
Abstract
Bone marrow mesenchymal stem cells (BMSCs) are thought to have great potential in the treatment of many diseases and may serve as a cell source for tissue engineering. These cells may be regulated by stromal cell-derived factor-1α (SDF-1α), which has been shown to promote the migration, proliferation, and osteogenic differentiation of BMSCs in inflammation-associated diseases. However, the specific mechanism underlying this process remains unclear. We herein transduced lentivirus carrying SDF-1α, empty vector, or siRNA-SDF-1α into mouse BMSCs and then performed transwell, CCK-8, cell cycle, alkaline phosphatase activity, and Alizarin Red staining experiments on the three groups of samples. Overexpression of SDF-1α promoted the migration, proliferation, and osteogenic differentiation of BMSCs, and SDF-1α upregulated the expression of Wnt pathway-related factors and downstream target genes as determined by western blot, real-time polymerase chain reaction, and immunofluorescence. The effect of low SDF-1α expression on BMSCs was significantly weakened. In addition, we transduced lentivirus carrying siRNA-Wnt3a into BMSCs and treated them with SDF-1 drugs. After inhibiting the Wnt pathway, SDF-1 significantly weakened the migration, proliferation, and osteogenic differentiation of BMSCs. From this, we concluded that high SDF-1 expression can promote the migration, proliferation, and osteogenic differentiation of BMSCs, at least in part by activating the Wnt pathway.
Introduction
Bone marrow mesenchymal stem cells (BMSCs) are nonhematopoietic stem cells that originate in the embryonic mesoderm and have the potential for self-replication, multidirectional differentiation, and regeneration [1,2,4]. In the 1970s, BMSCs were first discovered in the bone marrow by Friedenstein [5]. Studies have confirmed that under specific induction conditions, BMSCs have the ability to differentiate into various types of mature cells, such as osteoblasts and cells comprising the cartilage, spinal cord, nerves, fat, tendons, skin, and liver [3,4,6]. In addition, BMSCs can also secrete a variety of growth factors and cytokines and have low immunity and immunomodulatory functions [7]. With the development of cytology and tissue engineering techniques, BMSCs have been shown to have multidirectional differentiation and regeneration potential and are thus used as seed cells to promote the repair of damaged tissues, such as bone, cartilage, joint, heart, and liver tissue [8]. Therefore, BMSCs research has become a hot spot in the medical community.
Chemokines are a class of small cytokines or signaling proteins whose main function is to move chemotactic cells in a specific direction, and more than 60 members have been classified to date [9]. Stromal cell-derived factor-1 (SDF-1), an important member of the chemokine family, is expressed in two subtypes, SDF-1α and SDF-1β, with SDF-1α being the main subtype. SDF-1α is widely present in many tissues and organs of the human body, such as the lymph nodes, bone marrow, liver, lung, muscle, small intestine, kidney, and brain, and can sustainably exist in these organs and tissues [10 –14]. Studies have shown that SDF-1α plays an important role in the physiological functions of migration, distribution, development, differentiation, and apoptosis of various cells [15,16]. Moreover, SDF-1α plays a key role in the pathological process of some diseases, such as inflammation, tumor formation and metastasis, pathogen infection, and wound repair [17,18]. An increasing number of animal experiments have shown that SDF-1α can enhance the migration of BMSCs, mobilize BMSCs to diseased areas, and promote their proliferation and differentiation [19,20]. However, the specific mechanism of this effect of SDF-1α on BMSCs is still unclear.
During the migration, proliferation, and osteogenic differentiation of BMSCs, SDF-1α is regulated by multiple factors and multiple signaling pathways. Among them, the classical Wnt/β-catenin signaling pathway is crucial, affecting the migration, proliferation, and osteogenic differentiation of BMSCs through paracrine or autocrine methods [21]. The classical Wnt/β-catenin signaling pathway is a complex protein network that includes many proteins that can regulate the synthesis of Wnt signaling molecules, including Wnt glycoproteins (mainly Wnt1, Wnt3a, Wnt8, Wnt10b), β-catenin, frizzled receptors, low-density lipoprotein receptor-related proteins (LRP5/6), disheveled (Dsh), and glycogen synthesis kinase (GSK-3β). Activation of the classical Wnt/β-catenin signaling pathway is initiated by the Wnt protein combined with the frizzled receptor on the cell surface and LRP5/6 to form a complex. This complex activates Dsh in the cytoplasm, and the Dsh release signal inhibits the degradative activity of GSK-3β on β-catenin such that more free β-catenin accumulates in the cytoplasm and enters the nucleus to activate the LEF/TCF transcription factor and initiate downstream target genes [22 –26].
In this study, we transduced BMSCs with recombinant lentivirus carrying SDF-1α, empty vector, or siRNA-SDF-1α and observed that Lv-SDF-1α could enhance the migration, proliferation, and differentiation of BMSCs. We also observed a positive correlation between the expression of Wnt signaling pathway-related factors and SDF-1α. In addition, we used SDF-1α (100 ng/mL) to stimulate BMSCs transduced with siRNA-Wnt3a and observed that the migration, proliferation, and differentiation abilities of BMSCs were significantly reduced compared with those of the control group. Our results showed that SDF-1α promotes the migration, proliferation, and differentiation of BMSCs through the Wnt/β-catenin signaling pathway.
Materials and Methods
Cell culture and osteogenic differentiation
C57BL/6 mouse BMSCs were purchased from Cyagen Company (Guangzhou, China). BMSCs were placed in complete medium, which included basal medium, special 10% fetal bovine serum, 1% penicillin–streptomycin double antibody, and glutamine. All cells were cultivated at 37°C and 5% CO2 in a cell incubator. Upon reaching 80%–90% confluence, the BMSCs were digested and passaged, and the cell suspension was centrifuged at 1,000 rcf for 5 min. The supernatant was removed and resuspended in a medium, and the cells were used for further experiments; the BMSC passages did not exceed 10 generations.
To induce osteoblastic mineralization, upon reaching 60%–70%, BMSCs were cultured in osteogenic-inducing medium (OM) containing 10−8 M dexamethasone, 50 μg/mL ascorbic acid, and 5 mM β-glycerophosphate. The OM was replaced every 3 days for 3 weeks.
Lentivirus transduction of BMSCs and cell grouping
Third-generation BMSCs were selected and inoculated into 24-well plates at a density of 1 × 104 cells per well overnight in a 37°C incubator. After 24 h, LV-sdf-1-mus (GenePharma), LV-siRNA-sdf-1 (GenePharma), LV-siRNA-Wnt3a-mus (GenePharma) recombinant, and/or empty vector virus (GenePharma) were added to the medium according to the multiplicity of infection (MOI = 400). After 96 h of infection, green fluorescent protein was observed under a fluorescence inverted microscope (Olympus, Tokyo, Japan).
This experimental study was divided into two parts. The first part aimed to verify the effect of SDF-1α on the migration, proliferation, and differentiation of BMSCs. The cells were divided into three groups: the Lv-SDF-1α group (high SDF-1α expression), Null group (empty vector), and siRNA-SDF-1α group (low SDF-1α expression). The second part aimed to verify the effect of the Wnt pathway on the migration, proliferation, and differentiation of BMSCs by SDF-1. Cells were divided into three groups: the SDF-1+Null group (SDF-1 drug+empty vector), SDF-1+siRNA-Wnt3a group (SDF-1 drug + low Wnt3a expression), and siRNA-Wnt3a group (low Wnt3a expression). SDF-1 drugs were purchased from PeproTech.
Cell viability and cell cycle assays
The CCK-8 assay was used to detect cell viability and proliferation. BMSCs in each group were suspended, inoculated into 96-well plates (1.0 × 103 cells/well), and cultured in an incubator at 37°C and 5% CO2. Then, 10 μL of CCK-8 reagent (DOJINDO) was added to each well on days 1, 2, 3, 4, and 5. After incubation for 2 h, the absorbance at 450 nm was measured with a microplate reader.
The BMSCs in each group were selected and inoculated into 10-cm2Petri dishes (1 × 105 cells) and cultured for 5 days. The BMSCs were then harvested, washed with phosphate-buffered saline (PBS) (centrifuged at 2,000 rpm, 5 min), and made into a 1 × 106/mL single-cell suspension. The single-cell suspension was centrifuged, and the supernatant was removed and fixed with 75% cold ethanol at 4°C overnight. Next, the cells were centrifuged and washed with PBS and then incubated with 500 μL of PI/RNase (1:9) Staining Buffer (KeyGEN BioTECH, Nanjing, China) for 60 min. The cell cycle proportions (G1, S, and G2 phases) were detected by flow cytometry.
Transwell experiment
Cell migration was confirmed by the Transwell chamber (Corning) experiment. Transwells were seeded with 1 × 104 cells in the upper chamber, and 600 μL of serum-free medium containing SDF-1 drug (100 ng/mL) was added to the lower chamber at 37°C for 24 h. The next day, complete medium was removed, and the cells were washed with PBS and fixed with 4% paraformaldehyde for 10 min. Then, the polyethylene membrane was soaked in PBS, removed with a scalpel, and placed on a glass slide before being stained with Wright's-Giemsa stain (Jiancheng, Nanjing, China) for 8 min. Finally, the cells were observed and recorded under a microscope.
Alkaline phosphatase staining
BMSCs were inoculated on six-well plates and cultured for 7 days after osteogenic differentiation. The culture medium in the six-well plate was collected and subjected to alkaline phosphatase (ALP) activity measurement, rinsed with PBS, fixed in 4% paraformaldehyde for 15 min, and washed with PBS again. Then, ALP incubation solution (Solarbio, Beijing, China) was added to each well according to the manufacturer's instructions, and the cells were incubated in the dark for 20 min, rinsed with PBS, counterstained with Fast Red staining solution for 5 min, rinsed with PBS, and observed under a microscope.
Alizarin red staining
Mineral deposition was measured by Alizarin Red staining (ARS; Cyagen, Guangzhou, China) after the induction of osteogenic differentiation for 3 weeks. BMSCs were fixed with 4% paraformaldehyde for 30 min, washed with PBS twice, incubated with a 0.2% Alizarin Red solution for 5 min, and rinsed with PBS. The effect of osteogenic staining was observed under a microscope.
RNA extraction and quantitative real-time polymerase chain reaction
BMSCs were washed twice with PBS, and total RNA was extracted using a Total RNA Kit I (Omega) according to the manufacturer's instructions. The primers were provided by Sangon Biotech (Shanghai, China), the sequences of which are given in Table 1. Real-time polymerase chain reaction (PCR) was performed using green PCR SuperMix (TransGen, Beijing, China) on an Analytik Jena AG instrument (Qtower3G, Germany).
Primers Used for q-PCR
OCN, osteocalcin; q-PCR, quantitative real-time polymerase chain reaction; RUNX-2, runt-related transcription factor 2; SDF-1α, stromal cell-derived factor-1α.
Western blot analysis
To determine the protein expression of certain markers, proteins were extracted with RIPA lysis buffer (KeyGEN BioTECH) according to the manufacturer's instructions, and the protein concentrations in each sample were determined using the BCA Protein Assay Kit (KeyGEN BioTECH). Equal amounts of protein (50 μg) were loaded onto 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis gels and then transferred to polyvinylidine fluoride membranes (Millipore, Shanghai, China). The membranes were then blocked in 5% nonfat milk at room temperature for 1.5 h and incubated at 4°C overnight. The primary antibodies included anti-SDF-1 (ab9797, 1:2,000; Abcam), RUNX-2 (ab23981, 1:1,000; Abcam), OCN (DF12303, 1:500; Affinity, China), CyclinD1 (ab134175, 1:10,000, Abcam), c-Myc (ab32072, 1:300; Abcam), Wnt3a (ab219412, 1:1,000; Abcam), β-catenin (ab32572, 1:1,000; Abcam), and β-actin (bs-0061R, 1:5,000; Bioss, Beijing, China). After that, the membranes were washed and incubated with secondary antibodies (goat anti-rabbit IgG, 1:5,000) for 2 h at room temperature. Proteins were detected with the Enhanced Chemiluminescence Detection Kit (Millipore, Shanghai, China) and visualized by a Bio-Rad imaging system with Quantity One analysis software. Relative protein expression was normalized to the β-actin expression level.
Immunofluorescence
After BMSCs were cultured in a 24-well plate for 7 days, SDF-1α, Wnt3a, β-catenin, c-Myc, Cyclin D1, runt-related transcription factor 2 (RUNX-2), and osteocalcin (OCN) were detected by immunofluorescence (IF). Briefly, BMSCs were fixed in 4% paraformaldehyde for 30 min, permeabilized with 0.2% Triton X-100 for 15 min, and blocked with 1% bovine serum albumin for 30 min. Fixed BMSCs were washed and incubated overnight at 4°C with primary antibodies against SDF-1 (ab9797, 1:200; Abcam), RUNX-2 (ab23981, 1:400; Abcam), OCN (DF12303, 1:50; Affinity), Cyclin D1 (ab134175, 1:50; Abcam), c-Myc (ab32072, 1:100; Abcam), Wnt3a (ab219412, 1:100; Abcam), and β-catenin (ab32572, 1:250; Abcam). Subsequently, the cells were incubated with a fluorescence-conjugated secondary antibody (1:800; Jackson, PA) for 2 h at room temperature. Nuclei were stained with DAPI (ZSGB-BIO, Beijing, China) for 3 min. The presence of the proteins was examined under a fluorescence microscope (Olympus).
Statistical analysis
Statistical analysis was conducted using GraphPad Prism software (GraphPad Software, Inc.). The quantitative data are expressed as the mean ± standard deviation. When the data distribution was determined to be normal, the variance was uniform, and single-factor analysis of variance was used; a nonparametric test was used if these conditions were not satisfied. Semiquantitative data were obtained using the Friedman-M test of multiple samples designed by random blocks. P < 0.05 was considered statistically significant.
Results
Successful transduction of BMSCs and activity of SDF-1 in BMSCs
BMSCs were successfully transduced after lentivirus culture for 96 h, as shown by green fluorescence. When the MOI equaled 400, the green fluorescence intensity reached 80%–90% (Fig. 1A). BMSCs were transduced with lentiviruses carrying SDF-1α, empty vector, or siRNA-SDF-1α. Western blot (WB) and q-PCR experiments confirmed that the SDF-1α protein level in the Lv-SDF-1α group was higher compared with the Normal and Null groups, whereas that in the siRNA-SDF-1α group was lower compared with the Normal and Null groups; there was no difference between the Normal and Null groups (Fig. 1B, C). In subsequent experiments, we used the Null group as the negative control group. As shown in the fluorescence image, the expression level of SDF-1α was higher in the LV-SDF-1α group than in the Null group and lower in the siRNA-SDF-1α group than in the Null group (Fig. 1D).
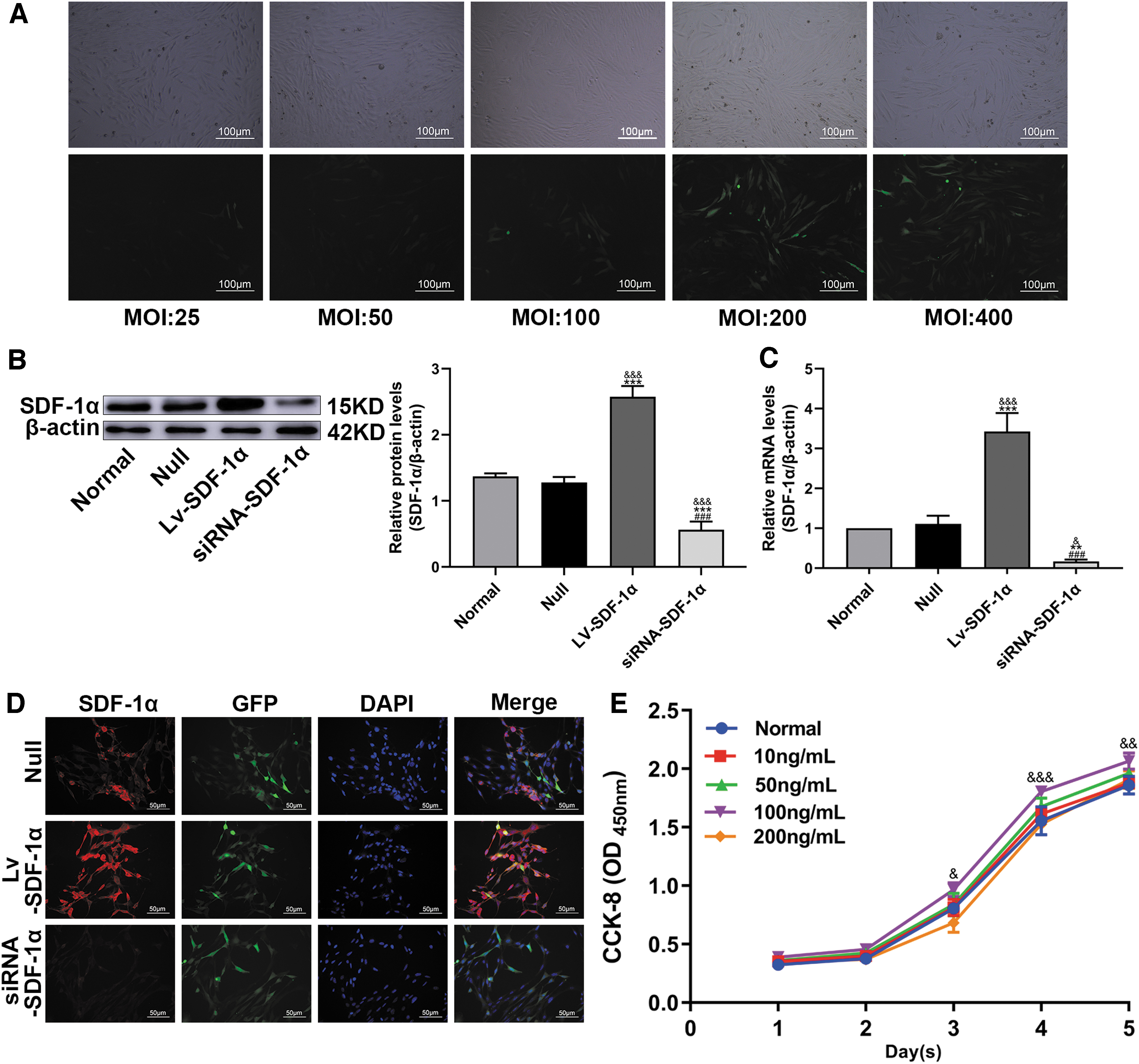
The effect of SDF-1 treatment on BMSC viability was assessed by the CCK-8 assay. BMSCs were treated with SDF-1 at concentrations of 10, 20, 50, 100, and 200 ng/mL for 1, 2, 3, 4, and 5 days of continuous culture; 100 ng/mL was determined to be the optimal concentration, as it had no effect on cell activity (Fig. 1E).
Lv-SDF-1α promotes the migration and proliferation of BSMCs and activation of the Wnt/β-catenin pathway
In the Transwell experiment, the number of migrating cells in the Lv-SDF-1α group was significantly higher compared with the Null group, whereas that in the siRNA-SDF-1α group was significantly lower compared with the Null group (Fig. 2A). This result shows that the high expression of SDF-1α induced by Lv-SDF-1α can promote the migration of BMSCs.
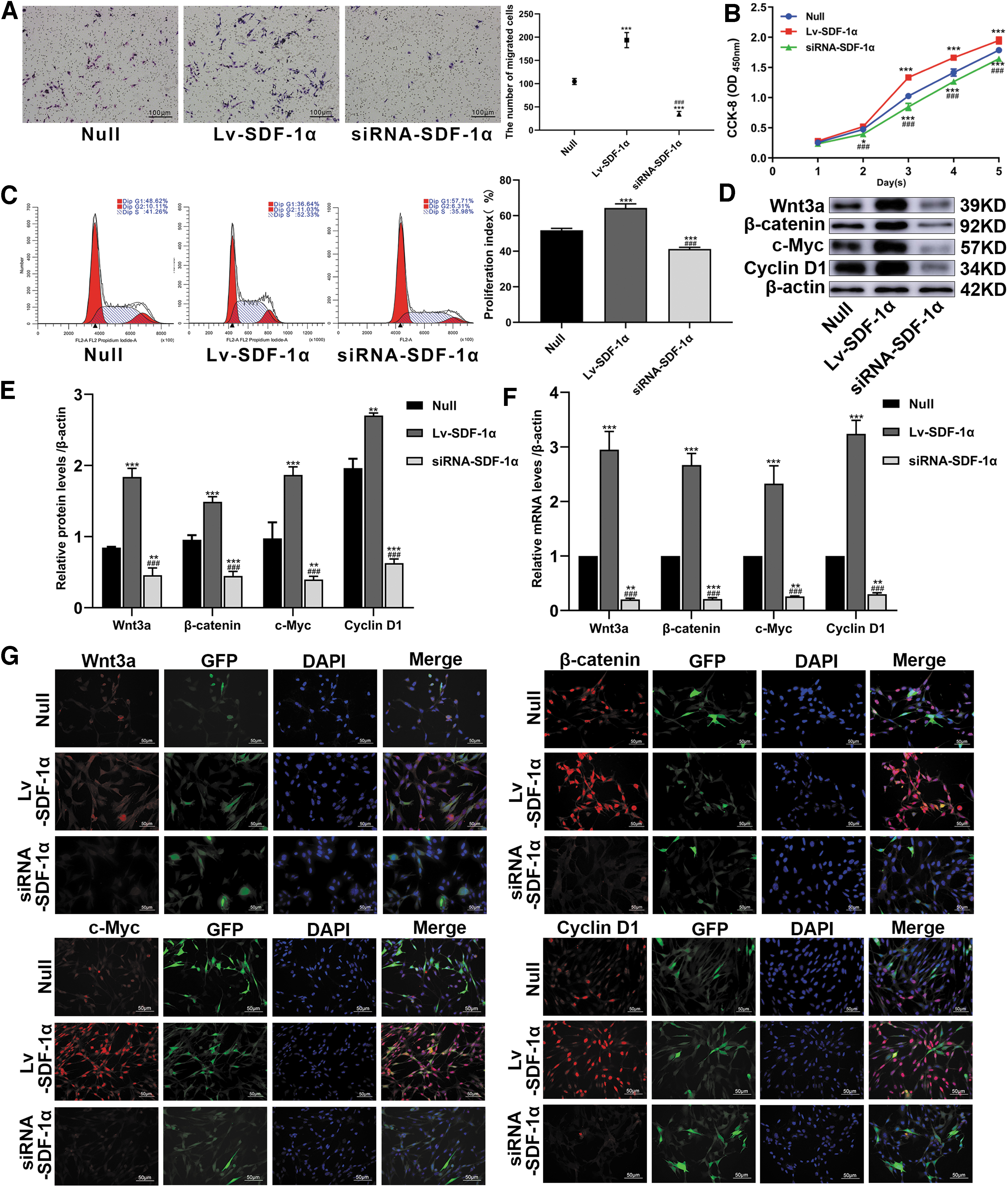
Lv-SDF-1α promotes the migration and proliferation of BMSCs. BMSCs were transduced with Lv-SDF-1α, empty vector, or siRNA-SDF-1α genes.
CCK-8 confirmed that Lv-SDF-1α promoted the proliferation of BMSCs compared with that in the Null group, whereas siRNA-SDF-1α delayed the proliferation of BMSCs compared with that in the Null group (Fig. 2B). According to the analysis of cell cycle detection by flow cytometry, the G1 phase was significantly shorter in the LV-SDF-1 group than in the Null group, and the Null group was shorter than the siRNA-SDF-1 group (36.64% vs. 48.62% vs. 57.71%). The G2+S phase was significantly longer in the LV-SDF-1 group than in the Null group, whereas the Null group was longer than the siRNA-SDF-1 group (11.03% +52.33% vs. 10.11% +41.26% vs. 8.31% +35.98%). The cell cycle test results were consistent with the CCK-8 results (Fig. 2C). WB and IF experiments confirmed that during the proliferation of BMSCs, Wnt3a, β-catenin, c-Myc, and Cyclin D1 protein levels were enhanced in the Lv-SDF-1α group but decreased in the siRNA-SDF-1α group, and the differences were significant (Fig. 2D, E, G). Compared with those in the Null group, the RNA levels of Wnt3a, β-catenin, c-Myc, and Cyclind1 were higher in the Lv-SDF-1α group and lower in the siRNA-SDF-1α group, as determined by q-PCR (Fig. 2F).
These results suggested that SDF-1 promoted the migration and proliferation of BMSCs, and we also found that Wnt pathway-related factors and downstream target genes were activated during this process.
Lv-SDF-1α can enhance BMSC osteogenic differentiation and activate the Wnt/β-catenin pathway
ALP and ARS showed that Lv-SDF-1α enhanced the osteogenic differentiation of BMSCs, whereas siRNA-SDF-1α inhibited the osteogenic differentiation of BMSCs (Fig. 3A, B). WB and IF experiments confirmed that during the osteogenic differentiation of BMSCs, the Wnt3a, β-catenin, c-Myc, RUNX-2, and OCN proteins were enhanced in the Lv-SDF-1α group and weakened in the siRNA-SDF-1α group, with significant differences (Fig. 3C, E). q-PCR confirmed that the RNA levels of Wnt3a, β-catenin, c-Myc, RUNX-2, and OCN were high in the Lv-SDF-1α group and low in the siRNA-SDF-1α group, with significant differences (Fig. 3D). These results suggest that SDF-1α can enhance the osteogenic differentiation of BMSCs and activate the Wnt signaling pathway.

Lv-SDF-1α can promote the osteogenic differentiation of BMSCs. BMSCs were transduced with Lv-SDF-1α, empty vector, or siRNA-SDF-1α genes.
Successfully transduced siRNA-Wnt3a gene into BMSCs
BMSCs were transduced with a lentivirus carrying siRNA-Wnt3a, resulting in low Wnt3a expression. WB and q-PCR confirmed that the expression level of Wnt3a in the siRNA-Wnt3a group was significantly reduced compared with that in the Normal and Null groups, whereas the differences between the Normal and Null groups were not statistically significant (Fig. 4A, B), and IF analysis also confirmed this result (Fig. 4C). Moreover, the protein SDF-1α was detected in these three groups by WB, and there was no significant difference between them (Fig. 4D).
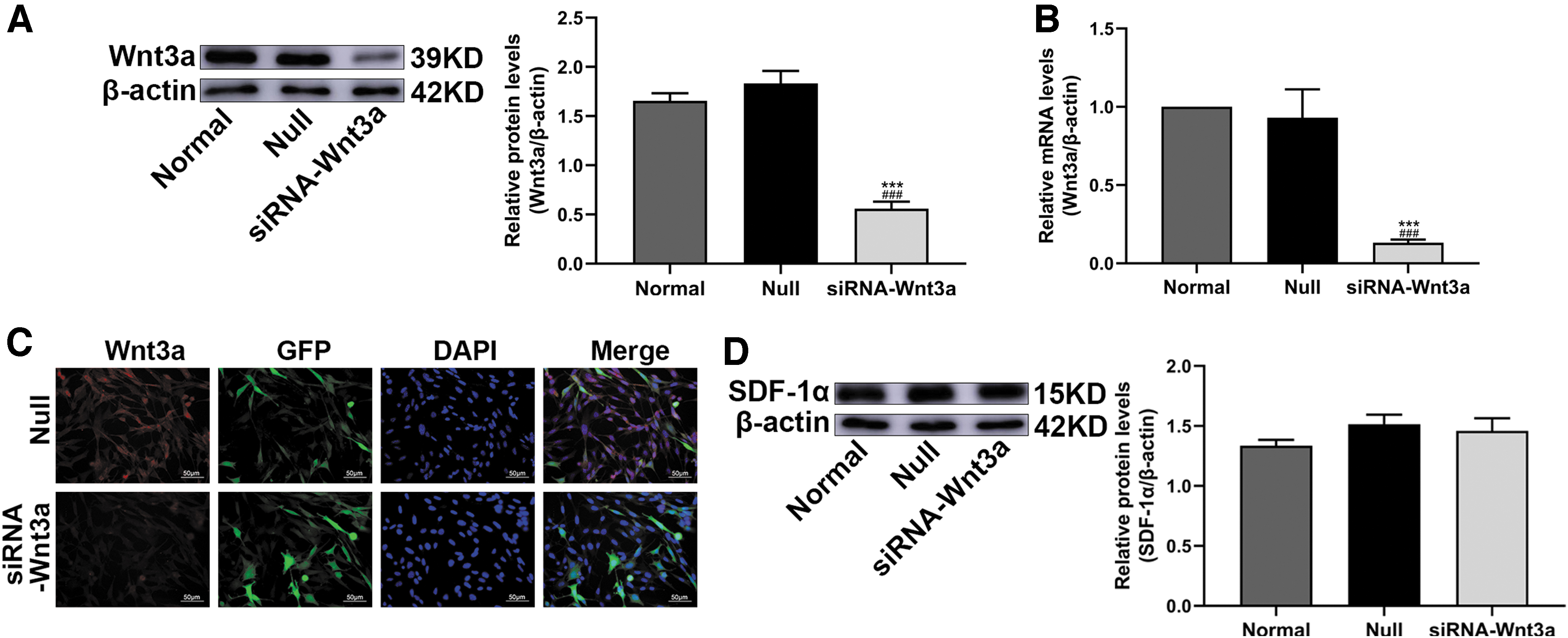
BMSCs were transduced with empty vector or siRNA-Wnt3a gene.
Inhibition of the Wnt pathway and the migration and proliferation of BMSCs were reduced by SDF-1 (100 ng/mL)
In the Transwell experiment, the number of migrated cells in the SDF-1+siRNA-Wnt3a and siRNA-Wnt3a groups was significantly lower than that in the SDF-1+Null group; however, there was no significant difference between the SDF-1+siRNA-Wnt3a and siRNA-Wnt3a groups (Fig. 5A), suggesting that the migration of SDF-1 to BMSCs is partially achieved through the Wnt pathway.

By inhibiting the Wnt/β-catenin signaling pathway, SDF-1 reduces the migration and proliferation of BMSCs. BMSCs were transduced with empty vector or the siRNA-Wnt3a gene. Then, the cells were treated with SDF-1 drug (100 ng/mL).
CCK-8 confirmed that the proliferation of BMSCs in the SDF-1+siRNA-Wnt3a and siRNA-Wnt3a groups was significantly reduced compared with that in the SDF-1+Null group; however, there was no significant difference between the SDF-1+siRNA-Wnt3a and siRNA-Wnt3a groups (Fig. 5B). According to the analysis of cell cycle detection by flow cytometry, the G1 phase in the SDF-1+Null group was significantly shorter compared with the SDF-1+siRNA-Wnt3a group and the siRNA-Wnt3a group (68.58% vs. 78.60% vs. 77.88%). The G2+S phase was significantly longer in the LV-SDF-1 group than in the SDF-1+siRNA-Wnt3a group and siRNA-Wnt3a group (5.66% +25.76% vs. 3.55% +17.85% vs. 3.74% +18.38%). The cell cycle test results were consistent with the CCK-8 results (Fig. 5C). WB and IF experiments showed that the β-catenin, c-Myc, and Cyclin D1 protein levels in the SDF-1+siRNA-Wnt3a and siRNA-Wnt3a groups were significantly lower than those in the SDF-1+Null group (Fig. 5D, F). The q-PCR experiment showed that the RNA levels of β-catenin, c-Myc, and Cyclin D1 were lower than those in the control group (Fig. 5E). The results indicate that the Wnt pathway is involved in the proliferation of BMSCs.
Inhibition of the Wnt pathway and osteogenic differentiation of BMSCs were reduced by SDF-1 (100 ng/mL)
ALP and ARS showed that in the SDF-1+siRNA-Wnt3a and siRNA-Wnt3a groups, SDF-1 significantly reduced the osteogenic differentiation ability of BMSCs compared with that in the SDF-1+Null group (Fig. 6A, B). WB and IF confirmed that the protein levels of β-catenin, c-Myc, RUNX-2, and OCN in the SDF-1+siRNA-Wnt3a and siRNA-Wnt3a groups were significantly lower than those in the SDF-1+Null group (Fig. 6C, D, F). q-PCR confirmed that the RNA levels of β-catenin, c-Myc, RUNX-2, and OCN in the SDF-1+siRNA-Wnt3a and siRNA-Wnt3a groups were significantly lower than those in the SDF-1+Null group (Fig. 6E). These results show that SDF-1 promotes the osteogenic differentiation of BMSCs through the Wnt pathway.
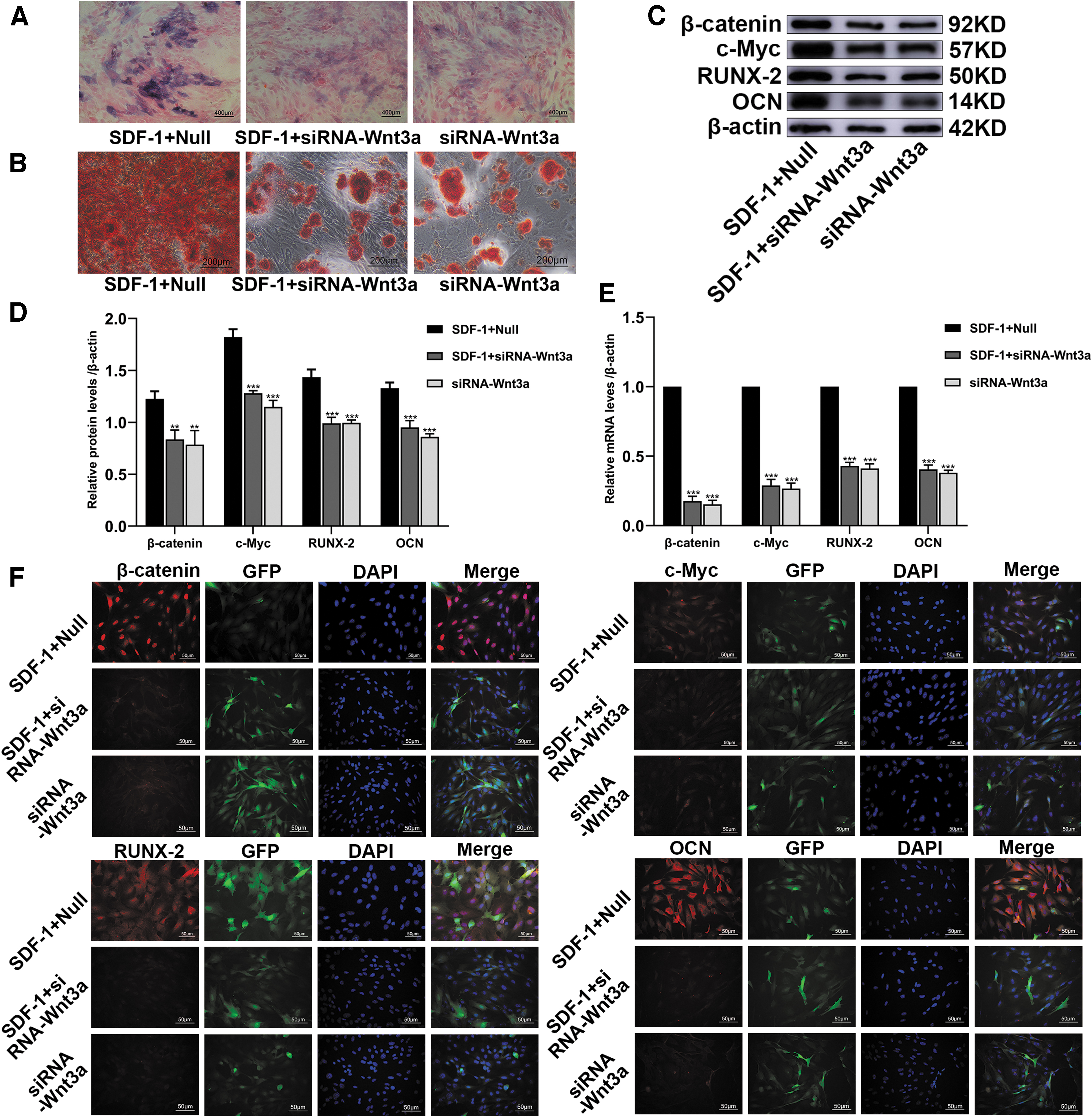
By inhibiting the Wnt/β-catenin signaling pathway, SDF-1 reduces the osteogenic differentiation of BMSCs. BMSCs were transduced with empty vector or siRNA-Wnt3a gene. Then, the cells were treated with SDF-1 drug (100 ng/mL).
Discussion
BMSCs are a group of stem cells with the potential for self-renewal and multidirectional differentiation. Under certain conditions, they have the ability to differentiate into osteoblasts, thereby maintaining the balance of bone metabolism [27]. SDF-1 is from a class of small-molecule proteins secreted by immune and nonimmune cells and plays an important role in bodily immune, allergy, inflammatory, and cell proliferation and differentiation processes [28]. SDF-1 is widely accepted to promote the proliferation and migration of BMSCs in inflammation-associated diseases and can promote osteogenic differentiation. However, the underlying molecular mechanism is unclear.
Lentiviral vectors are genetic vectors developed based on human immunodeficiency virus type 1 and have the ability to infect both dividing and nondividing cells. After foreign genes enter host cells, they are reverse transcribed and effectively integrated into the chromosomal genome to achieve durable and stable expression [29]. Lentiviral vectors do not produce any effective cellular immune response.
In our study, a Transwell experiment was used to confirm the migration of BMSCs, and high SDF-1 expression was shown to promote BMSC migration. Previous evidence supports that high SDF-1 expression can promote the migration of BMSCs and colonize bone marrow in a mouse bone marrow failure model, which is consistent with our results [30].
CCK-8 and cell cycle experiments were used to detect BMSC proliferation. CCK-8 is a highly sensitive, nonradioactive detection method for determining the number of viable cells in proliferation and/or toxicity experiments [31]. Cell cycle detection is commonly used for adherent or suspended cells, and G1, S, and G2 phases are detected by flow cytometry. The proliferation index (G2+S/G1+G2+S%) is used to indicate the rate of cell proliferation [32]. In our study, both the CCK-8 results and the proliferation index were significantly increased in the Lv-SDF-1α group, leading to the conclusion that high SDF-1 expression can promote BMSC proliferation. Studies have shown that endothelial cells self-proliferate thtrough autocrine SDF-1 and recruit mesenchymal stem cells to promote bone formation [33], which is consistent with our results. However, our new WB, q-PCR, and IF results show that high SDF-1 expression can upregulate the expression of Wnt3a, β-catenin, c-Myc, and Cyclin D1. Wnt3a, as the most common Wnt ligand, binds to the LRP5/6 receptor on the cell membrane through autocrine or paracrine forms, leading to more β-catenin accumulating in the cytoplasm of the cell that is then transferred to the nucleus [34]. Combined with specific target factors in the nucleus, specific activation of the downstream target gene c-Myc affects cell proliferation and differentiation [35]. CyclinD1 is a cyclin protein that allows cells to quickly pass through the cell cycle, thereby accelerating cell proliferation [36]. These experimental results indicate that the promotion of BMSC proliferation by SDF-1 may be related to the Wnt pathway.
ALP and ARS were used as indicators to evaluate the osteogenic differentiation of BMSCs. ALP is a phosphomonoester hydrolase, most of which is produced by bone cells and is commonly used as an indicator of early osteogenic differentiation [37]. ARS can react with calcium salts deposited on the surface of cells to produce a dark red compound, which is often used as a marker of advanced osteogenic differentiation [38]. According to our staining results, the intensities of both ALP and ARS were stronger in the Lv-SDF-1α group. Studies have shown that long noncoding RNA-H19 can enhance the osteogenic differentiation of BMSCs by upregulating the expression of SDF-1 [39], which is consistent with our results. In addition, in our osteogenic differentiation experiments, WB, IF, and q-PCR revealed a new phenomenon, as the levels of Wnt3a, β-catenin, c-Myc, RUNX-2, and OCN were enhanced in the Lv-SDF-1α group. RUNX-2 plays an important role in osteoblast differentiation, bone matrix protein production, bone formation, and reconstruction and is a recognized key transcription factor [40]. OCN is a vitamin K-dependent calcium-binding protein that plays an important role in regulating bone calcium metabolism and is a biochemical marker of late bone formation [41]. From our experimental results, we speculate that SDF-1 promotes the osteogenic differentiation of BMSCs and may be related to the Wnt pathway.
To verify this hypothesis, BMSCs were transduced with lentivirus carrying the siRNA-Wnt3a gene, resulting in low Wnt3a expression, and these cells were then treated with SDF-1 drug. In our study, the level of SDF-1α protein was not significantly reduced in the siRNA-Wnt3a group compared with the Normal group and the Null group. Based on this, we infer that the Wnt signaling pathway may be a downstream pathway of SDF-1. This is consistent with previous related reports [42]. The number of migrated cells in the Transwell, CCK-8, and cell cycle detection experiments as well as the ALP and ARS intensities were significantly lower in the SDF-1+siRNA-Wnt3a and siRNA-Wnt3a groups than in the SDF-1+Null group. In addition, WB, q-PCR, and IF revealed that in the SDF-1+siRNA-Wnt3a and siRNA-Wnt3a groups, the β-catenin, c-Myc, Cyclin D1, RUNX-2, and OCN expression levels were significantly reduced. However, these experimental results showed no significant difference in the SDF-1+siRNA-Wnt3a and siRNA-Wnt3a groups.
Therefore, our research demonstrates that SDF-1 promotes the migration, proliferation, and osteogenic differentiation of BMSCs at least in part through the Wnt signaling pathway. In the future, we will further use in vitro experiments to confirm this conclusion.
Conclusions
In summary, our results indicate the promotion of BSMC migration, proliferation, and osteogenic differentiation by SDF-1, at least partly through the Wnt signaling pathway. We herein provide a new strategy and possibility for clinicians in diseases such as nonunion fractures, bone defects, and osteoporosis.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the National Natural Science Foundation of China (no. 81860400) and General Hospital of Ningxia Medical University (Clinical Medicine Research Center of Autonomous Region) Open Project (no. 020007004127).