
Abstract
SARS-CoV-2 infection during pregnancy has been associated with poor maternal and neonatal outcomes and placental defects. The placenta, which acts as a physical and immunological barrier at the maternal–fetal interface, is not established until the end of the first trimester. Therefore, localized viral infection of the trophoblast compartment early in gestation could trigger an inflammatory response resulting in altered placental function and consequent suboptimal conditions for fetal growth and development. In this study, we investigated the effect of SARS-CoV-2 infection in early gestation placentae using placenta-derived human trophoblast stem cells (TSCs), a novel in vitro model, and their extravillous trophoblast (EVT) and syncytiotrophoblast (STB) derivatives. SARS-CoV-2 was able to productively replicate in TSC-derived STB and EVT, but not undifferentiated TSCs, which is consistent with the expression of SARS-CoV-2 entry host factors, ACE2 (angiotensin-converting enzyme 2) and TMPRSS2 (transmembrane cellular serine protease) in these cells. In addition, both TSC-derived EVT and STB infected with SARS-CoV-2 elicited an interferon-mediated innate immune response. Combined, these results suggest that placenta-derived TSCs are a robust in vitro model to investigate the effect of SARS-CoV-2 infection in the trophoblast compartment of the early placenta and that SARS-CoV-2 infection in early gestation activates the innate immune response and inflammation pathways. Therefore, placental development could be adversely affected by early SARS-CoV-2 infection by directly infecting the developing differentiated trophoblast compartment, posing a higher risk for poor pregnancy outcomes.
Introduction
During the COVID-19
Moreover, the maternal–fetal interface of women infected in the third trimester showed an associated strong immune response, with activation of natural killer and T cells, and increased expression of interferon (IFN)-related genes [14,15]. Strong systemic inflammatory responses in SARS-CoV-2-infected patients have been observed long after the initial phase of infection [16,17], potentially contributing to placental dysfunction in pregnancies past the active infection phase. Inflammatory lesions in placental villi, or chronic villitis, have been associated with placental dysfunction, particularly when severe (associated with perivillous fibrin deposition and fetal vascular obliterative lesions), leading to adverse effects on fetal development including fetal growth restriction and stillbirth [18].
Across gestation, the placenta performs multiple functions to sustain embryo and fetal development, which include forming a physical and immunological barrier at the maternal–fetal interface against potential pathogens infecting the mother. However, this barrier is not established until the end of the first trimester, and localized viral infection of the trophoblast compartment early in gestation can result in altered placental function and consequently suboptimal conditions for fetal growth and development, even in the absence of vertical transmission [19 –21].
Evidence indicates that infections with respiratory (ie, influenza virus) and other viruses [ie, cytomegalovirus (CMV), adeno-associated virus], considered to have little or no fetal pathogenicity, can impair trophoblast function and potentially contribute to pregnancy loss and other pregnancy complications, including preterm birth and preeclampsia [22 –24]. Therefore, evaluating the ability of SARS-CoV-2 to infect the trophoblast compartment of the early placenta and the consequent effects are of fundamental importance to understand the mechanisms behind poor pregnancy outcomes following viral infection.
The entry of SARS-CoV-2 into human cells occurs via the angiotensin-converting enzyme 2 (ACE2) receptor and assisted by the host transmembrane cellular serine protease (TMPRSS2) [25 –27]. ACE2 and TMPRSS2 are expressed at high levels in the trophoblast compartment of first trimester placentae and decrease as gestation progresses [14,28,29]. However, the effects of viral presence in the placenta during the early stages of development are difficult to study in ongoing pregnancies. It is currently unknown if the virus can actively replicate in the trophoblast compartment or if it can elicit an inflammatory response in the placenta, with potential adverse pregnancy outcomes.
Recently, human trophoblast stem cells (TSCs) have been derived from first trimester placental tissues [30]. These cells express typical cytotrophoblast (CTB) progenitor markers, including ITGA6 and EGFR, and can be differentiated in vitro into both invasive HLA-G+ extravillous trophoblast (EVT) and multinucleated hCG-secreting syncytiotrophoblast (STB), recapitulating early placental developmental processes. Thus, TSCs are a powerful model to investigate key mechanisms of early human placentation, including viral infection/replication in the trophoblast compartment.
In this study, we have investigated the effect of SARS-CoV-2 infection in early gestation placentae by using human TSCs and their EVT and STB derivatives to elucidate the potential consequences of viral infection on early placental development.
Materials and Methods
SARS-CoV-2 infection of undifferentiated and differentiated TSCs
Two undifferentiated TSC lines (1048 and 1049, both female) and TSC-derived EVT (d5) and STB (d6) were infected with SARS-CoV-2 USA-WA1/2020 strain, obtained from BEI Resources (NR-52281) at multiplicity of infection (MOI) = 1. After 1 h incubation at room temperature, the viral inoculum was removed, and cells were washed twice with phosphate-buffered saline (PBS) to remove unbound virus, followed by the addition of fresh TSC, EVT, or STB media, containing either vehicle (dimethyl sulfoxide) or 500 nM of remdesivir (considered time = 0 h). Cell lysates and culture supernatants were collected at 0, 2, 10, 24, and 48 h postinfection (h p.i.). All infections with SARS-CoV-2 were performed in a Biosafety Level 3 laboratory under the approval of the Sanford Burnham Prebys Medical Discovery Institute Biosafety Committee.
Plaque-forming assay
Supernatants from trophoblast cells infected with SARS-CoV-2 cells were collected at 0, 12, 24, and 48 h p.i. and stored at −80°C until used. Vero E6 cells (ATCC CRL-1586) were seeded in 12-well plates (6 × 105 cells per well) using the following cell media: Dulbecco's modified Eagle medium (Gibco) supplemented with 10% heat-inactivated fetal bovine serum (Gibco), 50 U/mL penicillin, 50 μg/mL streptomycin (both from Fisher Scientific), 1 mM sodium pyruvate (Gibco), 10 mM 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid (HEPES; Gibco), and 1 × minimum essential media (MEM) nonessential amino acids solution (Gibco). Vero E6 cells were then incubated overnight under 37°C and 5% CO2 conditions.
Confluent Vero E6 cells were washed once with 1 × PBS and infected with 100 μL of the collected TSC, STB, and EVT cell supernatants that were serially diluted 1:10. Plates were incubated 1 h at room temperature, followed by inoculum removal and addition of 1 mL overlay media [2 × MEM and 2.5% Avicel (RC-591 NF; FMC BioPolymer) at 1:1 ratio]. 2 × MEM contains 100 mL of 10 × MEM (Gibco), 10 mL of 100 × penicillin–streptomycin (Fisher Scientific), 10 mL of 100 ×
Bulk RNA-sequencing and analysis
Total RNA was quantified using Qubit Fluorometer (Thermo Fisher Scientific) and quality controlled on an Agilent TapeStation (Agilent, Santa Clara). RNA-seq libraries were prepared using the Ribo-Zero Plus rRNA Depletion kit (Illumina), followed by TruSeq Stranded Total RNA kit (Illumina) at the IGM Genomics Center at the University of California San Diego. Libraries were pooled and sequenced on NovaSeq 6000 S1 Flow Cell (Illumina) to an average depth of 23 million uniquely mapped reads. Quality control was performed using FastQC (v.0.11.8) and multiQC (v.1.11). Reads were mapped to GRCh38.p10 (GENCODE release 26) using STAR (v.2.7.3a; [31]) and annotated using featureCounts (subread v.1.6.3, GENCODE release 26 primary assembly annotation; [32]).
The STAR parameters used were as follows: –runMode alignReads –outSAMmode Full –outSAMattributes Standard –genomeLoad LoadAndKeep –clip3pAdapterSeq AGATCGGAAGAGC –clip3pAdapterMMp 1. The featureCounts parameters were as follows: -s 2 -p -t exon -T 13 -g gene_id. Ensembl genes without at least 2 samples with 10 or more reads were removed from analysis. Ensembl gene IDs were converted to HGNC gene symbols using BiomaRt (v. 2.42.1). Normalization was performed using the R (v.3.6.3) package DESeq2 (v.1.28.1; [33]). Qlucore Omics Explorer software version 3.1 (Qlucore AB, Lund, Sweden) was used to analyze genes expression data sets and generate principal component analysis (PCA) plots and heat maps with a defined variance of 0.0031 and two or multiple group comparisons. Genes with an adjusted P value of <0.05 and log2-fold change of >1 were considered differentially expressed. Differentially expressed genes were then subjected to Gene Ontology analysis using the Metascape web application [34].
Statistical analyses
All statistical tests were performed using GraphPad Prism® software (GraphPad software, CA). Tests described at the end of each method.
Data sharing
All RNA-seq data sets used in this study are available in the gene expression omnibus database under GSE216484.
Study approval
All work with human placental tissues was carried out under a protocol approved by the Human Research Protections Program Committee of the University of California San Diego Institutional Review Board.
Results
Expression of ACE2 and TMPRSS2 in human TSCs and its derivatives
To study the effect of SARS-CoV-2 infection on trophoblast cells during early development, we used TSCs derived from two different 6-week placental tissues (#1048 and #1049) and that have been recently characterized [35]. These cells recapitulate early trophoblast development and can be maintained undifferentiated in TSC media or differentiated into either STB or EVT, following previously published protocols [30]. Both lines were differentiated into STB and EVT, and differentiation was confirmed by cell type-specific marker expression, specifically downregulation of TSC markers TP63 and ITGA6 (Supplementary Fig. S1A) and upregulation of ASCL2 and ITGA1 for EVT and CGA and CGB for STB (Supplementary Fig. S1B, C).
First, we investigated the expression levels of SARS-CoV-2 entry proteins ACE2 and TMPRSS2 in both undifferentiated TSCs and TSC-derived STB and EVT. We observed significantly higher mRNA expression levels of ACE2 in both TSC-derived EVT and STB (1048: P < 0.001; 1049: P < 0.001) compared with undifferentiated TSCs (Fig. 1A). TMPRSS2 showed higher expression in EVT (1048: P < 0.05; 1049: P < 0.01) but not in STB (Fig. 1A) by quantitative real-time polymerase chain reaction (qPCR). Western blot showed high expression of both ACE2 and TMPRSS2 proteins in the differentiated cells and no or low expression in undifferentiated TSCs (Fig. 1B). These data correlate with expression levels of ACE2 and TMPRSS2 in first trimester placenta, where they are expressed in the mature trophoblast compartments (STB and EVT in the cell column) but not in the progenitor compartment (CTB) (Supplementary Fig. S1D) [14]. These data suggest that differentiated trophoblast, TSC-derived STB and EVT, but not progenitor CTB/undifferentiated TSCs, have the potential to support SARS-CoV-2 infection.
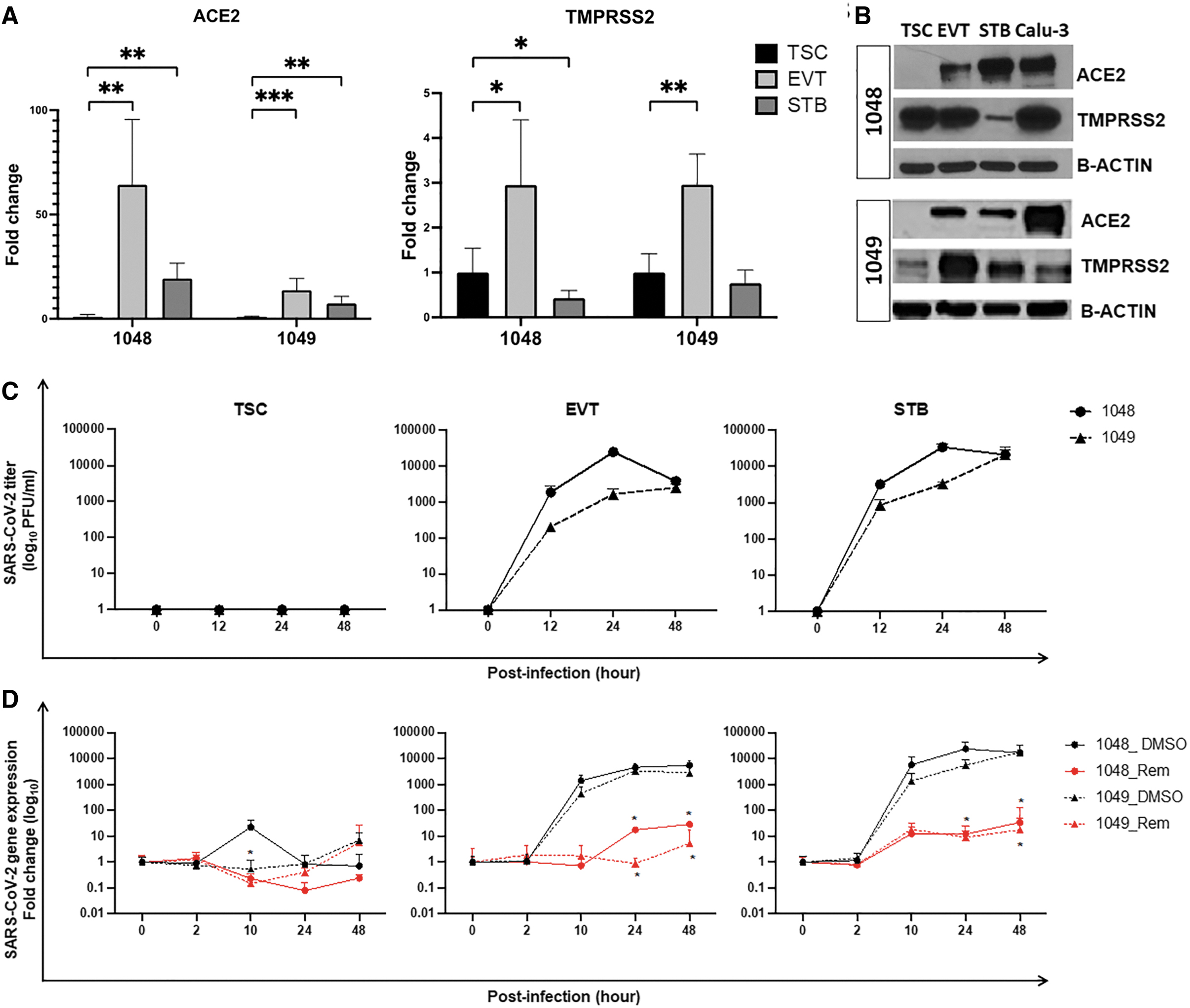
TSC-derived EVT and STB, but not undifferentiated TSCs, show active SARS-CoV-2 infection.
Differentiated EVT and STB cells support replication of SARS-CoV-2
We next sought to investigate if SARS-CoV-2 could infect and replicate within the trophoblast compartment of the developing placenta using TSC-derived cells. We exposed both undifferentiated TSCs and EVT and STB derivatives to SARS-CoV-2 (MOI = 1) for 1 h. After viral input removal (t = 0 h), we cultured the cells for additional 48 h with or without remdesivir, a nucleotide analog that inhibits active SARS-CoV-2 replication. We collected cell lysates and supernatants at the time point indicated postinfection (h p.i.) and evaluated viral growth as levels of released infectious viral particles measured by plaque assay and by qPCR for the SARS-CoV-2 nucleoprotein.
While increasing levels of infectious viruses were measured in EVT and STB cells, peaking at 24 h p.i. for both 1048 and 1049 cell lines, no plaques resulted from undifferentiated TSC supernatants (Fig. 1C). Consistent with the plaque assay data, no expression of viral mRNA was detected in undifferentiated TSCs, with or without remdesivir (Fig. 1D). In contrast, increasing levels of viral mRNA were detected in TSC-derived EVT and STB cells over time (Fig. 1D) and were significantly reduced in remdesivir-treated cells. Therefore, SARS-CoV-2 could productively infect the differentiated trophoblast compartments but not the stem cells/progenitors of early placenta. Collectively, these data suggest that SARS-CoV-2 can infect the trophoblast barrier of intact first trimester placenta.
Transcriptomic profiling of TSC-derived EVT after SARS-CoV-2 infection revealed activation of the IFN signaling pathway
To determine the overall response driven by the SARS-CoV-2 infection in trophoblast cells, we performed bulk RNA sequencing in noninfected (NI) cells and infected cells at 10 and 24 h p.i. (each group n = 6, three per cell line). PCA showed that each trophoblast cell type maintained different transcriptomic profiles (Supplementary Fig. S2A). As undifferentiated TSCs were not infected by SARS-CoV-2, we did not further evaluate these cells. PCA of the TSC-derived EVT samples showed that principal component (PC) 1 accounted for transcriptomic changes across the infection time points, whereas changes along PC2 were driven by line-to-line differences (Fig. 2A).
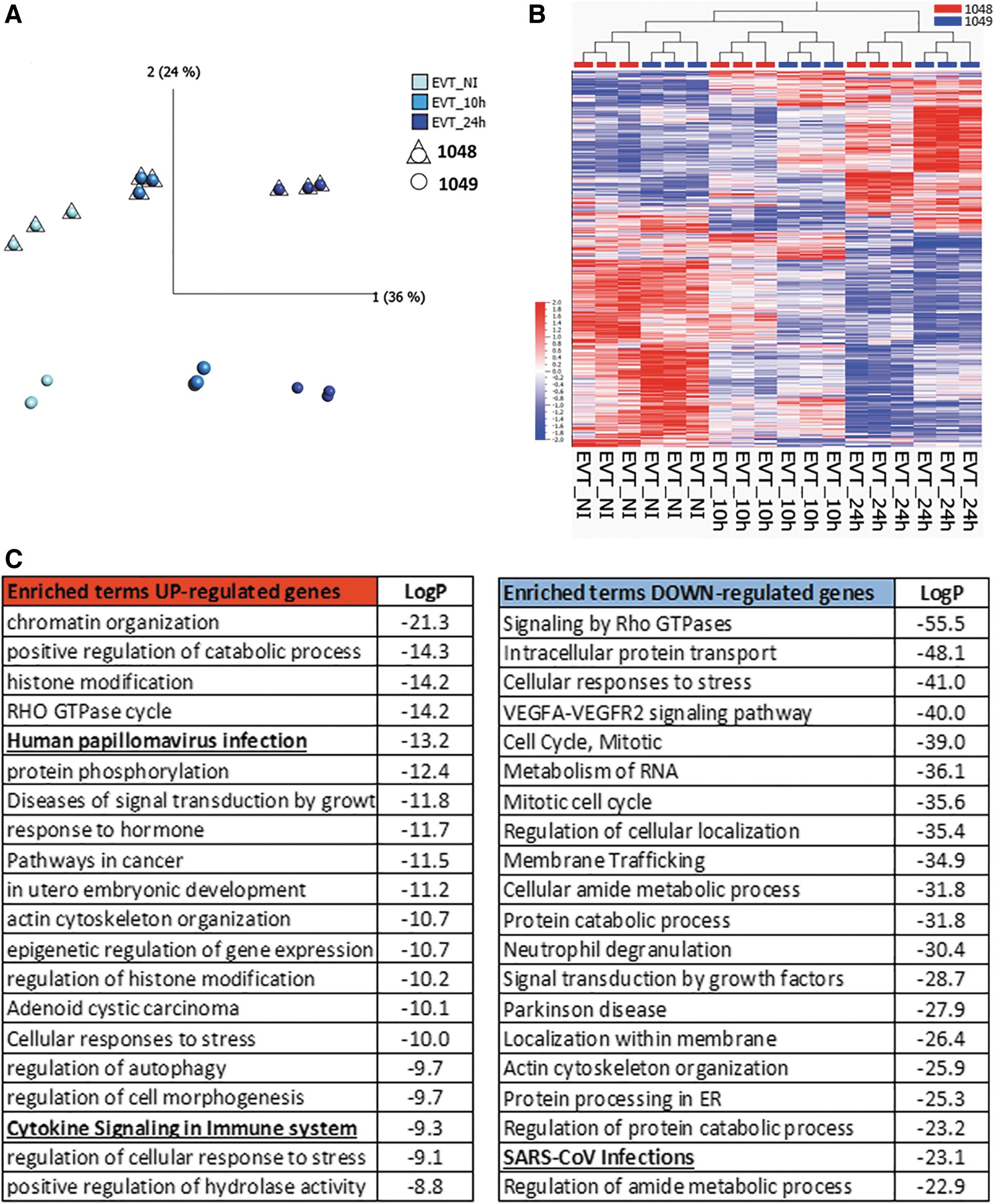
TSC-derived EVT showed gene expression changes in response to SARS-CoV-2 infection.
We found 2,425 differentially expressed genes (DEGs) between EVT NI and 24 h p.i. (q value <0.05) (Fig. 2B and Supplementary Data), of which 899 genes were upregulated and 1,526 were downregulated in EVT 24 h p.i. Gene Ontology and pathway analysis were performed for both set of genes using Metascape [34]. At 24 h p.i., the top 10 terms enriched in the upregulated gene list included chromatin remodeling, viral infections, cytokine signaling, and in utero embryonic development (Fig. 2C). Downregulated genes showed enrichment for cell cycle, membrane trafficking, and SARS-CoV-2 infection regulation (Fig. 2C).
Transcriptomic profiling of TSC-derived STB showed strong viral response to SARS-CoV-2 infection at 10 h p.i., but not at 24 h p.i.
TSC-derived STB showed a time-sensitive response to SARS-CoV-2 infection: at 10 h p.i., both cell lines showed a similar response, whereas their response at 24 h p.i. was dissimilar, with 1048 returning to a state more similar to NI and 1049 showing a unique signature (Fig. 3A). We found 682 differentially expressed genes between STB 10 h p.i. and NI, of which 118 were upregulated and 564 were downregulated (Supplementary Fig. S2B and Supplementary Data). Gene Ontology and pathway enrichment analysis for the upregulated genes showed enrichment in terms associated with IFN-stimulated genes (ISGs), cytokine signaling, and SARS-CoV-2-activated innate and adaptive immune response (Fig. 2B). Downregulated genes were enriched in terms for protein processing, vesicle-mediated transport, response to hormones, and cell signaling modulation (Supplementary Fig. S2C).

TSC-derived STB showed earlier peak of gene expression response to SARS-CoV-2 infection.
For the 24 h p.i. time point, we only evaluated changes in 1049 TSC-derived STB. We found 3,251 differentially expressed genes between 1049 STB 24 h p.i. compared with NI, of which 2,090 were upregulated and 1,161 were downregulated (Supplementary Fig. S2D). Gene Ontology and pathway enrichment analysis for both set of genes suggest maintenance of the STB status with arrest of cell cycle among the downregulated genes, and membrane trafficking and protein transport, as well as stress response, which could potentially be due to prolonged culture or in response to viral infection, enriched in upregulated genes (Supplementary Fig. S2E). Only 54 genes were shared between 1049 STB at 10 and 24 h p.i., presenting 46% of the upregulated genes at 10 h p.i. and 3% of the upregulated genes at 24 h p.i. (Supplementary Fig. S2F).
Interestingly, comparing the DEGs for EVT and STB at the time points with the strongest transcriptional changes following viral infection (EVT at 24 h p.i. and STB at 10 h p.i., as determined by Gene Ontology and pathway enrichment analysis), only 28 genes were upregulated in both (Fig. 3C and Supplementary Data). These 28 genes were involved in regulation of the NF-kappaB signaling and cellular response to stress. As excessive inflammation in the sites of infection has been associated with poorer outcomes with SARS-CoV-2, we decided to focus the next set of analyses on the Gene Ontology and pathway terms associated with inflammatory response at these trophoblast lineage-specific peak viral response times.
Inflammatory response of trophoblast cells to SARS-CoV-2 infection
We decided to focus on the inflammatory response elicited by SARS-CoV-2 infection in TSC-derived trophoblast. For EVT, we focused on the enriched term “Human papillomavirus infection” and “Cytokine Signaling in Immune system” in the upregulated terms (Fig. 4A, UP), and “SARS-CoV-2 infections” in the downregulated terms (Fig. 4A, DOWN). Interestingly, under the enrichment analysis for “Human Papillomavirus infection,” we found terms associated with different viruses including SARS-CoV-2, Ebola virus, Epstein–Barr virus, human CMV, and Herpes simplex virus, suggesting that the response elicited by SARS-CoV-2 in TSC-derived EVT is part of a common antiviral response (Supplementary Fig. S3).

hTSC-derived EVT and STB showed IFN-associated response to SARS-CoV-2 infection.
The DEGs associated with this response belonged mainly to the IFN response (Fig. 4A), including eight ISGs: interferon alpha-inducible protein 6 (IFI6), interferon-stimulated gene 15 (ISG15), EIF2AK2, IFI27, interferon regulatory factors 7 (IRF7), TNF receptor-associated factor 3 (TRAF3), interferon-induced transmembrane protein 1 (IFITM1), and IRF3 (Fig. 4B). Conversely, genes associated with the immune response that were downregulated in EVT 24 h p.i. included CAVX, IFNGR1, KPNA2, and NUP155, suggesting a compensatory or modulatory cellular effort on the IFN pathway (Fig. 4C).
In TSC-derived STB, we focused on “SARS-CoV-2 activates/modulates innate and adaptive immune responses,” “Antiviral mechanism by IFN-stimulated genes,” and “Regulation of I-kappaB kinase/NF-kappaB signaling” (Fig. 4D). These included MX1, ABCE1, TRIM25, and KPNA4 genes, which have been previously identified as regulators or effectors of the cellular antiviral response (Fig. 4E) [36,37]. The 28 genes upregulated in both EVT 24 h p.i. and STB 10 h p.i. included TRIM25 and TRAF4, involved in antiviral innate immunity [38,39]. Collectively, our data on TSC-derived trophoblast are consistent with the data previously reported on placental SARS-CoV-2 infection and its effect on IFN signaling as main inflammatory response pathway [40].
Inflammatory response in placentas at term from pregnancy infected with SARS-CoV-2
We then decided to investigate whether these inflammatory pathway-associated genes were differentially misregulated in placentas of patients with known SARS-CoV-2 infection in the first trimester of pregnancy and delivering at term with chronic inflammatory patterns of placental injury. Specifically, we picked placentas with severe chronic villitis (n = 6, Supplementary Fig. S4A), characterized by maternal cell infiltration in the placental villi (V) with associated perivillous fibrin deposition and avascular villi. We identified a full-thickness section from each that showed the histological lesion, isolated RNA from them, and compared them to placentae from pregnancies without SARS-CoV-2 infection or villitis, delivered at term.
To probe the presence of an inflammatory response in these samples, we evaluated the expression of a panel of 28 cytokines using a commercially available kit. Six proinflammatory (IL1B, IL6, IL8, IL16, IL1A, and TNF-α) and two anti-inflammatory cytokines (IL5 and IL10) were within the range of detection (Supplementary Fig. S4B). Most of the detectable cytokines were significantly upregulated in SARS-CoV-2-infected placentae with villitis, compared with noninfected placentae. Interestingly, we did not detect expression of any of the eight ISGs included in the panel (data not shown), and TNF was not upregulated compared with control samples (Supplementary Fig. S4B).
Of the inflammation-associated genes upregulated secondary to SARS-CoV-2 infection in TSC-derived EVT or STB, we tested for the presence of seven (EVT) or four (STB) such genes, which are mostly involved in the type I IFN downstream signaling. Interestingly, all of them were either downregulated or unchanged compared with controls (Supplementary Fig. S4C, D), suggesting that, while the inflammatory cells potentially recruited by the initial infection persist in the placenta long after resolution of the infection, the early regulation of ISGs during the initial infection might be difficult to capture, particularly in a setting where the effects of maternal infection in early pregnancy can only be evaluated months later upon placenta examination after term delivery.
Discussion
Placental function is crucial for both maternal and fetal health during pregnancy. Among other functions, the placenta serves as both a physical and an immunological barrier to pathogen invasion. Nevertheless, an uncontrolled and sustained inflammatory response within the placenta can contribute to injury of this important interface, which, in turn, can lead to both adverse pregnancy outcomes and abnormalities in fetal growth and development [41 –45]. SARS-CoV-2 infections during gestation have been associated with multiple patterns of placental injury, including chronic inflammation (villitis/intervillositis), perivillous fibrin deposition, fetal and maternal vascular malperfusion, and trophoblast necrosis, with the most consistent pattern being chronic inflammation in combination with perivillous fibrin and trophoblast necrosis, which has been deemed diagnostic of “COVID-19 placentitis” [4]. Currently, little is known about the mechanisms by which maternal SARS-CoV-2 infection early in gestation might affect placental function and pregnancy outcomes.
In agreement with published studies [14,29], we observed ACE2 and TMPRSS2 expression in the STB compartment of first trimester placentae as well as in the developing EVT in the cell column. We also noted that these entry proteins were lacking in CTB, the compartment containing trophoblast progenitors. This is in accordance with studies showing viral particles in the STB compartment with an associated inflammatory response and trophoblast necrosis [12,46], although in situ infection of EVT has yet to be reported.
It is currently unknown if SARS-CoV-2 can indeed infect and replicate within the trophoblast compartment of first trimester placentae. Therefore, we assessed if a recently developed in vitro model of first trimester trophoblast, namely human TSC, could be used to study SARS-CoV-2 infection within this compartment. Our TSC lines were derived from first trimester placental tissues, resemble cells similarly derived by Okae et al. based on RNA profiling [30], and can be differentiated into both functional STB and EVT to recapitulate the trophoblast compartment in the early stages of placental development. Similar to the data on first trimester placentae, undifferentiated TSCs showed low or no expression of ACE2 and TMPRSS2, whereas these proteins were detected in TSC-derived STB and EVT, confirming their suitability as a model to study SARS-CoV-2 infection in the trophoblast compartment of the developing placenta.
Consistent with the ACE2 and TMPRSS2 expression data, we found that SARS-CoV-2 was able to infect TSC-derived STB and EVT, but not the undifferentiated cells. Moreover, we observed viral replication and growth within the mature trophoblast cells, which was decreased by treatment with the viral replication inhibitor, remdesivir. To the best of our knowledge, this is the first study showing productive viral replication in trophoblast cells representative of the early gestation placenta. Previous studies have shown potential viral replication using a SARS-CoV-2 virus reporter in term CTB but not STB [14]. Moreover, Karvas et al. used three-dimensional (3D) organoids generated using naive pluripotent stem cell-derived and placenta-derived TSCs and showed limited infection by a SARS-CoV-2 pseudovirus as well as the intact virus [47].
However, as the authors admitted, organoids might not represent an ideal model to study infection in the STB compartment, due to their inside out structure, which shields the STB compartment from the viral particles. Similar to our data, their experiment with two-dimensional (2D) TSCs showed no infection in undifferentiated cells. However, they also noted a low infection rate with TSC-derived STB in 2D, although this was performed only with the SARS-CoV-2 pseudovirus using staining of the Spike protein as readout. Our study used the actively replicating virus in both TSC-derived STB and EVT and showed not just expression of the SARS-CoV-2 nucleoprotein in TSC-derived trophoblast but dampened expression following remdesivir treatment. Moreover, we went a step further, showing release of newly formed infectious SARS-CoV-2 viruses into the supernatant over time by conducting plaque assay evaluation at 0, 2, 10, 24, and 48 h p.i. Taken together, our data strongly suggest that SARS-CoV-2 can infect and replicate productively in the differentiated trophoblast compartment within first trimester placentas.
RNA-seq analysis of infected TSC-derived STB and EVT showed a cellular response to the SARS-CoV-2 infection, but with different peak times and mechanisms between the two cell types. TSC-derived EVTs response was greatest at 24 h p.i., although some response was noted already at 10 h p.i. Gene Ontology analysis showed various cellular responses to stress, including response to different viral infections and cytokine immune response. Genes associated with viral infections and upregulated in EVT included ISG15, IFI6 and IFI27, IFITM1, as well as IRF3 and IRF7, and TRAF3. These genes are part of the innate immune system, a primary host defense strategy to directly suppress viral infections and stimulate the host inflammatory response, including the secretion of proinflammatory cytokines/chemokines and IFNs [48]. Most of these genes have been observed in the response of other cell types to SARS-CoV-2 infection, including alveolar cells [49], as well as cellular responses to other viral infections, including influenza A virus.
However, while most of these genes have an antiviral effect, some are known to promote infection and inflammation in the case of SARS-Cov-2. For example, IFITM1 has been associated with more efficient SARS-CoV-2 infection rather than blockage [50]. Moreover, ISG15 upregulation in SARS-CoV-2 infection might be associated with an enhanced inflammatory response, potentially increasing the infiltration of immune cells at the maternal–fetal interface and causing inflammatory patterns of placental injury observed in the long term [51]. Interestingly, we did not observe upregulation of IFN genes by RNA-seq. However, evidence suggests that some viruses, such as CMV, can regulate ISGs via IRF3 signaling in an IFN/JAK/STAT-independent manner [52].
TSC-derived STBs response to viral infection showed a more rapid onset, observed to peak at 10 h p.i., compared with that of EVT. Gene Ontology analysis showed enrichment in terms associated with IFN, cytokine, and NF-kappaB signaling and overall regulation of viral life cycle. Genes upregulated at 10 h p.i. in STB included MX1, TRIM25, ABCE1, and KPNA4. These genes are involved in the modulation of the innate immune response by IFN proteins [48].
The IFN-mediated response is usually observed at the onset of the local immune response upon viral infection and triggers a cascade of events resulting in the activation of antiviral ISGs and the upregulation of chemokines and cytokines, which have both local and systemic effects [53]. While the latter have been observed in SARS-CoV-2 patients, the IFN-mediated response is much more difficult to detect in a clinical setting where the temporal separation between initial viral infection and tissue examination is significantly long. Therefore, the IFN-mediated response appeared to be dampened/absent in placentas from SARS-CoV-2-affected pregnancies, delivering at term, despite histological evidence of chronic villitis.
We selected cases based on two main criteria: (1) SARS-CoV-2 infection in the first trimester, as confirmed by nasal swab, and (2) histopathological evidence of chronic villitis in such placentas, compared with those from noninfected patients, as potential sign of adverse effect from SARS-CoV-2 infection. Higher expression of both pro- and anti-inflammatory cytokines confirmed the presence of an inflammatory lesion in these samples. While we are not able to confirm that the trophoblasts were in fact affected or that inflammation in these placentae was the direct result of infection by SARS-CoV-2, the persistence of inflammatory cells at the maternal–fetal interface might suggest an earlier local induction of the innate immune response. Our in vitro data suggest a potential way in which SARS-CoV-2 infection of the trophoblast compartment could have triggered an IFN-mediated response, which resulted in maternal immune cell infiltration in the placental bed.
Unfortunately, the current TSC model does not allow maintenance of the differentiated trophoblast cell types for longer time periods in culture, and alternate trophoblast organoid models, with STB on their outer layer, will be required to test the long-term effects of the IFN-mediated response following SARS-CoV-2 infection in this compartment. Moreover, more complex placental models, including both trophoblast and nontrophoblast cells, will be required to investigate the full response of this organ to a potential local infection of the trophoblast compartment. Despite the limitations of this study, including specifically the long temporal gap between infection and placental evaluation in our in vivo (clinical) cohort, our findings provide an important first step toward understanding the mechanisms by which SARS-CoV-2 infection in early gestation could result in the increased risk of pregnancy complications, as reported by multiple groups during the pandemic [1,2,4].
Conclusions
In summary, our study of the maternal–fetal interface in the context of maternal SARS-CoV-2 infection provides evidence that first trimester differentiated trophoblast could be infected and serve as a reservoir for the replicating virus in the developing placenta. Moreover, both early EVT and STB elicit an IFN-mediated innate immune response similar to other cell types infected with SARS-CoV-2 [54,55]. While this response may be transient and/or be altered over time, it could initiate a cascade of events resulting in some of the placental lesions observed in SARS-CoV-2-affected pregnancies. Overall, we have shown that placenta-derived TSCs are a robust in vitro model to investigate the effect of this virus specifically in the trophoblast compartment of the placenta. In the future, the development of mixed culture models, including both trophoblast and nontrophoblast placental cells, and/or optimized 3D organoids could help better model long-term effects of SARS-CoV-2 and other viral infections in the developing placenta.
Footnotes
Acknowledgment
We thank Dr. Parast for the support and mentorship on this project and the valuable comments and feedback on this article.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This publication includes data generated at the University of California San Diego Institute for Genomic Medicine (IGM) Genomics Center utilizing an Illumina NovaSeq 6000 that was purchased with funding from the National Institutes of Health SIG grant (#S10 OD026929).
Supplementary Material
Supplementary Data
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Table S1
Supplementary Table S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.