28th day in our research. The peptide Ang-(1-7), signaling by Mas, has been found to exert antifibrotic and antihypertrophic effects and can facilitate improvement in LV functions post-MI. Inhibition of the ACE2/Ang (1-7) axis and hyperactivity of the ACE/AngII axis may consequently lead to activation of EPR1/2 and protein kinase C (PKC) pathway [28]. Therefore, these may underlie reasons as to why we observed that fibrosis was relieved and that measures of heart function were enhanced. After exosomal based therapies for the respective treatment group, we found that measures of activation abilities of RAS promoters were affected, and that the more meaningful approach was to accelerate the conversion of adverse factors to favorable factors, thereby forming persistent myocardial protection.
Large numbers of reactive oxygen species (ROS) can be produced during myocardial ischemia and reperfusion. These ROS can consequently cause apoptosis in cardiomyocytes due to oxidative stress and can lead to cardiac remodeling and cardiac dysfunction [29]. H2O2 as a kind of ROS that can directly oxidize lipids and proteins located upon cellular membranes, can react with iron ions in the cell to produce more active free radicals, and can induce apoptosis through its impacts and roles in a variety of pathways. Research has also indicated that H2O2 can induce apoptosis in cardiomyocytes through increasing levels of oxidative stress [30]. After the action of H2O2 upon H9c2 cells, we found that measures of the activity of cells decreased; however, measures of the activity of the exosomes treatment group were higher than for the control group, and measures of apoptosis rates were reduced. These findings signified that exosomes improved the activity of cardiac muscle cells and enhanced the ability of myocardial cells to resist oxidative stress-related injury. Our findings are supported by previous research that confirmed myocardial damage occurred after myocardial infarction and was related to activation of RAS. Findings based around AngII in this study indicted it could promote H9c2 hypertrophy and apoptosis, which is consistent with the claims made by research from Gavras [31]. Measures related to cellular activity of the exosome treatment group were higher than were observed in the control treatment group, and cell morphology was similar to what was observed in the normal treatment group. Thus, exosomes can facilitate the alleviation of adverse effects of AngII upon cardiomyocytes. Previous research has confirmed that AngII initiates expression of nuclear genes by way of activating mitogen-activated protein kinases (MAKPs), which consequently cause a series of changes, such as cell proliferation, cytokine secretion (TGF-β), extracellular matrix expression, and protein synthesis. Thus, initiating the transcription and expression of nuclear fibrosis and proliferation-related genes can result in the promotion of proliferation of cardiac fibroblasts and increases in collagen synthesis [32,33]. Thus, whether or not exosomes can regulate AngII by way of influencing gene expression requires further investigation. More important was that targeting deletion of AngII receptors may consequently increase the risk of heart rupture in AMI rats. Whether or not the inhibition of AngII action that we observed in this study would have consequently increased the possibility of cardiac rupture in AMI remains unclear and is an important area for future research.
Conclusion
In this study, we confirmed the positive effects of exosomes in treatment of myocardial infarction, and that they could reduce the ischemic area and the risk of heart failure, alleviate fibrosis, and improve myocardial perfusion. Furthermore, exosomes were found to have affected the levels of expression of RAS-related factors in plasma in examinations of samples with myocardial infarction. In vitro, exosomes protected H9c2 cells from damage caused by either H2O2 or AngII. Protective effects of exosomes in myocardial infarction may have worked by way of regulating the balance of the RAS system.
Footnotes
Author Disclosure Statement
The authors declare no potential conflicts of interest.
Funding Information
This article was supported by National Natural Science Foundation of China (No.81370242), Medical Scientific Research Foundation of Guangdong Province of China (No.A2019089), and The Stem Cell Preclinical Research Projects of the Affiliated Hospital of Guangdong Medical University (No.2018PSSC006).
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.