
Abstract
At present, the number of diabetes patients has exceeded 537 million worldwide and this number continues to increase. Stem cell therapy represents a new direction for the treatment of diabetes; the use of stem cells overcomes some shortcomings associated with traditional therapies. Functional β cells play an important role in the pathogenesis of diabetes. As therapeutic targets, functional β cells are restored by a variety of stem cells, including pluripotent stem cells, mesenchymal cells, and urine-derived stem cells. Although all types of stem cells have their own characteristics, they mainly promote the repair and regeneration of β cells through directional differentiation, immunomodulation, and paracrine signaling after homing to the injured site. However, stem cell therapy still faces many obstacles, such as low long-term cell survival rate after transplantation, low maintenance time of blood glucose homeostasis, immune rejection, and tumorigenesis. Recently, genetically edited pluripotent stem cells and the cotransplantation of mesenchymal stem cells and islet cells have made significant progress in improving the efficacy of stem cell transplantation processes, also providing powerful tools for the study of the mechanisms underlying diabetes and disease modeling. In this review, we first focused on: (1) stem cells as a pool for the differentiation of insulin-producing cells; (2) stem cells as a source for regenerative repair of damaged islets and as a potential cotransplanted population with islets; (3) the potential of combining gene editing with stem cell therapy; and (4) selection of the stem cell transplantation approach. Based on these topics, we discuss the challenges within the field of adapting stem cell-supported and stem cell-derived transplantations and the promising routes for overcoming these problems.
Introduction
The prevalence of diabetes has increased dramatically, and this trend will not be reversed in the coming decades. More than 537 million adults aged 20–79 years suffer from diabetes globally (1 out of 10 people); it is estimated that by 2045, the number of patients with diabetes will reach 783 million [1]. Therefore, diabetes, as a chronic metabolic disease, is an urgent and growing global health problem. With the progression of the disease, the vast majority of diabetes patients will have serious chronic complications, which will severely affect their quality of life or even shorten their life span, and pose a tremendous economic burden to families, societies, and ultimately to countries.
To date, there is no cure for diabetes. All patients with type 1 diabetes mellitus (T1DM) and those with late type 2 diabetes mellitus (T2DM) have been treated with exogenous insulin injection to control their blood glucose levels. However, there are some disadvantages associated with the current method of subcutaneous insulin injection, which does not achieve the efficient glycemic control provided by endogenous insulin secretion [2]. Insufficient insulin doses could lead to hyperglycemia, which promotes the occurrence and development of chronic complications of diabetes. Excessive insulin doses lead to hypoglycemia, hypoglycemic coma, and even death [3,4].
Furthermore, exogenously administered insulin lacks C-peptide, which plays an important role in the prevention and amelioration of diabetic microvascular lesions, especially those associated with diabetic nephropathy [5].
In recent years, advances in islet isolation and purification methods, as well as in immunosuppressive regiments, have made islet transplantation a therapeutic option for patients with T1DM to achieve a more physiologic form of glycemic control [4]. Islet transplantation can restore pancreatic endocrine function. Moreover, it does not cause trauma or pose risks to the patients, as observed in case of pancreas transplantation [6].
However, the broad application of clinical islet transplantation is greatly limited due to several major obstacles, including the shortage of donor islets, potential complications of immunosuppressive drugs, and poor graft function after transplantation [4,7,8]. According to the current standard of islet transplantation (Edmonton protocol), at least two donors are required for a single islet transplant [9]. In summary, its widespread application is impractical at present. Therefore, there is an urgent need to find new effective ways to treat diabetes and diabetic chronic complications. The results referred above have promoted efforts to generate functional β cells through differentiation of stem cells or by inducing endogenous regeneration.
Stem cell transplantation has emerged as an alternative treatment for diabetes. In this review, we focus on several promising routes of stem cell transplantation to reestablish endogenous insulin secretion, summarize the recent progress of research on stem cell therapies for DM, and discuss the potential mechanisms underlying their therapeutic effects.
Role of Functional β Cells
The islets of Langerhans constitute the endocrine part of the pancreas, and ovoid clusters of cells are scattered throughout the pancreas. The main function of islets is to maintain blood glucose homeostasis through the coordinated release of hormones, including glucagon, insulin, and somatostatin. It is now widely recognized that β cells in the islets play a crucial role in the pathogenesis of DM. As an essential hormone for regulating plasma glucose levels, insulin is produced and secreted by β cells.
Hence, when insulin levels are insufficient due to the lack of β cell mass or function, or both, it can lead to hyperglycemia and diabetes [10]. In autoimmune-mediated T1DM, β cell function appears to be impaired in a long-term gradual way in prediabetes, and the β cell mass starts to decrease rapidly before the diabetic stage. Although β cell function can be temporarily relieved after initial insulin treatment, continued autoimmunity and elevated workload will eventually lead to recurrence of cellular exhaustion and cell death. In T2DM mediated by metabolic mechanisms, the increased demand for insulin during the initial insulin resistance period leads to the compensation of β cell function and mass, but once β cell work is overloaded, it will result in dedifferentiation, functional failure, and even β cell death [11,12].
C-peptide, a bioactive peptide also derived from β cells, may exert a notable function in the treatment of diabetic microangiopathy, especially diabetic nephropathy [13]. Thus, β cell damage is the primary prognostic determinant of DM. As a result, therapies designed to prevent or limit β cell damage and/or promote β cell repair or regeneration have great clinical significance.
Stem Cells
Stem cells are an undifferentiated population of cells that have the unique ability to self-renew and maintain multipotency and clonogenicity. The potential for stem cells' differentiation into either multiple lineages or limited cell types is determined by their pluripotency [14]. According to their developmental origin, stem cells can be divided into three major categories: embryonic stem cells (ESCs), induced pluripotent stem cells (iPSCs), and adult stem cells (ASCs). Stem cell transplantation is the most promising cure for diabetes (Table 1). Its main repair mechanism may include direct differentiation, paracrine action, and immunomodulation [15].
Summary of Reports on Stem Cells: Results Observed Following In Vivo Stem Cell Transplantation
Human.
Allogeneic.
AT, adipose tissue; BMSC, bone marrow mesenchymal stem cell; ESC, embryonic stem cell; hESC, human embryonic stem cell; IC, islet cell; IPCs, insulin-producing cells; iPSC, induced pluripotent stem cell; MSC, mesenchymal stem cell; NOD, nonobese diabetic; pIPCs, precursors of insulin-producing cells; SC, stem cell; SCID, severe combined immunodeficient; SD, Sprague Dawley; USC, urine-derived stem cell; WS, Wolfram syndrome.
Cell Sources for Differentiation Into Insulin-Producing cells
Embryonic stem cells
ESCs are pluripotent stem cells derived from embryos that are isolated from the inner cell mass of the blastocyst [16]. Human embryonic stem cells (hESCs), which are specialized cells with the potential to differentiate into all three major germ layers, have the ability to display unlimited proliferation and directional differentiation.
According to the four-stage protocol introduced by Zhang et al. [17], as a modification to the five-stage protocol described by D'Amour et al. [18], hESCs can differentiate into endocrine cells that express pancreatic hormones but do not respond to glucose stimulation in vitro, further to mature insulin-producing cells (IPCs) that secrete insulin/C-peptide in response to glucose stimulation. Rezania et al. [19] verified that hESCs could first differentiate into pancreatic progenitor cells and then further mature into functional IPCs in diabetic mice treated with insulin and maintain normal blood glucose levels for 2.5–4 months after the withdrawal of exogenous insulin.
However, these protocols usually lead to polyhormonal (insulin+/glucagon+/somatostatin+) cells at the end of in vitro differentiation. Moreover, it takes about 23 weeks for pancreatic progenitor cells to mature in vivo and resolve diabetes. Therefore, the seven-stage protocol of Rezania et al. [20] first introduces vitamin C to produce PDX1+/NKX6.1+ pancreatic progenitor cells as the indispensable precursors and then uses a particular combination of ALK5 inhibitors, AXL inhibitors, and thyroid hormone (T3) to obtain highly differentiated monohormonal IPCs in vitro. These cells can express key markers of mature β cells, including MAFA (a basic leucine zipper transcription factor), and result in the control of diabetes in mice within 40 days after transplantation.
To obtain β cells with dynamic insulin secretion properties, Nair et al. formed enriched β-clusters by sorting and reaggregating immature endocrine cells at the final stage of differentiation, driving mitochondrial oxidative respiration to induce metabolic maturation. Under these conditions, the derived β cells also showed an increase in mitochondrial energization and calcium signaling under glucose stimulation [21]. Meanwhile, the reduction in cell cluster size, the elimination of transforming growth factor β inhibitors, and the use of serum-free medium in the latest differentiation protocol resulted in the dynamics insulin release of derived cells, which greatly improved glucose tolerance after transplantation into mice [22]. To a large extent, the native structure of islets is generalized in 3D (three-dimensional) differentiation cultures.
Notably, Hogrebe et al. [23] overcame the requirement for 3D culture in stem cell differentiation by manipulating actin cytoskeleton, thus robustly produced glucose-responsive IPCs in planar culture. After transplantation underneath the kidney capsule of streptozotocin (STZ)-induced diabetic mice, the hESC-derived β cells produced by the planar protocol reversed severe preexisting diabetes better than the suspension protocol.
However, immunologically unmatched hESCs face difficulties in avoiding cell destruction caused by autoimmune reactions and allogeneic transplantation reactions [24]. Recently, Sui et al. [25] obtained matching pluripotent stem cells from the skin cells of patients with T1DM through somatic cell nuclear transfer. This patient-specific nuclear transfer embryonic stem (NT-ES) cell can not only differentiate into C-peptide-positive cells but also show better differentiation efficiency than some iPSC strains. NT-ES-derived β cells also display glucose regulation similar to that of human pancreatic islets cells following transplantation. Nevertheless, ethical issues and teratoma formation are still obstacles to the clinical application of ESCs, and the effectiveness and safety of their transplantation still need to be ascertained through further research [26].
Induced pluripotent stem cells
The discovery of iPSCs is a milestone in medicine and life science. By simultaneously transducing four genes, namely OCT4, KLF4, SOX2, and c-MYC, terminally differentiated fibroblasts derived from mice or humans can be reprogrammed into stem cells with characteristics similar to those of ESCs [27,28]. Therefore, iPSCs have become a new source of stem cells for therapies aimed at treating diabetes. Similar to ESCs, they can also be transformed into pancreatic β-like cells in vitro and act by releasing insulin and C-peptide [29].
By reprogramming adult fibroblasts with three transcription factors (SOX2, OCT4, KLF4), iPSCs that could differentiate into IPCs from patients with T1DM were generated in 2009. However, these cells had low transformation efficiency, lack of function in vivo and in vitro, and could not correctly express β cell genes [30]. The source of iPSCs can also influence the success of β cell differentiation. The study by Jeon et al. [24] compared embryonic fibroblasts and NOD mouse pancreas-derived epithelial cells (NPEs) from nonobese diabetic mice and found that NPE-iPSCs could more efficiently differentiate into IPCs. In addition, NPE-derived β cells could express more pancreas-specific genes and restore blood glucose after transplantation.
Pagliuca et al. [31] successfully differentiated hPSCs into functional human pancreatic β cells. These monohormonal IPCs from nondiabetic (ND) donors could not only remain monohormonal after transplantation but also secrete insulin in response to glucose in vivo. Subsequently, Millman et al. [32] extended this method to produce IPCs from T1DM patients. They found that T1DM IPCs did not differ functionally from ND IPCs in vivo and in vitro, and both expressed certain β cell markers, such as PDX1 and NKX6.1. Furthermore, these T1DM IPCs could reverse alloxan-induced hyperglycemia in mice after transplantation. This method of obtaining IPCs from diabetic patients themselves makes it possible to reduce immune rejection after transplantation.
Mesenchymal stem cells
Mesenchymal stem cells (MSCs) are multipotent ASCs capable of multilineage differentiation. They usually reside in the bone marrow stroma, but have also been found in adipose tissue, spleen, liver, umbilical cord blood, peripheral blood, and other mesenchymal tissues [33]. Over the years, MSCs have been extensively studied in animal experiments and clinical trials as a cell therapy for diabetes because of their easy availability, immunomodulatory properties, nutritional activity, and anti-inflammatory effects.
In 2004, three groups of researchers reported that murine bone marrow-derived MSCs could be successfully differentiated into IPCs [34 –36]. Timper and her group [37] reproduced these early observations through using human adipose tissue-derived mesenchymal stromal cells (hAT-MSCs). Gabr et al. [38] induced 5%–10% human MSCs to differentiate into IPCs through a three-stage differentiation protocol in vitro, and the transplantation of these cells into nude mice with STZ-induced diabetes rapidly reversed their diabetes within 7–10 days.
However, although Xin et al. [39] modified and improved the induction strategy to improve the differentiation efficiency of MSCs, it is still less compared with pluripotent stem cells. The life span of differentiated IPCs and the need for repeated injections of these cells after transplantation remain to be verified. Moreover, it is still unknown whether differentiated MSCs can maintain their immune regulatory properties [40].
Urine-derived stem cells
Urine-derived stem cells (USCs), a subpopulation of cells isolated and expanded from urine [41], have surface markers similar to those of MSCs. Therefore, USCs display similar biological characteristics as stem cells, such as self-renewing ability, high scalability, multidifferentiation potential, and paracrine properties. Compared with MSCs, USCs are advantageous in that they can be obtained through a noninvasive, simple, safe, and low-cost procedure regardless of the age and health status of the donor. In addition, the high telomerase activity in urine cells contributes to improve reprogramming efficiency but not to teratomas or tumors in vivo [42].
Recently, Hwang et al. [43] have successfully differentiated human USCs (hUSCs) into IPCs. Although not as well as islet cells, the IPCs could increase insulin secretion in response to high glucose concentrations. However, it remains to be further investigated whether diabetes can be reversed in vivo.
Regenerative repair and cotransplantation
In addition to differentiating into IPCs to treat diabetes, MSCs and USCs are also sources of regenerative repair in damaged islets and potential cotransplantation groups with islets.
In 2006, Lee et al. [44] observed that a small proportion of the intracardiac infused bone marrow mesenchymal stem cells (BMSCs) spontaneously differentiated and expressed PDX-1 and human insulin after homing in to the pancreas of diabetic mice. More importantly, they played a role in reversing diabetes by increasing the number of islets and IPCs in diabetic mice and may have beneficial effects on other tissues throughout the body.
Several studies have suggested that AT-MSCs have more advantages in clinical applications than BMSCs. In addition to being obtained through minimally invasive methods and easy to isolate, AT-MSCs show a greater immunomodulatory capacity, and their growth and differentiation potential does not decline with age [45 –47]. Amer et al. [48] found that 8 weeks after the injection of IPCs differentiated from AT-MSCs into diabetic rat pancreas, there was no significant difference in the blood glucose and insulin levels between rats in the transplantation group and normal rats. This may be due to the fact that the IPCs became more mature in vivo or that they facilitated the regeneration of endogenous islets in rats.
Arany et al. [49] showed that hematopoietic stem cells (HSCs) and MSCs have different mechanisms of action, which are mainly manifested in their different time courses for improving glycemic control in diabetic mice. Improvement of glucose tolerance and an increase of β cell mass were observed in mice 3 weeks after transplant of HSCs, indicating that HSCs may achieve its therapeutic effect by enhancing β cell regeneration of progenitor cells and the proliferation of residual β cells in islets.
Moreover, it was found that single injections of MSCs were rapidly removed from the pancreas, suggesting that it is most likely to have a rapid impact on the function of the remaining β cells in diabetic mice through the release of trophic factors. However, multiple intravenous MSC infusions also led to the restoration of the structure and number of islets and to the continuous improvement of glucose tolerance and insulin secretion in T2DM rats [50]. For T1DM, the shift in cytokine profile to anti-inflammatory and the balance between Th1 and Th2 may explain why MSCs could achieve long-term reversal of diabetes [51].
The performance of IPCs derived from USCs after transplantation remains to be verified, but recent studies suggest that the injection of USCs may be beneficial to the improvement of diabetes and its complications.
Zhao et al. [52] found that the injection of hUSCs through the tail vein can prolong the survival time of mice with high-dose-STZ-induced diabetes and improve glucose tolerance and insulin levels in mice with multiple low-dose-STZ-induced diabetes. These results suggest that hUSCs not only have a protective effect on damaged β cells but also promote their functional recovery, by providing signals as a cell source and eliciting paracrine functions to promote pancreatic islet revascularization. In a study by Dong et al. [53], the homing of intravenous hUSCs was only observed in the pancreas and kidneys of diabetic rats. However, they are known to protect the myocardium, bladder, and glomeruli by inhibiting apoptosis and improving fibrosis, so as to prevent the occurrence of diabetic complications.
The contact and coculture of MSCs and islet cells can provide a microenvironment conducive to the functional and morphological reconstruction of damaged islets [54]. Therefore, some recent studies have chosen to cotransplant islet cells with MSCs to explore whether the long-term control of diabetes could be achieved.
Navaei-Nigjeh et al. [55] found that cotransplantation of islet cells and AT-MSCs needed only half of the number of cells required for islet transplantation alone, but the effect was similar in a diabetic rat model. The mechanisms underlying the above phenomenon may be associated with the fact that MSCs maintain a more complete islet morphology, enhance the revascularization of islets, and, through paracrine signaling, lead to the secretion of a variety of nutritional and immune regulatory factors. The therapy can relatively reduce mass required for successful islet transplantation and provide ideas for alleviating the shortage of donor islet materials.
When cotransplanted with islets, autologous MSCs seem to produce local immune privilege sites and delay allograft rejection in vivo, thereby promoting graft survival [56]. Westenfelder et al. used the strategy of cocultivating MSCs and islet cells into new islets. After transplantation, they could achieve complete blood sugar control in diabetic mice and reduce insulin consumption in diabetic pet dogs [57,58]. In a clinical study conducted by Wang et al. [59], three patients with chronic pancreatitis who underwent total pancreatectomy were infused with autologous MSCs through the portal vein during islet transplantation. During the 12-month follow-up period after transplantation, they showed improved glycemic control, and no direct adverse events associated with this cell therapy were observed.
Gene editing
The field of gene engineering is developing rapidly. IPCs derived from iPSCs have been generated by correcting pathogenic genes in monogenetic diabetes, which performed better than the previous patient-derived iPSCs in vivo and in vitro. Ma et al. [60] generated iPSCs from skin fibroblasts of a patient with permanent neonatal diabetes mellitus, and these differentiated into insulin-negative cells. After gene editing of these cells using CRISPR/Cas9 system, about 53% of these cells were insulin positive, which could maintain the blood glucose homeostasis of STZ-induced diabetic mice after transplantation.
In Maxwell's study, iPSCs from two patients with Wolfram syndrome (WS) were studied, and the differentiation of autologous highly functional IPCs was achieved using the CRISPR/Cas9 technique to correct for diabetes-causing variants in WS. This protocol is a six-stage differentiation process that is modulated by the cytoskeleton of the pancreatic cells. After correction, the function and differentiation efficiency of these cells were significantly improved, and the expression levels of chronic endoplasmic reticulum stress-related genes that have been shown to mediate β cell apoptosis were significantly reduced. These IPCs maintained blood glucose homeostasis for up to 6 months after transplantation in mice with previously severe diabetes [61].
The advantage of iPSC transplantation is that autologous cells can be edited based on the patient's disease-inducing genes, which can not only eliminate alloimmunological rejection but also provide the patient with individualized cell sources. However, it is possible that autologous iPSCs may induce a T cell-dependent immune response [62]. Furthermore, in most cases, diabetes is multicausal, and the application of gene editing is limited.
Routes of stem cell transplantation
Experiments to explore the effect of stem cell transplantation on blood sugar control in diabetic animal models have adopted a variety of approaches. Kono et al. [63] showed that hAT-MSCs can be transplanted into a diabetic mouse model by tail vein injection. Dong et al. [53] further proved that it is feasible to transdifferentiate stem cells into IPCs through intravenous injection. However, the venous route lacks reliable and good target tissue absorption because stem cells in the venous system may be trapped in the capillaries of the lung. Hence, stem cells transplanted using this approach mainly regulate the pathophysiological process through a combination of local paracrine and systemic effects.
The liver, as the body's blood pool, has sufficient nutrients and space for islet cell survival. Hepatic infusion through the portal vein is also convenient to perform, with little damage and high safety. Therefore, the liver is currently the preferred site for clinical islet transplantation. However, some studies have indicated that this implantation site has the characteristics of low oxygen tension, instant blood-mediated inflammatory reaction, and active innate immune system, which render it unsuitable to provide the best microenvironment for the survival of islet cells [64 –66].
Transplantation under the renal capsule is also a widely used approach in stem cell transplantation experiments. Compared with other transplantation approaches, this approach tends to be less prone to rejection, has better transplantation effects, and higher stability [67]. Moreover, it is easier to perform nephrectomy and facilitate histological examinations [68]. Nevertheless, due to the limited space in the human renal capsule, the number of transplantable cells is relatively small, and fewer islets can survive and function after transplantation [69]. In addition, the clinical translation of subcapsular renal transplantation also faces the challenge of common diabetes-related renal complications [70].
The omentum and striated muscle can also be considered as transplantation sites because of their accessibility and high angiogenesis ability. Svensson et al. [71] showed that muscles can provide implanted islets with three times the number of blood vessels provided by the subcapsular area of the kidney. Bartholomeus et al. found that the immature porcine β cell implants transplanted into the omentum could have twice as much insulin reserves and proliferative activity as those transplanted into the renal capsule [72].
In summary, there is lack of evidence on which modality is the most effective or has the least number of complications. To define the best approach for transplantation, specific studies should be designed to track the effects of stem cell homing and different transplantation sites on stem cell proliferation.
Conclusions and Future Prospects
In recent years, stem cell-based therapies have provided a new research direction for the treatment of diabetes, and each candidate stem cell has its own advantages and disadvantages. Moreover, some issues need to be solved (Fig. 1) with regard to the clinical application of stem cell therapies.
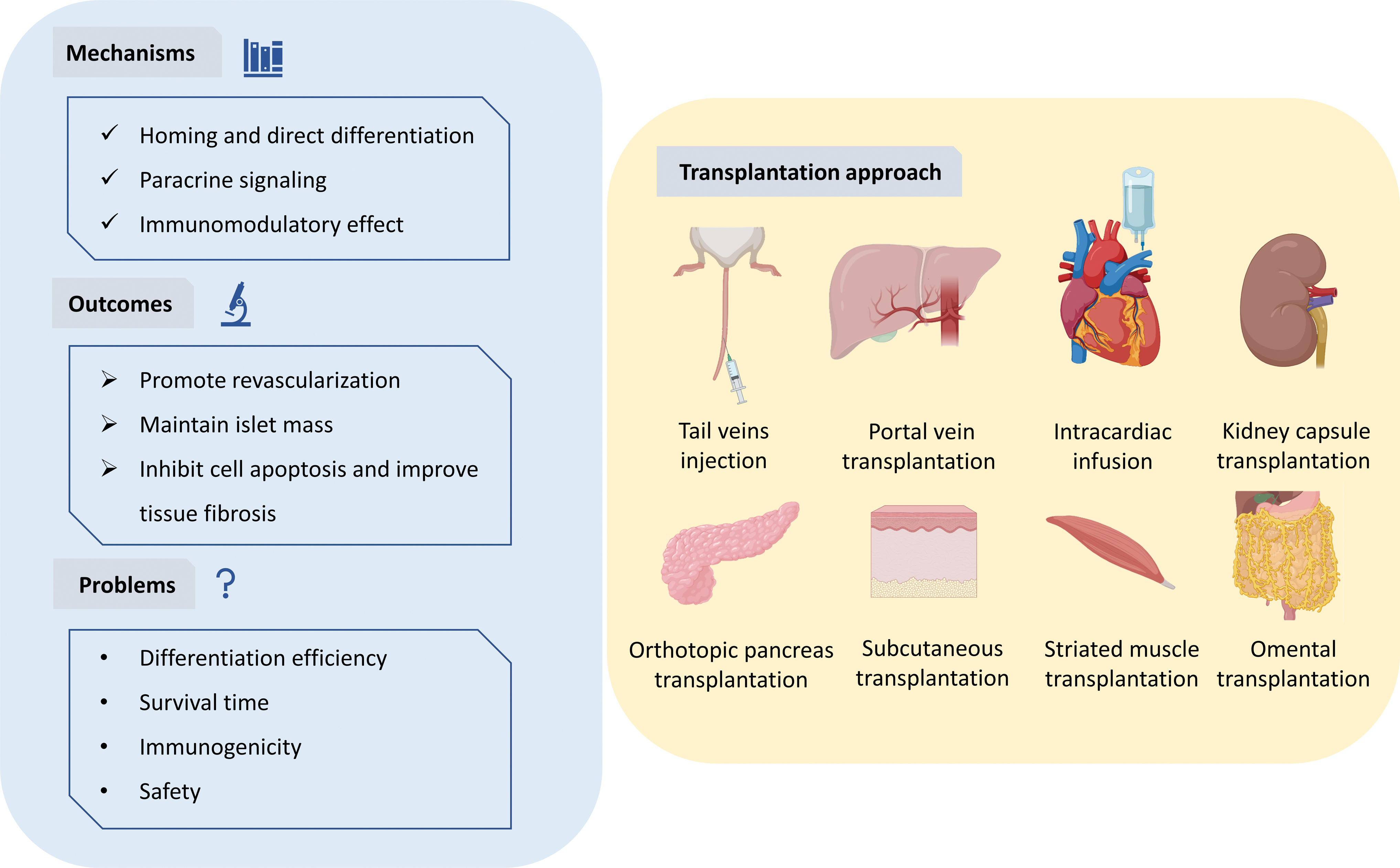
The summary of stem cell transplantation for treating diabetes mellitus. The main mechanism of stem cell therapy lies in the homing of stem cells to the kidney or pancreas, differentiating into insulin-producing cells, paracrine signals, and immunomodulatory effects. These can reverse diabetes through promoting the reconstruction of blood vessels around the cells, maintaining the mass of islets, and inhibiting apoptosis and improving tissue fibrosis. Stem cells can be infused through blood vessels, such as the tail vein, portal vein, and even the left ventricle; they can also be transplanted to the kidney capsule, pancreas, subcutaneous, striated muscle, and omentum. Color images are available online.
First, the microenvironment under the conditions of diabetes plays a vital role in the survival, differentiation, migration, and repair of stem cells. It has been shown that the decreased synthesis of glycosaminoglycans and proteoglycans in the surrounding tissues and the generation of advanced glycosylation end products can inhibit the proliferation of stem cells in diabetes patients [73,74]. Oxidative stress may also affect the paracrine effects of MSCs, and high glucose levels directly impair their migration capacity [15].
Hence, it is important to increase the cell's retention ratio and ensure their long-term effects after transplantation. Further research should consider the use of biological materials, such as porous scaffolds combined with growth factors, oxygen-generating particles, and transferable microvascular networks, or low-frequency electrical stimulation and other physical stimulation methods to promote revascularization and prolong the life span of the implanted cells in vivo.
Another main obstacle to the transformation of stem cell therapy into clinical applications is the immunogenicity of stem cells. Autologous cell transplantation is affected by the state and quantity of autologous cells. For the transplantation of MSCs, Zhang et al. proposed that allogeneic MSCs might be a more appropriate choice than autologous MSCs. Systemic diseases, such as diabetes, can change the inherent properties of MSCs and, thus, impair their protective function. Therefore, it is difficult to isolate a sufficient number of highly active autologous MSCs for patients with these diseases [75,76].
However, allogeneic transplantation rejection is difficult to avoid, especially the autoimmune rejection associated with T1DM. Immunosuppressive drugs can reduce the potency of cells and are associated with side effects, while the use of biological packaging materials is limited due to delayed insulin release kinetics and poor peripheral blood supply. Therefore, more experiments are needed to determine whether the use of immunosuppressants or their minimum dosages, as well as better packaging devices, is required. It should be more clearly discussed at which stage the differentiation of MSCs into IPCs can achieve glycemic control with maximal immunosuppressive effects.
Recently, gene editing and cotransplantation have become popular topics in the field of stem cells. The CRISPR/Cas9 gene editing tool can help circumvent immune rejection by dismantling major histocompatibility complex-related components [77], and it can also correct for diabetic variants to enable the robust differentiation of autologous cells [61]. Cotransplantation can promote synergy among the different cell types and help compensate for their weaknesses.
These methods provide novel ideas for solving the problems stated above. In a recent ongoing clinical trial of pluripotent stem cell-derived beta cells, Vertex Pharmaceuticals has achieved positive results in the first patient with T1DM from the phase 1/2 clinical trial using fully differentiated pancreatic islet cell replacement therapy. At day 90 after receiving single-dose treatment, this patient has resumed insulin production and decreased daily exogenous insulin use by 91%. This is the first trial data to confirm that such a cell therapy can significantly restore islet cell function in a patient with T1DM [78]. And ViaCyte and CRISPR Therapeutics are enrolling patients for the first allogeneic, gene-edited cell replacement therapy to enter clinical trials; the phase 1 clinical trial aims to evaluate its tolerability, safety, and immune evasion in patients with T1DM [79].
For T2DM patients characterized by the combination of insulin resistance and relative insulin deficiency, current treatments cannot alleviate inflammation or reverse their β cell damage [80]. MSC transplantation seems to be a good strategy for the treatment of T2DM due to their properties. In addition to the mechanisms of differentiation into IPCs, repair of β cells, and immunosuppression, MSCs have also been found to improve insulin resistance [81,82] and regulate hepatic glucose metabolism [83]. Some clinical trials have shown that T2DM could be treated by MSC transplantation. However, the duration of therapeutic effectiveness, the need for multiple injections, and the route of administration need to be further studied [84].
In summary, ideal stem cells for the treatment of diabetes should have the following properties: (1) simple acquisition; (2) ability to achieve the long-term reversal of hyperglycemia and to resist the pressure of diabetes both in vitro and in vivo; and (3) safety for clinical use, that is, they must not cause immune rejection and teratomas. To widely use stem cells in the treatment of diabetes in the future, more relevant animal experiments and clinical studies with longer monitoring time are needed; these will help ascertain the safety and effectiveness of new stem cell-based treatment methods.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
The work of the authors was supported by a grant from the Wenzhou Science and Technology Bureau (Y20170004).