
Abstract
Urine-derived stem cells (USCs) are adult stem cells isolated from urine with strong proliferative ability and differentiation potentials. Cell transplantation of USCs could partly repair liver injury. It has been reported that the proliferative ability of bone mesenchymal stem cells in patients with chronic liver failure is significantly lower than in patients without liver disease. The aim of this study was therefore to evaluate the biological characteristics of USCs from end-stage liver disease patients (LD-USCs, USCs from patients with liver disease) compared with those from normal healthy individuals (N-USCs, USCs from normal individuals), with a view to determining whether autologous USCs can be applied to the treatment of liver disease. In this study USCs were isolated from urine samples of male patients with end-stage liver disease. Adherent USCs exhibit a spindle- or rice grain-like morphology, and express CD24, CD29, CD73, CD90, and CD146 surface markers, but not CD31, CD34, CD45, and CD105. We observed no differences in cell morphology or cell surface marker profile between LD-USCs and N-USCs. LD-USCs exhibited similar proliferative, colony-forming, apoptotic, and migratory abilities to N-USCs. Both USCs demonstrated similar capacities for osteogenic, adipogenic, and chondrogenic differentiation. When USCs were transplanted into CCl4 treatment-induced acute and chronic liver fibrosis mouse models, we observed a decrease in liver index, recovery of alanine aminotransferase and aspartate aminotransferase levels, alleviation of liver tissue injury, and dramatic improvement of liver tissue structure. USC transplantation can effectively recover liver function and improve liver tissue damage in acute or chronic liver injury mouse models. According to the results, we concluded that the biological characteristics of LD-USCs are not affected by basic liver disease. This study provides further evidence of the stem cell characteristics and liver repair function of LD-USCs, which may serve as a theoretical and experimental foundation for autologous USC transplantation technology in the treatment of liver failure and end-stage liver diseases.
Introduction
End-stage liver disease is the most common of the clinical liver disease syndromes, and it is associated with a high mortality [1,2]. Liver transplantation is the only curative treatment for patients with end-stage liver disease [3], but its use is limited by the lack of donors and the high risk of graft rejection [4,5]. One alternative to orthotopic liver transplantation is hepatocellular transplantation, which is an important cell-based therapy that utilizes isolated hepatocytes to compensate for liver function and repair damaged liver tissue [6,7].
Stem cells exhibit strong differentiation potential, self-renewal ability, immunoregulation, and targeted therapeutic function [8]. A number of studies have revealed that several kinds of stem cells can be used in hepatocyte transplantation, including adult hepatocyte stem cells (also known as hepatocyte oval cells or hepatocyte progenitor cells), mesenchymal stem cells (MSCs), and embryonic stem cells [6,7,9 –11]. Indeed, stem cells from different sources are widely used in hepatocyte transplantation to improve liver injury and they show promising results in basic research and clinical applications [12].
However, the stem cells are usually obtained from health donors in an invasive manner, leading to ethical controversy. Although the antigenicity of allogeneic stem cells is relatively low, patients may still be at risk of immune rejection and transplantation-related side effects. Ideally, seed cells should be isolated from the patient's own body, so that they are not rejected by the patient's immune system [13,14].
As the name suggests, urine-derived stem cells (USCs) are derived from urine, and possess stem cell characteristics and multidirectional differentiation potential [15,16]. As they are autologous stem cells that originate from the patient themselves, USCs are not subject to immune rejection after transplantation. Moreover, compared with other stem cell types, such as adipose-derived stem cells (ASCs) or bone marrow-derived mesenchymal stem cells (BM-MSCs), USCs have several advantages including derivation from a convenient source that involves noninvasive sample collection, as well as easy isolation and cell culture. These features simplify the preparation and use of autologous stem cells, which makes USCs one of the most promising alternative seed cell types for stem cell-based therapeutic strategies [17,18].
Studies have suggested that USCs are derived from kidney renal parenchyma, and it is difficult to obtain healthy USCs from patients with kidney disease whose renal tissue structure is damaged [19,20]. Theoretically, patients with chronic liver disease but normal renal tissue structure and function should produce unaffected USCs capable of normal stem cell functions, such as proliferation and differentiation. However, there is evidence that the in vitro proliferative ability of BM-MSCs is significantly lower in patients with chronic liver failure compared with patients without liver disease [21]. Therefore, before autologous USCs can be used in the treatment of liver disease, it is first necessary to determine whether the biological characteristics of USCs are affected in patients with basic liver disease.
In this study, we compared the in vitro biological characteristics and in vivo liver repair function of USCs derived from normal individuals (N-USCs) with USCs derived from patients with liver disease (LD-USCs), to determine whether the biological characteristics of LD-USCs were affected by liver disease. This study provides further evidence of the stem cell characteristics and liver repair function of LD-USCs, which may serve as a theoretical and experimental foundation for autologous USC transplantation technology in the treatment of liver failure and end-stage liver diseases.
Materials and Methods
Isolation and culture of USCs
USCs were isolated from urine samples of 20 male patients with end-stage liver disease in the affiliated hospital of Chongqing medical university (men, ranging from 32 to 78 years old, average age = 55.6 ± 14.5) and 20 normal health person (men, ranging from 24 to 64 years old, an average age = 50.5 ± 12.7). All subjects involved in this study gave their informed consent for inclusion. This study was conducted under the approval of Institutional Review Board of Chongqing Medical University.
Urine samples were centrifuged at 1,500 rpm for 5 min at room temperature. The cell pellets were washed by phosphate-buffered saline (PBS) (Solarbio, Beijing, China), and resuspended with 24 mL of medium specific for USCs culture with 10% fetal bovine serum (FBS; Hyclone, Logan, Utah), 100 U/mL penicillin, 100 μg/mL streptomycin, and then plated in 24-well plates at 37°C and 5% CO2. USCs in urine sample were separated into single cell, cell colony were counted within 7 days after first colony formation. Colony numbers per 200 mL were calculated. After 10–14 days, total cells in a 24-well plate were trypsinized and passaged to 60 mm dishes, and then 100 mm dishes.
Cells at passages 3–5 were used, in each experiment, at least 3 USCs from patient with liver disease (LD-USCs) and 3 USCs from normal individuals (N-USCs) were randomly selected from 20 patients and 20 normal persons, and each USCs samples was confirmed to be used in at least one experiment.
Flow cytometry analysis
Cells were trypsinized and washed by PBS twice, followed by resuspension at a concentration of 1 × 106 cells/mL in PBS. Cells suspensions were incubated in the dark at 4°C with FITC or PE-labeled antibodies (murine anti-human CD24-FITC, CD29-PE, CD31-FITC, CD34-PE, CD45-FITC, CD73-PE, CD90-PE, CD105-FITC, CD146-PE; Becton, Dickinson and Company) [22,23]. After 30 min of incubation, cell suspensions were washed twice and resuspended in 300 μL PBS for flow cytometry.
Cell proliferation potential
Cells were separately plated on 60 mm dishes at a density of 1 × 106 cells starting at passage 1. Cells were trypsinized and counted with a hemocytometer every 3 days until passage 10. Following each passage, cells were replanted at the same density for further culture. Population doublings (PD) were calculated using the formula: PD = log2 (Nh/Ni). The population doubling time (Dt) was calculated with the formula: Dt = Ct/log2 (Nh/Ni), where Nh is cell harvest number, N0 is the inoculun cell number, and Ct is the culture time.
Colony formation assay
Cells were planted in 12-well plates in duplicate at 200 cells per well and cultured for ∼10 days. Cells were fixed with 4% paraformaldehyde then stained with 0.1% crystal violet. Cell colonies containing >50 cells were counted under an inverted microscope (Nikon ECLIPSE Ti, Japan). The plate clone formation efficiency was calculated as follows: (numbers of colonies/numbers of seeded cells) × 100%.
Cell migration ability
Wound healing assay was performed to detect parallel cell migration ability. Cells were plated in six-well plates and reached 100% confluence. A scratch wound in the surface of confluent monolayers was created by a pipette tip, suspend cells were removed by washing with PBS. Bright-field images of the same wound field were captured at indicated time points.
Transwell assay was performed to detect vertical cell migration ability. Cells were resuspended with serum-free culture medium and then 200 μL cell suspension (3 × 104 per well) was seeded into the upper Transwell chambers (8.0 μm; Corning, New York). Medium containing 10% FBS was added into the lower chambers. After 48 hours, cells in the upper chamber were carefully removed, and the remaining cells that migrated and invaded to the lower surface were fixed in 4% paraformaldehyde for 30 min at room temperature and washed in PBS, then stained with 0.1% crystal violet (Beyotime, Shanghai, China) for 20 min at room temperature. Five independent fields of view were randomly quantified under a microscope (Nikon, Tokyo, Japan). The stain was dissolved in absolute ethanol and the absorbance value was measured at 570 nm with a microplate reader (Thermo Scientific, MA).
Multiple differentiation potential
Cells were plated in 24-well plates at a density of 1 × 105/well, and respectively cultured in appropriate induction medium (Cyagen Biosciences, Inc., Guangzhou, China) to evaluate multiple differentiation potential along osteogenic, adipogenic, and chondrogenic lineages. Induction medium was changed every 3 days. Early osteogenic differentiation was detected by alkaline phosphatase (ALP) staining (Solarbio) after 7 days of induction, adipogenic and late osteogenic differentiation were detected by Oil red O staining (Beijing Reagan Biotechnology Co., Ltd.) and Alizarin red staining (Solarbio), respectively, after 14 days of induction, and chondrogenic differentiation was stimulated by using Alcian blue staining (Beijing Reagan Biotechnology Co., Ltd.). Each staining assay was carried out according to the manufacturer's instruction.
Animal model and cell transplantation
All procedures involving animals were reviewed and approved by the Animal Care and Use Committee of Chongqing Medical University. Nude mice [8 weeks old, male, 20–26 g, qualified number: SCXK (Beijing) 2014-0004] were purchased from Tengxin Institute of Biotechnology (Chongqing, China). All mice were randomly divided into acute liver injury group (n = 42) and chronic liver injury group (n = 24). Owing to the short period of modeling and strong liver compensation ability, the liver of acute animal model showed a certain degree of repair. To detect the change of liver index, aspartate aminotransferase (AST) and alanine aminotransferase (ALT) level, and liver structure, two time points D3 and D7 were set up in acute animal model. There were 12 animals in each CCl4-treated acute model and 6 animals in each CCl4-treated chronic model. Total 42 mice in acute liver injury were randomly divided into control group (n = 6), CCl4 model group (n = 12), N-USCs transplanted group (n = 12), and LD-USCs transplanted group (n = 12). Total 24 mice in chronic liver fibrosis experiments were randomly divided into control group (n = 6), CCl4 model group (n = 6), N-USCs transplanted group (n = 6), and LD-USCs transplanted group (n = 6).
CCl4 (Aladdin Bio-Chem Technology Co., Ltd., Shanghai, China) reagent were freshly prepared and diluted to 10% (vol/vol) by olive oil. To induce acute liver failure mice model, nude mice were intraperitoneally injected with 10% CCl4 at the dose of 20 mL/kg body weight, and mice in control group were intraperitoneally injected an equal volume of olive oil [24]. After CCl4 treatment for 24 and 72 h, 2 × 106 USCs suspended in 0.2 mL of PBS were transplanted through caudal vein, and the CCl4 group were administrated with an equal amount of PBS.
To induce chronic liver fibrosis, nude mice were intraperitoneally injected with 10% CCl4 at the dose of 10 mL/kg body weight twice a week for 8 weeks, and mice in control group were intraperitoneally injected an equal volume of olive oil at the same time. Our previous study found that CCl4 treatment for 8 weeks can successfully obtain chronic liver injury model with stable and serious liver fibrosis. To improve the implantation rate of USCs transplantation and repair liver injury, USCs were treated twice a week for 2 weeks [25]. After CCl4 treatment for 8 weeks, 2 × 106 USCs suspended in 0.2 mL of PBS were injected through caudal vein twice a week for 2 weeks, and the CCl4 group were administrated with an equal amount of PBS.
Assessment of liver function and histopathology
Mice were killed at the day 1 (D1), D3, and D7 after the treatment of CCl4 in acute liver injury mice model, and were killed 3 days later after the last USCs transplantation in chronic liver fibrosis mice model. The serum levels of ALT, AST samples were measured. Liver index was calculated as follows: Liver index = liver wet weight (g)/body weight (g) × 100%. Liver tissues were fixed in 4% paraformaldehyde, embedded in paraffin. The sections were harvested at 4 μm and detected using hematoxylin and eosin (H&E) staining (Beijing Reagan Biotechnology Co., Ltd.) and Masson staining (Beijing Reagan Biotechnology Co., Ltd.). The expression of alpha-smooth muscle actin (α-SMA) (1:800; Bioss, Beijing, China), myeloperoxidase (MPO) (1:200; Boster, China), and 8-hydroxy-2′-deoxyguanosine (8-OHdG) (1:400; Bioss) were detected by immunohistochemistry.
Statistical analysis
All data were quantitative variables. As the sample size was small, a Shapiro–Wilk test was used to evaluate the normal distribution of the data. Student's t-test was used for the comparisons of normally distributed variables between two groups and one-way analysis of variance and a Student-Newman-Keul's post-test were used to measure significant differences among more than two groups. If the data were not a normal distribution (Figs. 1A, 3A, 4B, and 7A), Wilcoxon signed-rank test was conducted as a nonparametric statistical test. P value <0.05 was considered as statistically significant.

N-USCs and LD-USCs exhibit similar morphologies and characteristics.
Results
Morphology and characteristics of LD-USCs and N-USCs
Primary USCs began to adhere ∼3 days after seeding, and cell colonies were observed after 7 days in culture. Although the cell clones were related to the time and temperature of urine placement in vitro, cell clones were obtained from every urine sample. The primary cell clones per 200 mL urine in the patient group (23.96 ± 12.3) was higher than that of the healthy group (21.2 ± 5.2), but no statistical differences were found between N-USCs and LD-USCs (P>0.05; Fig. 1A). Adherent USCs exhibited a spindle-like or rice grain-like morphology, and the cells reached a confluency of 50%–60% within 10–14 days of culture (Fig. 1B). With consistent passaging, the cells grew quickly and expanded to 80%–90% confluency within 3–4 days, and there was a progressive increase in the proportion of spindle-like cells and a corresponding decrease in the number of rice grain-like cells. The growth rate of USCs in vitro diminished significantly after passage 10, and most flat cells lost their proliferative ability (Fig. 1C). There was no difference in cell morphology or the ratio of the two cell types in samples from the healthy compared with patient group (10 samples).
Flow cytometry analysis results indicated that the USCs expressed MSCs surface markers, including CD24, CD29, CD73, and CD90, and the endothelial cell surface marker CD146. The hematopoietic cell surface markers CD31, CD34, CD45, and the endothelial cell marker CD105 were expressed at low levels. No significant difference in the expression levels of the surface markers was observed between the N-USC and LD-USC groups (P>0.05; Fig. 2). These data indicated that we had successfully isolated and cultured USCs from the urine of patients with end-stage liver disease, and that these cells exhibited cell morphologies and stem cell marker characteristics that were similar to those of the N-USCs.

Flow cytometric analysis of cell surface markers. Quantitative analysis and comparison of cell surface marker expression levels in N-USCs and LD-USCs. Data are given as mean ± standard deviation of three independent experiments. P > 0.05. Color images are available online.
Proliferative and migratory abilities of N-USCs and LD-USCs
During 10 generations of consecutive culture, the Dt of each passage showed no significant difference between the N-USC and LD-USC groups (P>0.05; Fig. 3A). The population doubling (PD) time of N-USCs and LD-USCs at P10 was 25.65 ± 2.33 and 26.93 ± 1.96 h, respectively.

N-USCs and LD-USCs exhibit similar proliferative abilities and apoptosis rates.
The clone formation rate reflects two important cellular traits, namely cell population dependence and proliferative ability. The colony formation abilities of N-USCs (9.92% ± 2.27%) and LD-USCs (9.13% ± 2.88%) were not significantly different (P>0.05; Fig. 3B). As is typical of stem cells, the baseline level of apoptosis among the USCs was relatively low, and no significant difference in apoptosis rate was found between N-USCs and LD-USCs (P>0.05; Fig. 3C). The wound healing rate and the number of cells that migrated through a transwell membrane were also not significantly different in LD-USCs compared with N-USCs (P>0.05; Fig. 4A, B).

N-USCs and LD-USCs possess similar migratory abilities.
The above results revealed that LD-USCs had similar cell proliferative, colony forming, apoptotic, and migratory abilities to N-USCs, indicating that the self-renewal capabilities of USCs were not affected by chronic liver disease in the donor.
Multiple differentiation potential of N-USCs and LD-USCs
Multipotential differentiation is an important feature of stem cells. We therefore examined the osteogenic, chondrogenic, and adipogenic differentiation potential of USCs grown in different induction media, using specific stains to determine functional activity (Fig. 5). After 7 days of osteogenic induction, 30%–40% of the cells were positive for ALP staining, displaying purple-blue staining in the cytoplasm, which is an early indicator of osteogenic differentiation. After 14 days of osteogenic induction, some of the cells (∼20%) stained positive for Alizarin Red S. Calcium nodules displayed red staining, which is a feature of late osteogenic differentiation and calcium salt deposition by USCs. After 14 days of chondrogenic induction, almost all the cells stained positive for Alcian Blue, in both the cytoplasm and cell membrane. After 14 days of adipogenic induction, ∼45%–50% of cells exhibited vacuolar, lipid deposition in the cytoplasm, Oil Red O staining, and visible lipid droplets in many cells. Therefore, these data suggest that USCs have great potential to differentiate into osteoblasts, chondrocytes, and adipocytes. Here too, there was no significant difference in the differentiation capacity of the N-USCs compared with the LD-USCs.

N-USCs and LD-USCs have similar capacities for multipotential differentiation. USCs seeded in 24-well plates were exposed to osteogenic, chondrogenic, or adipogenic differentiation media. ALP, scale bar = 25 μm; Alizarin Red, scale bar = 100 μm; Oil Red O, scale bar = 100 μm; and Alcian Blue, scale bar = 25 μm. ALP, alkaline phosphatase. Color images are available online.
N-USCs and LD-USCs exhibit similar repair efficiency in a mouse model of acute liver injury
Intraperitoneal injection of 10% CCl4 at the dose of 20 mL/kg body weight was performed once to induce acute liver injury in mice of model group. N-USCs and LD-USCs were transplanted in vivo through the tail vein after CCl4 treatment for 24 and 72 h, respectively. Compared with the control group, the liver index and serum levels of ALT and AST were significantly increased in mice of model group (P < 0.05; Fig. 6A). The results from H&E staining showed regularly arranged cells and a normal hepatic structure in liver tissues from the control group (Fig. 6B). However, on D1 after CCl4 treatment, liver tissues from the mice of model group exhibited enlarged hepatocytes, vacuolar or ballooning degeneration, loose and transparent cytoplasm, and even disappearance of the nuclear envelope and the nucleus itself. On D3 after CCl4 treatment, the serum levels of ALT/AST and tissue structure exhibited obvious signs of recovery, and the hepatocyte swelling was reduced, although the vacuolar degeneration was still evident (Fig. 6A, B). Compared with the model group, the serum levels of ALT/AST in mice of USC-transplanted groups were significantly lower (P < 0.05; Fig. 6A), with obvious recovery of the tissue structure, and an increase in the nucleus/cytoplasm ratio. There was no difference in the liver index between the mice in model group and USC-transplanted groups (P > 0.05; Fig. 6A). On D7 after CCl4 treatment, N-USCs or LD-USCs exhibiting significant decrease in the liver index with mice transplanted twice. ALT/AST serum levels recovered to the normal range, and no differences were found among the three groups at D7 (Fig. 6A, P > 0.05). There was an evident recovery of the hepatic tissue structure, although with slightly swollen hepatocytes. The tissue structure and arrangement of hepatocytes in the USC-transplanted groups were regular without obvious abnormalities (Fig. 6B). There were no statistical differences in liver index, ALT/AST, or pathological changes between the N-USC- and LD-USC-transplanted groups (P > 0.05; Fig. 6B).
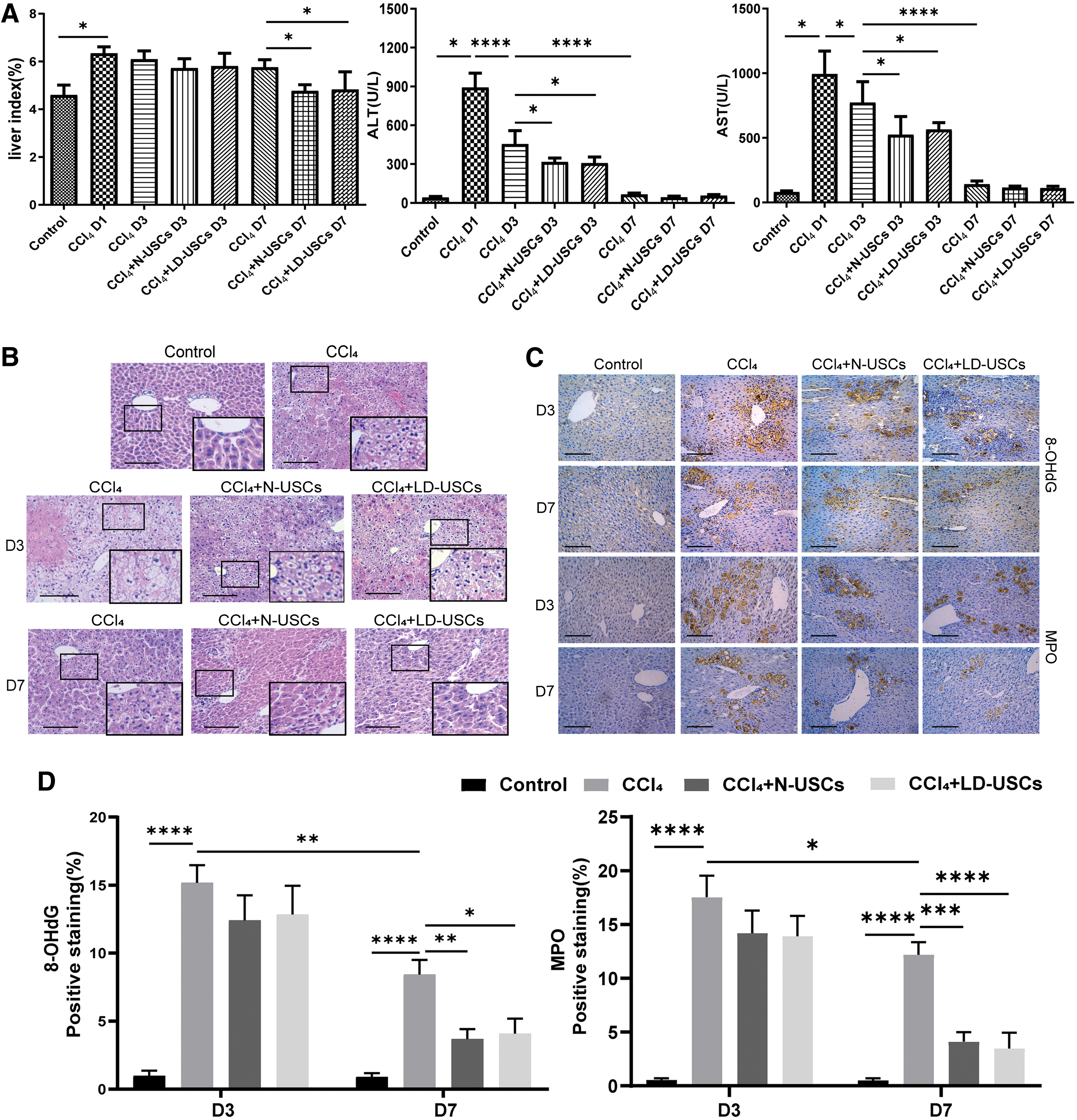
N-USCs and LD-USCs exhibit similar repair efficiency in a mouse model of acute liver injury.
The immunohistochemical analyses showed that the expression levels of the oxidative stress-related genes, MPO and 8-OHdG, were low in normal liver tissue, but significantly increased in the model group (P < 0.05; Fig. 6C, D). On D3 after CCl4 treatment, the expression levels of MPO and 8-OHdG were markedly reduced, but no difference was observed between the model and the USC-transplanted groups (P > 0.05; Fig. 6D). On D7 after CCl4 treatment, the expression levels of MPO and 8-OHdG decreased further, but were higher in tissues from the mice in model group than those from mice of USC-transplanted groups (P < 0.05; Fig. 6D), indicating that liver function and tissue can self-repair during acute liver injury. USC transplantation in our model of acute liver disease was able to decrease the liver index, alleviate liver tissue injury, and promote liver recovery. LD-USCs exhibited a recovery efficiency that was similar to that of the N-USCs.
N-USCs and LD-USCs exhibit similar recovery efficiency in a mouse model of chronic liver fibrosis
Intraperitoneal injection of 10% CCl4 was performed for 8 weeks to induce chronic liver fibrosis in nude mice of model group. N-USCs and LD-USCs were transplanted in vivo through the tail vein twice a week for 2 weeks. The liver index and serum levels of ALT/AST in mice in the model group were significantly higher than those in the control group (P < 0.05; Fig. 7A). USC transplantation reduced the liver index and the level of ALT slightly, but no statistical difference was found between the three groups (P > 0.05; Fig. 7A). The level of AST in the USC-transplanted groups were lower than that in the model group (P < 0.05; Fig. 7A), but there were no statistically significant differences between the N-USC and LD-USC groups (P > 0.05; Fig. 7A). The liver tissue structure of control nude mice was normal, and the hepatic lobules were clear and intact (Fig. 7B). After 8 weeks of CCl4 treatment, the tissue structure was disordered and there was evidence of hepatocyte degeneration, nuclear pyknosis or disappearance, island pseudolobules, and infiltration of inflammatory cells in the portal area. The results from Masson staining showed blue-stained fibrous tissue around the pseudolobules, extending from the central vein to the edge of the hepatocyte space (Fig. 7B). After USC transplantation for 2 weeks, the liver tissue structure was partially restored, no obvious pseudohepatic lobules were observed, and the blue-stained fibrotic area in the liver tissue had decreased (P < 0.05; Fig. 7D). There was no significant difference in H&E and Masson staining between the N-USC and LD-USC groups (P > 0.05; Fig. 7D). As given in Fig. 7C, the expression levels of the fibrosis marker, smooth muscle actin (α-SMA), and the oxidative stress-related genes, MPO and 8-OHdG (all of which are expressed at very low levels in normal liver tissue), demonstrated obvious increases in the model group to control group (P < 0.05; Fig. 7D). With USC transplantation, the expression levels of α-SMA, MPO, and 8-OHdG were markedly reduced, but no differences were found between the N-USC and LD-USC groups (P > 0.05; Fig. 7D). These results indicated that USCs transplanted through the tail vein could partly colonize in liver tissue and were able to achieve some remediation of liver function in mice with chronic liver failure (Supplementary Fig. S1). In particular, USC transplantation produced a dramatic amelioration in the structure and fibrosis of the damaged liver tissue. Of importance, there was no difference in repair capability between the N-USC and LD-USC groups.

N-USCs and LD-USCs exhibit similar repair efficiency in a mouse model of chronic liver fibrosis.
Discussion
Stem cells have a capacity for self-renewal and the potential to differentiate into more mature, specialized cells, and they have demonstrated great potential in the treatment of a variety of tissue damage phenotypes, tumors, and end-stage diseases [8,26,27]. Adult stem cells are undifferentiated cells that exist in niche within differentiated tissue, and which are able to self-renew and differentiate into a certain range of cell types. Compared with embryonic stem cells, adult stem cells are safer and are easily obtained without ethical controversy. Autologous adult stem cells also have the advantage of bypassing immune rejection following transplantation, realizing the promise of individualized treatment in clinical applications [28,29]. Many types of adult multipotent stem cells have been isolated from different tissues and identified, such as hematopoietic stem cells, bone marrow MSCs, neural stem cells, liver stem cells, skin epidermal stem cells, ASCs, retinal stem cells, and pancreatic stem cells [30,31].
It has been a topic of some debate as to whether stem cells exist in urine. In 2008, Professor Zhang at the Wake Forest Institute for Regenerative Medicine first reported the presence of a small number of highly proliferative cells in urine that possessed certain mesenchymal characteristics, and which he named USCs [15]. Compared with BM-MSCs, USCs are easier to obtain by noninvasive and low-cost methods, have higher telomerase activity, and their capacity for self-renewal and cell proliferation is much better, allowing them to maintain a consistent population doubling rate [32,33]. From pooled urine samples collected over 24 h, it is possible to obtain ∼140 USC clones. After several generations of culture, ∼5.3 × 1010 cells (at P5) can be produced, which is sufficient for clinical applications. Even after ∼20 successive cell generations, USCs do not exhibit chromosomal aberrations or tumor formation in vivo [15,34]. In addition, USCs can secrete nutritional and angiogenic factors, as well as cytokines, act as tumor suppressors, and confer immunomodulatory effects that inhibit the proliferation of peripheral blood T cells, B cells, and monocytes. Together, these characteristics indicate that USCs are a new type of fully functional stem cell [35,36]. In our previous study, we demonstrated that USCs have a cell phenotype, self-renewal potential, and multilineage differentiation capacity that is similar to that of MSCs [37]. USC transplantation can effectively improve liver function and liver tissue damage in acute or chronic liver injury mouse models, indicating that USCs are an ideal and reliable cell source for hepatic cell transplantation and liver tissue engineering [25]. Autologous USCs originating from patients with liver diseases are exposed to pathophysiological conditions that could alter their function; therefore, we considered it necessary to evaluate the safety, feasibility, and functional capacity of LD-USCs before their clinical application in the treatment of liver disease.
The patients recruited for this study had end-stage chronic hepatitis B, liver cirrhosis, gastrointestinal hemorrhage, hepatoblastoma, or hepatocellular carcinoma. Based on the primary data from blood serum tests for ALT, AST, thiobarbituric acid, total bilirubin, and albumin levels, the hypohepatia index of blood tests of patients was evaluated as severe liver function damage [38 –40]. USCs are thought to originate from kidney glomerular parietal cells; therefore, patients with abnormal kidney function were excluded. Stem cells in urine must have sufficient proliferative activity to adhere to the plate and eventually form colonies, and the presence of cell colonies thus reflects the proliferative ability of primary USCs [34,41,42]. The cell colony number per 200 mL urine was not significantly different between the N-USC and LD-USC groups, but there were two discrete samples of interest in the LD-USC group, which came from two patients with liver cirrhosis. As a result of abdominal dropsy, the volume of collected urine from the patients with liver cirrhosis was much less in comparison with the other patients [43], and the colony number data were therefore high after normalization by urine volume. Biliary atresia mostly occurs in infants within 1 year of age [44]. Although it can be difficult to collect urine samples from patients this young, we obtained 48 cell colonies from 20 mL urine samples of 5-month-old patients with biliary atresia; the colony number was therefore 480, and these data had to be excluded as an outlier. Therefore, biliary atresia was not included in this study. The population doubling (PD) level refers to the total number of times the cells in the population have doubled since their primary isolation in vitro. The Dt is the period of time required for cells to double the population, which is a standard measure applied to accurately assess cell growth ability [45,46]. Over the course of 10 passages, there were no significant differences in PD and Dt between the two groups. Based on the calculation method, the values for cell colony number are affected by the urine volume; nevertheless, we have demonstrated that the proliferative ability of USCs was not affected by end-stage liver diseases.
Migration is a very important characteristic of stem cells, which can be affected by variant signals at the lesion site; transplanted stem cells have an inherent tropism toward injured tissue, where they then differentiate into specific cells or affect the microenvironment through autocrine or paracrine signaling [47 –49]. As multipotent stem cells, USCs have a high capacity for differentiation into osteogenic, chondrogenic, and adipogenic cell lineages [50,51]. We have previously demonstrated that USCs can also differentiate into hepatocyte-like cells capable of liver function [25]. In this study, compared with N-USCs, LD-USCs exhibited similar capacities for proliferation, migration, and differentiation, highlighting their potential for use as autoallergic stem cells in clinical applications such as liver regeneration.
In the second part of our study, we established acute and chronic liver injury mouse models to compare the repair efficiency of N-USCs and LD-USCs after transplantation through the tail vein. An acute liver injury model produced by a single CCl4 treatment resulted in a high liver index, increased levels of serum AST and ALT, and hepatocyte degeneration and necrosis [52,53]. Thanks to the strong self-healing ability of the mouse liver, in the absence of continuous damage, mice in the acute liver injury model exhibited recovery of the liver index and liver structure to near normal levels after 10 days [54]. The experimental period was therefore shortened to 7 days. Nevertheless, USCs that were successfully implanted in the liver of acute injury mice exhibited significant improvement in liver function and structure, especially with two USC transplants. No significant differences were found between the two USC-treated groups. To establish a chronic liver injury model, CCl4 was administered for 8 weeks, after which the structural disorder of the liver tissue, fibrosis, and the formation of false lobules were irreversible even when CCl4 was withdrawn [39,55,56]. This CCl4-induced chronic liver fibrosis model was stable and served as a valuable tool to evaluate the repair efficiency of USC transplantation. Owing to the powerful compensatory function of the liver, the liver index and the levels of serum ALT and AST in the chronic injury mice increased slightly compared with those in the normal group, but the numerical values were much lower than those from mice in the acute injury group. Therefore, no statistical difference in liver function was observed between the three groups. However, after four USC transplantations within 2 weeks, we observed an obvious reduction in blue-stained fibers in the liver tissue, false lobules, and liver tissue structure disorder, and the expression levels of the fibrosis marker, α-SMA, and the oxidative stress-related genes, MPO and 8-OHdG [57 –59], decreased remarkably. There was no difference between the N-USC and LD-USCs transplanted groups. The above data suggest that USC transplantation can partially repair the pathological changes resulting from chronic liver injury induced by CCl4. USCs isolated from patients with liver disease did not exhibit any loss of their repair function in vivo. One limitation of this study is that female patients were not included because of the risk of sample contamination and the difficulty of USC isolation from female urine samples. A relative sterile collection process and urine sample treatment as quickly as possible after in vitro might solve this problem. Another is the relatively small sample size, which should be expanded for further evaluation, including young patients with end-stage liver diseases, such as biliary atresia cirrhosis, hepatoblastoma, and viral hepatitis. It is also worth noting that the mechanism of USC transplantation in the treatment of liver injury remains unclear. Transdifferentiation may be an acceptable mechanism of stem cells in the treatment of liver diseases. This differentiation ability comes from the multiple-differentiation potential of stem cells. Stem cells with low immunogenicity can also inhibit the proliferation and maturation of immune cells. Stem cells can migrate to the injured liver under the action of chemokines and play a role as the paracrine control to secrete growth factors or anti apoptotic cytokines to promote hepatocyte proliferation. In addition, stem cells can regulate the phenotypic transformation of Kupffer cells to improve liver injury and cirrhosis [60 –64]. However, the specific mechanisms underlying the curative effect still require further investigation.
Conclusions
USC transplantation can effectively recover liver function and improve liver tissue damage in acute or chronic liver injury mouse models. In this study, we further demonstrated that there are no differences between LD-USCs and N-USCs in biological characteristics in vitro and repair capacity in vivo. Taken together, autologous USCs obtained by a noninvasive and safe method may be used as an ideal and reliable seed cell source for hepatocellular transplantation, and show good application prospect in the clinical treatment of the end-stage liver diseases.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by research grants from the Chongqing Science and Technology Commission (No. csct2018jscx-msybX0084 to Y.B.).
Supplementary Material
Supplementary Figure S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.