
Abstract
Branching morphogenesis is an important process in shaping the arborized structures of several organs. However, the driving force that directs this process from progenitor pools remains incompletely understood. In this lineage tracing study, we investigated the role of Id2+ embryonic progenitor cells in branching organs such as the pancreas, kidney, mammary gland, thyroid gland, and salivary gland. We found that a subset of Id2+ distal progenitor cells in the embryonic pancreas and kidney can give rise to multiple lineages of progeny cells during branching morphogenesis. Id2-labeled cells also supported the postnatal development of the mammary glands. However, Id2+ cells did not contribute to the development of the salivary and thyroid glands. We found the Id2+ cells located in the tip progenitor pools of pancreas and kidney have self-renewal potential and contribute descendants to multiple epithelial cell lineages. Our findings enrich the current model of distal progenitor pools driving branching morphogenesis and provide a new marker to investigate the regularity of branching in these organs.
Introduction
Reciprocal epithelial–mesenchymal signaling drives the process of epithelial structure remodeling called branching morphogenesis, wherein an epithelial rudiment extends and branches iteratively to generate an arborized tree-like structure [1].
Epithelial branching morphogenesis shapes the development of multiple organ systems during embryonic development, such as the lungs, pancreas, kidneys, mammary glands, and salivary glands [2,3]. Each of these organs has unique branching strategies, although the final organ trees appear partially similar [4]. Significant progress has been made in revealing the driving factors during branching morphogenesis, and lineage tracing has shown that branching tips comprise multipotent progenitors with a higher proliferation index than proximal cells [5 –7]. In the mammalian lung, Sox2+ proximal progenitor cells contribute to the formation of conducting airways, and Sox9+ distal progenitor cells give rise to alveolar epithelial cells during pseudoglandular stages (E12.5–E16.5) [8].
Likewise, the multipotent pancreatic progenitors located at the tip of the branching tree during E9.5–E14 give rise to all adult pancreatic cell types. The expansion of tip progenitors leaves behind the trunk progeny that possesses lineage-restricted differentiation potential [9,10]. The embryonic kidney has a unique branching pattern in which Six2+ nephron progenitor cells and Foxd1+ interstitial progenitor cells coordinate with each other in the nephrogenic niche and drive renal organogenesis [11,12]. The early phase of mammary gland development is similar to that of the salivary gland because they both initiate branching morphogenesis as a solid epithelial mass and hollow into double-layered structures of luminal cells and basal cells [1].
The mammary gland is particularly characterized by activated proliferation of terminal end buds during puberty and expansion of alveoli during lactation, whereas the salivary gland finishes branching by repeated cleaving during embryogenesis [13]. Although many studies have revealed the organ-specific and shared mechanism of branching morphogenesis, it is largely unknown whether these organs have a common developmental pattern or similar progenitor cells.
The helix-loop-helix (HLH) proteins are a superfamily of eukaryotic transcriptional regulators that share a common HLH domain [14]. They contribute to driving cell differentiation and lineage commitment by forming dimers [15]. Inhibitors of DNA binding (Id), which possess a conserved HLH domain but lack an effective DNA binding domain, are dominant negative regulators that work by forming nonfunctional heterodimers with other HLH transcription proteins [16,17]. In general, Id proteins play a positive regulatory role in cell proliferation and a negative regulatory role in cell differentiation, and their repression of lineage commitment during development plays a significant role in stem/progenitor cell maintenance [18 –21].
Id2 protein is a member of the Id protein family and is distributed more ubiquitously than other members. It is highly expressed in the mesoderm and endoderm during postgastrulation and then limited to the epithelium in organs arising from the primitive gut [22]. One of its major functions is to control the epithelial cell phenotype [23,24], tissues that undergo epithelial–mesenchymal interactions tend to express Id genes, as observed in the process of branching morphogenesis [25,26]. Inspired by the evidently higher expression of Id2 in the distal tip epithelium, Emma L. Rawlins discovered that the Id2+ distal tip lung epithelium contained embryonic progenitor cells [27].
Although high expression of the Id2 protein indicates active morphogenic activities in the epithelia, the mechanism by which Id2 signaling transforms a simple epithelial bud into a complex arborized network of tubules is unknown in organs generated by branching morphogenesis. The positive effects of Id2 on cell proliferation are extensive and well documented. TGF-β-Id2 axis regulates the proliferation transitions in airway basal stem cells during development and regeneration [28]. Id genes can also sense Smad signals and create a permissive or refractory nuclear environment that defines decisions of cell fate and proliferation [29]. Thereby, we postulated that Id2 expression may mark the branching progenitors covering not only epithelial progenitors, but also some mesenchymal progenitors, especially early embryonic multipotent progenitors during branching morphogenesis.
To clarify whether Id2 protein marked embryonic epithelial progenitor cells and whether these Id2+ progenitor cells were retained in adult organs, we used “knock-in” transgenic mice in which the tamoxifen-dependent CreERT2 recombinase was targeted to the Id2 locus to trace the fate of these cells at different times during organ branching morphogenesis. By inducing and lineage-labeling these transgenic mice at specific time points, we analyzed the temporal and spatial potential of Id2+ epithelial progenitor cells, revealing the distinct regulation pattern of the Id2 protein during development in various organs. Our study also delineates the role of a population of Id2+ epithelial progenitor cells in pancreas, kidney, and mammary gland formation, and not in the formation of thyroid and salivary glands.
Methods
Mice
All mice were maintained in an SPF facility (25°C, 12 h light/dark cycle, food and water ad libitum). All animal procedures were approved by the Animal Ethics Committee of Cheeloo College of Medicine, Shandong University. Also, our study does not involve clinical trial registration. The B6.129S(Cg)-Id2tm1.1(cre/ERT2)Blh/ZhuJ (JAX Stock No:016222, Id2-CreERT2/+ ) mouse line was purchased from Jax Laboratory. Id2-CreERT2/+ heterozygous mice were viable and fertile, but homozygous embryos were miscarried due to defective lung alveolarization. Reporter mice B6.129X1-Gt (ROSA)26Sortm1(EYFP)Cos/J (JAX Stock No: 006148, R26R-EYFP) were obtained from Shanghai Model Organism Center Inc.
All mice were maintained on the C57BL/6 background by back-crossing with C57BL/6 mice. For genotype identification, genomic DNA was extracted from transgenic mouse tails using a DNA extraction kit (Tiangen, China). Genotypes of the transgenic mice were confirmed through PCR using genomic DNA. Allele-specific primers are as follows: Id2-CreERT2 : P1 5′-CTT CCT ACG AGC AGC AT-3′; P2, 5′-CTC ACC TGC AAG GAC AGG AT-3′; P3 5′-GCT CCT GGA CAG GAA TCA AG-3′. R26R-EYFP: P1 5′-AAG ACC GCG AAG AGT TTG TC-3′; P2 5′-AAA GTC GCT CTG AGT TGT TAT-3′; P3 5′-GGA GCG GGA GAA ATG GAT ATG-3′. PCR products were used for genotype analysis.
Animal procedures
For time mating, the mice were mated overnight by placing one male with three females. The day of vaginal plug observation was considered as E0.5, and the day of birth was defined as P0. Pregnant females were gavaged with tamoxifen (2 mg/mouse) (Sigma-Aldrich, St. Louis, MO) dissolved in corn oil to activate Cre recombinase and administered progesterone (1.5 mg/mouse) (Sigma-Aldrich) at the same time. The gestation period lasted ∼19 days, and embryos were delivered by cesarean section at E18.5, due to the high rate of miscarriage in tamoxifen-treated pregnant mice, and the neonates were nursed by lactating mice. Adult mice (P30 onward) received tamoxifen (Sigma-Aldrich) at a dose of 0.25 mg/g body weight at the indicated times.
Immunofluorescence
Mice were anaesthetized and then perfused with 1 mL cold 4% paraformaldehyde (PFA) through the heart to fix the fresh organs for 10 min. The dissected organs from the transgenic mice were placed in phosphate-buffered saline (PBS) and washed thrice, after which the organs were immersed in 4% PFA solution overnight. After fixation, tissues were dehydrated in gradient sucrose in PBS at 4°C, embedded in OCT (Sakura, Japan), and sectioned at 8–10 μm. Slices were stored at −20°C until use. For staining of frozen sections, the dried sections were treated with 0.2% Triton X-100 in PBS and then blocked with 5% donkey serum in PBS.
The following primary antibodies were used at 1:50 ∼ 1:200 dilution for staining: GFP/YFP/BFP (ab6673; Abcam, Waltham, MA); insulin (ab181547; Abcam); glucagon (15954-1-AP; Proteintech, Rosemont, IL); α-amylase (A8273; Sigma-Aldrich); Aquaporin1 (PA5-78805; Thermo Fisher, Waltham, MA); Aquaporin2 (PA5-78809; Thermo Fisher), and WT1 (ab89901; Abcam). Sections were stained with primary antibodies at 4°C overnight, washed with PBS thrice, and incubated with Alexa-conjugated secondary antibodies (Thermo Fisher) for 30 min at room temperature (20°C–25°C). After washing, the sections were mounted with DAPI (ab104139; Abcam) before confocal imaging.
Confocal microscopy
A Zeiss LSM 880 Meta laser scanning confocal microscope was used to visualize the optical sections and analyze the protein colocalization.
Results
Lineage tracing of early/late embryonic Id2+ progenitor cells in branching organs
Despite differences in the regulatory mechanism and cellular responses in various branching organs, common regulatory programs can be identified in the branching morphogenetic control of multiple organs. In this study, we chose the pancreas, kidney, salivary gland, thyroid gland, and mammary gland as typical models for their similarities in driving force and dissimilarities in cellular interplay. To trace the fate of Id2+ cells derived from mouse embryos, we crossed the cell-specific CreERT2 mouse line Id2-CreERT2 with the reported mouse line R26R-EYFP.
When pregnant mice were treated with tamoxifen at a certain point, the inducible Cre-loxP recombination was activated in Id2-CreERT2 ; R26R-EYFP double-positive embryos, leading to permanent labeling of enhanced yellow fluorescent protein (EYFP) in Id2+ cells and all their descendants. First, we conducted a short phase tracing to show the usability of the transgenic mice (Supplementary Fig. S1 in the supplementary material). When pregnant mice were treated with tamoxifen at E10.5, EYFP, and Id2 protein almost completely overlapped in the pancreas of embryos at E13.5.
To determine the proliferation ability and differentiation potential of Id2+ cells at different time points, we chose E10.5 and E14.5 as the early/late induction time points considering the branching schedule of the major organs and finally dissected the double-positive offspring to detect EYFP expression at postnatal periods P7, P15, and P30. EYFP expression was not observed in the control group without tamoxifen treatment. The labeling frequency was highest in the pancreas among these branching organs, and EYFP expression persisted till P30 in the pancreas regardless of the induction time (Fig. 1a–d).
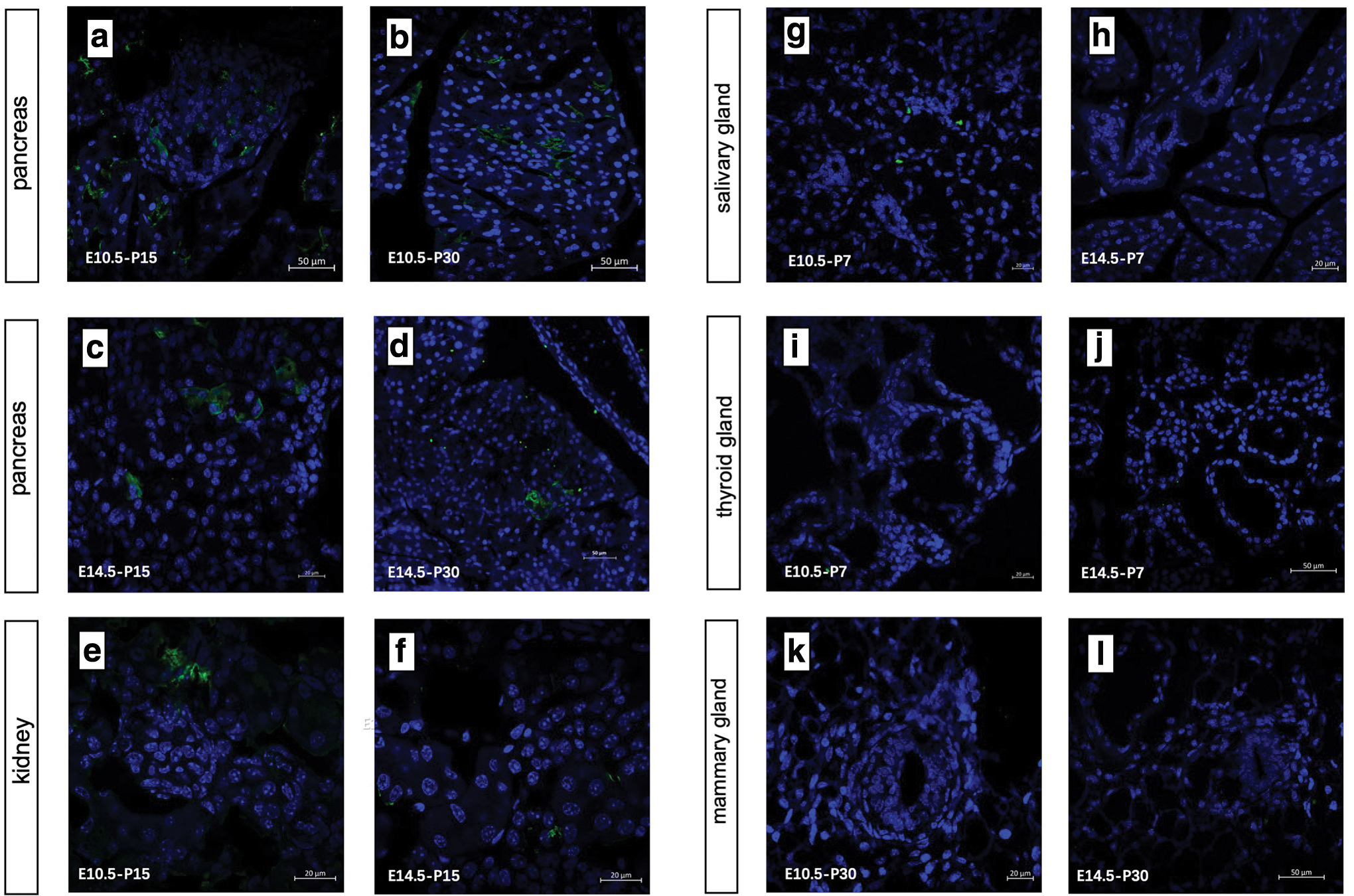
Lineage tracing of early/late embryonic Id2+ progenitor cells in branching organs.
However, the induction frequency was higher in the early embryonic stage than in the late stage, which is in accordance with the time of activity of embryonic progenitor cells. The positive cells in the kidney were detected at P30 when induced at E10.5 and P15 at the latest when induced at E14.5 (Fig. 1e, f). A cluster of several positive cells may descend from a common progenitor. Similar to the pancreas, active renal branching morphogenesis also occurred before E14.5, but Id2 scarcely tagged lineage-restricted progenitors in the kidney during late embryogenesis.
The mammary gland is a special organ with the majority of its branching process occurring during puberty. Therefore, we dissected the mammary glands of adolescent mice, which were treated with tamoxifen at E10.5 or E14.5, but no EYFP was detected in the specimens from all double-positive female mice (Fig. 1k, l). Lineage tracing-targeted Id2 promoter ruled out the possibility that embryonic progenitors were retained in situ and contributed to adult morphogenesis in the mammary gland. Furthermore, we did not find any distinct EYFP cell clusters in the salivary and thyroid glands at P7 above the background (Fig. 1g–j).
One reason may be the differential expression of Id2 proteins in different embryonic organs. More importantly, the negative results in these two organs reflected a distinct branching model that cleaves after solid epithelial outgrowth remodeled the organ morphogenesis. In conclusion, Id2 indicated different self-renewal and proliferation potentials depending on the specific organ, and it can be used as a tool to study the early development of pancreatic and renal progenitor cells.
Id2 expression in embryonic pancreas and kidney development
Previous studies have shown that the Id2 protein is highly expressed within the epithelium during mouse embryogenesis, and its expression level may indicate active morphogenesis. Evidence from a previous study revealed that Id2 marked a subset of self-renewing embryonic progenitors in the developing lung. Apart from the lung, the pancreas is also a typical organ generated from repeated epithelial branching. Since our experiment proved that Id2+ cells contribute to pancreatic and kidney development, we continued to explore the expression of Id2 in embryonic pancreas and kidney.
To further confirm the dynamic expression trend of Id2 in embryonic organs, we performed immunofluorescence for the Id2 protein (red), epithelial marker E-cadherin (green), and DAPI (blue) in the pancreas and kidney from E12.5–E18.5, mainly covering the period of branching morphogenesis.
We noted extensive co-localization of Id2 and E-cadherin for the entire observation duration. Whether pancreas or kidney, Id2 was highly expressed at E12.5, when most cells were in an undifferentiated state (Fig. 2a, d). The red fluorescence signal gradually decreased with time from E12.5 to E15.5 (Fig. 2b, e). At E18.5, the expression of Id2 protein was too low to be detected (Fig. 2c, f). More importantly, Id2 expression was clearly restricted to the distal part of the pancreas, rather than the proximal cells in the fetal pancreas, which indicates that Id2 holds the potential for multipotent progenitor cells or at least lineage-restricted progenitor cells in mouse embryonic pancreas and kidney.

Immunofluorescence of Id2 expression in embryonic pancreas and kidney.
To visually demonstrate the contribution of Id2+ cells during embryonic branching development, we made a short phase tracing in the embryonic stage. We treated pregnant mice with tamoxifen at E10.5, and dissected embryos to at fixed time intervals. In this study, we chose E12.5 and E14.5 as the observation time, for the short time intervals can show the contribution of Id2+ cells to branching events. At E12.5, the labeled EYFP cells reflected the basal expression level of Id2 expression, which also proved the availability of the transgenic mice (Fig. 2g, i). After 48 h, the EYFP expression tended to spread over a wider area (Fig. 2h, j). This distribution pattern of fluorescent cells reflected development characteristics of Id2+ cells. Labeled cells in terminal differentiation may not last for long, but those with ability of self-renewal will extend distally.
Lineage restriction of embryonic Id2+ pancreatic progenitor cells
The pancreas undergoes specialized differentiation into two components during development: 99% exocrine part mainly producing digestive enzymes and 1% endocrine part producing hormones for glucose control. To further investigate the contribution of different time-stamped embryonic Id2+ progenitors, we stained sections of the pancreas with major differentiation markers.
We discovered that Id2+ cells labeled at E10.5 can spin off both exocrine and endocrine progeny, because EYFP+ cells were found to co-express insulin/glucagon (endocrine) and amylase (exocrine) in the P30 pancreatic tissue section under a laser confocal microscope (Fig. 3a–c). The same result was observed at P15, but the result at day 30 was more indicative of the long-term differentiation potential of Id2+ progenitors. The expression of Id2 proteins at E10.5 indicated its multilineage differentiation potential.
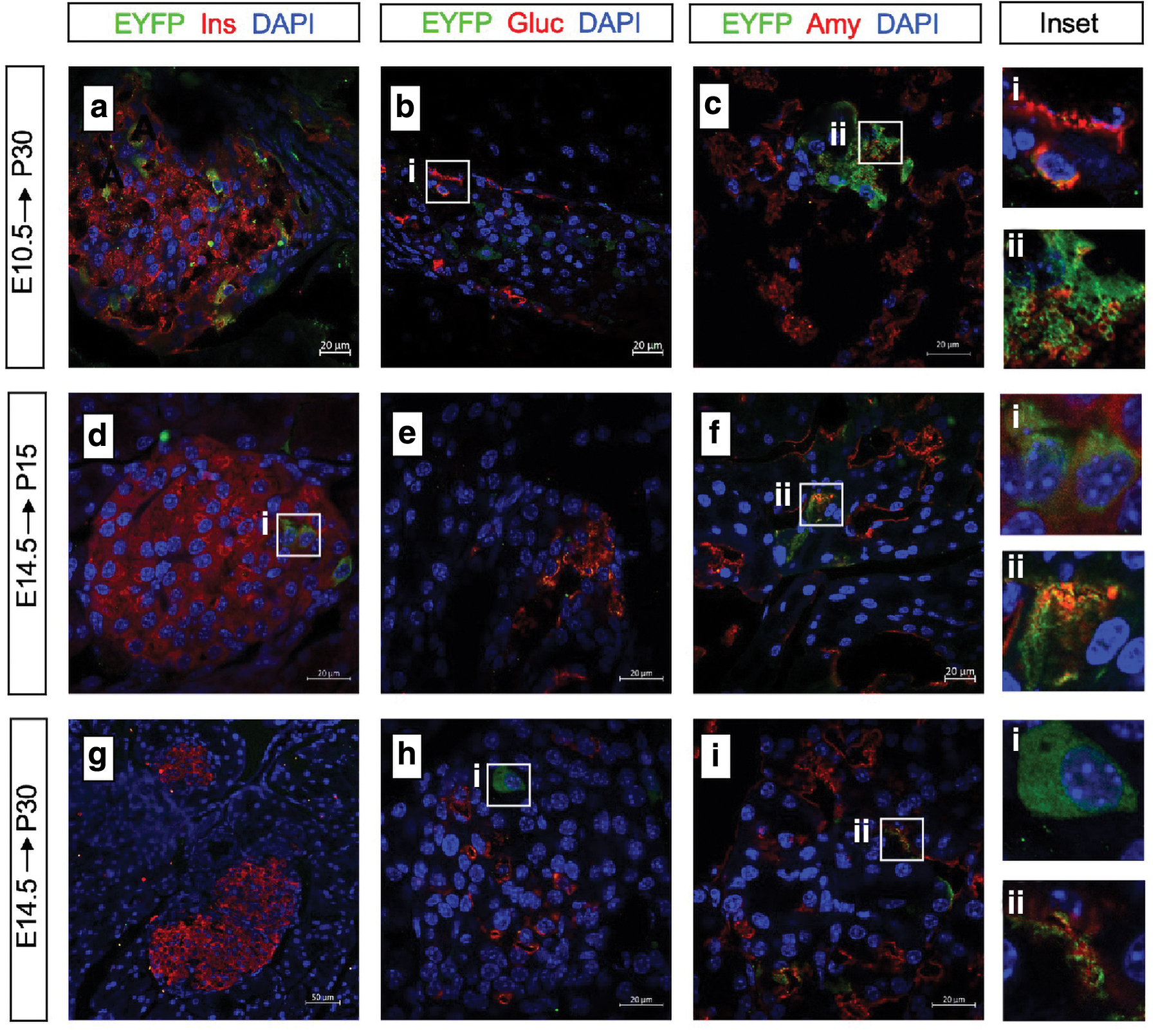
Lineage restriction of embryonic Id2+ pancreatic progenitor cells.
Next, we sought to determine whether the late Id2+ progenitor cells had the same multidirectional differentiation potential. Consequently, we stained sections of the pancreas with the same markers at E14.5, when the organs underwent a second transition of lineage specialization. Although very few EYFP cells co-expressed glucagon and insulin at P15 (Fig. 3d–f), the labeling efficiency of endocrine cells was reduced to zero at P30, and almost all of the labeled daughter cells were positive for amylase or distributed around the duct (Fig. 3g–i), indicating that late Id2+ progenitor cells had exocrine-biased lineage restriction compared to the early Id2+ progenitor cells.
This result was consistent with previous reports claiming that tip progenitor cells have multidirectional differentiation potential, and we confirmed that Id2 expression at a later stage had exocrine potential. Although multiple sections were analyzed to avoid errors, we could not completely rule out stochastic changes in the labeling efficiency due to Cre expression efficiency.
Tracing the fate of embryonic Id2+ progenitor cells in kidney development
The kidney is a sophisticated organ whose formation is initiated by the interaction between the ureteric bud (UB) and metanephric mesenchyme (MM). Mesenchymal progenitors located in the MM undergo a mesenchyme-to-epithelial transition and form renal vesicles that eventually develop into nephrons. This is a continuum shift due to the lack of definitively distinguished markers. Lineage-tracing experiments confirmed that MM can produce all parts of the nephron, and UB contributes to the collecting duct.
Inefficient labeling of postnatal renal sections revealed that Id2 was difficult to label mesenchymal progenitors in the embryonic kidney because of its low expression in the renal mesenchyme. However, several fluorescent cells formed a cluster rather than presenting a scattered distribution. Yet there were obvious fluorescence markers in renal tissue sections on P0, which indicates that Id2 can still label part of progenitors (Supplementary Fig. S2a in the supplementary material). To study the differential potential of positive cells, we chose AQP1, WT1, E-cadherin, and AQP2 for immunofluorescence staining. AQP1 was expressed in the proximal tubules and the thin limbs of the loop of Henle, whereas AQP2 was expressed exclusively in the collecting ducts of the adult kidney.
WT1 and E-cadherin mark the podocytes and distal tubules of the kidney, respectively. We found that the majority of the EYFP+ cells expressed AQP1 and a small number of EYFP+ cells have co-expression with WT1 or E-cadherin. There was little evidence to support co-expression with AQP2 (Fig. 4a–h). AQP1, WT1 and E-cadherin positive cells are derived from MM, whereas AQP2 positive collecting duct are derived from the branching UB. Correspondingly, Id2 tends to express in the nephrogenic zone that extends as a band along the entire surface of a fetal kidney in E14.5 (Supplementary Fig. S2b in the supplementary material). Therefore, Id2 could have labeled a fraction of the tip progenitors in the fetal kidney, and this part of progenitors may have differentiation potential other than collecting duct.
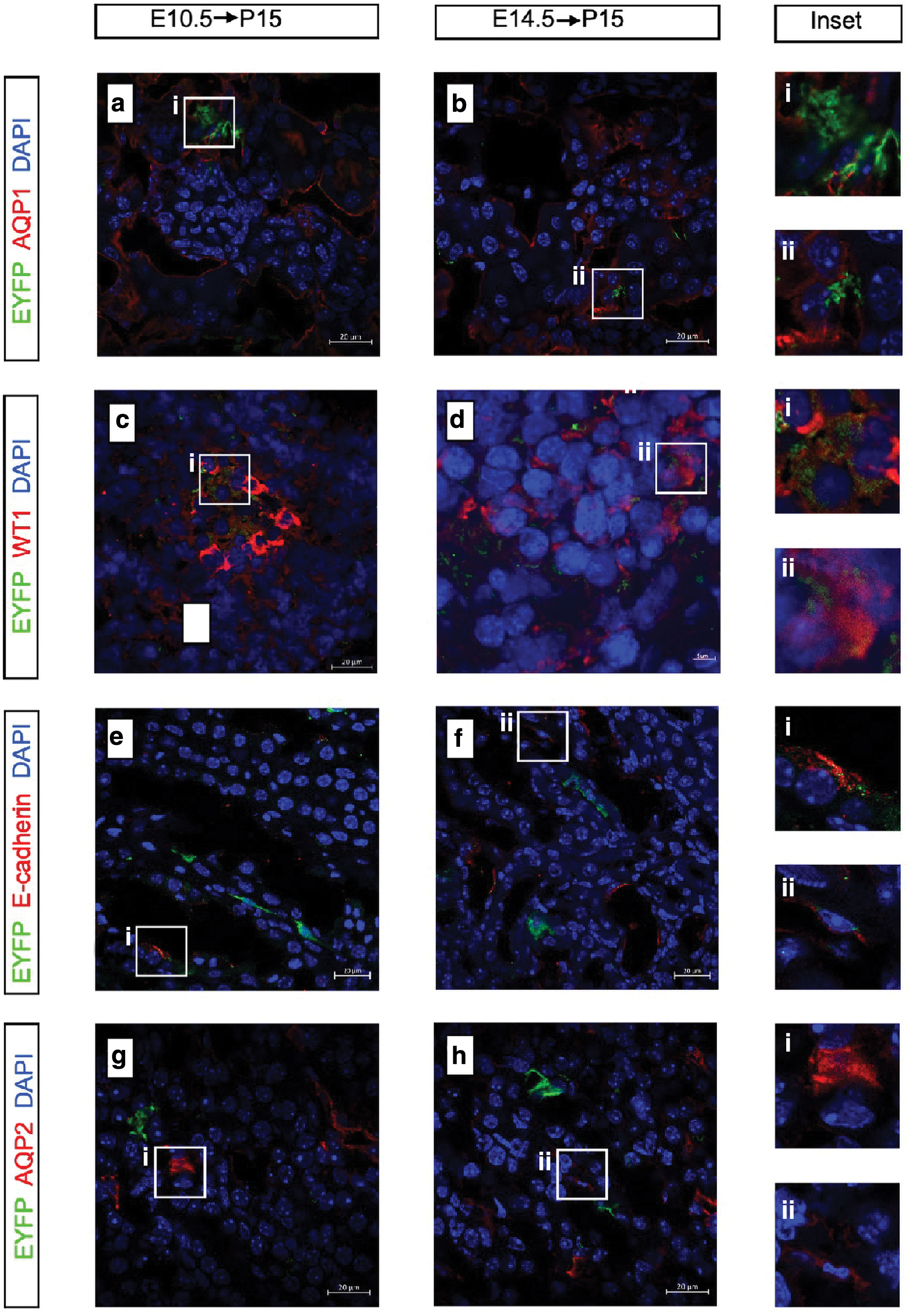
Tracing the fate of embryonic Id2+ progenitor cells in kidney development.
Lineage tracing of Id2+ cells in adult organs
The identification of adult stem cells with therapeutic potential has been a hot topic of research for several years. The lineage-tracing experiment using the Cre/loxp system has been successfully applied to mark adult stem cells, with the goal of choosing a specific gene for a particular cell population. We have found that Id2 can mark embryonic progenitors in certain organs, including the pancreas and kidney. Next, we wanted to explore whether Id2 expression in adult organs could label adult stem/progenitor cells and give rise to multiple cell lineages for a long time. Double-positive mice were given 5 mg tamoxifen at P30 and dissected after 5 or 60 days.
There were some small cells scattered in the pancreas, kidney, and salivary gland after 5 days, reflecting the initial expression pattern in these organs. After tracing for 60 days, no obvious fluorescent signal was detected in any of the tissue sections (Fig. 5a–d). The thyroid gland was hardly labeled with any cells on day 5, which suggested low efficiency of recombination under the Id2 promoter in the adult thyroid gland. Together, these results suggest that the ability of Id2 to label progenitor cells is related to the branching process and does not show such specificity in adulthood. Notably, the mammary gland is a special organ since it achieves full development after birth (Fig. 5e, f).
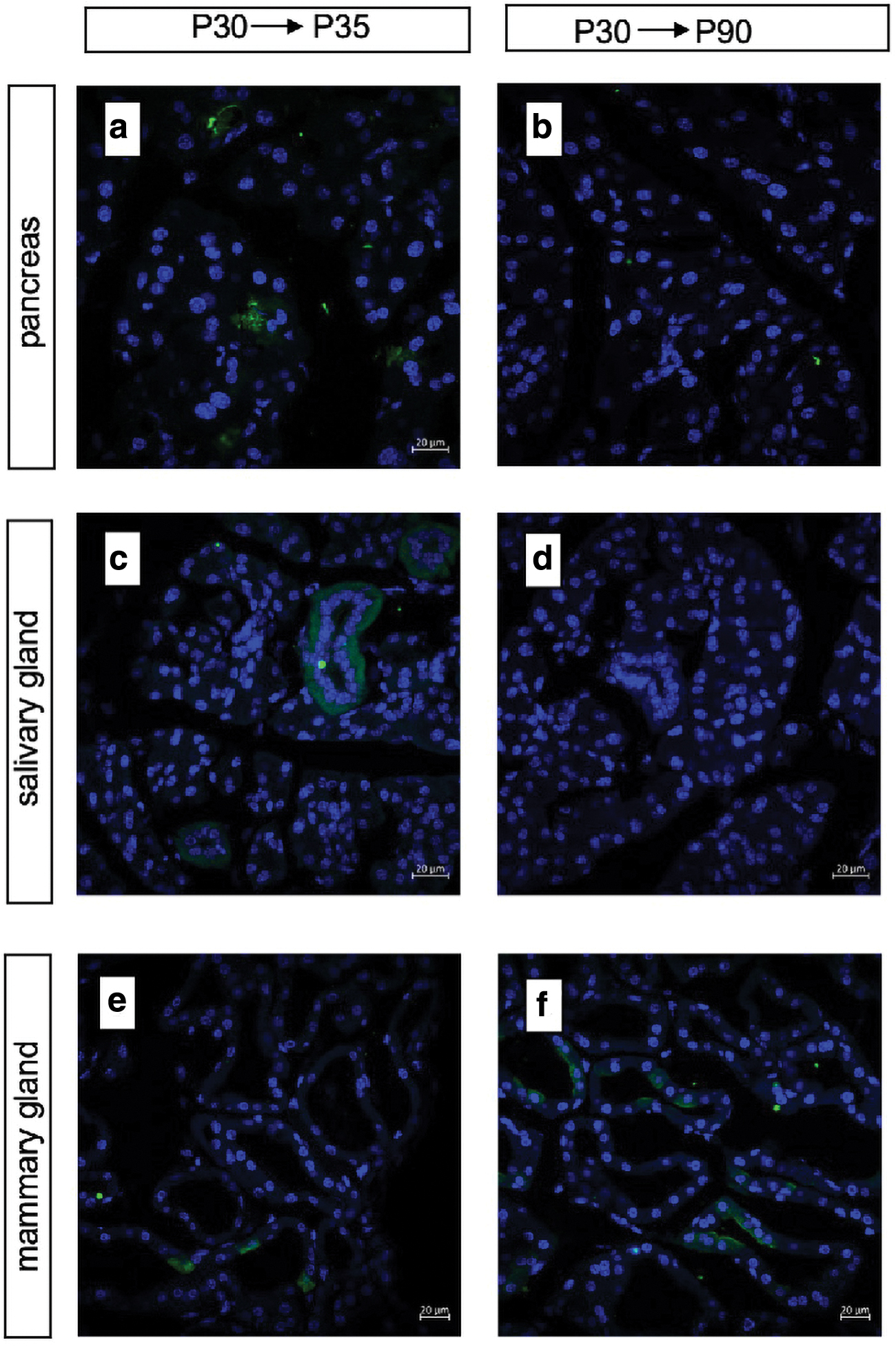
Lineage tracing of Id2+ cells in adult organs.
In this study, we treated prepubescent mice with single gavage administration and analyzed the mammary glands at 0.5 day of lactation. Meanwhile, mice at mid-pregnancy that were administered the same feed were chosen as the control group. The experimental mammary glands showed higher labeling efficiency than the control. Not all acini were labeled, but the positive cells existed in bunches usually within the acinus. Although Id2 is differentially expressed in epithelial cells, stromal cells and immune cells under homeostasis, we found that persistently labeled cells were present in the acinar during pregnancy. In summary, Id2 can partially label mammary gland stem and progenitor cells that contribute to morphology remodeling during adolescence.
Discussion
Our results revealed the commonality and diversity of various branching organs using the Id2-specific tracing method. First, we found that the distal tip progenitor pools in the embryonic pancreas and kidney contain a subset of Id2+ progenitors that can give rise to multiple lineages of progeny cells during branching morphogenesis. Id2 proteins at E10.5 indicated its multilineage differentiation, whereas E14.5 Id2+ progenitor cells had exocrine-biased lineage restriction in fetal pancreas. Id2 could label tip progenitors in the fetal kidney, which mainly differentiated into proximal tubules. The potentiality of multidirectional differentiation decreased gradually with development and disappeared completely in adulthood.
On the contrary, no stable Id2+ cells appeared in the postnatal salivary gland and thyroid gland, thereby reflecting the common branching pattern of the two organs. They initialized branching as an epithelial mass and hollowed into the lumen structure, and Id2 protein was dispensable in this process. Furthermore, treatment of preadolescent mammary glands and analysis during lactation also supported the notion that Id2 plays an important role in mammary gland morphogenesis. Although the pregnant mice were gavaged with sufficient tamoxifen to induce Cre expression, we cannot exclude the possibility that a small proportion of positive cells were not labeled due to Cre recombinase efficiency.
A previous study has demonstrated the regulatory effect of Id2 due to its high expression in epithelial–mesenchymal bidirectional interactions [30]. Epithelial Id2 regulates the specification of intestinal Lgr5+ progenitors in a cell-intrinsic manner [31], and it also contributes to enhancing the proliferation of neural progenitor cells [32,33]. Our research expands the knowledge regarding Id2 by showcasing its role in branching morphogenesis and provides direct evidence of Id2+ epithelial progenitors in the pancreas, kidney, and mammary gland. Notable, not all postnatal branching organs had embryonic Id2+ progeny cells, although Id2 was widely expressed in organs derived from the primitive gut [22].
This phenomenon indicated the heterogeneity of Id2-positive cells depending on their distribution in various organs, whereas the specific mechanism causing the discrepancy between the organs needs to be investigated in depth.
On the contrary, the potential ability of multidifferentiation decreased with the induction time, going downward from E10.5 to E14.5. This change resulted from both intrinsic factors and microenvironment remodeling. The key components of cellular transition still remain unknown. It should be emphasized that Id2 protein expression during the embryonic or postnatal period indicated differences in cell status and function, and we found that proliferative ability only existed in fetal organs. Therefore, Id2 protein expression can indicate progenitor potency only in the embryonic phase in certain organs, such as the lung, pancreas, and kidney.
Identification of adult stem cells for organ repair and generation has been a research target in recent years. Many recent studies have provided evidence of the multipotency and self-renewal capacity of bona fide stem cells. Bronchioalveolar stem cells located in the bronchioalveolar-duct junctions can contribute to lung regeneration after lung injury [34]. Procr+ progenitors found in mouse pancreas can form islet-like organoids in vitro and reverse diabetes in vivo [35].
However, our studies did not confirm that Id2+ embryonic progenitors reside in situ or Id2+ adult stem cells exist during homeostasis. One exception to this rule is the mammary gland, whose branching morphogenesis occurs during adolescence. Prepubescent treatment showed higher labeling efficiency than mid-pregnancy, which indicated that prepubescent-labeled cells had the capacity to expand during postnatal development. The mechanism of Id2+ cell expansion to support lactation in a specific microenvironment requires further study.
In summary, our study advances the model of branching morphogenesis by demonstrating that Id2+ progenitors situated at distal progenitor pools in the pancreas, and kidneys can proliferate and give rise to multiple progeny cells; meanwhile, Id2-marked cells in the prepubescent mammary glands contribute to postnatal structural remodeling. This study will be helpful in the pursuit of identification of adult stem cells needed for organ repair and regeneration. By bringing to light the role of Id2 in branching morphogenesis in multiple organs, it prompts investigative research into ascertaining the similarity and differences in the branching mechanisms of various organs.
Conclusion
In summary, our study investigated the role of Id2+ embryonic progenitor cells in the branching process of the pancreas, kidney, mammary gland, thyroid gland, and salivary gland by using a transgenic mouse model. We discovered that Id2+ marked distal progenitor cells in the embryonic pancreas and kidneys, as well in the mammary glands of adolescent mice. Our study reveals Id2 protein has different self-renewal and proliferation potentials depending on the specific organ, and it can be used as a tool to study the early development of pancreatic and renal progenitor cells.
Data Availability
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Footnotes
Acknowledgments
We also thank Dr. Meiling Wu and Jie Zhang from the Translational Medicine Core Facility of Shandong University for technical assistance.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by grants from the Rongxiang Regenerative Medicine Foundation of Shandong University [2019SDRX-02].
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.